Responses of Korean Pine to Proactive Managements under Climate Change
by
 ,
,
Kai Liu
1, 
Hongshi He
1,2,*,
Wenru Xu
2,
Haibo Du
1,
Shengwei Zong
1,
Chao Huang
3,
Miaomiao Wu
1,
Xinyuan Tan
1 and
Yu Cong
4 1
Key Laboratory of Geographical Processes and Ecological Security in Changbai Mountains, Ministry of Education, School of Geographical Sciences, Northeast Normal University, Changchun 130024, China
2
School of Natural Resources, University of Missouri, 203 ABNR Bldg, Columbia, MO 65211, USA
3
CAS Key Laboratory of Forest Ecology and Management, Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang 110016, China
4
Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China
*
Author to whom correspondence should be addressed.
Forests 2020, 11(3), 263; https://doi.org/10.3390/f11030263
Submission received: 21 December 2019
/
Revised: 21 February 2020
/
Accepted: 25 February 2020
/
Published: 27 February 2020
(This article belongs to the Special Issue Adaptive Forest Management and Decision Making Models under Climate Change (Risk and Uncertainty))
Abstract
:Proactive managements, such as the resistant and the adaptive treatments, have been proposed to cope with the uncertainties of future climates. However, quantifying the uncertainties of forest response to proactive managements is challenging. Korean pine is an ecologically and economically important tree species in the temperate forests of Northeast China. Its dominance has evidently decreased due to excessive harvesting in the past decades. Understanding the responses of Korean pine to proactive managements under the future climates is important. In this study, we evaluated the range of responses of Korean pine to proactive managements under Representative Concentration Pathway (RCP) 8.5 scenarios from four General Circulation Models (GCMs). We coupled an ecosystem process-based model, LINKAGES, and a forest landscape model, LANDIS PRO, to simulate scenarios of management and climate change combinations. Our results showed that the resistant and the adaptive treatment scenarios increased Korean pine importance (by 14.2% and 42.9% in importance value), dominance (biomass increased by 9.2% and 25.5%), and regeneration (abundance <10 years old increased by 286.6% and 841.2%) throughout the simulation. Results indicated that proactive managements promoted the adaptability of Korean pine to climate change. Our results showed that the variations of Korean pine response to climate change increased (ranging from 0% to 5.8% for importance value, 0% to 4.3% for biomass, and 0% to 85.4% for abundance) throughout the simulation across management scenarios. Our result showed that regeneration dictated the uncertainties of Korean pine response to climate change with a lag effect. We found that the effects of proactive managements were site-specific, which was probably influenced by the competition between Korean pine and the rare and protected broadleaf tree species. We also found that the adaptive treatment was more likely to prompt Korean pine to migrate into its suitable habitats and promoted it to better cope with climate change. Thus, the adaptive treatment is proposed for Korean pine restoration under future climates.
1. Introduction
The leading edge of tree species limited by temperature are expected to migrate poleward and upward under warming change [1,2,3,4]. Species unsuitable for future climate could extirpate from current distribution, and forest composition might include new tree species adaptive to future climate [5,6]. Increasing evidence suggests that tree species may fail to keep up with the rate of climate change owing to their limited dispersal capacity and long regeneration cycle [7,8,9].
Management could hinder or reinforce the transition of forest composition in accordance with various objectives in the face of climate change [10,11]. Proactive managements, such as the resistant and the adaptive treatments, have been proposed to mediate the effects of future climates [10,12,13]. The resistant treatment reduces the occurrence of management to maintain existing forest composition, whereas the adaptive treatment increases the occurrence of management to promote species turnover [10,14]. Generally, thinning releases growing space for remaining trees and alleviates competition for resources [15], which may result in a high diversity of both composition and structure [16]. High diversity is hypothesized to enhance forest adaptive capacity under future uncertainties [16,17]. A recent study showed that proactive managements with diverse objectives could help with the adaptation of US Central Hardwood Forests to climate change [17]. Increasing frequency of disturbance catalyzed the adaptation of forest ecosystems to changing climate [18]. Likewise, frequent managements may promote forest adaptation [19,20]. However, there still exist numerous uncertainties in forest response to proactive managements, which is one of the impediments for the implementation of effective adaptation measures [21].
Korean pine is a dominant species in the mixed coniferous and broadleaf forests in the temperate forests of Northeast China. Meanwhile, Korean pine is a species with high economic value in providing timber and food for wildlife habitats [22]. However, Korean pine dominance has dramatically decreased as a consequence of extensive and excessive timber harvesting for nearly a half century [23,24]. Currently, early- and mid-succession species occupy most of the growing space in the mixed Korean pine and broadleaf forests except some natural reserves [25]. In general, Korean pine seed needs adequate light to generate [26,27]. Most young Korean pines die of light limitation as forest canopy has been closed for a period of time [23], resulting in poor regeneration of Korean pine [28,29]. Thus, restoring Korean pine is an urgent issue of management in the temperate forests of Northeast China.
Drought induced by warming has limited Korean pine growth [30]. Accumulating evidence revealed that temperatures in Northeast China have substantially increased and precipitation has slightly decreased since the 1950s [31,32,33], especially for the mixed Korean pine and broadleaf region. Dendrochronological studies showed that rising temperature and reduced precipitation could further limit Korean pine growth [5,34]. However, pressure from drought on Korean pine decreased with altitude and latitude increase [30]. A recent study showed that Korean pine radial growth decreased at lower elevations under climate change characterized by warming and drought, but increased at higher elevations [5]. Furthermore, it was reported that Korean pine growth decline induced by recent warming gradually decreased with latitude increase [35]. Recent studies revealed that warming climate altered the natural habitat of Korean pine and promoted it migrating northward [36,37]. These results indicated that Korean pine suitable distribution may shift upward and northward under warming climates [35].
Tree harvest might play a more significant role in species distribution than climate change [38,39]. A recent study suggested that partial harvest increased available light for oak regeneration while appearing to buffer against the effects of drought under a warming climate [40]. Likewise, tree harvest might mitigate Korean pine pressure from drought induced by increasing temperature and decreasing precipitation. A recent study showed that climate warming decreased Korean pine growth in the closed forest, but promoted the growth in forest gaps [41].
Future restoration for Korean pine has numerous uncertainties in the context of climate change. Although multiple General Circulation Models (GCMs) predicted increases in temperature and precipitation in Northeast China, these predictions varied seasonally [42,43]. A recent study suggested that future climates increased Korean pine aboveground carbon sequestration rates and promoted its dominance [44]. Furthermore, tree harvest interacting with climate change could have synergistic effects on tree species [9,45]. Tree harvest would accelerate tree species colonization by providing colonization opportunities and ameliorate extinction by reducing competition under climate change [46]. Recent studies showed that tree harvest increased the dominance of Korean pine in Northeastern China since it was prohibited from harvesting [47,48]. As reported, tree harvest accelerated Korean pine regeneration and would assist it to shift northward under a warming climate [49]. Given the uncertainties of future climates and the synergistic effects between climate change and tree harvest, it is imperative to assess the range of uncertainties of Korean pine responses to proactive managements under future projections.
Thus, in this study, we employed a forest landscape model, LANDIS PRO, to evaluate the response ranges of Korean pine dominance and regeneration to the resistant and the adaptive treatments under climate change. We expected that the resistant treatment would maintain the dominance and regeneration of Korean pine and that the adaptive treatment would promote its dominance and regeneration. Specifically, we intended to determine: (1) what were the ranges of variation in Korean pine importance value under alternative managements? (2) to what extent could the resistant treatment maintain the dominance and regeneration of Korean pine? and (3) to what extent could the adaptive treatment promote its dominance and regeneration?
2. Materials and Methods
2.1. Study Area
Our study area is located in the Small Khingan Mountains of Northeast China (47°50′–49°10′ N, 127°50′–130°10′ E), covering approximately 1.5 million hectares and ranging in elevation from 139 to 1429 m (Figure 1). Annual mean temperature ranges from 1.0 to −1.0 °C and annual total precipitation decreases from 700 to 550 mm from south to north [22]. Small Khingan Mountains forests lie in the transition between the boreal and the temperate forest biomes and are sensitive to change in climate since Korean pine and some hardwood species are currently at their leading edges. The principal soil of this area is Haplic Luvisols, while Mollic Gleysols, Gleyic Phaeozems, Gleyic Luvisols, and Haplic Phaeozems are minor. Timber harvesting in the past has evidently decreased Korean pine dominance, and white birch and poplar widely distribute at present. Most forests in this region are in the stem exclusion stage with the dense and closed forest canopy. To restore Korean pine, the Natural Forest Conservation Program was implemented in 1998. Currently, the commercial harvest is excluded, and thinning from below is permitted in the General Ecological Welfare Forest and the Commercial Forest (Figure 1). The Special Ecological Welfare Forest precludes any managements.
