Nitrogen and Phosphorus Concentration in Leaf Litter and Soil in Xishuangbanna Tropical Forests: Does Precipitation Limitation Matter?


1
CAS Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, 88 Xuefu Road, Kunming 650223, China
2
Pitchandikulam Forest Consultants, Auroville 605 101, Tamil Nadu, India
*
Author to whom correspondence should be addressed.
Forests 2019, 10(3), 242; https://doi.org/10.3390/f10030242
Submission received: 18 February 2019
/
Revised: 4 March 2019
/
Accepted: 5 March 2019
/
Published: 8 March 2019
(This article belongs to the Special Issue Impact of Global Change on Soil Carbon Storage and Biogeochemical Cycles in Tropical Forest Ecosystems)
Abstract
:Tropical forests are generally expected to be limited by the availability of nitrogen (N) and phosphorus (P), and these nutrient limitations could be increased by changes in forest biogeochemistry due to limited precipitation. This effect is presumed to be enhanced in the forests predominated by monsoon climate. The present study examined the impacts of monthly precipitation on total N and P in leaf litter and soil of Xishuangbanna tropical forests. Litterfall and top soil were sampled from each of the five 20 × 20 m plots established in the primary (PTF) and secondary tropical forests (STF), at monthly interval for one year. Soils were strongly acidic and showed significant differences between the sites. The monthly amounts of soil and leaf litter nutrients showed great variations between the PTF and STF. Leaf litter N and P were associated with precipitation in both dry and rainy seasons. Soil N and P were not significantly related to precipitation, indicating that changes in vegetation composition and litterfall production together accounted for variation in soil N and P. Our results suggest that the precipitation limitation may affect the leaf litter N and P changes, but did not support the prediction that precipitation limitation can immediately lead to effects on soil N and P in the Xishuangbanna tropical forests.
1. Introduction
In tropical forest ecosystems, nitrogen (N) and phosphorus (P) are the most important limiting nutrients for carbon sequestration [1]. Carbon (C) constitutes the basic structure of plants and accounts for ca. 50% of plant biomass, N is an essential component of enzymes, and P is an essential element of nucleic acids and membrane lipids [2]. Low P availability is usually expected [3] in tropical forest ecosystems although N limitation is also common [4]. These nutrient limitations could be exacerbated by changes in forest biogeochemistry in response to less rainfall [5]. Consequently, ecosystems with very high precipitation often have low nutrient availability because of slow decomposition and high leaching loss [6,7]. Climate and soil quality are widely recognized as major drivers of virtually all properties of ecosystems and communities. However, the effects of climate on soil properties are not widely appreciated, despite major advances in the understanding of soil formation and ecosystem dynamics [8].
C fluxes will become increasingly vulnerable during the 21st century in tropical forests due to reduced mean annual precipitation for large areas of the tropics predicted by the different Earth-system models [9,10]. Many studies indicated that C cycling is commonly limited by N and/or P from nutrient manipulation experiments in tropical forests [4,11,12,13] and such limitations will be expected to change forest biogeochemistry in response to limited precipitation [5,14]. Other climatic factors, such as temperature, appear to be a partial driver of variations in senesced-leaf N and P [15], and thermal environment can also influence geographic patterns in leaf N and P [16]. In contrast, drought severity was also a key climatic factor correlated with leaf N and P, which was more appropriate for evaluating the impact of water status than precipitation [17].
The soil chemical variations can be primarily explained by climate (e.g., leaching intensity and duration of droughts, including El Nino) and geology (geological age, silicate clays, Fe oxides, and bedrock P concentration) [18]. However, topography, soil texture, and stand characteristics contributed the most to spatial variations in soil organic C, and total N and P [19]. Spatial patterns of total P strongly resembled woody vegetation cover throughout the soil profile [20], but differed from total N predominately regulated by biotic factors (plant residues and microbial communities) [21], especially in subsurface soils. The availability of P and cations typically declined in old, highly weathered soils, as they have been leached out of the system or become bound in unavailable forms [22]. Therefore, P or relatively mobile cations may limit biological processes and regulate N cycling [23,24] because the soil N undergoes continuous transformation and loss in/from soil through the combined processes of nitrification and denitrification, leaching, and erosion [25]. The C:N ratio was usually lower in the driest forest soils affected by fire due to the higher proportion of C lost relative to N during burning [26]. Consequently, the more distinct altitudinal trend of N, particularly for the deep layer, could also be due to the effect of fire [27]. Soil disturbance that increases the calcium content of the topsoil could also reduce the soil N and P availability [28].
In China’s terrestrial ecosystems, the leaf litter N and P concentrations increase with annual precipitation and temperature [29], but soil P limits primary production [30,31,32]. On the other hand, N deposition increases soil acidity, decreases litter decomposition rates, and suppresses soil respiration, and therefore favors soil C accumulation [33,34]. Plant chemistry, soil biogeochemical processes, and vegetation composition, each of which can directly influence plant N or P in spite of temperature and precipitation gradients [15,35,36,37]. In Xishuangbanna, southwestern China, the maximum loss of all nutrients was found in stems followed by branches, roots, and leaves [38], as a result of higher litter decomposition rate [39]. Studies on the relationship between monthly precipitation and nutrient content provide insights to examine the potential effects of seasonal drought on local forests under a monsoon climate. Hence, the main objective of the present study is to quantify the relationship between soil and leaf litter N and P, and precipitation limitation and temperature in two different forest types. We hypothesized that the precipitation is primarily limiting N and P in leaf litter and soil of Xishuangbanna tropical forests.
