1. Introduction
Populus tremula L. (known as common, European, or Eurasian aspen) has the largest native range of any species from the
Populus genus, being one of the most widely-distributed trees globally [
1]. This species occurs from 40° to 70° N latitude [
2] on the Eurasian continent, across a wide variety of soils, elevations, and climatic conditions. In Poland, common aspen is considered as the only forest species of the
Populus genus. Being an early successional species,
P.
tremula L. plays an important role in the first generation of a forest on previously non-forested areas, or following considerable forest disturbances. Tree stands with common aspen as the dominant tree species cover only 0.8% of all forest area in Poland; however, owing to its high ecological value,
P. tremula constitutes a respected supplementary (co-occurring) tree species. In addition, because of its relatively short life span,
P. tremula provides numerous birds and mammals with habitats and food, thereby contributing to the enhancement of forest biodiversity. Aspen’s advantage over all other native tree species of Poland manifests however in large part through its rapid growth, especially in the juvenile phase. Through its favorable wood properties, low lignin, and high carbohydrate content along with fibers of small diameter and thin wall,
P. tremula comprises a suitable and desirable tree species for the production of pulp and paper [
3]. These qualities appear to be particularly significant in consideration of the projections showing that paper production is expected to increase worldwide from the current value of 400 million Mg annually, to 700–900 million Mg by 2050 [
4]. Thus, plantations of fast-growing aspen may represent a promising source of wood for satisfying the increasing demand for wood-based products, providing the opportunity to reduce the timber harvest from natural forests. Moreover, the latter possibility provides added incentive to improve both the growth rates and wood properties of fast-growing species such as aspen, because of the limited wood resources that have been increasingly exploited for papermaking, as well as other cellulose-based products and timber purposes.
Taking into account the considerable genetic diversity in natural populations of aspen and its predisposition to hybridization, the most promising results of aspen improvement might be achieved through breeding, inter-specific hybridization, and cloning [
5,
6,
7,
8,
9]. Hybrid aspens, in particular the offspring of geographically distant species (e.g.,
P. tremula with
P. tremuloides Michaux), demonstrate superior performance over the average of both their genetically distinct parents [
10]. This phenomenon, known as heterosis, or hybrid vigor, constitutes a multi-genetic complex trait, and can be extrapolated as the sum of multiple physiological and phenotypic traits, including the magnitude and rate of growth, flowering time, yield, and resistance to biotic and abiotic environmental factors [
11,
12]. However, despite the ability of numerous studies to achieve desirable increases in the growth of F
1 hybrids of
Populus spp. including aspen [
13,
14], the genetic mechanism underlying heterosis remains incompletely understood.
The hybrid vigor phenomena in aspen breeding began to be widely utilized in Europe in the early 20th century [
10,
15,
16,
17,
18]. At the end of the 1950s,
P. tremula ×
P. tremuloides hybrids were already produced in almost all European countries [
17], as well as in the USA and Canada [
8,
19]. In Poland, the first studies on poplar hybridization within the
Populus section began in 1953 [
10]. The main objective was to cross aspen (
P. tremula) with native white poplar (
P. alba L.) or trembling (American) aspen (
P. tremuloides). Concurrently, an independent breeding program for aspen was also initiated in the Polish forestry. Although the goals of this program were never fully realized, over 50 trees were phenotypically selected, the majority of which originated from the Białowieża Forest. Owing to their high qualitative and quantitative value, these selected
P. tremula genotypes were used as the mother trees in the hybridization program for control crosses with
P. tremuloides or
P. alba as the paternal parents.
The primary factor that supports the improvement and testing of a fast-growing species as a feedstock source for papermaking, as well as renewable energy, is based on the restrictions placed on the harvesting of natural forests, stemming from ecological needs and social expectations concerning the use of forests. Nevertheless, fast-growing aspen and its hybrids have attracted relatively little research attention in Poland.
The aims of this study were therefore to (1) evaluate the productivity of aspen and its hybrids in the environmental conditions of Poland, and (2) investigate the effects of P. tremula with P. tremuloides and P. alba hybridization on the growth and wood quality for papermaking purposes, in the context of further genotypes selection and developing recommendations for industrial application.