2.2. Climate Data
We included four climate scenarios based on the Climate Model 3 of the Geophysical Fluid Dynamics Laboratory (GFDL-CM3) of the National Oceanic and Atmospheric Administration, the Hadley Center Global Environment Model version 2 Earth System (HadGEM2-ES) model of the United Kingdom Meteorological Office Hadley Center, the Model for Interdisciplinary Research on Climate Version 5 (MIROC5) of the Japan Agency for Marine Earth Science and Technology, Atmosphere and Ocean Research Institute of the University of Tokyo and the National Institute for Environmental Studies, and the Meteorological Research Institute Climate General Circulation Model 3 (MRI-CGCM3) of the Meteorological Research Institute of Japan general circulation models under radiative forcing scenarios, Representative Concentration Pathway (RCP) 8.5. The RCP 8.5 is the highest emission scenario in the Intergovernmental Panel on Climate Change Fifth Assessment Report and is close to the current emission trajectories. The reason we chose the four GCMs was that they predicted climate conditions with a high spatial resolution (longitude × latitude ranging from 1.1° × 1.1° to 2.5° × 2.0°). Additionally, they reflected different seasonal temperature and precipitation patterns, which ensured that climate ensembles captured the projection uncertainties (Figure 2).
To consider the heterogeneity of environment in this study area, we generated eight land types based on elevation (more or less than 600 m), aspect (south or north), and active accumulated temperature (more or less than 2200 °C*d) [48]. We assumed abiotic factors, such as climate and soil, were homogeneous in one land type but heterogeneous among land types. The principal soil in one land type was used to represent this land type [50]. We needed daily precipitation, maximum and minimum temperature, mean surface wind speed, and incident solar radiation on each land type under current and future climate scenarios. We obtained current daily climate data (1980–2009) from the China National Meteorological Information Center. We obtained future daily climate data (2070–2099) from the Coupled Model Inter-Comparison Project phase 5. We interpolated current climate data and downscaled future climate data to each land type using the R package ‘meteoland’ [50]. Compared with the current climate, the four GCMs projected that the multiple-year seasonal mean precipitation dramatically increased in summer ranging from 60 to 180 mm and less than a 50 mm increase in the other seasons (Figure 2a). Mean temperature increased by 1.5 to 5 °C during 2070–2099 under the RCP 8.5 emission scenario (Figure 2b).
2.3. LINKAGES 3.0 Model and Parameterization
LINKAGES is a biophysical process model that simulates tree growth, competition, seedling establishment, nutrient cycling, evapotranspiration, and soil hydrology at the site scale [51]. The LINKAGES model simulates the physiological effects of abiotic factors including climate and soil on species establishment and growth. LINKAGES 3.0 needs the main parameters, as follows, daily climate data, initial soil data, and species biological traits. We employed the LINKAGES model to estimate seedling establishment probability (SEP) for each species and carrying capacity (maximum growing space occupied, MGSO) on eight land types under each climate scenario. We assumed seedling establishment probability and carrying capacity were uniform within one land type but different among land types. Since daily climate data were used to parameterize LINKAGES, it could capture species’ responses to the seasonal patterns of climate projections.
Daily climate data were input into LINKAGES and were obtained as described in the Climate data section, including maximum and minimum temperature, precipitation, mean incoming solar radiation, and mean wind speed. We derived measures of soil attributes from a soil dataset including layers, thick, organic matter, nitrogen, clay, sand, rock, and root abundance [52]. We further calculated the wilting point and the field moisture capacity by the Century model soil calculator. We derived species biological traits required by LINKAGES (Table 1), such as maximum and minimum growing degree-day requirements (DMAX and DMIN), parameter of Richard function (B2 and B3), growth factor (G) scaling growth rates, drought tolerance (D3), mean January temperature tolerant threshold (FROST), leaf litter type (TL), and foliage retention time (FRT) from the Scientific Database of China Plant Species and a previous study [51]. We obtained the start and the end growing season days from the Forest Phenology Products of Northeast China [53].
We estimated seedling establishment for each species on each land type by simulating seedling establishment and growth from the bare ground over 30 years. Then, we used species biomass and the maximum of the sum of each species biomass under each climate scenario to calculated SEPs on each land type [39,54]. To estimate MGSO on each land type under alternative climate scenarios, we simulated all species establishment and growth from the bare ground over 300 years and used the maximum biomass among climate scenarios [39]. We linearly interpolated values of SEP and MGSO before the year 2100, and we held these values constant after the year 2100. SEP and MGSO were key parameters in LANDIS PRO that simulated species demography and responses to climate change. Although LINKAGES is a daily model whereas LANDIS is a yearly to decadal model, the different temporal resolution of LINKAGES and LANDIS would not affect the modeling coupling that has been used in this and previous studies [9,54].
2.4. LANDIS PRO Model and Parameterization
LANDIS is a raster-based forest landscape model that records tree species distribution (absence or presence) and abundance (tree number and basal area) by age cohort in each raster cell [55]. LANDIS simulates species demography, competition, and forest management over large temporal and spatial extents with flexible resolutions. Inputs for LANDIS PRO contain spatial data, mainly including raster maps of species composition, land type, stand and management area, and non-spatial data mainly including species biological traits, growth curve, SEP, and MGSO.
Species demography including growth, fecundity, dispersal, colonization, and mortality, is mainly driven by species ecological traits such as growth rate, longevity, maturation age, shade tolerance, maximum dispersal distance, maximum diameter at breast height, maximum stand density index, and number of potential germination seeds per mature tree (Table 2). We obtained these species attributes from previous studies in the temperate forests [44,48,49]. Growth is regulated by the growth rate quantified by the relationship between age and diameter at breast height. Seed abundance on one cell is determined by fertile tree abundance, species reproductive rate, habitat connectivity, and dispersal capacity from the surrounding cells. Seed dispersal generates spatial interaction among pixels and makes it possible to simulate changes in species distribution and abundance. When species’ SEP is greater than a randomly generated 0.0–1.0 number, and meanwhile there exists enough growing space, individuals will colonize on that cell. Competition is initiated once MGSO is reached, and the mortality caused by competition is characterized as decreasing tree number with increasing diameter, which is determined by Yoda’s thinning theory [55,56]. Additionally, LANDIS simulates the mortality induced by longevity and disturbances. We simulated forest management using a management area map and a stand map in the LANDIS harvest module [57]. The management area map provides boundaries to constrain specific harvest events and there are different objectives in each management area. Management activities are implemented in each stand that is the minimum treatment unit.
We derived an initial species composition map, including number of trees by age cohort for tree species in each cell, from inventory data providing age information and a stand composition map detailing species distribution and abundance [48]. We verified initial forest biomass and ensured that there was no significant difference between inventory data and parameterized LANDIS data [47,48]. We generated the management area map and the stand map based on actual forest management data. The land type map was generated as described in the climate data section. SEP and MGSO on each land type were calculated as described in the LINKAGES 3.0 model and parameterization section. The resolution of all the input raster was 100 × 100 m. We simulated thinning from below in the General Ecological Welfare Forest and the Commercial Forest (Figure 1) and prohibited harvesting Korean pine, manchurian ash, manchurian walnut, amur corktree, and amur linden according to the current management requirement [47].
2.5. Experimental Design
Reducing stand density or increasing forest compositional and structural diversity are usually considered to cope with future climate change [10,12,14]. Generally, reducing stand density could maintain current species composition, and high diversity is hypothesized to promote forest with a greater range of potential responses to future uncertainty [17]. We designed two proactive treatments (Table 3), that is the resistant and the adaptive treatments, corresponding to the above two categories of management to cope with uncertainties of climate change. We also considered a baseline treatment excluding any management to check the effects of the two proactive treatments on Korean pine. Thus, we formulated the above three treatment scenarios. All commercial harvests (i.e., thinning from above) have been excluded and thinning from below is currently the only permitted management in this region. The resistant treatment scenario introduced the current management regime, and the minimum and the residual stand stocking were referred to the previous study [47]. The current management thinned 10% of forest area per decade and had 100 years of rotation, which were derived from the China Regulation for Tending of Forest and the consultation with local experts [47]. For both the resistant and the adaptive treatment scenarios, only the thinning rotation was modified, while all other simulation parameters, such as the minimum and the residual stand stocking, were kept constant. The resistant treatment scenario extended the current management rotation by 50 years to reduce disturbance on the landscape, and it harvested 7% of forest area per decade to ensure the managed area was equal to forest area within 150 years. Whereas, the adaptive treatment scenario shortened the current management rotation by 50 years to increase disturbance, and it harvested 20% of forest area per decade to ensure the managed area was equal to forest area within 50 years.
We simulated three treatment scenarios under four climate scenarios, for a total of 12 simulation scenarios. We started LANDIS PRO with the same initial forest composition for the 12 simulation scenarios to project the changes in Korean pine for 150 years (from 2010 to 2160 with a 10-year step). We assessed the effects of three treatment scenarios on Korean pine importance value (IV = (individual species density/total density + individual species basal area/total basal area)/2), relative dominance, and regeneration represented by the abundance of juveniles less than 10 years old. To better evaluate results among different scenarios, we took the baseline treatment as the reference and calculated the relative change of importance value, biomass, and abundance under the three treatment scenarios. We calculated the relative change for treatment scenario i as the ratio between RVi (response variable i) and RVbaseline. We calculated the mean and the standard deviation of relative change of each response variable under the four climate scenarios for each treatment scenario to evaluate the range of uncertainties of Korean pine response to future climate. We also analyzed relative changes of these response variables for the short term (2010–2040), the medium term (2050–2100), and the long term (2110–2160) under the three treatment scenarios. The separate results for each land type were similar to the entire study area, thus we did not present individual results for each land type.