2. Materials and Methods
2.1. Study Area
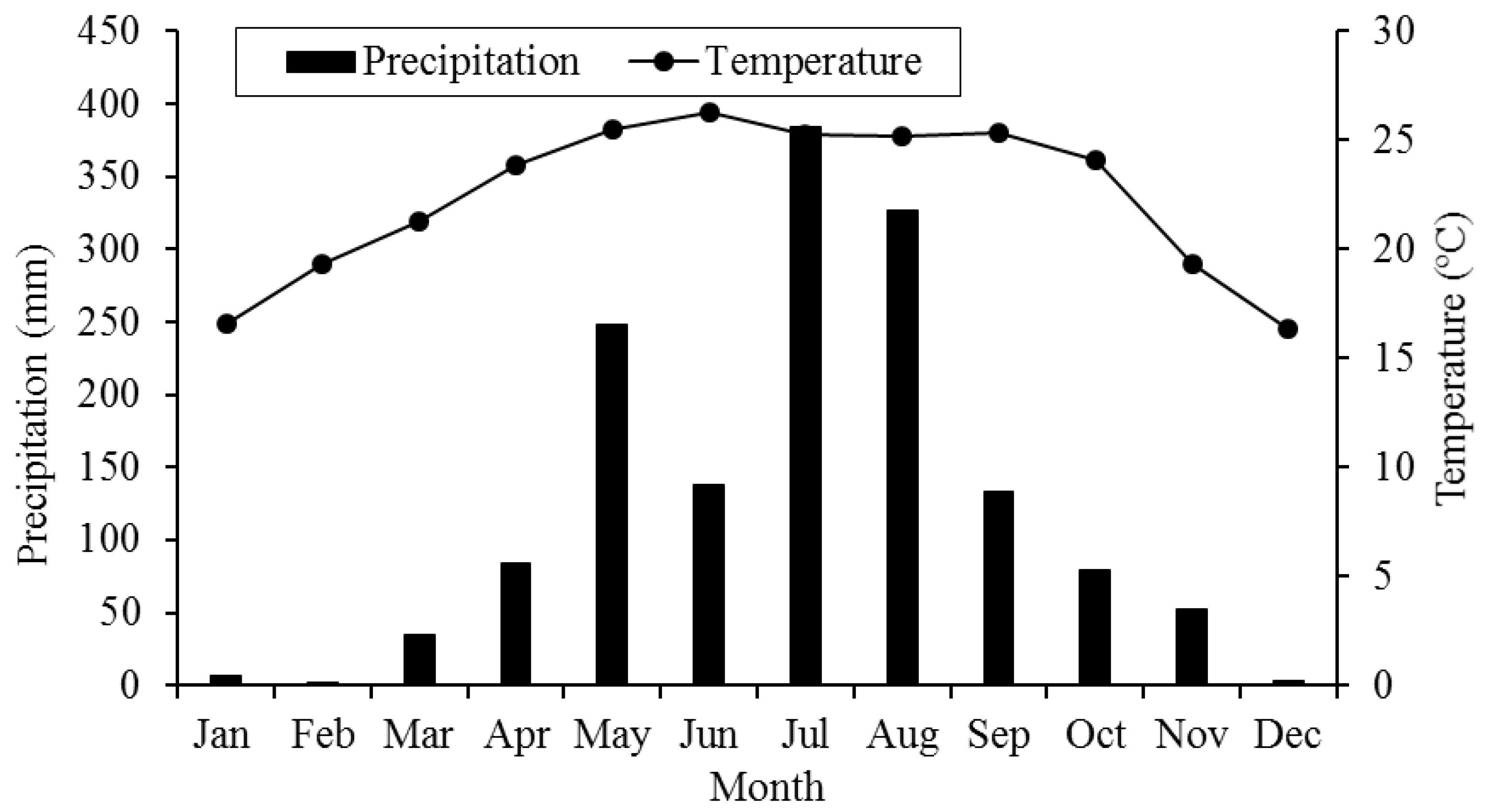
Xishuangbanna (21°08′–22°36′ N, 99°56′–101°50′ E) is located in the southwestern Yunnan Province of China. This region has alternating dry (November–April) and rainy (May–October) seasons, a typical monsoon climate. Monthly precipitation and temperature data were gathered from Xishuangbanna Station for Tropical Rainforest Ecosystem Studies (21°54′ N, 101°16′ E, 560 m.a.s.l.) near the plots selected for the study during the period 2008–2009 (Figure 1). Mean annual precipitation is about 1500 mm, and about 87% of precipitation occurs in the rainy season. Mean annual temperature is 21.7 °C, with monthly temperatures ranging from 15.6 to 25.3 °C. In addition, the absolute average amount of annual fog drip was 89.4 mm, which contributes nearly 5% of the annual rainfall, with 86% of the fog drip occurring in the dry season in the Xishuangbanna forest stand (November–April) [40]. Tree seedlings and understory species also partially rely on fog moisture for seedling growth, especially at the peak of the dry season due to strong seasonality of soil water content in this area [41].
Soils in Xishuangbanna are derived from both igneous and sedimentary rocks [42]. There are three main soil types in the Xishuangbanna region: (1) Laterite soil developed from siliceous rocks, such as granite and gneiss, which occurs below 1000 m elevation with a deep solum and thin humus horizon, (2) lateritic red soil, derived from sandstone substrates, which occurs in areas above 1000 m elevation, and (3) limestone hills which have soil derived from hard limestone substrate of Permian origin with a pH of 6.75 [43]. Soil bulk density is 1.2 g cm−3 with a pH of 5.4 [44], and potential splash erosion rate is 2552 g m–2 in the Xishuangbannna tropical rainforest [45].
The climax vegetation type in this area is tropical seasonal rainforest (primary forest) occurring in the valleys and lowlands below 1000 m elevation, and the south-subtropical evergreen broad-leaved forest, which occurs on the mountains above 1000 m elevation. This study was conducted in a tropical seasonal rainforest (primary tropical forest, hereafter PTF) and a secondary forest (STF) evolved from PTF disturbance. Details about the physical geography, vegetation characteristics and tree diversity are reported in Cao et al. [46] and Zhang and Cao [47].
2.2. Litterfall and Soil Sample Collection
A total of five 20 × 20 m plots with an interdistance of 50 m were established in each of the two sites (PTF and STF). The plots within each site were chosen on the basis of stand structure, vegetation type and disturbance. The distance between the two sites is 15 km and there was no significant difference in rainfall characteristics between the sites. Litterfall was collected from five traps measuring 1 × 1 m (2-mm mesh) in each study plot. All traps were permanently fixed at a 1-m height above the ground, and the accumulated litter in the traps was collected at monthly intervals (once in the middle of every month) for the entire year. Damaged traps were replaced immediately. Collected litterfall was brought to the laboratory and separated into (i) recently fallen litter, (ii) leaf fragments including twigs ≤1 cm diameter, and (iii) miscellaneous pieces, including flowers and fruits. All samples were weighed after drying at 80 °C for 24 h. Only recently fallen litter layer samples were taken for chemical analysis because the traps were emptied monthly to prevent significant decomposition and leaching loss of litter nutrients. Samples were pooled and were analyzed plot wise. Five soil samples of 10 × 10 × 10 cm (top soil) were taken randomly in each plot and pooled for monthly nutrient analysis over a period of 1 year. All soil samples were air dried for 24 h, and roots and stones were removed by hand. Litterfall data was adopted and discussed from Mani and Cao [48] to find relationships with precipitation, leaf litter and soil N, and leaf litter and soil P across forest types.
2.3. Laboratory Analysis
Soil samples were sieved (<2 mm) and subsamples were taken for chemical analysis. Soil pH and moisture were measured monthly using pH and moisture probe meters, respectively. Total nitrogen (N), phosphorus (P), potassium (K) and calcium (Ca) were analyzed from the collected subsamples. Total N values in litter and soil were determined using a C–N Analyzer (Vario MAX CN, Hanau, Germany). Total P, K, and Ca were determined by Inductively Coupled Plasma Atomic-emission Spectrometer (ICP-AES; Thermo Jarrell Ash Corporation, Waltham, MA, USA).