2. Materials and Methods
2.1. Experimental Location, Soil, and Climatic Condition
The experimental area of the study was located in northern Poland (54°4ʹ26ʺ N, 20°30ʹ4ʺ E) in the proximity of Lidzbark Warmiński. Average annual temperature is 8.0 °C with an annual precipitation of 683 mm. The experiment was initiated in April 2011 on post-agricultural land. According to the World Reference Base for Soil Resources (WRB) the main soil type was Luvisol. The soil texture was determined as sandy clay with a pH in H2O of 4.7–5.0 (acidic), and the C:N ratio of 8.7–9.3 in the top 20 cm. The soil preparation was done by plowing, and the saplings were planted in holes, created by an earth auger powered by a tractor.
2.2. Planting Material
Planting material consisted of 15 hybrid aspen clones of
P. tremula × P. tremuloides (TA × TE), four hybrid clones of
P. tremula × P. alba (TA × A), and one clone of
P. tremula (TA × TA)
, which performance was tested previously in Germany. Additionally, a mixture of 30 clones of
P. tremula (TA) plus trees from wild populations in the Białowieża Forest was used as a reference (
Table 1). Hybrid clones had been crossed and selected in Poland and Germany. The German genotypes were propagated by tissue culture. Polish clones used in the experiment were crossed and selected at the Forest Research Institute, and were propagated vegetatively from root cuttings. Prior to planting, all saplings were maintained for one year in the forest nursery.
2.3. Study Design
The study layout comprised a randomized complete block design with four replicates. Each block was divided into 21 plots equal to the number of the tested group of clones. A total of 25 saplings of a given clone were planted within each plot with a spacing of 2.5 × 3.0 m, resulting in a planting density of 1333 saplings ha−1. A bordering row was planted around the experimental area. The area was fenced to prevent browsing by wild animals. During the first three years, the plantation was weeded mechanically once annually. No irrigation or fertilization was applied to the tested area.
2.4. Measurement of Tree Characteristics
The diameter at breast height (DBH; measured at a height of 1.3 m) of all trees was measured after four, five, six, and seven growing seasons on the plantation. The basal area for each tree was calculated, based on its DBH. Height was recorded for 20% of trees systematically in each plot and year. The height curve was constructed separately for each clone in a given block and year, according to the following function [
20]:
where
h represents tree height (m),
DBH is the diameter at breast height (cm), and
α,
β are the fitted coefficients.
The estimated coefficients (
α,
β) of the regression function for each clone in each block were used to estimate the height of trees from the entire range of DBH, which was utilized in the volume equation. Tree volume was calculated based on the function developed by Wróblewski and Zasada [
21] based on the volume table of Orłow (elaborated by Czuraj [
22]):
where
V represents an individual tree stem volume,
h is tree height, and
DBH is breast height diameter.
Based on individual tree volumes, we calculated tree volume on an area basis and the mean annual increment (MAI) for each clone at a given age. The survival rate was assessed based on the number of living trees.
2.5. Properties of Cellulosic Pulp
At the age of seven years, three clones representing all species combination types and characterized by the good performance (‘Wä 13′ TA × TE, ‘IBL 91/78′ TA × A, and ‘W 3′ TA) were selected to evaluate the properties of cellulosic pulp. From each selected clone in a given replication, three trees (total number 9 trees per clone), representing average parameters, were felled. The ‘average trees’ were identified according to the basal area of trees. From harvested trees, 70 cm long samples were taken at the middle of every 2 m section, beginning from the stem base to the tree top. The total volume of the samples represented approximately 50% of the volume of the whole stem. All samples were numbered and transferred to the laboratory immediately after collection. The wood samples were manually debarked and deprived of knots, and subsequently sawn into chips using an electric Milwaukee MD 304 saw (Milwaukee Electric Tool Corporation, Hilden, Germany).
Poplar cellulosic pulp was prepared by the Kraft method [
23,
24] from air-dried woodchips (25 mm × 15 mm × 8 mm). Dry weight (d.w.) of all materials was determined prior to pulping. Pulping processes were conducted in a 0.015 m
3 stainless steel reactor with regulated temperature (PD-114, Danex, Rosko, Poland) with agitation (3 swings min
−1, swinging angle of 60°). Suspensions of woodchips (1000 g d.w. of wood chips in 0.004 m
3 of alkaline sulfate solution) were heated for 120 min to achieve the temperature of 165 °C, and incubated at this temperature for a further 120 min. Then, the temperature was decreased to ambient temperature (22 ± 1 °C) using a jacket with cold tap water, and the insoluble residue was separated by filtration within the reactor, washed with approximately 0.050 m
3 of demineralized water, and incubated overnight (12 h) in demineralized water to remove the residues of the alkali-soluble fractions.