3. Results
3.1. Variations in Korean Pine Importance Value (IV)
Korean pine IV were 14.2% and 42.9% higher on average under the adaptive and the resistant treatment scenarios than the baseline treatment scenario throughout the simulation (Figure 3a). The difference in IV between the baseline scenario and either the resistant or the adaptive treatment scenarios gradually increased. The maximum difference between the baseline and the adaptive treatment scenarios was 64.2% at the end of the simulation year, and the maximum difference between the baseline and the resistant treatment scenarios was 21.6%. Predictions of IV had the greatest uncertainties under the baseline treatment scenario, followed by the resistant and the adaptive treatment scenarios. The predicted uncertainties gradually increased over time and ranged from 0% to 5.8%, 0% to 4.9%, and 0% to 4.6% under the baseline, the resistant and the adaptive treatment scenarios, respectively.
Spatial patterns of Korean pine IV were similar under alternative treatment scenarios, but IV under the adaptive treatment scenario evidently was higher than the resistant and the baseline treatment scenarios across the entire region (Figure 4a–c). Korean pine was more dominant in the center and the northeast of its distribution area and was less dominant in the south irrespective of treatment scenario. The uncertainties of IV across the entire region were largest under the adaptive treatment scenario, followed by the resistant and the baseline treatment scenarios (Figure 5a–c).
3.2. Variations in Korean Pine Dominance
Korean pine biomass were 25.5% and 9.2% higher on average under the adaptive and the resistant treatment scenarios than the baseline treatment scenario throughout the simulation (Figure 3b). The difference in biomass between the baseline treatment scenario and either the resistant or the adaptive treatment scenarios gradually increased. The maximum difference between the adaptive and the baseline treatment scenarios was 46.7% at the end of the simulation year, and the maximum difference between the resistant and the baseline treatment scenarios reached 16.9%. Predictions of biomass had the largest uncertainties under the baseline treatment scenario, followed by the resistant and the adaptive treatment scenarios. Predicted uncertainties increased over time and ranged from 0% to 4.3%, 0% to 4.2%, and 0% to 3.9% under the baseline, the resistant, and the adaptive treatment scenarios, respectively.
Korean pine biomass showed similar spatial patterns under alternative treatment scenarios, but biomass was larger under the adaptive treatment scenario than those under the resistant and the baseline treatment scenarios across the whole area (Figure 4d–f). The high biomass area was concentrated in the center and the northeast of its distribution area regardless of treatment scenario, whereas biomass in the south was minor. The uncertainties of biomass were largest under the adaptive treatment scenario, followed by the resistant and the baseline treatment scenarios across the region (Figure 5d–f).
3.3. Variations in Korean Pine Regeneration
The abundance of Korean pine juveniles were 841.2% and 286.6% higher on average under the adaptive and the resistant treatment scenarios than the baseline treatment scenario throughout the simulation, especially for the short term (Figure 3c). Predictions of abundance had the greatest uncertainties under the adaptive treatment scenario, followed by the resistant and the baseline treatment scenarios. The predicted uncertainties increased in fluctuation ranging from 0% to 22.8%, 0% to 45.8%, and 0% to 85.4% under the baseline, the resistant, and the adaptive treatment scenarios, respectively. Compared with the baseline treatment scenario, both the resistant and the adaptive treatment scenarios increased abundance of juveniles across the distribution area, especially for the adaptive treatment scenario (Figure 4g–i). The uncertainties of abundance across the whole region were largest under the adaptive treatment scenario, followed by the resistant and the baseline treatment scenarios (Figure 5g–i).
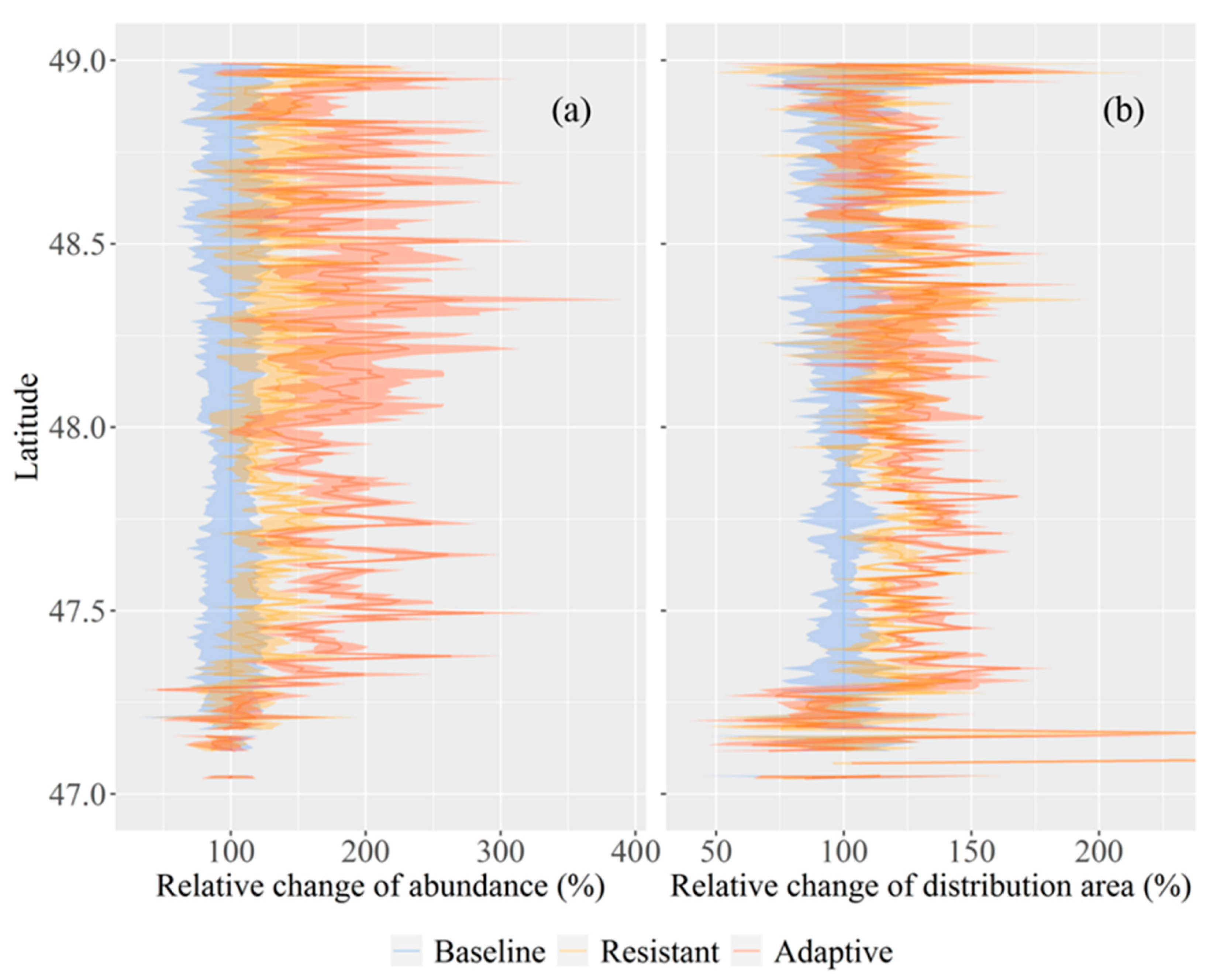
The average abundance of Korean pine juveniles was similar among alternative treatment scenarios in low latitude regions (latitude less than 47.3°) (Figure 6a). However, the abundance of juveniles in high latitude regions (latitude greater than 47.3°) were 80.1% and 29.4% higher on average under the adaptive and the resistant treatment scenarios than the baseline treatment scenario. The uncertainties of abundance were largest under the adaptive treatment scenario with a mean of 36.0%, followed by the resistant and the baseline treatment scenario with 25.8% and 21.8%. The distribution area of Korean pine juveniles was analogous in low latitude regions (latitude less than 47.3°) for all treatment scenarios (Figure 6b). Yet, the distribution area in high latitude regions (latitude greater than 47.3°) were 25.4% and 16.7% higher on average under the adaptive and the resistant treatment scenarios than the baseline treatment scenario. The differences of uncertainties of distribution area were minor among the three treatments with mean of 12.8%.
4. Discussion
As expected, proactive managements promoted Korean pine IV, dominance, and regeneration under a warming climate (Figure 3 and Figure 4). Results indicated that proactive managements could enhance the adaptability of Korean pine to climate warming. Recent studies in the Mediterranean region reported similar findings that thinning enhanced the radial increment of Pinus pinaster and Scots pine stands under climate change [15,58]. Although Korean pine dominance could be maintained with prohibiting from harvesting under the current management regime [24], thinning could further release growing space for Korean pine and mitigate competition from other species, which could promote Korean pine growth and regeneration [41]. A recent study showed that thinning increased compositional and structural diversity of red pine and hardwood forests in eastern US [16], and forest with high diversity was generally assumed to have a large potential to cope with climate change [16,17]. Moreover, thinning decreased the mortality caused by competition and increased the proportion of species adaptive to climate change, which further increased forest adaptability [15,58,59]. Although thinning was able to enhance forest adaptability to climate change, it might decrease forest aboveground carbon storage. Thus, there is a trade-off between carbon storage and adaptability to climate change, which should be considered in forest management [58,60].