2.4. Data Analysis
An independent t-test was used to test the difference between sites. The coefficient of variation (CV, %) was calculated to find the relative variability between the sites. A Kolmogorov–Smirnov (K-S) non-parametric test was performed to observe normal distribution, and log10 transformation was explored to improve the normality of litterfall, pH, total N and P data before multiple linear regression analysis. Bivariate relationships between climate (precipitation and temperature) and total N and P pools were examined with Pearson’s correlation. All statistical analyses were performed using STATISTICA 10 (Stat Soft. Inc., Tulsa, OK, USA).
3. Results
3.1. Variation in Soil pH, Litterfall, Leaf Litter and Soil N and P
Soils are strongly acidic and showed significant differences between the sites (t = 2.83, p < 0.01, n = 12). The degrees of variation (CV, %) for total litterfall, leaf litter N and P were higher in PTF than STF, and those for soil N and P were higher in STF than PTF. In addition, leaf litter P (t = 2.83, p < 0.01, n = 12), soil N (t = −7.09, p < 0.0001, n = 12) and soil P (t = −12.77, p < 0.0001, n = 12) in STF were found significantly higher than PTF (Table 1).
3.2. Relationship between Main Climatic Factors and Leaf Litter and Soil Nutrients
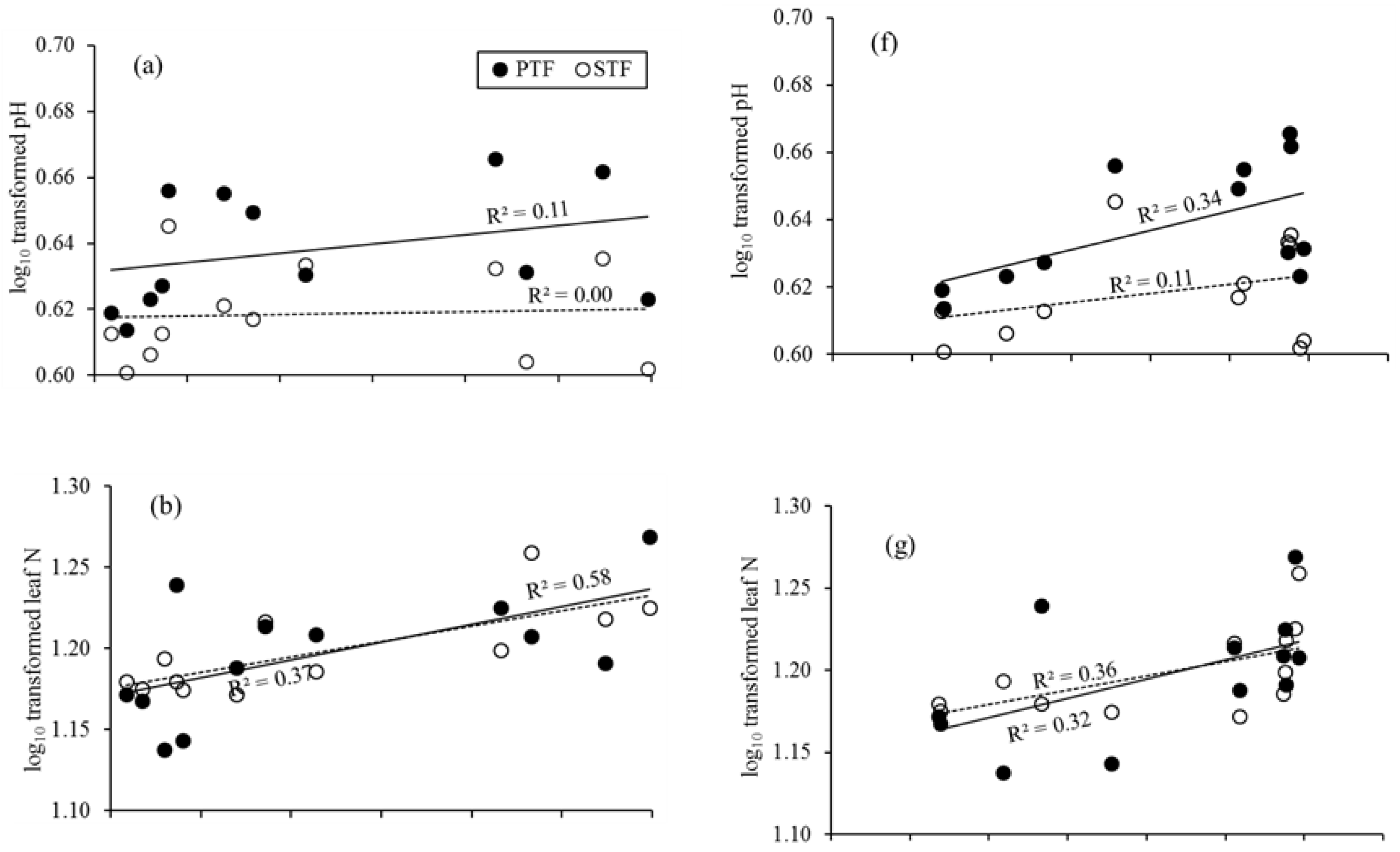
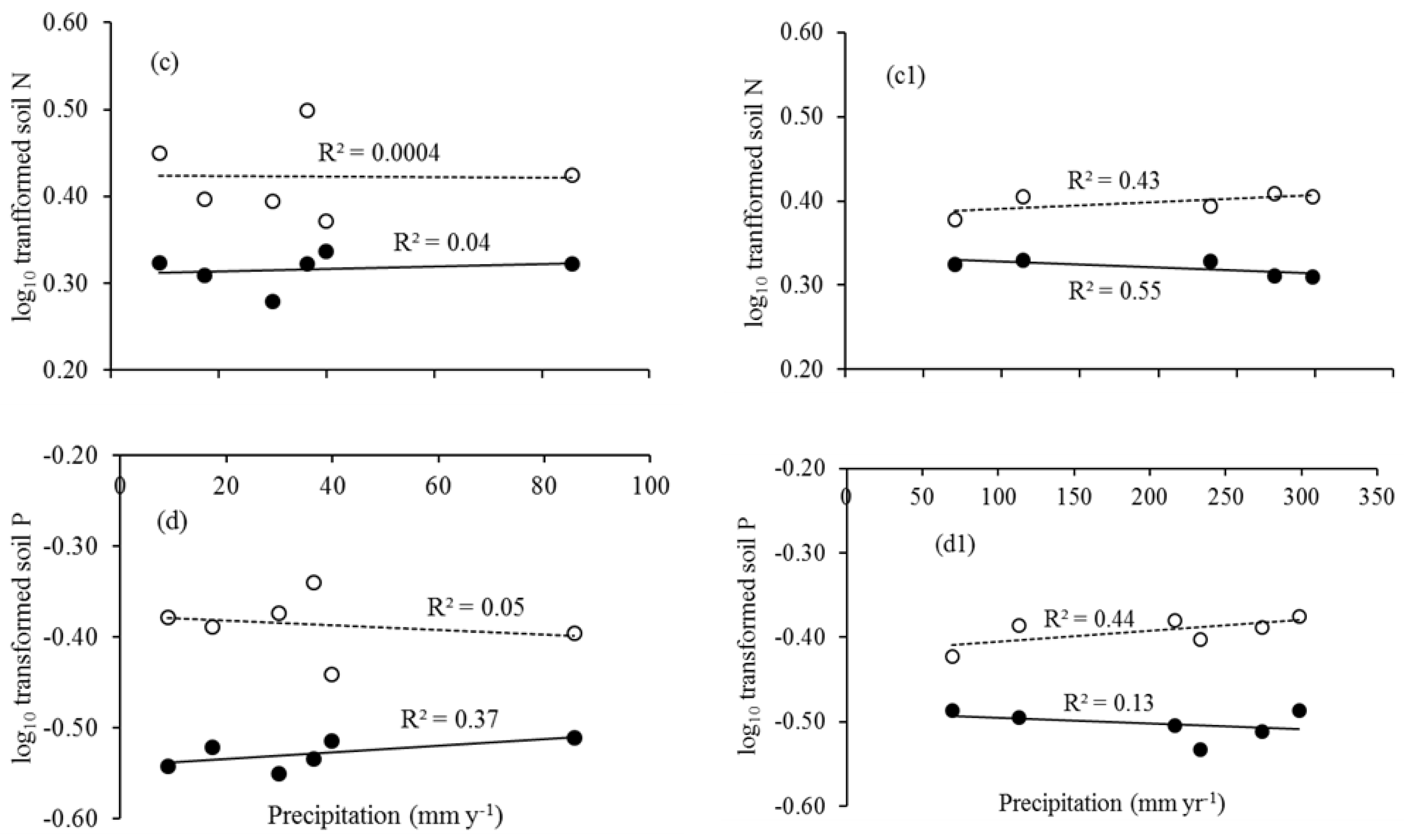
Total litterfall was not positively associated with precipitation (Figure 2). Leaf litter N and P were positively correlated with precipitation (Table 2; Figure 3). Leaf litter N in the dry season (November to April), and leaf litter P in both the dry and rainy seasons (May to October) were found to be positively correlated with precipitation in the STF. Leaf litter P had a significant negative relationship with precipitation in the dry season in the PTF (Figure 4).
4. Discussion
4.1. Litterfall Production and Climatic Variables
Total litterfall was not related to precipitation. Mani and Cao [48] reported that the total litterfall production increased in the latter part of the dry season (March–April) and there was a second minor peak at the end of the rainy season (September–October). Litterfall production increased in the dry season may be related to precipitation seasonality and landscape-level vegetation variation. Low water availability in the dry season and ambient temperatures initiate the heavy litterfall during this period [49,50,51,52]. Litterfall production may vary with local topography due to the effect of drought periods. In particular, the forests on well drained soils experience a more acute water shortage during dry months [53,54,55]. The higher litterfall production in STF mainly resulted from twigs and reproductive parts due to greater variations in canopy architecture and tree species. The dead branches usually remain on trees for a long period in the tropical forests due to its complex structure and canopy architecture, which leads to higher variations in twig litterfall when they occasionally fall on the forest floor [56,57,58]. The variety of mature trees in STF may regularly produce a high quantity of flowers, fruits, and seeds in a masting year, resulting in higher variation in the reproductive parts which were observed from a decade study in Xishuangbanna [56]. However, species composition is the most important factor influencing litter production within a climate zone [54,59]. Furthermore, the litter quality had an important influence on the rate of litter decomposition, but the decomposition environment could override differences in litter quality [60].
