1. Introduction
On a global scale, forests are known as one of the most dominant terrestrial ecosystems, covering 13 billion hectares or approximately 30.6% of the Earth’s total land area [
1]. Forests contribute in the storage of a large amount of biomass and play an important role in maintaining atmospheric carbon balance on Earth [
2]. Researchers have been facing a challenge for decades to enhance the accuracy of tree biomass and carbon estimations, which are crucial for understanding climate change, nutrient cycling, forest productivity, forest health, etc. [
3,
4,
5]. Measuring the actual weight of each tree component is unquestionably known as the most precise approach for estimating tree biomass. However, it is time-consuming, costly, and destructive. Thus, developing allometric biomass equations is acknowledged as a greater alternative means to estimate forest biomass [
6,
7,
8]. To date, a large number of allometric biomass models have been established for more than several hundreds of tree species in different ecological and geographical regions worldwide [
9,
10,
11,
12,
13].
When developing biomass equations to estimate components, subtotals, and total biomass of an individual tree, there are two common traits as a result of applying the different approach in fitting the biomass data, which are well-known as non-additive and additive. The former model separately fit the component, subtotal, and total biomass data disregarding the intrinsic correlations between components from the same individuals [
6,
14,
15]. Contrarily, the latter models simultaneously fit the component, subtotal, and total biomass data reflecting the intrinsic correlations between tree components of the same individuals. As a result, the subtotal or total biomass predictions will be equivalent to the summation of biomass component estimations [
8,
15,
16]. Several parameter estimation techniques have been proposed by researchers to address the compatibility property for a system of biomass equations [
6,
17,
18,
19,
20]. Among these methods, seemingly unrelated regression (SUR) and nonlinear seemingly unrelated regression (NSUR) appear to be more popular than others since they are more general and flexible in application [
17,
19,
21,
22,
23,
24].
To date, several model configurations have been developed in order to attain the additivity property between the component and subtotal or total biomass equations. Affleck and Diéguez-Aranda [
25] introduced an aggregated model system (denoted as AMS0), in which a number of N tree component nonlinear biomass equations are fitted jointly without using any parameter restriction (i.e., aboveground or total biomass). This model structure appeared as an alternative to the previous approach proposed by Parresol [
17]. Parresol’s approach also specifies a nonlinear biomass model for N tree components, and then aboveground or total biomass summation is defined as a parameter restriction in the aggregated model system (denoted as AMS1). Dong et al. [
19] implemented another model structure that was mentioned and recommended by Parresol [
17], in which the usage of parameter restriction is also necessary to ensure the additivity property between tree components. However, instead of using one parameter restriction, this aggregated model system (denoted as AMS3) employed three parameter restrictions defining the crown, aboveground, and total biomass as the summation of tree components. For the sake of comparison, here, these aggregated model systems are calculated by jointly fitting all N, N+1, and N+3 equations using weighted NSUR for AMS0, AMS1, and AMS3, respectively.
There are direct and indirect methods for determining the carbon stock of an individual tree. The direct method requires the carbon concentration to be measured initially by using the carbon analyzer to burn the tree samples [
26,
27]. Then, the obtained carbon concentration value can be utilized as the conversion factor to be multiplied by the actual dry biomass. On the contrary, the indirect method employs a generic carbon conversion factor multiplied by dry biomass value [
28]. There are several generic carbon conversion factor values that have been accepted globally: (1) the general carbon concentration value of 0.50 and (2) 0.45 for both non-woody and woody components [
29,
30]. However, recent studies revealed that the carbon concentration varies within a range of 0.44–0.56 depending upon the biomass components and the tree species. Thus, approximately 10% bias would be generated if one is relying on 0.50 as a generic carbon conversion factor to estimate carbon stock [
31,
32,
33]. Clearly, species-, component-, and even region-specific carbon concentration measurements are absolutely needed in order to decrease the inaccuracy of carbon stock quantification [
34].