Our results showed that the uncertainties of Korean pine response to climate change increased over time (Figure 3 and Figure 5). This was consistent with the uncertainty of forest landscape prediction increase over time [61]. Korean pine had minor uncertainties in response to climate change before regeneration from the initial stand or existing seedlings reached a mature stage (age less than 40 years old). These seedlings gradually matured and started to produce seeds as stands developed, which contributed to greater uncertainties of Korean pine. This was because uncertainties associated with the regeneration of Korean pine were greater under climate change with a lagging phenomenon. More Korean pine regenerated in the regions with higher abundance due to adequate seeds, and thus, higher uncertainties of Korean pine response to climate change concentrated in those regions. Compared with mature individuals, Korean pine juveniles could rapidly respond to climate change [62,63]; thus, Korean pine regeneration was more sensitive than dominance.
Our results showed that proactive managements facilitated the leading edge of Korean pine to shift poleward, but had less impact on its trailing edge. This result was in agreement with a recent finding that tree species at the leading edge migrated poleward, while the trailing edge had no evident change [63]. Generally, moisture availability and temperature were critical factors in limiting species distribution [64]. Temperature limited the leading edge of Korean pine in the mixed coniferous and broadleaf forests located in the humid temperate forests [22]. A recent study showed that tree harvest and climate change interacted to affect species distribution, and the interactive effect between tree harvest and climate change acted in the same direction on species distribution [9]. The previous study showed that tree harvest accelerated Korean pine shifting northward under a warming climate [49]. Hence, investigations on species distribution needed to incorporate tree harvest besides climatic factors such as temperature and precipitation.
Our result showed that the positive effects of proactive managements on Korean pine were site-specific, which was in line with a previous study [59]. Compared with the high-latitude region (latitude more than 47.3°), proactive managements did not evidently promote the dominance and abundance of Korean pine in the low latitude region (latitude less than 47.3°) (Figure 4d–i and Figure 6). There were several rare and protected broadleaf tree species, such as manchurian ash, manchurian walnut, amur corktree, and amur linden, which were also prohibited from harvesting. Proactive managements facilitated both Korean pine and these species, which increased competition for Korean pine and thereby limited its growth and regeneration. A recent study showed that the effect of thinning varied at different elevations. Compared with low elevation, the increasing drought at high elevation decreased the positive effect of thinning [59]. Thus, the effect of thinning on forest adaptability to climate change was influenced by species composition and drought tolerance.
Our results showed that the adaptive treatment (increasing diversity) with the high frequency of thinning obviously promoted dominance and regeneration of Korean pine greater than the resistant treatment (reducing density), which suggested that increasing thinning rotation could make Korean pine better cope with climate change. Recent studies showed that light and moderate thinning maintained current composition while heavy thinning promoted forest to adapt to climate change, which indicated that changing thinning intensity also could affect species adaptability [19,59]. Moreover, the previous study found that harvesting species maladaptive to climate change increased the abundance of adaptive species and promoted forest adaptability [65]. Except for reducing stand density or increasing stand compositional and structural diversity, facilitating species composition transition was also used to cope with climate change. The transition treatment focused on facilitating species composition or stand structure to a more adaptive state [10,12]. Generally, the transition treatment included planting species adaptive to future climate outside of their native ranges or drastically altering the disturbance regime and structural composition of a forest to promote a shift to new functional states [12,66,67]. A recent study showed that planting future-adapted species would help systems maintain productivity and functional diversity [67]. Thus, future forest management should consider the combinations of intensity, rotation, harvesting species, and climate-suitable planting to enhance forest adaptability to climate change.
Forest management interacted with multiple scale processes to affect forest adaptability. Climatic factors, such as temperature and precipitation, determined species distribution at the regional scale. Temperature generally limited latitudinal shift whereas precipitation limited longitudinal shift [64]. Dispersal at the landscape scale enabled species to change their distribution to adapt to changing environmental conditions [8]. Additionally, competition for resources at the stand scale also impacted individual growth, mortality, and regeneration [46]. Traditional experimental studies have limited capacity to include environmental heterogeneity over the large spatial extent while being limited in monitoring for a sufficient length of time to observe the effect of management on forest adaptability to climate change [58]. However, forest landscape models incorporating the effects of spatial heterogeneity of abiotic factors, landscape-processes (e.g., dispersal and forest management), and stand dynamics such as growth, mortality, and regeneration on forest adaptability [68], presented a viable approach in studying forest response to climate change.
Several factors excluded in this study might contribute to uncertainties of our simulation. We did not consider the effect of fire on Korean pine. A recent study showed that fires were forecast to globally increase in the context of climate change [69]. Generally, young trees were more susceptible to fire than mature trees [70], thus omitting fire would increase the uncertainties of Korean pine regeneration. Moreover, tree harvest and fire interacted to affect forest dynamics [71], the interactive effect also could increase uncertainties of prediction. Compared with niche models and process-based ecosystem models, the LANDIS PRO model did not directly input climate data while coupled with an ecosystem model to incorporate the effects of climate change. We considered the effects of climate change on species demography including temperature, precipitation, radiation, wind, growing season length, and drought, but ignoring nitrogen deposition and CO2 fertilization, which had the significant impact on tree species distribution and primary production [72,73]. We assumed that forest management policy and the boundaries of management area and stand kept constant, however these assumptions varied in different periods, which would contribute to uncertainties of simulation. Although these factors and assumptions increased uncertainties of prediction, our results still clarified the response of Korean pine forest to proactive managements under climate change and provided a valuable reference for restoration and protection of Korean pine mixed forest.
5. Conclusions
In this study, we evaluated the range of Korean pine to proactive treatments under climate change by coupling an ecosystem process-based model, LINKAGES, and a forest landscape model, LANDIS PRO. The adaptive treatment increases the adaptability of Korean pine to climate change more than the resistant treatment, and we recommend applying the adaptive treatment to promote Korean pine under future climatic conditions. The uncertainties of Korean pine response to management treatments increase over time under climate change, and the uncertainties are associated with the regeneration of Korean pine. The effects of proactive treatments on Korean pine are site-specific, and the competition between Korean pine and the rare and protected broadleaf tree species might offset the positive effects. Proactive treatments promote the leading edge of Korean pine to shift poleward, while they had less impact on its trailing edge. The adaptive treatment is more likely to prompt Korean pine to migrate into its suitable habitats under climate change. Future forest management alternatives, except for thinning from below, also should include thinning from below to promote Korean pine under climate change.
Author Contributions
Conceptualization, K.L. and H.H.; formal analysis, K.L., W.X., H.D., S.Z., C.H., and M.W.; funding acquisition, H.H.; resources, K.L., W.X., and C.H.; supervision, H.H.; writing—original draft, K.L.; writing—review and editing, H.H., W.X., H.D., S.Z., M.W., X.T., and Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China, grant number 2017YFA0604403-3 and 2016YFA0602301, the Joint Fund of National Natural Science Foundation of China, grant number U19A2023, and the National Natural Science Foundation of China, grant number 31800408.