In the PTF site, litterfall production did not correlate with climatic variables (precipitation and temperature), indicating that the precipitation was not a limiting factor observed by both short-term [48] and long-term study [56] in the Xishuangbanna tropical seasonal rainforests. This is consistent with the result of litterfall production along successional and altitudinal gradients of subtropical forests in China, suggesting that precipitation only controls the vegetation growth and thus the litterfall production through increasing soil water content and atmospheric humidity [61]. Chave et al. [60] found that annual rainfall did not explain any variation in annual litterfall from a meta-analysis of 81 South American tropical forest sites. Therefore, there were no precipitation or lagged precipitation effects on litterfall production in the stands without a seasonal shortage of water supply [17,61].
4.2. Leaf Litter N and P and Precipitation Limitation
There were relatively higher variances (CV, %) in leaf litter N and P in the PTF, indicating the presence of monthly irregular fluctuations. In contrast, low variance in the STF indicates that the monthly variation was little (Table 1). The higher variances in the PTF was likely associated with precipitation regime (Figure 3) and favor faster litter decomposition. Leaf litter N was significantly related to precipitation in both the dry and rainy season in the STF (Figure 4). Senesced-leaf litter N and P were generally significantly related to mean annual temperature and mean annual precipitation, but temperature appears to be a partial driver of variations in senesced-leaf litter N and P assessed from a global large dataset [15].
There were significant relationships between leaf litter N, P and precipitation (Figure 3). As long as plant nutrients are usually water soluble and large amounts lost by leaching at high mean annual precipitation in tropical forest sites [15]. Yet, there was a positive relationship between leaf litter N and dry season precipitation, and leaf litter P and dry season precipitation in the secondary forest (Figure 4). This is consistent with our early findings from the same forest types that the leaf N/P was altered by seasonal control followed by floristic variation and organic matter turnover rather than precipitation [50]. However, evidence of seasonal controls found that foliar N:P values differed by 25% between wet and dry seasons in Costa Rica [61]. In contrast, results differed from a trend of declining foliar N:P values with increasing rainfall in Hawaiian montane forest [62] and Panama [63], suggesting that total rainfall may have an influence on nutrient supply. This link to canopy chemistry’s influence is minor compared to those caused by species differences and soil type [61].