Mongolian oak (
Quercus mongolica Fisch. ex Ledeb.) is a medium- to large-sized deciduous broadleaf tree species that is native to forested areas in eastern Asia and easternmost Russia. In China, it occupies the largest natural secondary forest regions, which is extensively distributed in the mountainous regions of Daxing’an, Xiaoxing’an, Zhangguangcai, and the northernmost end of the Changbai mountain area [
35]. Wang [
36] has developed the allometric equations of tree components, aboveground, and total biomass, and Zhang et al. [
37] have examined the variations of carbon concentration in biomass tissues for Mongolian oak in northeast China. However, both of these studies used a relatively small sample size, and particularly the biomass models developed by Wang [
36] were not additive. Therefore, the aims of this research were: (1) to construct three aggregated biomass model systems (i.e., AMS0, AMS2, and AMS3) with weighted NSUR based on different combinations of independent variables for Mongolian oak in northeast China; (2) to verify the predictive performance of the three systems using jackknifing techniques; (3) to evaluate six procedures of quantifying carbon stock of an individual tree; and (4) to compare the aggregated model systems developed in this study against the previously developed biomass equations.
4. Discussion
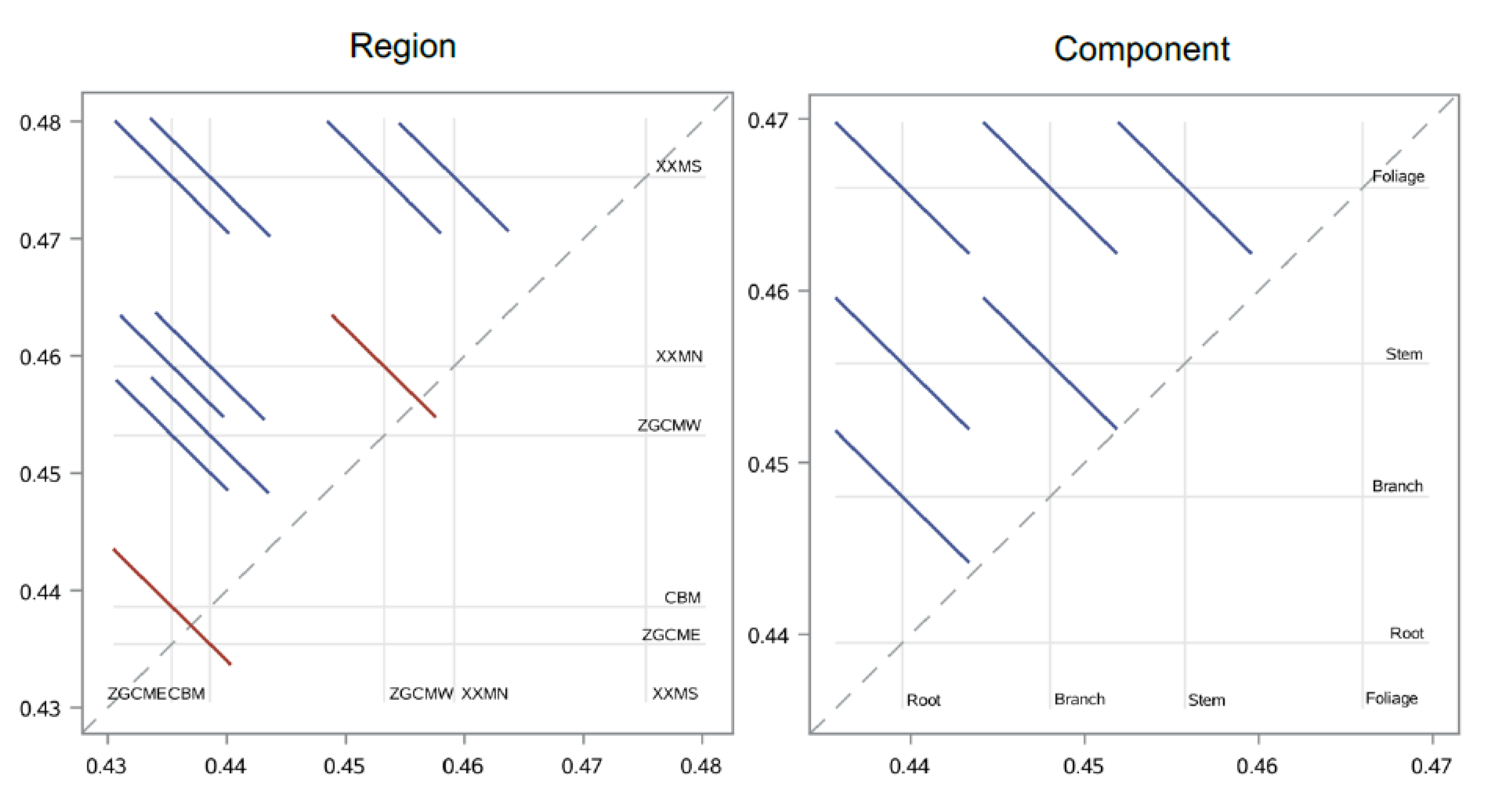
In the present study, the development of biomass allometric models and the analysis of carbon concentrations were based on a set of data having a broad range of tree diameters and a wide geographic coverage, allowing estimations to be more precise. Specifically, this comprehensive sampling strategy enabled the effects of various tree components on carbon concentrations of