Acknowledgments
We thank Jiangtao Xiao and Guohua Song for their suggestions on the parameterization of LANDIS and harvest specifications. We are grateful to Wen J. Wang for her suggestions on the parameterization of LINKAGES.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Elsen, P.R.; Tingley, M.W. Global mountain topography and the fate of montane species under climate change. Nat. Clim. Chang. 2015, 5, 772. [Google Scholar] [CrossRef]
- Alexander, J.M.; Chalmandrier, L.; Lenoir, J.; Burgess, T.I.; Essl, F.; Haider, S.; Kueffer, C.; McDougall, K.; Milbau, A.; Nuñez, M.A. Lags in the response of mountain plant communities to climate change. Glob. Chang. Biol. 2018, 24, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Liu, J.; Zhou, L.; Zhou, W.; Fang, X.; Wei, Y.; Jiang, S.; Dai, L. Spatial variation and temporal instability in the climate–growth relationship of Korean pine in the Changbai Mountain region of Northeast China. For. Ecol. Manag. 2013, 300, 96–105. [Google Scholar] [CrossRef]
- Taylor, A.R.; Boulanger, Y.; Price, D.T.; Cyr, D.; McGarrigle, E.; Rammer, W.; Kershaw, J.A., Jr. Rapid 21st century climate change projected to shift composition and growth of Canada’s Acadian Forest Region. For. Ecol. Manag. 2017, 405, 284–294. [Google Scholar] [CrossRef]
- Sittaro, F.; Paquette, A.; Messier, C.; Nock, C.A. Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits. Glob. Chang. Biol. 2017, 23, 3292–3301. [Google Scholar] [CrossRef]
- Liang, Y.; Duveneck, M.J.; Gustafson, E.J.; Serra-Diaz, J.M.; Thompson, J.R. How disturbance, competition, and dispersal interact to prevent tree range boundaries from keeping pace with climate change. Glob. Chang. Biol. 2018, 24, e335–e351. [Google Scholar] [CrossRef]
- Wang, W.J.; Thompson, F.R., III; He, H.S.; Fraser, J.S.; Dijak, W.D.; Jones-Farrand, T. Climate change and tree harvest interact to affect future tree species distribution changes. J. Ecol. 2019, 107, 1901–1917. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- Locatelli, B.; Evans, V.; Wardell, A.; Andrade, A.; Vignola, R. Forests and climate change in Latin America: Linking adaptation and mitigation. Forests 2011, 2, 431–450. [Google Scholar] [CrossRef]
- Nagel, L.M.; Palik, B.J.; Battaglia, M.A.; D′Amato, A.W.; Guldin, J.M.; Swanston, C.W.; Janowiak, M.K.; Powers, M.P.; Joyce, L.A.; Millar, C.I. Adaptive silviculture for climate change: A national experiment in manager-scientist partnerships to apply an adaptation framework. J. For. 2017, 115, 167–178. [Google Scholar] [CrossRef]
- Fischer, A.P. Adapting and coping with climate change in temperate forests. Glob. Environ. Chang. 2019, 54, 160–171. [Google Scholar] [CrossRef]
- Fischer, A.P. Characterizing behavioral adaptation to climate change in temperate forests. Landsc. Urban Plan. 2019, 188, 72–79. [Google Scholar] [CrossRef]
- Aldea, J.; Bravo, F.; Bravo-Oviedo, A.; Ruiz-Peinado, R.; Rodríguez, F.; Del Río, M. Thinning enhances the species-specific radial increment response to drought in Mediterranean pine-oak stands. Agric. For. Meteorol. 2017, 237, 371–383. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Bradford, J.B.; Fraver, S.; Palik, B.J. Forest management for mitigation and adaptation to climate change: Insights from long-term silviculture experiments. For. Ecol. Manag. 2011, 262, 803–816. [Google Scholar] [CrossRef]
- Olson, M.G.; Knapp, B.O.; Kabrick, J.M. Dynamics of a temperate deciduous forest under landscape-scale management: Implications for adaptability to climate change. For. Ecol. Manag. 2017, 387, 73–85. [Google Scholar] [CrossRef]
- Thom, D.; Rammer, W.; Seidl, R. Disturbances catalyze the adaptation of forest ecosystems to changing climate conditions. Glob. Chang. Biol. 2017, 23, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Sohn, J.A.; Hartig, F.; Kohler, M.; Huss, J.; Bauhus, J. Heavy and frequent thinning promotes drought adaptation in Pinus sylvestris forests. Ecol. Appl. 2016, 26, 2190–2205. [Google Scholar] [CrossRef]
- Bréda, N.; Brunette, M. Are 40 years better than 55? An analysis of the reduction of forest rotation to cope with drought events in a Douglas fir stand. Ann. For. Sci. 2019, 76, 29. [Google Scholar] [CrossRef] [Green Version]
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest adaptation to climate change—Is non-management an option? Ann. For. Sci. 2019, 76, 48. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y. Vegetation in the Small Khingan Mountains of China; Science Press: Beijing, China, 1994. [Google Scholar]
- Li, J.; Li, J. Regeneration and restoration of broad-leaved Korean pine forests in Lesser Xing′ an Mountains of Northeast China. Acta Ecol. Sin. 2003, 23, 1268–1277. [Google Scholar]
- Yu, D.; Zhou, L.; Zhou, W.; Ding, H.; Wang, Q.; Wang, Y.; Wu, X.; Dai, L. Forest management in Northeast China: History, problems, and challenges. Environ. Manag. 2011, 48, 1122–1135. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhou, W.; Zhou, L.; Dai, L. Exploring the history of the management theory and technology of broad-leaved Korean pine (Pinus koraiensis Sieb. et Zucc.) forest in Changbai Mountain Region, Northeast China. Chin. J. Appl. Ecol. 2019, 30, 1426–1434. [Google Scholar]
- Zhu, J.; Wang, K.; Sun, Y.; Yan, Q. Response of Pinus koraiensis seedling growth to different light conditions based on the assessment of photosynthesis in current and one-year-old needles. J. For. Res. 2014, 25, 53–62. [Google Scholar] [CrossRef]
- Sun, Y.; Zhu, J.; Yu, L.; Yan, Q.; Wang, K. Photosynthetic characteristics of Pinus koraiensis seedlings under different light regimes. Chin. J. Ecol. 2009, 28, 850–857. [Google Scholar]
- Qi, L.; Yang, J.; Yu, D.; Dai, L.; Contrereas, M. Responses of regeneration and species coexistence to single-tree selective logging for a temperate mixed forest in eastern Eurasia. Ann. For. Sci. 2016, 73, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhu, J.; Sun, O.J.; Yan, Q. Photosynthetic and growth responses of Pinus koraiensis seedlings to canopy openness: Implications for the restoration of mixed-broadleaved Korean pine forests. Environ. Exp. Bot. 2016, 129, 118–126. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Ji, Y.; Li, Z.; Li, M.; Zhang, Y. Temperature signals in tree-ring width and divergent growth of Korean pine response to recent climate warming in northeast Asia. Trees 2017, 31, 415–427. [Google Scholar] [CrossRef]
- Zhao, W.; Du, H.; Wang, L.; He, H.S.; Wu, Z.; Liu, K.; Guo, X.; Yang, Y. A comparison of recent trends in precipitation and temperature over Western and Eastern Eurasia. Q. J. R. Meteorol. Soc. 2018, 144, 604–613. [Google Scholar] [CrossRef]
- Piao, S.; Ciais, P.; Huang, Y.; Shen, Z.; Peng, S.; Li, J.; Zhou, L.; Liu, H.; Ma, Y.; Ding, Y. The impacts of climate change on water resources and agriculture in China. Nature 2010, 467, 43. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Lu, X. Weak cooling of cold extremes versus continued warming of hot extremes in China during the recent global surface warming hiatus. J. Geophys. Res. Atmos. 2018, 123, 4073–4087. [Google Scholar] [CrossRef]
- Wang, X.; Pederson, N.; Chen, Z.; Lawton, K.; Zhu, C.; Han, S. Recent rising temperatures drive younger and southern Korean pine growth decline. Sci. Total Environ. 2019, 649, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.; Wang, X.; Zhang, Y.; Li, Z. Different responses of Korean pine (Pinus koraiensis) and Mongolia oak (Quercus mongolica) growth to recent climate warming in northeast China. Dendrochronologia 2017, 45, 113–122. [Google Scholar] [CrossRef]
- Li, G.; Bai, F.; Sang, W. Different responses of radial growth to climate warming in Pinus koraiensis and Picea jezoensis var. komarovii at their upper elevational limits in Changbai Mountain, China. Chin. J. Plant Ecol. 2011, 35, 500–511. [Google Scholar] [CrossRef]
- Gao, L.; Wang, X.; Zhao, X. Response of Pinus koraiensis and Picea jezoensis var. komarovii to climate in the transition zone of Changbai Mountain, China. Chin. J. Plant Ecol. 2011, 35, 27–34. [Google Scholar] [CrossRef]
- Li, X.; He, H.S.; Wu, Z.; Liang, Y.; Schneiderman, J.E. Comparing effects of climate warming, fire, and timber harvesting on a boreal forest landscape in northeastern China. PLoS ONE 2013, 8, e59747. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; He, H.S.; Thompson, F.R., III; Fraser, J.S.; Hanberry, B.B.; Dijak, W.D. Importance of succession, harvest, and climate change in determining future composition in US Central Hardwood Forests. Ecosphere 2015, 6, 1–18. [Google Scholar] [CrossRef]