4.3. Soil pH, N, P, and Precipitation Limitation
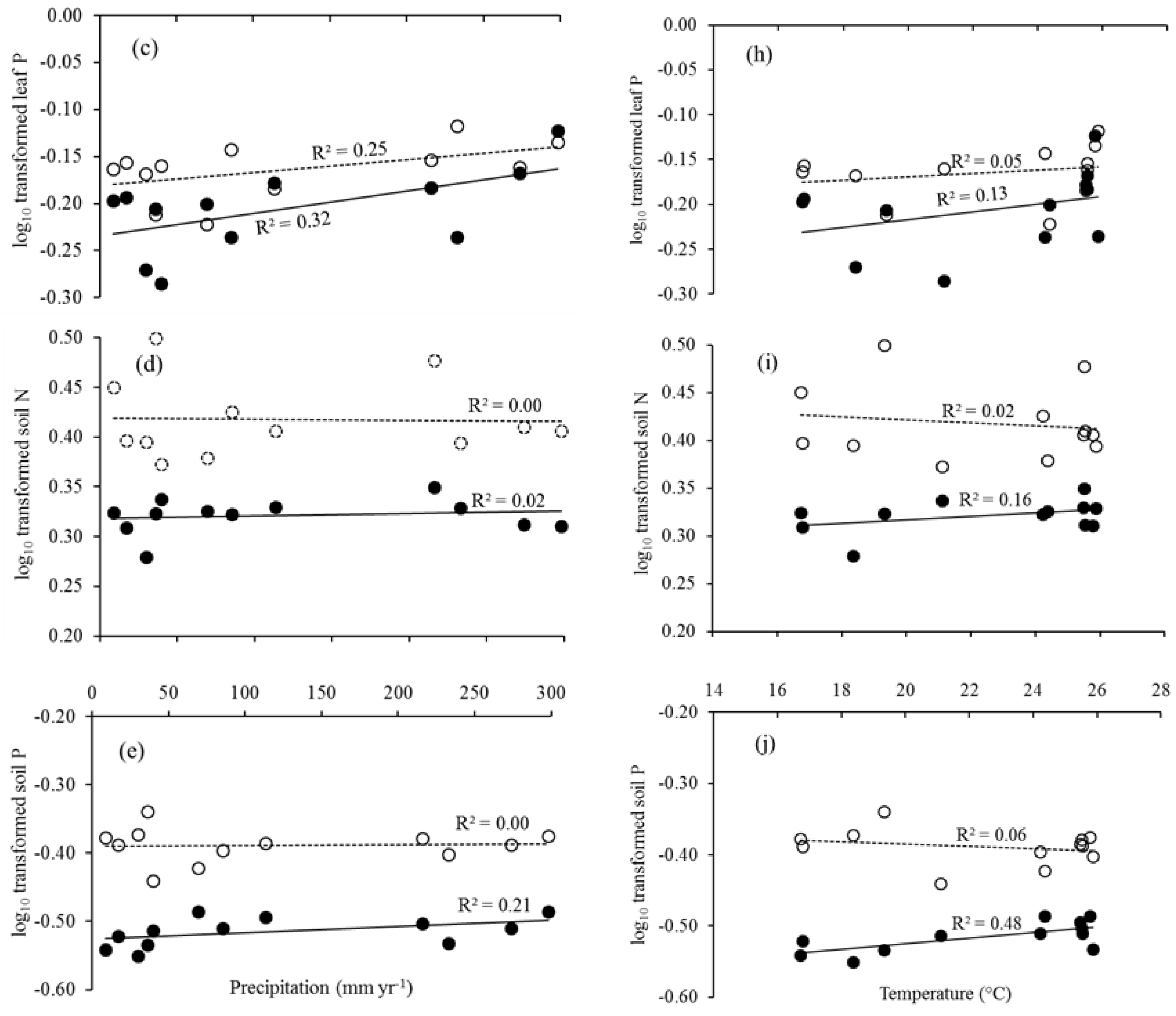
Soil pH was extremely acidic ranging from 4.2 to 4.4 and not related to precipitation (Table 1; Figure 3). Previous study reported that phosphate declines when soil pH falls below 5.5 [64]. So, more acidic soils with less total P in our study sites are consistent with the result of heavily weathered and nutrient-poor soils in the Central Amazon [65]. There was no positive correlation between pH and soil P, and this contrasted with a previous study that found positive relationships among total P, soil pH, and tree diversity in another PTF in Xishuangbanna [66]. Xue et al. [67] found that P was the most limiting nutrient and that the acidic soils further limit P availability.
Our data showed that there were no relationships between soil N and precipitation, as well as soil P and precipitation (Figure 3). Soil properties like pH, clay, and sand were also more important in explaining variation in total N and P than fine root density in the subsurface soils [20]. Soil parent materials could strongly modulate the N and P limitations [68,69] as well as soil nutrient availability under local and global scales [70]. The relationships between leaf litter N and P, and soil N and P were not detected in the study sites. This could be due to the changes in vegetation composition and precipitation limitation that together explained the variation in leaf litter nutrients at the local scale. However, in tropical ecosystems with low availability of N (especially in sandy soils) that are normally depleted in 15N, this pattern could be associated with the presence of N-fixing species [71].
5. Conclusions
Leaf litter N and P were associated with precipitation in both dry and rainy seasons, suggesting that precipitation limitation (moisture availability) is an important control for litter decomposition during the dry season, particularly in the STF. The decrease in precipitation would have profound consequences on the N and P cycles through changes in leaf litterfall nutrients, which could have long-term negative effects on net primary production and future carbon storage in these forest types. Eventually, variations in leaf N and P, and soil N and P were associated with litterfall production and floristic composition even at local scale.
Author Contributions
S.M. and M.C. conceived and designed the work. S.M. conducted the field, laboratory, and data analysis. S.M. and M.C. wrote the manuscript.
Funding
This research was funded by National Key Basic Research Program of China (grant number—2014CB954100) and the Applied Fundamental Research Foundation of Yunnan Province (grant number—2014GA003).
Acknowledgments
We extend our thanks to D. Chen, X. Deng and Y. Fu for their help. We thank Vijayalaxmi Kinhal and anonymous reviewers for helpful comments on earlier drafts of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Agren, G.I.; Wetterstedt, J.A.M.; Billberger, M.F.K. Nutrient limitation on terrestrial plant growth–Modeling the interaction between nitrogen and phosphorus. New Phytol. 2012, 194, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Bejarano-Castillo, M.; Campo, J.; Roa-Fuentes, L.L. Effects of increased nitrogen availability on C and N cycles in tropical forests: A meta-analysis. PLoS ONE 2015, 10, e0144253. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, W.H.; Dietze, M.C.; Jackson, R.B.; Phillips, R.P.; Rhoades, C.C.; Rustad, L.E.; Vose, J.M. Forest biogeochemistry in response to drought. Glob. Chang. Biol. 2016, 22, 2318–2328. [Google Scholar] [CrossRef] [PubMed]
- Zak, D.R.; Grigal, D.F. Nitrogen mineralization, nitrification and denitrification in upland and wetland ecosystems. Oecologia 1991, 88, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Turner, D.R.; Kitayama, K. Foliar nutrients during long-term soil development in Hawaiian montane rain forest. Ecology 1995, 76, 712–720. [Google Scholar] [CrossRef]
- Huston, M.A. Precipitation, soils, NPP, and biodiversity: Resurrection of Albrecht’s curve. Ecol. Monogr. 2012, 82, 277–296. [Google Scholar] [CrossRef]
- Neelin, J.D.; Münnich, M.; Su, H.; Meyerson, J.E.; Holloway, C.E. Tropical drying trends in global warming models and observations. Proc. Natl. Acad. Sci. USA 2006, 103, 6110–6115. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; 1535p. [Google Scholar]
- Elser, J.J.; Bracken, M.E.S.; Gruner, D.S.; Harpole, W.S.; Hillebrand, N.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary production in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.S.; Becklund, K.K.; Gei, M.G.; Iyengar, S.B.; Meyer, R.; O’Connell, C.S.; Schlling, E.M.; Smith, C.M.; Waring, B.G.; Werden, L.K. Nutrient addition effects on tropical dry forests: A mini-review from microbial to ecosystem scales. Front. Earth Sci. 2015, 3, 34. [Google Scholar] [CrossRef]
- Campo, J. Shift from ecosystem P to N limitation at precipitation gradient in tropical dry forests at Yucatan, Mexico. Environ. Res. Lett. 2016, 11, 095006. [Google Scholar] [CrossRef]
- Yuan, Z.; Chen, H.Y. Global trends in senesced-leaf nitrogen and phosphorus. Glob. Ecol. Biogeogr. 2009, 18, 532–542. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef]
- Zhao, W.; Reich, P.B.; Yu, Q.; Zhao, N.; Yin, C.; Zhao, C.; Li, D.; Hu, J.; Li, T.; Yin, H.; et al. Shrub type dominates the vertical distribution of leaf C:N:P stoichiometry across an extensive altitudinal gradient. Biogeosciences 2018, 15, 2033–2053. [Google Scholar] [CrossRef]