Q. mongolica in five different growing regions, along with their variation within tree sizes, to be thoroughly and duly scrutinized. Carbon concentrations were found to be significantly different within tree components and tree-growing regions, ranging from 0.4194 ± 0.0171 in the root of ZGCME region to 0.4858 ± 0.0177 in the foliage of XXMS region. This finding is in accordance with those that have been reported elsewhere in the literature, which show significant differences in carbon concentration in components of the balsam fir in Canada [
43], 10 Chinese temperate tree species in northeast China [
37], and eight subtropical tree species in southern China [
44]. Moreover, Rodríguez-Soalleiro et al. [
34] confirmed the importance of biomass components and tree species as the source of variation in carbon concentration of several broadleaved species, which were found to explain about 52% and 17% of the variance, respectively. Overall, carbon concentration was found highest in foliage (0.4670), lowest in root (0.4406), and slightly lower in branches (0.4491) than stem (0.4568). Thomas and Malczewski [
26] and Zhang et al. [
37] studied the carbon content of the same specific species in northeast China. Thomas and Malczewski [
26] focused on analyzing the stem wood carbon content, while Zhang et al. [
37] reported the carbon content in all tree components. By comparison, the total range of carbon concentration across whole components in this study was figured to be more stable and narrower than reported by Zhang et al. [
37] (0.4620–0.5140), and the stem carbon concentration in this study was about 4% smaller than Thomas and Malczewski [
26].
To date, there are two completely opposing ways to calculate carbon stocks. Some researchers have used a constant generic 0.50 carbon concentration value to be multiplied by a biomass value [