The obtained fibrous biomass was disintegrated using a laboratory JAC SHPD28D propeller pulp disintegrator (Danex, Rosko, Poland) at 10,000 rpm and screened using a PS-114 membrane screener (Danex, Rosko, Poland) (0.2 mm gap). Fibers were collected, dried for 48 h in ambient conditions (21 ± 1 °C), then weighed. Analysis of the chemical composition of cellulosic pulps included a quantification of extractives, lignin, cellulose, hemicelluloses, and ash. The lignin mass fraction was determined by a gravimetric method in compliance with the TAPPI T222 standard after the removal of extractives (TAPPI T204). The holocellulose mass fraction was determined in accordance with the TAPPI T249 standard. Cellulose was quantified as alpha cellulose (TAPPI T203). The hemicelluloses mass fraction was calculated as the difference between the holocellulose and cellulose mass fractions. Ash content was determined by a gravimetric method in compliance with TAPPI T211. All these assays were performed in triplicate for each raw material.
2.6. Properties of Paper Sheets
Sheets of paper were produced under laboratory conditions from rewetted pulp samples (22.5 g d.w. samples were soaked in water for at least 8 h) that were subjected to disintegration using the laboratory JAC SHPD28D propeller pulp disintegrator at 20,000 rpm, according to the ISO 5263-1 (2004) standard. The disintegrated pulps were concentrated to a dry weight mass fraction of 10% and refined in a JAC 12DPFIX PFI mill (Danex, Rosko, Poland) under standard conditions (ISO 5264-2 (2011)). The ultimate standard freeness of the pulp was approximately 30° Schopper-Riegler. Water retention values were determined according to the ISO 23714 (2014) standard. Dimensions of fibers were measured using a Morfi Compact Black Edition apparatus (Techpap, Saint Martin d′Hères, France) according to ISO 16065-2 (2014).
Paper sheets (grammage of approximately 80 g m−2) were produced from the refined and unrefined pulps, using a Rapid-Koethen class apparatus (Danex, Rosko, Poland) according to ISO 5269-2 (2004). Mechanical properties of paper were determined only for the sheets with a grammage of 80 ± 1 g m−2.
The sheets were stored for 24 h at the relative moisture mass fraction of 50 ± 2% and temperature of 23 ± 1 °C (ISO 187 (1990)), prior to the determination of mechanical properties such as tensile index, stretch, tensile energy absorption (ISO 1924-2 (2008)), and tear index (ISO 2758 (2014)). Other parameters measured included bulk (ISO 534 (2011)), brightness (TAPPI T452), and opacity (TAPPI T519).
2.7. Data Analysis
To assess differences in biometric characteristics and productivity (volume and MAI) between clones and the interaction between clones over time, generalized linear mixed models with a log link function and normal-distributed errors were used (the GLIMMIX procedure). The choice of the optimal model was based on the Akaike Information Criterion. The model was expressed using the following equation:
where
represents the general mean,
b is the mean of random effect,
is the
j-clone effect,
is the
k-year effect,
represents the
kj-interaction between clones and time, and
Bl is the
l-random block effect. The log link function represented the multiplicative form of the explanatory variables. The type III tests of fixed effects were used to determinate the significance of specified effects. The size of these effects was calculated as means ratios according to the base level achieved for TA clones from wild populations (reference) in Białowieża Forest. The same generalized linear mixed models and subsequent procedures were implemented for comparisons between groups of different species crosses (clones were grouped in accordance with their parental species). In this case, the pure species TA (= clone mixture from wild populations in the Białowieża Forest and clone ‘W 3′) was used as a reference.
The papermaking traits between species crosses were compared using Analysis of Variance (ANOVA) with the PROC GLM procedure. The model assumed:
where
represents a dependent variable, μ is a general mean,
Sj is a
j-clone effect, and ε
i is a random component from the normal distribution. Such model choice was dictated by the fact that data were collected on an annual basis (after seven years of tree growth), with the results relating to optimized processes (without considering the block effect). The
post-hoc comparisons between clones/species combination types were performed using the Tukey HSD test. Moreover, the SAS CORR procedure was used to determine the correlations between papermaking traits for clones. All statistical analyses were performed with SAS/STAT (rel. 14.3) statistical package (SAS Institute Inc., Cary, NC, USA).