- Fujii, K.; Shibata, M.; Kitajima, K.; Ichie, T.; Kitayama, K.; Turner, B.L. Plant–soil interactions maintain biodiversity and functions of tropical forest ecosystems. Ecol. Res. 2018, 33, 149–160. [Google Scholar] [CrossRef]
- Jiang, F.; Wu, X.; Xiang, W.; Fang, X.; Zeng, Y.; Ouyang, S.; Lei, P.; Deng, X.; Peng, C. Spatial variations in soil organic carbon, nitrogen and phosphorus concentrations related to stand characteristics in subtropical areas. Plant Soil 2017, 413, 289–301. [Google Scholar] [CrossRef]
- Zhou, Y.; Boutton, T.W.; Wu, X.B. Soil phosphorus does not keep pace with soil carbon and nitrogen accumulation following woody encroachment. Glob. Chang. Biol. 2018, 24, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Sanford, R.L., Jr. Nutrient cycling in moist tropical forest. Ann. Rev. Ecol. Syst. 1986, 17, 137–167. [Google Scholar] [CrossRef]
- Jackson, R.B.; Manwaring, J.H.; Caldwell, M.M. Rapid physiological adjustment of roots to localized soil enrichment. Nature 1990, 344, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, O.A.; Derry, L.A.; Vitousek, P.M.; Huebert, B.J.; Hedin, L.O. Changing sources of nutrients during four million years of ecosystem development. Nature 1999, 397, 491–497. [Google Scholar] [CrossRef]
- Berhe, A.A.; Torn, M.S. Erosional redistribution of topsoil controls soil nitrogen dynamics. Biogeochemistry 2017, 132, 37–54. [Google Scholar] [CrossRef]
- Vega, J.A.; Fonturbel, T.; Merino, A.; Fernandez, C.; Ferreiro, A.; Jimenez, E. Testing the ability of visual indicators of soil burn severity to reflect changes in soil chemical and microbial properties in pine forests and shrubland. Plant Soil 2013, 369, 73–91. [Google Scholar] [CrossRef]
- Mastrolonardo, G.; Certini, G.; Krebs, R.; Forte, C.; Egli, M. Effects of fire on soil organic matter quality along an altitudinal sequence on Mt. Etna, Sicily. Catena 2013, 110, 133–145. [Google Scholar] [CrossRef]
- Bahr, A.; Ellström, M.; Schnoor, T.K.; Påhlsson, L.; Olsson, P.A. Long-term changes in vegetation and soil chemistry in a calcareous and sandy semi-natural grassland. Flora 2012, 207, 379–387. [Google Scholar] [CrossRef]
- Tang, Z.; Xu, W.; Zhou, G.; Bai, Y.; Li, J.; Tang, X.; Chen, D.; Liu, Q.; Ma, W.; Xiong, G.; et al. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Zhang, C.; Tian, H.; Liu, J.; Wang, S.; Liu, M.; Pan, S.; Shi, X. Pools and distributions of soil phosphorus in China. Glob. Biogeochem. Cycles 2005, 19, GB1020. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, B.; Davis, M.; Sardans, J.; Peñuelas, J.; Billings, S. Long-term nitrogen deposition linked to reduced water use efficiency in forests with low phosphorus availability. New Phytol. 2015, 210, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.; Zhang, W.E.; Zhu, W.; Gundersen, P.E.; Fang, Y.; Li, D.; Wang, H.U. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Lauenroth, W.K. Relative abundance of plant functional types in grasslands and shrublands of North America. Ecol. Appl. 1996, 6, 1212–1224. [Google Scholar] [CrossRef]
- Carrera, A.L.; Sain, C.L.; Bertiller, M.B. Patterns of nitrogen conservation in shrubs and grasses in the Patagonian Monte, Argentina. Plant. Soil 2000, 224, 185–193. [Google Scholar] [CrossRef]
- Bertiller, M.B.; Sain, C.L.; Carrera, A.L.; Vargas, D.N. Patterns of nitrogen and phosphorus conservation in dominant perennial grasses and shrubs across an aridity gradient in Patagonia, Argentina. J. Arid Environ. 2005, 62, 209–223. [Google Scholar] [CrossRef]
- Shanmughavel, P.; Sha, L.; Zheng, Z.; Cao, M. Nutrient cycling in a tropical seasonal rain forest of Xishuangbanna, Southwest China. Part 1: Tree species: Nutrient distribution and uptake. Bioresour. Technol. 2001, 80, 163–170. [Google Scholar] [CrossRef]
- Zheng, Z.; Shanmughavel, P.; Sha, L.; Cao, M.; Warren, M. Litter decomposition and nutrient release in a tropical seasonal rain forest of Xishuangbanna, Southwest China. Biotropica 2006, 38, 342–347. [Google Scholar] [CrossRef]
- Liu, W.J.; Zhang, Y.P.; Li, H.M.; Liu, Y.H. Fog drip and its relation to groundwater in the tropical seasonal rain forest of Xishuangbanna, Southwest China: A preliminary study. Water Res. 2005, 39, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liu, W.; Li, P.; Duan, W.; Li, H. Dry season water uptake by two dominant canopy tree species in a tropical seasonal rainforest of Xishuangbanna, SW China. Agric. For. Meteorol. 2010, 150, 380–388. [Google Scholar] [CrossRef]
- Cao, M.; Zou, X.; Warren, M.; Zhu, H. Tropical forests of Xishuangbanna, China. Biotropica 2006, 38, 306–309. [Google Scholar] [CrossRef]
- Wang, W.F.; Qui, D.Y.; Wu, J.C.; Ye, H.M. The Soils of Yunnan; Yunnan Science and Technology Press: Kunming, China, 1996. (In Chinese) [Google Scholar]
- Lu, H.Z.; Wu, B.X.; Sha, L.Q.; Wang, J. Seasonal variation of soil respiration and its components in tropical rainforest and rubber plantation in Xishuangbanna, Yunnan. Chin. J. Appl. Ecol. 2009, 20, 2315–2322. [Google Scholar]
- Liu, W.; Luo, Q.; Li, J.; Wang, P.; Lu, H.; Liu, W.; Li, H. The effects of conversion of tropical rainforest to rubber plantation on splash erosion in Xishuangbanna, SW China. Hydrol. Res. 2015, 46, 168–174. [Google Scholar] [CrossRef]
- Cao, M.; Zhang, J.H.; Feng, Z.L.; Deng, J.W.; Deng, X.B. Tree species composition of a seasonal rain forest in Xishuangbanna, south-west China. Trop. Ecol. 1996, 37, 183–192. [Google Scholar]
- Zhang, J.H.; Cao, M. Tropical forest vegetation of Xishuangbanna SW China and its secondary changes, with special reference to some problems in local nature conservation. Biol. Conserv. 1995, 73, 225–238. [Google Scholar] [CrossRef]