45,
46,
47], in line with the recommendation of Roy et al. [
48] and Wenzl [
49]. On the contrary, some authors prefer to multiply the biomass value with the total weighted mean carbon concentration (TWMCc) of an individual tree [
50,
51], while others prefer to used respective mean component carbon concentration (RMCCc) as a multiplying factor to obtain carbon stock estimates [
52]. These second forms of approaches are considered a better approach since many studies have reported that carbon concentration varies greatly depending upon the type of tree species and tissues [
37,
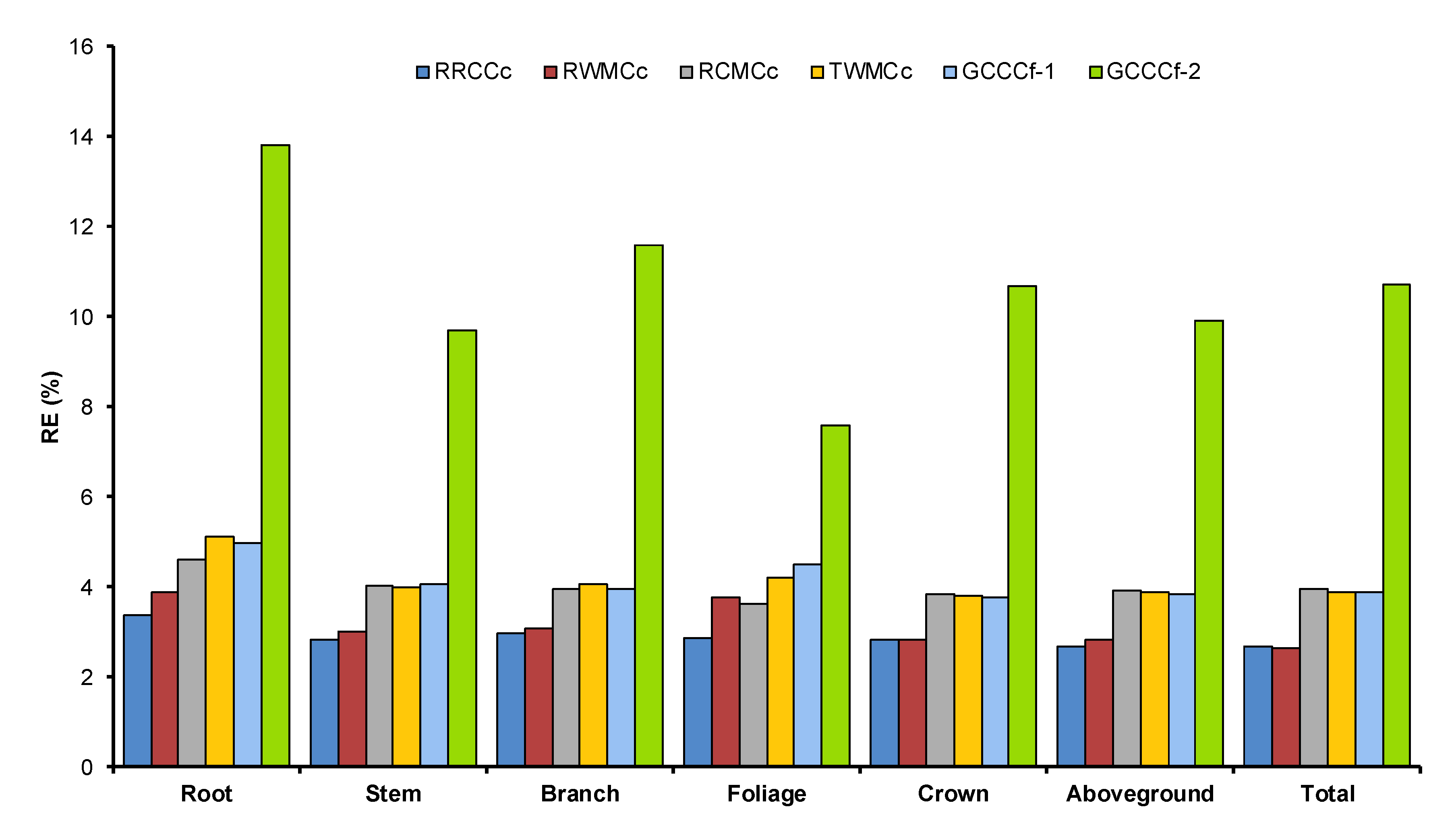
53]. Accordingly, using a generic carbon concentration conversion factor (0.50) may cause serious errors in estimating carbon stock. In the current study, six different alternative procedures of quantifying carbon stock were tested for Mongolian oak. The regional respective component carbon concentration (RRCCc) was the best procedure to estimate the carbon stock of an individual tree though implementing the RRCCc requires the comprehensive data of carbon concentration according to the tissue types and specific growing region. Hence, we propose the usage of a regional weighted mean carbon concentration (RWMCc) procedure as an appropriate alternative approach to quantify carbon stock, which performed equally as well as RRCCc. Our result evidence that employing RWMCc will only give 0.25% higher relative error compared with RRCCc. Our data and analyses firmly recommend against utilizing the generic carbon concentration proportion of 0.50 for Mongolian oak as it will likely lead to significant biases in yielding carbon stock estimation, just as it has been reported in other studies for other tree species [
28,
32,
39].