4. Discussion
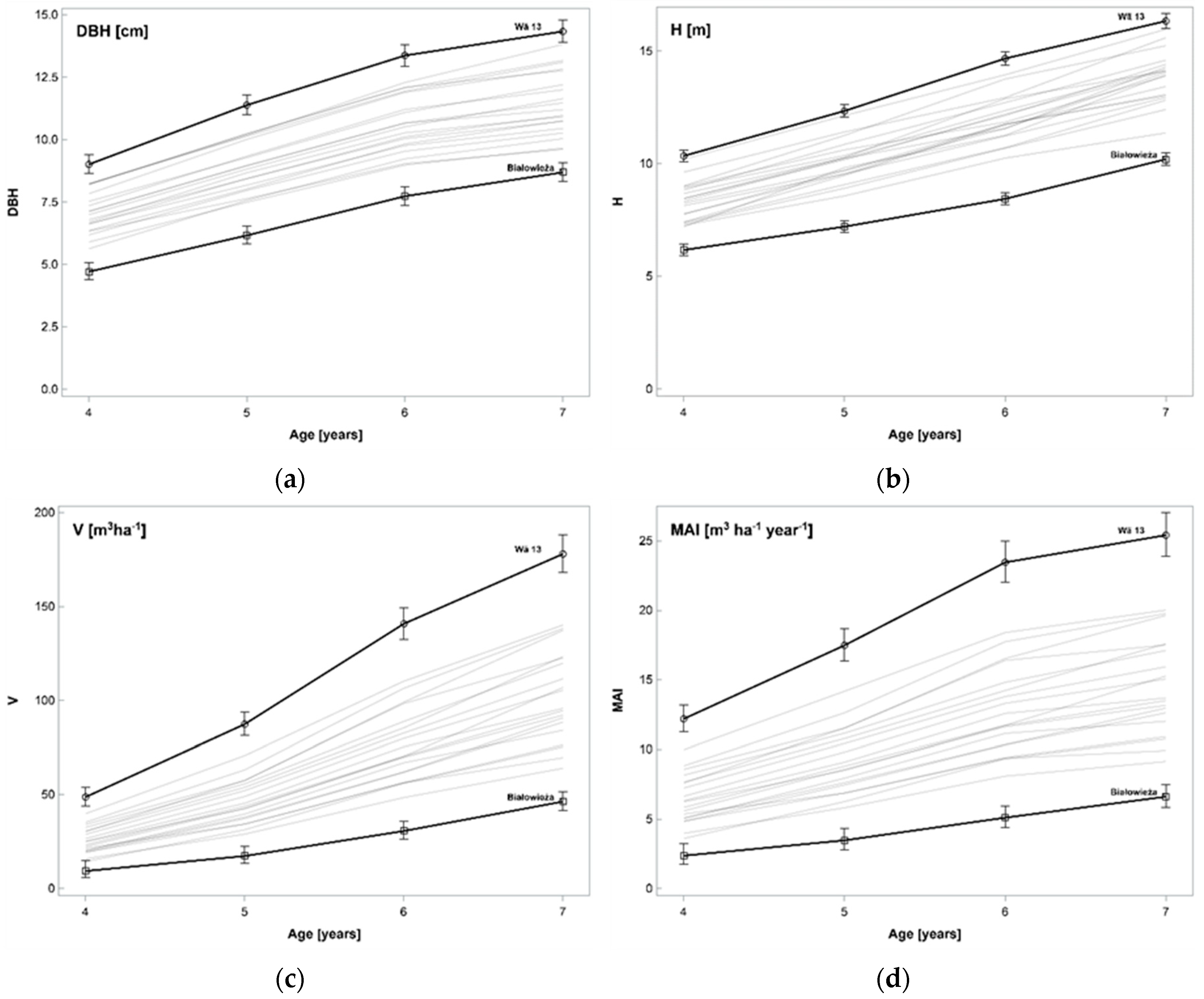
Aspen is among the most widely used poplars for papermaking purposes in the Northern Hemisphere. Because hybrid aspens may benefit from heterosis, the majority of practically oriented hybridization programs are aimed at using this phenomenon for the improvement of growth and wood quality parameters. This is also confirmed by the present study, which showed that during a seven-year long experiment, all hybrid aspen clones were characterized by superior growth and productivity in comparison with those of wild species.