- Mani, S.; Cao, M. Lagging effect of nutrient release in tropical seasonal forest soils in Xishuangbanna, south-west China. J. Trop. For. Sci. 2016, 28, 217–226. [Google Scholar]
- Arunachalam, A.; Arunachalam, K.M.; Pandey, H.N.; Tripathi, R.S. Fine litterfall and nutrient dynamics during forest regrowth in the humid subtropics of north-eastern India. For. Ecol. Manag. 1998, 110, 209–219. [Google Scholar] [CrossRef]
- Dutta, R.K.; Agarwal, M. Litterfall, litter decomposition and nutrient release in five exotic plant species planted on coal mine spoils. Pedobiologia 2001, 45, 298–312. [Google Scholar] [CrossRef]
- Daws, M.I.; Mullins, C.E.; Burslem, D.F.R.P.; Paton, S.R.; Dalling, J.W. Topographic position affects the water regime in a semi deciduous tropical forest in Panama. Plant. Soil 2002, 238, 79–90. [Google Scholar] [CrossRef]
- Gibbons, J.M.; Newbery, D.M. Drought avoidance and the effect of local topography on trees in the understorey of Bornean lowland rain forest. Plant. Ecol. 2003, 164, 1–18. [Google Scholar] [CrossRef]
- Baltzer, J.L.; Thomas, S.C.; Nilus, R.; Burslem, D.F.R.P. Edaphic specialization in tropical forest trees: Physiological correlates and responses to reciprocal transplantation. Ecology 2005, 86, 3048–3062. [Google Scholar] [CrossRef]
- Facelli, J.M.; Pickett, S.T.A. Plant litter: Its dynamics and effects on plant community structure. Bot. Rev. 1991, 57, 1–32. [Google Scholar] [CrossRef]
- Maass, J.M.; Martínez-Yrízar, A.; Patińo, C.; Sarukhán, J. Distribution and annual net accumulation of above-ground dead phytomass and its influence on throughfall quality in a Mexican tropical deciduous forest ecosystem. J. Trop. Ecol. 2002, 18, 821–834. [Google Scholar] [CrossRef]
- Tang, J.W.; Cao, M.; Zhang, J.H.; Li, M.H. Litterfall production, decomposition and nutrient use efficiency varies with tropical forest types in Xishuangbanna, SW China: A 10-year study. Plant. Soil 2010, 335, 271–288. [Google Scholar] [CrossRef]
- Paoli, G.D.; Curran, L.M. Soil nutrients limit fine litter production and tree Growth in mature lowland forest of southwestern Borneo. Ecosystems 2007, 10, 503–518. [Google Scholar] [CrossRef]
- Dent, D.H.; Bagchi, R.; Robinson, D.; Majalap-Lee, N.; Burslem, D.F. Nutrient fluxes via litterfall and leaf litter decomposition vary across a gradient of soil nutrient supply in a lowland tropical rain forest. Plant. Soil 2006, 288, 197–215. [Google Scholar] [CrossRef]
- Zhou, G.; Guan, L.; Wei, X.; Zhang, D.; Zhang, Q.; Yan, J.; Wen, D.; Liu, J.; Liu, S.; Huang, Z.; et al. Litterfall production along successional and altitudinal gradients of subtropical monsoon evergreen broadleaved forests in Guangdong, China. Plant. Ecol. 2007, 188, 77–89. [Google Scholar] [CrossRef]
- Chave, J.; Navarrete, D.; Almeida, S.; Álvarez, E.; Aragão, L.E.O.C.; Bonal, D.; Châtelet, P.; Silva-Espejo, J.E.; Goret, J.-Y.; von Hildebrand, P.; et al. Regional and seasonal patterns of litterfall in tropical South America. Biogeosciences 2010, 7, 43–55. [Google Scholar] [CrossRef]
- Townsend, A.R.; Cleveland, C.C.; Asner, G.P.; Bustamante, M. Controls over foliar N:P ratios in tropical rain forests. Ecology 2007, 88, 107–118. [Google Scholar] [CrossRef]
- Schuur, E.A.G.; Matson, P.A. Net primary productivity and nutrient cycling across a mesic to wet precipitation gradient in Hawaiian montane forest. Oecologia 2001, 128, 431–442. [Google Scholar] [CrossRef]
- Santiago, L.S.; Schuur, E.A.G.; Silvera, K. Nutrient cycling and plant-soil feedbacks along a precipitation gradient in lowland Panama. J. Trop. Ecol. 2005, 21, 461–470. [Google Scholar] [CrossRef]
- Young, A. Tropical Soils and Soil Surveys; Cambridge University Press: Cambridge, UK, 1976; p. 468. [Google Scholar]
- Laurance, W.F.; Fearnside, P.M.; Laurance, S.G.; Delamonica, P.; Lovejoy, T.E.; Rankin-de Merona, J.M.; Chambers, J.Q.; Gascon, C. Relationship between soils and Amazon forest biomass: A landscape-scale study. For. Ecol. Manag. 1999, 118, 127–138. [Google Scholar] [CrossRef]
- Xu, W.; Liu, L.; He, T.; Cao, M.; Sha, L.; Hu, Y.; Li, Q.; Li, J. Soil properties drive a negative correlation between species diversity and genetic diversity in a tropical seasonal rainforest. Sci. Rep. 2016, 6, 20652. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.Y.; Tang, J.W.; Sha, L.Q.; Meng, Y. Soil nutrient contents and their characteristics of seasonal changes under Shorea chinensis forest in Xishuangbanna. Acta Phytoecol. Sin. 2003, 27, 373–379. [Google Scholar]
- Davies, J.; Tipping, E.; Rowe, E.; Boyle, J.; Graf Pannatier, E.; Martinsen, V. Long-term P weathering and recent N deposition control contemporary plant-soil C, N, and P. Glob. Biogeochem. Cycle 2016, 30, 231–249. [Google Scholar] [CrossRef]
- Augusto, L.; Achat, D.L.; Jonard, M.; Vidal, D.; Ringeval, B. Soil parent material-A major driver of plant nutrient limitations in terrestrial ecosystems. Glob. Chang. Biol. 2017, 23, 3808–3824. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, M.; Huang, Z.; Lin, T.C.; Vadeboncoeur, M.A.; Searle, E.B.; Chen, H.Y. Temporal changes in soil C-N-P stoichiometry over the past 60 years across subtropical China. Glob. Chang. Biol. 2018, 24, 1308–1320. [Google Scholar] [CrossRef]
- Martinelli, L.A.; Piccolo, M.C.; Townsend, A.R.; Vitousek, P.M.; Cuevas, E.; McDowell, W.; Robertson, G.P.; Santos, O.C.; Treseder, K. Nitrogen stable isotopic composition of leaves and soil: Tropical vs. temperate forests. Biogeochemistry 1999, 46, 45–65. [Google Scholar] [CrossRef]
Figure 1.