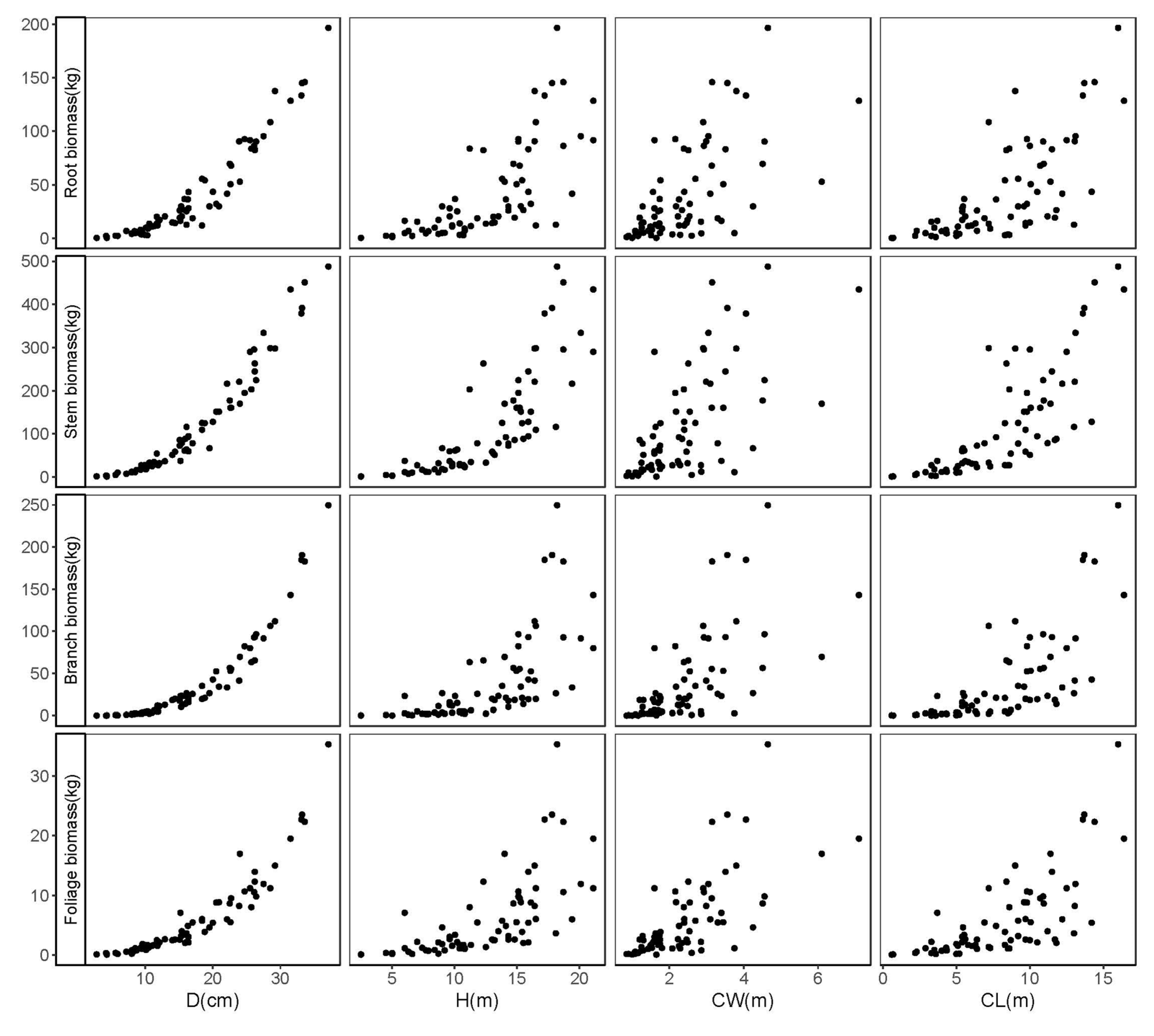
Tree diameter at breast height (D) is an indispensable predictor in biomass modeling and indubitably appears as among the most fundamental elements in forest inventory practice [
54,
55]. As a sole predictor in estimating tree biomass, D has always been providing an accurate model prediction for many species across numerous regions and delivering a simple model structure that is acquainted with being easy-to-use for forest practitioners [
56,
57]. The insertion of tree height (H) and crown characteristics (i.e., crown length [CL] and crown width [CW]) into biomass allometric models has been shown to generate a significant improvement in the model fitting and performance. In the current study, D, D and H, and D, H, and CL were used as a predictor(s) to develop each of the three systems of additive biomass model (Equations (1)–(4)). One of our available crown attribute predictors, CW, was intentionally eliminated in the model development since it was not significantly improving the model fitting performance for all tree component biomass equations. Our results noted that the inclusion of H and H and CL in the stem and foliage biomass equations, respectively, could improve the overall model fitting performance. This finding is in accordance with previous literature [
6,
15,
58,
59].