The MAI of the stem wood of the best performing clone ‘Wä 13′ (TA × TE) was at the highest expected level for climatic conditions of the Baltic Region, which encompasses central and northern European countries. Numerous studies have reported that the MAI of hybrid aspen in the southern part of the Nordic region and the Baltic area may reach up to 20 m
3 ha
−1 year
−1 during rotation periods of 20–25 years [
25,
26,
27,
28,
29,
30,
31]. The highest MAI measured to date, 25.8 m
3 ha
−1 year
−1, was recorded for a 23 year old hybrid aspen (TA × TE) plantation in southern Sweden [
32]. For hybrid aspen trials established in north-western Germany, the modeled MAI reached its maximum at the age of 24 years [
33]. In the present study, the majority of aspen hybrid clones examined at the age between four and seven years demonstrated a high MAI, with that for the best clone amounting to 25 m
3 ha
−1 year
−1 at the age of seven years, which progressed in each consecutive year of the analysis. Considering that the MAI for aspen plantations is not expected to culminate before the age of 20 years [
34], we can anticipate further improvement of this MAI in subsequent years.
In Poland, a rotation period of native stands of
P. tremula typically comprises 40 years, which in the present case can be assigned to vegetatively propagated offspring of wild populations from the Białowieża Forest. Taking into account that in the present study inter-specific crosses resulted in 1.4-fold greater mean annual increment, as compared to the clones of trees from wild populations of TA, and ranging from 1.6- to 4.6-fold enhancement for particular hybrid clones, in comparison to the reference, our data suggest that the rotation age for selected hybrid aspen clones might be reduced to 20 years. Similar conclusions were provided by Li et al. [
8], who postulated that the expected rotation for aspen clones in the north-eastern part of the United States might also be limited to 20 years. This remains consistent with the considered periods of 30 years in the Swedish short-rotation program [
35] along with the estimated period for reaching merchantable volumes of American aspen crosses in Alberta, Canada [
5].
The reduced rotation periods for superior hybrid aspen clones may complement traditional, multifunctional forest management with the long-rotation periods, widely implemented in Central Europe. Shortened production cycles may serve as a partial remedy for the loss of stability of forest ecosystems in conditions of rapidly progressing climate change. Over the period 1951–2008, the mean temperature in Poland increased by 0.24 °C per 10 years [
36]. Climate change not only involves directional changes in the mean values of climatic variables (e.g., global warming) but the frequency and magnitude of various extreme climatic events [
37,
38]. In many regions of Europe, extreme weather events (heat waves, heavy precipitation, droughts, heavy storms) are likely to intensify in the coming decades [
39,
40]. In the case of hurricanes/storms, which in recent years frequently cause serious damages in Polish and in European forestry [
41], it was proved that a greater stand age and a taller stand height increase stand susceptibility [
41,
42,
43].
Replacement of such high-risk stands, and associated economic losses, can be made by introduction of stands with shortened production cycles, which have also the potential to significantly contribute to both increasing biomass supply and capturing carbon from the atmosphere [
44,
45,
46,
47].
Understanding the growth increment curve of various tree species in the juvenile period holds exceptional importance for silviculture [
48], determining e.g., the choice of spacing and term of first thinning. Among fast-growing species, aspen is considered to achieve the highest annual increment, albeit somewhat later than poplars from the
Aigeiros or
Tacamahaca sections or species of the
Salix genus [
49]. However, the stem volume produced by hybrid aspen in northern Poland at the age of seven years did not reflect inferior productivity in comparison to that of hybrid poplars at the same age, growing in the same soil and climatic condition [
50]. As highlighted by Liesebach et al. [
51], considerable growth increment is produced by aspen from the age of 6 years onward with the differences occurring at a very early stage of tree growth. This may explain the comparable results of productivity achieved by species crosses from the
Populus section in the present study with those of the different crosses among the
Aigeiros or
Tacamahaca sections.