Measured monthly precipitation (mm) and temperature (°C) at the study site.

Figure 2.
Litterfall in relation to precipitation. Solid circles represent the site primary tropical forest (PTF) and empty circles the site secondary tropical forest (STF).
Figure 2.
Litterfall in relation to precipitation. Solid circles represent the site primary tropical forest (PTF) and empty circles the site secondary tropical forest (STF).

Figure 3.
Leaf litter N and P, and soil N and P in relation to precipitation and temperature. pH, leaf litter N, leaf litter P, soil N, and soil P in relation to precipitation (a–e, respectively) and temperature (f–j, respectively). Solid trend lines represent the site PTF and dotted lines the site STF.
Figure 3.
Leaf litter N and P, and soil N and P in relation to precipitation and temperature. pH, leaf litter N, leaf litter P, soil N, and soil P in relation to precipitation (a–e, respectively) and temperature (f–j, respectively). Solid trend lines represent the site PTF and dotted lines the site STF.


Figure 4.
Leaf litter N (a,a1) and P (b,b1), and soil N (c,c1) and P (d,d1) in the (i) dry and (ii) rainy season. Solid trend lines represent the site PTF and dotted lines the site STF.
Figure 4.
Leaf litter N (a,a1) and P (b,b1), and soil N (c,c1) and P (d,d1) in the (i) dry and (ii) rainy season. Solid trend lines represent the site PTF and dotted lines the site STF.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of mean variables from the study sites.
| Variable (n = 12) | Primary Tropical Forest (PTF) | Secondary Tropical Forest (STF) | t-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean ± SE | CV (%) | Skewness | K-S Test | Mean ± SE | CV (%) | Skewness | K-S Test | ||
| Precipitation (mm year−1) | 1421.5 ± 9.8 | 90.5 | 0.69 | 0.20 | 1421.5 ± 9.8 | 90.5 | 0.69 | 0.20 | - |
| Temperature (°C) | 22.4 ± 0.8 | 16.5 | −0.61 | 0.27 * | 22.4 ± 0.8 | 16.5 | −0.61 | 0.27 * | - |
| pH | 4.4 ± 0.1 a | 4.2 | 0.34 | 0.23 | 4.2 ± 0.1 b | 3.5 | 0.51 | 0.16 | 2.83 ** |
| Total litterfall (t ha−1 yr−1) | 6.03 ± 0.5 a | 44.3 | 0.83 | 0.18 | 8.3 ± 0.50 a | 37.8 | 1.20 | 0.20 | −1.91 |
| Leaf litter N (g kg−1) | 15.8 ± 0.4 a | 8.9 | 0.35 | 0.10 | 15.8 ± 0.3 a | 6.3 | 1.28 | 0.18 | −0.04 |
| Leaf litter P (g kg−1) | 0.6 ± 0.1 a | 10.3 | 0.13 | 0.15 | 0.7 ± 0.1 b | 6.7 | −0.48 | 0.19 | −2.67 ** |
| Soil N (g kg−1) | 2.1 ± 0.1 a | 3.9 | −0.82 | 0.20 | 2.6 ± 0.2 b | 9.4 | 1.24 | 0.25 * | −7.09 *** |
| Soil P (g kg−1) | 0.3 ± 0.03 a | 4.9 | −0.05 | 0.13 | 0.4 ± 0.04 b | 5.8 | −0.12 | 0.18 | −12.77 *** |
* Significant at p < 0.05; ** p < 0.01; *** p < 0.0001. Different lowercase letters (a and b) indicate that means significantly differ using t-test, SE—Standard Error, CV—Co-efficient of variation, K-S test—Kolmogorov–Smirnov test.
Table 2.
Correlations (Pearson’s) between climate (PPT and TEMP) and leaf litter; and soil nutrient pools from the study sites.
Table 2.
Correlations (Pearson’s) between climate (PPT and TEMP) and leaf litter; and soil nutrient pools from the study sites.
| Variable (n = 12) | PPT | TEMP | pH | LF-PTF | LF-STF | Leaf Litter N | Leaf Litter P | Soil N | Soil P |
|---|---|---|---|---|---|---|---|---|---|
| PPT | 1 | ||||||||
| TEMP | 0.81 * | 1 | |||||||
| pH | 0.23 | 0.50 | 1 | ||||||
| LF-PTF | −0.17 | 0.27 | 0.60 * | 1 | |||||
| LF-STF | 0.54 | 0.60 * | 0.16 | 0.37 | 1 | ||||
| Leaf litter N | 0.79 * | 0.68 * | −0.12 | −0.18 | 0.54 | 1 | |||
| Leaf litter P | 0.77 * | 0.43 | −0.20 | −0.46 | 0.43 | 0.71 * | 1 | ||
| Soil N | 0.01 | −0.01 | 0.14 | −0.12 | −0.15 | 0.28 | 0.07 | 1 | |
| Soil P | 0.32 | 0.19 | −0.22 | −0.47 | 0.31 | 0.54 | 0.48 | 0.52 | 1 |
* Significant at p < 0.05; PPT—Precipitation, TEMP—Temperature, LF—Litterfall, PTF—Primary Tropical Forest, STF—Secondary Tropical Forest.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mani, S.; Cao, M. Nitrogen and Phosphorus Concentration in Leaf Litter and Soil in Xishuangbanna Tropical Forests: Does Precipitation Limitation Matter? Forests 2019, 10, 242. https://doi.org/10.3390/f10030242
AMA Style
Mani S, Cao M. Nitrogen and Phosphorus Concentration in Leaf Litter and Soil in Xishuangbanna Tropical Forests: Does Precipitation Limitation Matter? Forests. 2019; 10(3):242. https://doi.org/10.3390/f10030242
Chicago/Turabian StyleMani, S., and Min Cao. 2019. "Nitrogen and Phosphorus Concentration in Leaf Litter and Soil in Xishuangbanna Tropical Forests: Does Precipitation Limitation Matter?" Forests 10, no. 3: 242. https://doi.org/10.3390/f10030242
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.