- Kellner, K.F.; Swihart, R.K. Timber harvest and drought interact to impact oak seedling growth and survival in the Central Hardwood Forest. Ecosphere 2016, 7, e01473. [Google Scholar] [CrossRef]
- Zhu, L.; Yang, J.; Zhu, C.; Wang, X. Influences of gap disturbance and warming on radial growth of Pinus koraiensis and Abies nephrolepis in Xiaoxing’an Mountain, Northeast China. Chin. J. Ecol. 2015, 34, 2085–2095. [Google Scholar]
- Liu, Z.; Mehran, A.; Phillips, T.J.; AghaKouchak, A. Seasonal and regional biases in CMIP5 precipitation simulations. Clim. Res. 2014, 60, 35–50. [Google Scholar] [CrossRef]
- Chen, L.; Frauenfeld, O.W. Surface air temperature changes over the twentieth and twenty-first centuries in China simulated by 20 CMIP5 models. J. Clim. 2014, 27, 3920–3937. [Google Scholar] [CrossRef]
- Ma, J.; Hu, Y.; Bu, R.; Chang, Y.; Deng, H.; Qin, Q. Predicting impacts of climate change on the aboveground carbon sequestration rate of a temperate forest in northeastern China. PLoS ONE 2014, 9, e96157. [Google Scholar] [CrossRef] [PubMed]
- García-Valdés, R.; Svenning, J.C.; Zavala, M.A.; Purves, D.W.; Araújo, M.B. Evaluating the combined effects of climate and land-use change on tree species distributions. J. Appl. Ecol. 2015, 52, 902–912. [Google Scholar] [CrossRef]
- Wang, W.J.; Thompson, F.R., III; He, H.S.; Fraser, J.S.; Dijak, W.D.; Spetich, M.A. Population dynamics has greater effects than climate change on tree species distribution in a temperate forest region. J. Biogeogr. 2018, 45, 2766–2778. [Google Scholar] [CrossRef]
- Liu, K.; Liang, Y.; He, H.; Wang, W.; Huang, C.; Zong, S.; Wang, L.; Xiao, J.; Du, H. Long-Term Impacts of China’s New Commercial Harvest Exclusion Policy on Ecosystem Services and Biodiversity in the Temperate Forests of Northeast China. Sustainability 2018, 10, 1071. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Liang, Y.; He, H.S.; Thompson, J.R.; Wang, W.J.; Fraser, J.S.; Wu, Z. The formulations of site-scale processes affect landscape-scale forest change predictions: A comparison between LANDIS PRO and LANDIS-II forest landscape models. Landsc. Ecol. 2017, 32, 1347–1363. [Google Scholar] [CrossRef]
- Bu, R.; He, H.S.; Hu, Y.; Chang, Y.; Larsen, D.R. Using the LANDIS model to evaluate forest harvesting and planting strategies under possible warming climates in Northeastern China. For. Ecol. Manag. 2008, 254, 407–419. [Google Scholar] [CrossRef]
- De Caceres, M.; Martin-StPaul, N.; Turco, M.; Cabon, A.; Granda, V. Estimating daily meteorological data and downscaling climate models over landscapes. Environ. Model. Softw. 2018, 108, 186–196. [Google Scholar] [CrossRef]
- Dijak, W.D.; Hanberry, B.B.; Fraser, J.S.; He, H.S.; Wang, W.J.; Thompson, F.R. Revision and application of the LINKAGES model to simulate forest growth in central hardwood landscapes in response to climate change. Landsc. Ecol. 2017, 32, 1365–1384. [Google Scholar] [CrossRef]
- Shangguan, W.; Dai, Y.; Liu, B.; Zhu, A.; Duan, Q.; Wu, L.; Ji, D.; Ye, A.; Yuan, H.; Zhang, Q. A China data set of soil properties for land surface modeling. J. Adv. Model. Earth Syst. 2013, 5, 212–224. [Google Scholar] [CrossRef]
- Yu, X.; Zhuang, D.; Jiang, D.; Qiankun, W. Forest phenology products of Northeast China. Glob. Chang. Res. Data Publ. Repos. 2014. [Google Scholar] [CrossRef]
- He, H.S.; Mladenoff, D.J.; Crow, T.R. Linking an ecosystem model and a landscape model to study forest species response to climate warming. Ecol. Model. 1999, 114, 213–233. [Google Scholar] [CrossRef]
- Wang, W.J.; He, H.S.; Spetich, M.A.; Shifley, S.R.; Thompson, F.R., III; Dijak, W.D.; Wang, Q. A framework for evaluating forest landscape model predictions using empirical data and knowledge. Environ. Model. Softw. 2014, 62, 230–239. [Google Scholar] [CrossRef]
- Yoda, K. Self-thinning in overcrowded pure stands under cultivated and natural conditions (Intraspecific competition among higher plants. XI). J. Biol. Osaka City Univ. 1963, 14, 107–129. [Google Scholar]
- Fraser, J.S.; He, H.S.; Shifley, S.R.; Wang, W.J.; Thompson, F.R., III. Simulating stand-level harvest prescriptions across landscapes: LANDIS PRO harvest module design. Can. J. For. Res. 2013, 43, 972–978. [Google Scholar] [CrossRef]
- Ameztegui, A.; Cabon, A.; De Cáceres, M.; Coll, L. Managing stand density to enhance the adaptability of Scots pine stands to climate change: A modelling approach. Ecol. Model. 2017, 356, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Elkin, C.; Giuggiola, A.; Rigling, A.; Bugmann, H. Short-and long-term efficacy of forest thinning to mitigate drought impacts in mountain forests in the European Alps. Ecol. Appl. 2015, 25, 1083–1098. [Google Scholar] [CrossRef]
- Wagner, S.; Nocentini, S.; Huth, F.; Hoogstra-Klein, M. Forest management approaches for coping with the uncertainty of climate change: Trade-offs in service provisioning and adaptability. Ecol. Soc. 2014, 19, 32. [Google Scholar] [CrossRef]
- Xu, C.; Gertner, G.Z.; Scheller, R.M. Uncertainties in the response of a forest landscape to global climatic change. Glob. Chang. Biol. 2009, 15, 116–131. [Google Scholar] [CrossRef]
- Zhu, K.; Woodall, C.W.; Clark, J.S. Failure to migrate: Lack of tree range expansion in response to climate change. Glob. Chang. Biol. 2012, 18, 1042–1052. [Google Scholar] [CrossRef]
- Woodall, C.W.; Oswalt, C.M.; Westfall, J.A.; Perry, C.H.; Nelson, M.D.; Finley, A.O. An indicator of tree migration in forests of the eastern United States. For. Ecol. Manag. 2009, 257, 1434–1444. [Google Scholar] [CrossRef]
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Sci. Adv. 2017, 3, e1603055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenberg, J.W.; Duinker, P.N.; Bush, P.G. Exploring adaptation to climate change in the forests of central Nova Scotia, Canada. For. Ecol. Manag. 2011, 262, 2316–2327. [Google Scholar] [CrossRef]
- Stein, B.A.; Staudt, A.; Cross, M.S.; Dubois, N.S.; Enquist, C.; Griffis, R.; Hansen, L.J.; Hellmann, J.J.; Lawler, J.J.; Nelson, E.J. Preparing for and managing change: Climate adaptation for biodiversity and ecosystems. Front. Ecol. Environ. 2013, 11, 502–510. [Google Scholar] [CrossRef]
- Duveneck, M.J.; Scheller, R.M. Climate-suitable planting as a strategy for maintaining forest productivity and functional diversity. Ecol. Appl. 2015, 25, 1653–1668. [Google Scholar] [CrossRef]
- Wang, W.J.; He, H.S.; Thompson, F.R., III; Spetich, M.A.; Fraser, J.S. Effects of species biological traits and environmental heterogeneity on simulated tree species distribution shifts under climate change. Sci. Total Environ. 2018, 634, 1214–1221. [Google Scholar] [CrossRef]
- Jolly, W.M.; Cochrane, M.A.; Freeborn, P.H.; Holden, Z.A.; Brown, T.J.; Williamson, G.J.; Bowman, D.M. Climate-induced variations in global wildfire danger from 1979 to 2013. Nat. Commun. 2015, 6, 7537. [Google Scholar] [CrossRef]
- He, H.S.; Mladenoff, D.J.; Gustafson, E.J. Study of landscape change under forest harvesting and climate warming-induced fire disturbance. For. Ecol. Manag. 2002, 155, 257–270. [Google Scholar] [CrossRef]
- Huang, C.; He, H.S.; Liang, Y.; Wu, Z.; Hawbaker, T.J.; Gong, P.; Zhu, Z. Long-term effects of fire and harvest on carbon stocks of boreal forests in northeastern China. Ann. For. Sci. 2018, 75, 42. [Google Scholar] [CrossRef] [Green Version]
- Griepentrog, M.; Eglinton, T.I.; Hagedorn, F.; Schmidt, M.W.; Wiesenberg, G.L. Interactive effects of elevated CO 2 and nitrogen deposition on fatty acid molecular and isotope composition of above-and belowground tree biomass and forest soil fractions. Glob. Chang. Biol. 2015, 21, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Ma, S.; He, H.S.; Liu, Z.; Thompson, F.R., III; Jin, W.; Wu, Z.F.; Spetich, M.A.; Wang, L.; Xue, S. Effects of rising atmospheric CO2, climate change, and nitrogen deposition on aboveground net primary production in a temperate forest. Environ. Res. Lett. 2019, 14, 104005. [Google Scholar] [CrossRef]
Figure 1.
Geographical location of the study area. SEWF, Special Ecological Welfare Forest; GEWF, General Ecological Welfare Forest; CF, Commercial Forest.
Figure 1.
Geographical location of the study area. SEWF, Special Ecological Welfare Forest; GEWF, General Ecological Welfare Forest; CF, Commercial Forest.

Figure 2.
Changes in average seasonal (a) precipitation and (b) temperature of future climate (2070–2099) from the four GCMs (GFDL-CM3, HadGEM2-ES, MIROC5, MRI-CGCM3) under the RCP 8.5 scenario compared with current climate (1980–2009).
Figure 2.
Changes in average seasonal (a) precipitation and (b) temperature of future climate (2070–2099) from the four GCMs (GFDL-CM3, HadGEM2-ES, MIROC5, MRI-CGCM3) under the RCP 8.5 scenario compared with current climate (1980–2009).