Taking into account differences in productivity among various parental species and combinations of species within the Populus section, we revealed that inter-specific hybrids of TA × TE and TA × A performed significantly better than vegetative progenies of TA. Notably, however, on the clone level, statistically significant differences between the selected TA clone ‘W 3′, and other clones of inter-specific hybrid aspen crosses, in terms of growth parameters, were not found. This finding suggests that selection and testing genotypes in the area of the future utilization may result in good performing clones, with adaptive ability to specific climatic and soil conditions. Furthermore, it seems reasonable to add the clone ‘W 3′ to further stages of the tree improvement program, thereby enriching the native population of P. tremula, and providing new genotypes for future mating strategies.
Hybrid aspen clones were characterized by more desirable properties related to papermaking traits than those of the clone of the pure species TA. Moreover, the chemical composition of the cellulosic pulp was diversified among the clones. It is notable that the cellulose mass fraction was greater in the pulps of both hybrid aspen clones than that in the pulp of the pure species, indicating the enhanced desirability of the former. This result impacts yield and Kappa number, as reflected in the positive correlation with the cellulose mass fraction and a negative correlation with the lignin mass fraction. As indicated by Hart et al. [
5], the further consequences of particular carbohydrate concentrations can be revealed in ethanol biofuel applications. Therefore, the obtained differences in chemical composition (yield) between clones may represent important factors when considering the utilization of the biomass of a particular clone for paper and biofuel production at the industrial scale.
The present results regarding the absolute values of screened yield and Kappa number are comparable with the findings of several previous studies. It has to be mentioned however, that a much higher yield of hardwood pulps can be achieved by trees older than those we tested in our study [
52]. When considering yields and characteristics of cellulosic pulps obtained from multiple softwood and hardwood species, along with fast growing grasses in Poland [
24], the yield of cellulosic pulp from aspen clones obtained in the present study confirms their high suitability for pulp production. In comparison, the only alternatives for hybrid aspen pulp in Poland appear to comprise pulps obtained from silver birch (
Betula pendula Roth.) and the hybrid poplar
P. maximowiczii ×
P. trichocarpa ‘NE-42′ (syn. ‘Hybrid 275′).
Fiber properties of hybrid aspen clones were enhanced and more desirable, compared to those of pure species fibers, albeit they remained typical for the
Populus section [
45]. According to Francis et al. [
53], 31 year old aspens produced much longer fibers than those of 7.5 year old poplars. Consequently, fiber and strength properties are expected to increase with age. This observation highlights the need for a reassessment of pulp and paper properties at the end of the rotation period in the experimental plantation in northern Poland. The differences in fiber morphology between aspen crosses resulted in different physical properties of paper sheets, which is consistent with the results of a previous study by Gurnagul et al. [
54].
Generally, sheet properties were affected by fiber morphology in manners that could be predicted from the principles of paper physics [
54,
55]. In particular, tearing resistance, as the most important strength parameter, was directly correlated with fiber length.
5. Conclusions
Overall, the high evaluated productivity identified in the present study clearly indicated that short rotation plantation of hybrid aspen might be considered as an important additional source of woody biomass for pulp and paper products in Poland. Such plantations may complement traditional, multifunctional forest management, in which conservation approaches play an increasing role. The reduced rotation periods for superior hybrid aspens would likely contribute to an increase in economic benefits, and partly mitigate the uncertainty accompanying the long-rotation periods applied in traditional silviculture. In addition, this aspect appears to be particularly important with regard to rapid climate change and its impact on forests.
Furthermore, significant improvement of hybrid aspen traits related to growth, pulp, and paper properties was observed in the present study, which dominated over those of pure species. The effect of hybrid vigor manifested as 1.4-fold greater MAI compared to that of wild species, with particular hybrid clones exhibiting a 1.6- to 4.6-fold enhancement. The superiority of hybrid aspen also translated into papermaking properties, as the chemical properties of the pulp, yield, fiber morphology, and physical properties of the final paper sheets were more desirable for P. tremula hybrids with both P. tremuloides and P. alba, than those for pure P. tremula. Together, our findings confirmed that a good knowledge of the maximal growth parameters that could be achieved with particular site conditions; species, or hybrid selection, combined with the concise relationship between chemistry, fiber morphology, and sheet properties, could facilitate the optimal clone choice for each specific region for industry purposes.

 ,
, 



{kind=link}
{kind=link}
{kind=link}