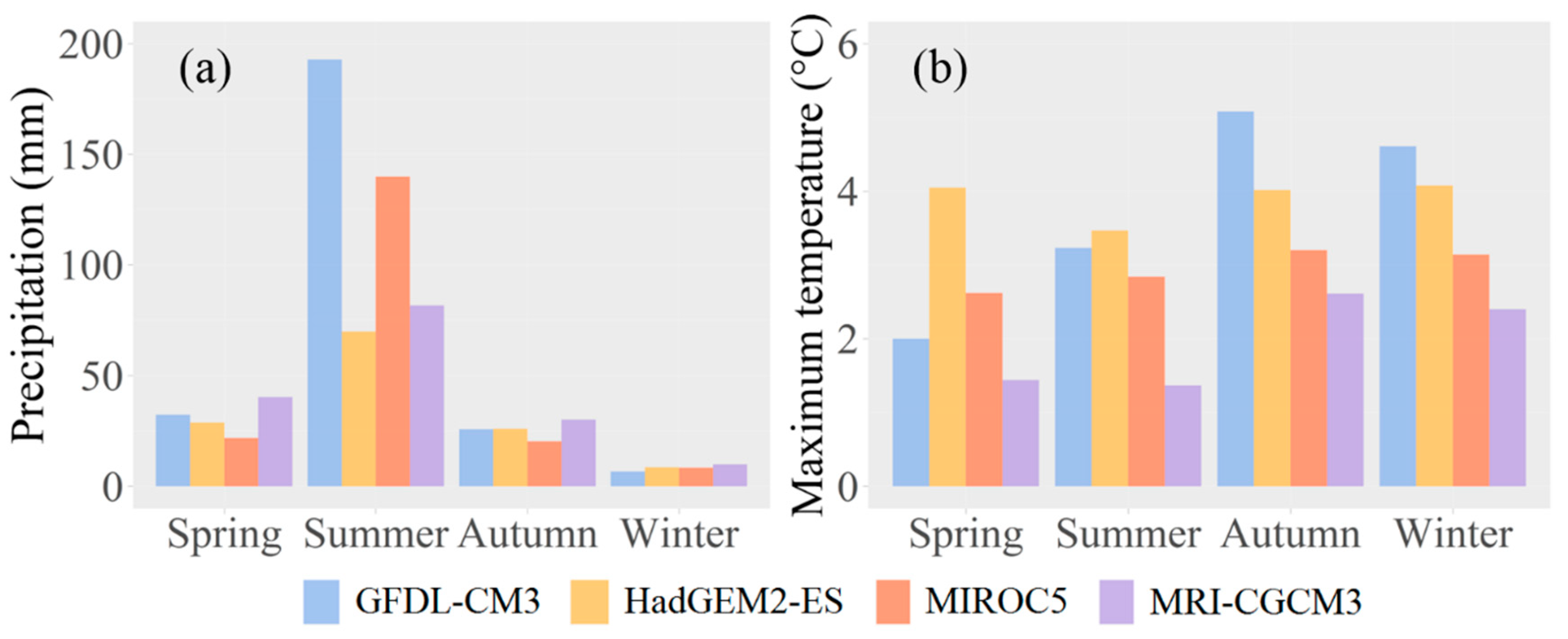
Figure 3.
The relative changes of Korean pine importance value (IV) (a), biomass (b), the abundance of juveniles (age less than 10 years old) (c), at the regional scale over time under the baseline, the resistant and the adaptive treatment scenarios, respectively. Error bands represented one standard deviation of each variable among four climate scenarios under each treatment scenario.
Figure 3.
The relative changes of Korean pine importance value (IV) (a), biomass (b), the abundance of juveniles (age less than 10 years old) (c), at the regional scale over time under the baseline, the resistant and the adaptive treatment scenarios, respectively. Error bands represented one standard deviation of each variable among four climate scenarios under each treatment scenario.
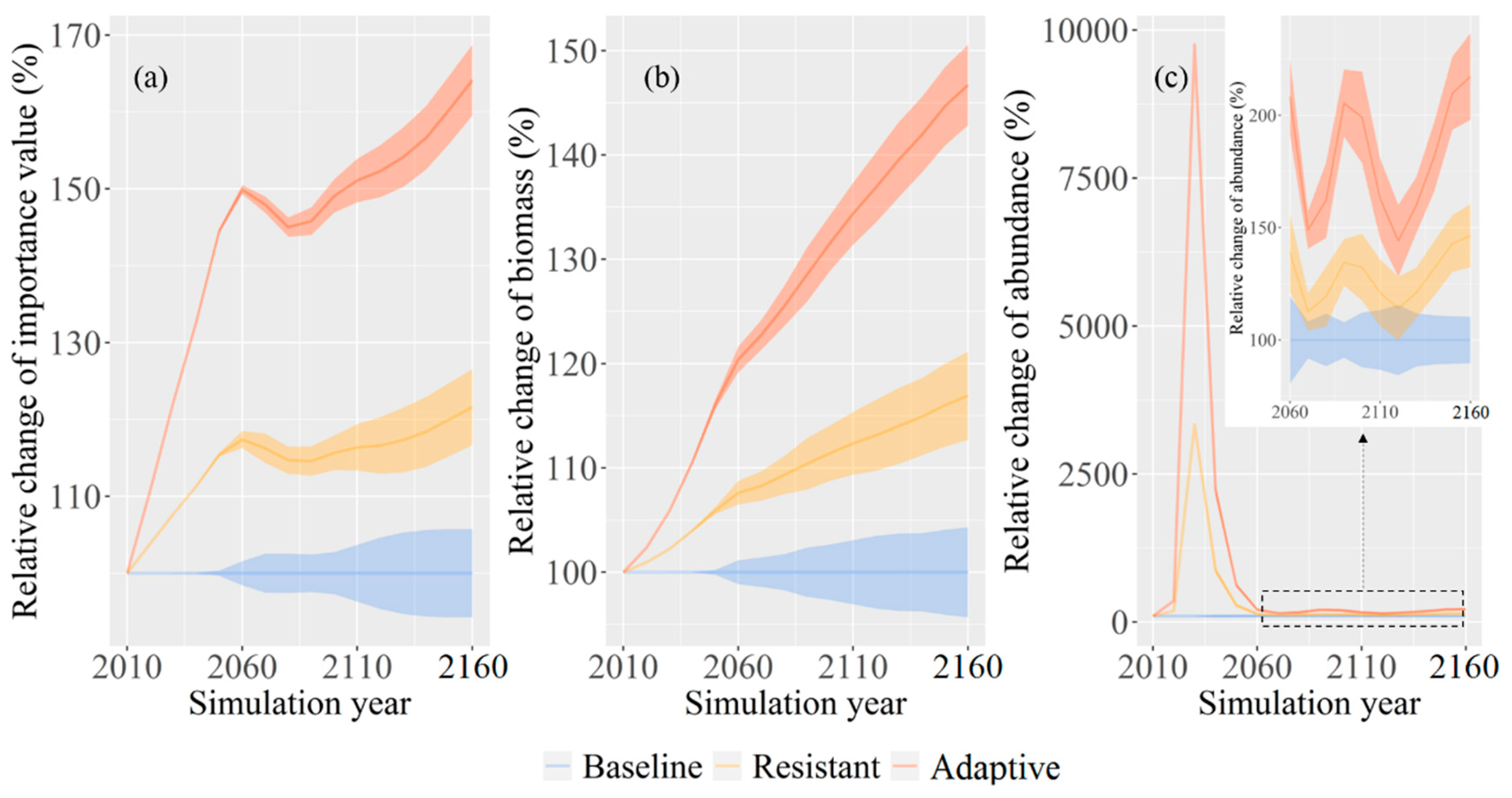
Figure 4.
Spatial pattern of the relative changes of mean Korean pine importance value (IV) (a–c), biomass (d–f), the abundance of juveniles (age less than 10 years old) (g–i), among four climate scenarios under the baseline, the resistant, and the adaptive treatment scenarios at the end of the simulation year, respectively.
Figure 4.
Spatial pattern of the relative changes of mean Korean pine importance value (IV) (a–c), biomass (d–f), the abundance of juveniles (age less than 10 years old) (g–i), among four climate scenarios under the baseline, the resistant, and the adaptive treatment scenarios at the end of the simulation year, respectively.
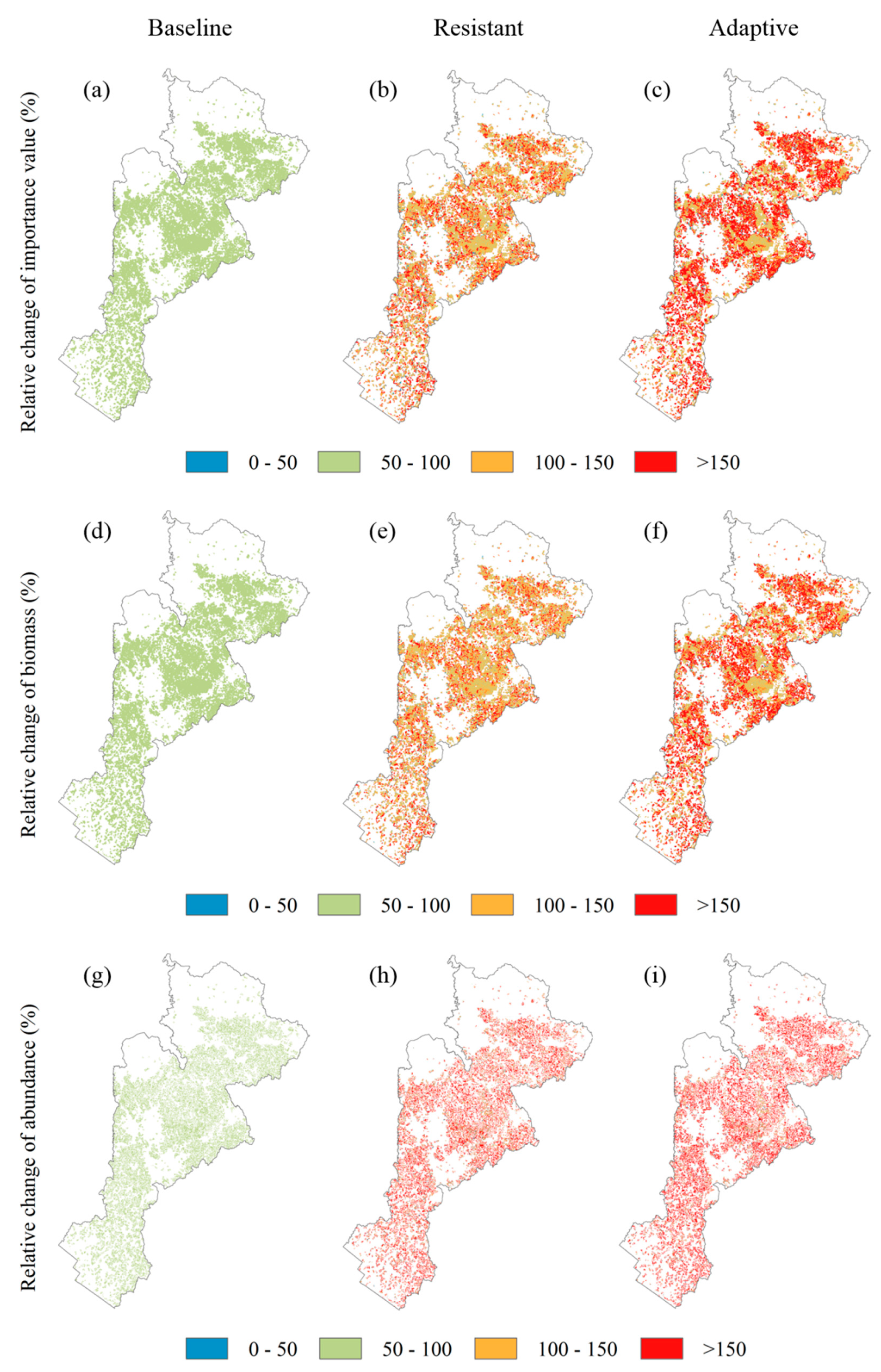
Figure 5.
Spatial pattern of uncertainties in the relative changes of Korean pine importance value (IV) (a–c), biomass (d–f), the abundance of juveniles (age less than 10 years old) (g–i), under the baseline, the resistant, and the adaptive treatment scenarios at the end of the simulation year, respectively. Uncertainty was a standard deviation of each variable among four climate scenarios under each treatment scenario.
Figure 5.
Spatial pattern of uncertainties in the relative changes of Korean pine importance value (IV) (a–c), biomass (d–f), the abundance of juveniles (age less than 10 years old) (g–i), under the baseline, the resistant, and the adaptive treatment scenarios at the end of the simulation year, respectively. Uncertainty was a standard deviation of each variable among four climate scenarios under each treatment scenario.

Figure 6.
The relative changes of the abundance (a) and distribution area (b) of Korean pine juveniles under the baseline, the resistant and the adaptive treatment scenarios at the end of the simulation year, respectively. Abundance was the mean values within each 1 km zones along the longitudinal direction, and distribution area was the mean cell number of existing juveniles (less than 10 years old) within the corresponding zones. Error bands represented one standard deviation of each variable among four climate scenarios under each treatment scenario.
Figure 6.
The relative changes of the abundance (a) and distribution area (b) of Korean pine juveniles under the baseline, the resistant and the adaptive treatment scenarios at the end of the simulation year, respectively. Abundance was the mean values within each 1 km zones along the longitudinal direction, and distribution area was the mean cell number of existing juveniles (less than 10 years old) within the corresponding zones. Error bands represented one standard deviation of each variable among four climate scenarios under each treatment scenario.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The major species parameters in the LINKAGES 3.0 model.
| Species | DMAX | DMIN | B3 | B2 | G | D3 | FROST | TL | FRT |
|---|---|---|---|---|---|---|---|---|---|
| Korean pine, Pinus koraiensis | 2500 | 900 | 0.29 | 62.96 | 104.53 | 0.28 | −35 | 12 | 2 |
| Korean spruce, Picea koraiensis and Picea jezoensis | 2500 | 600 | 0.35 | 63.62 | 87.79 | 0.19 | −34 | 11 | 3 |
| Khingan fir, Abies nephrolepis | 1800 | 400 | 0.40 | 67.36 | 87.65 | 0.23 | −34 | 10 | 3 |
| Larch, Larix gmelinii | 1900 | 400 | 0.32 | 60.27 | 87.92 | 0.42 | −38 | 12 | 1 |
| Manchurian ash, Fraxinus mandschurica | 2800 | 1000 | 0.34 | 67.26 | 121.90 | 0.19 | −31 | 2 | 1 |
| Manchurian walnut, Juglans mandshurica | 2650 | 850 | 0.23 | 41.40 | 72.89 | 0.28 | −32 | 2 | 1 |
| Amur corktree, Phellodendron amurense | 3200 | 1000 | 0.23 | 43.43 | 79.53 | 0.23 | −32 | 8 | 1 |
| Mongolian oak, Quercus mongolica | 3100 | 1100 | 0.32 | 60.27 | 87.92 | 0.51 | −34 | 9 | 1 |
| Black elm, Ulmus davidiana | 2700 | 900 | 0.35 | 63.62 | 105.35 | 0.18 | −33 | 5 | 1 |
| Mono maple, Acer mono | 3200 | 1000 | 0.52 | 62.10 | 89.69 | 0.23 | −32 | 2 | 1 |
| Ribbed birch, Betula costata | 1900 | 700 | 0.35 | 63.62 | 105.35 | 0.19 | −35 | 4 | 1 |
| Dahur Birch, Betula dahurica | 3100 | 600 | 0.75 | 74.52 | 118.67 | 0.49 | −35 | 4 | 1 |
| Amur linden, Tilia amurensis | 2400 | 800 | 0.40 | 67.36 | 87.65 | 0.23 | −33 | 2 | 1 |
| White birch, Betula platyphylla | 3100 | 600 | 0.75 | 74.52 | 118.67 | 0.41 | −38 | 4 | 1 |
| Poplar, Populus davidiana | 3000 | 700 | 0.66 | 78.77 | 146.52 | 0.33 | −34 | 7 | 1 |
DMAX, degree day maximum (degree); DMIN, degree day minimum (degree); B2 and B3 are growth parameter of Richard function; G, scales the growth rate; D3, proportion of growing season species can withstand drought; FROST, minimum January temperature species can withstand (celsius degree); TL, leaf litter type; FRT, foliage retention time (year).
Table 2.
Major species biological traits in the Small Khingan Mountains, Northeastern China.
| Species | MT/Long | ST | MD | MDBH | MSDI | NPGS |
|---|---|---|---|---|---|---|
| Korean pine | 40/300 | 4 | 150 | 110 | 550 | 20 |
| Korean spruce | 30/300 | 4 | 150 | 90 | 600 | 20 |
| Khingan fir | 30/300 | 4 | 150 | 85 | 650 | 20 |
| Larch | 20/300 | 2 | 300 | 95 | 650 | 30 |
| Manchurian ash | 30/250 | 3 | 300 | 100 | 600 | 25 |
| Manchurian walnut | 20/250 | 2 | 200 | 90 | 650 | 25 |
| Amur corktree | 20/250 | 3 | 300 | 95 | 650 | 25 |
| Mongolian oak | 20/300 | 2 | 200 | 95 | 600 | 20 |
| Black elm | 20/250 | 3 | 800 | 90 | 600 | 25 |
| Mono maple | 20/200 | 3 | 200 | 60 | 700 | 25 |
| Ribbed birch | 20/250 | 3 | 800 | 90 | 650 | 25 |
| Dahur Birch | 15/150 | 2 | 800 | 50 | 750 | 25 |
| Amur linden | 30/300 | 3 | 200 | 85 | 650 | 20 |
| White birch | 15/150 | 1 | 2000 | 50 | 800 | 30 |
| Poplar | 15/150 | 1 | 2000 | 60 | 800 | 30 |
MT, mature, age of tree species sexual maturity (year); Long, longevity, mean maximum age of tree species (year); ST, shade tolerance class (including 1–5, 1 and 5 mean the least and the most tolerance, respectively); MD, maximum dispersal distance (m); MDBH, maximum diameter at breast height (cm); MSDI, maximum stand density index (number of standard trees per ha, which is 10-inch tree); NPGS, number of potential germination seeds per mature tree (number/single cell).
Table 3.
Forest management treatments.
| Treatments | Removal Method | Minimum, Residual Stand Stocking (m3 ha−1) | Percent Area for Harvest, Rotation (Years) |
|---|---|---|---|
| Baseline | — | — | — |
| Resistant | Thinning from below | 46, 44 | 7%, 150 |
| Adaptive | Thinning from below | 46, 44 | 20%, 50 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, K.; He, H.; Xu, W.; Du, H.; Zong, S.; Huang, C.; Wu, M.; Tan, X.; Cong, Y. Responses of Korean Pine to Proactive Managements under Climate Change. Forests 2020, 11, 263. https://doi.org/10.3390/f11030263
AMA Style
Liu K, He H, Xu W, Du H, Zong S, Huang C, Wu M, Tan X, Cong Y. Responses of Korean Pine to Proactive Managements under Climate Change. Forests. 2020; 11(3):263. https://doi.org/10.3390/f11030263
Chicago/Turabian StyleLiu, Kai, Hongshi He, Wenru Xu, Haibo Du, Shengwei Zong, Chao Huang, Miaomiao Wu, Xinyuan Tan, and Yu Cong. 2020. "Responses of Korean Pine to Proactive Managements under Climate Change" Forests 11, no. 3: 263. https://doi.org/10.3390/f11030263
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.