In this study, the aboveground (stem, branch, and foliage) and belowground (root) biomass data were obtained across a range of diameter (1.70–31.90 cm) and height (2.40–27.00 m). Previous research revealed that Mongolian oak might grow to exceed the diameter and height range investigated in this study [
60]. However, the existences of those large sizes of Mongolian oak are extremely rare in practice, especially in natural secondary forest. Moreover, the Chinese Government has strictly forbidden to cut the large size trees (D > 50 cm) for any kind of purposes. Thus, our newly developed aggregated biomass model systems are inappropriate for estimating the tree components, subtotals, and total biomass exceeding the diameter and height range provided in the present study.
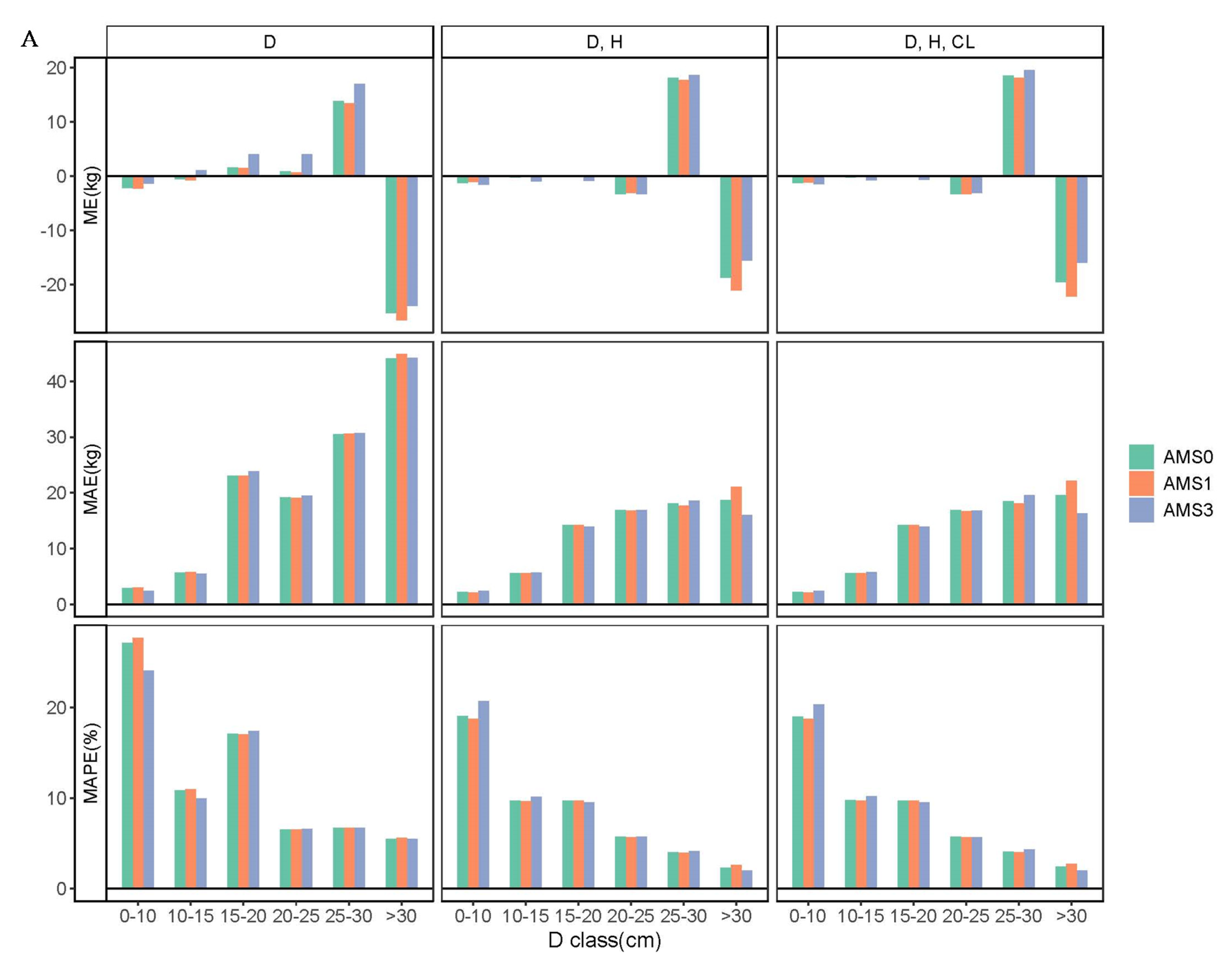
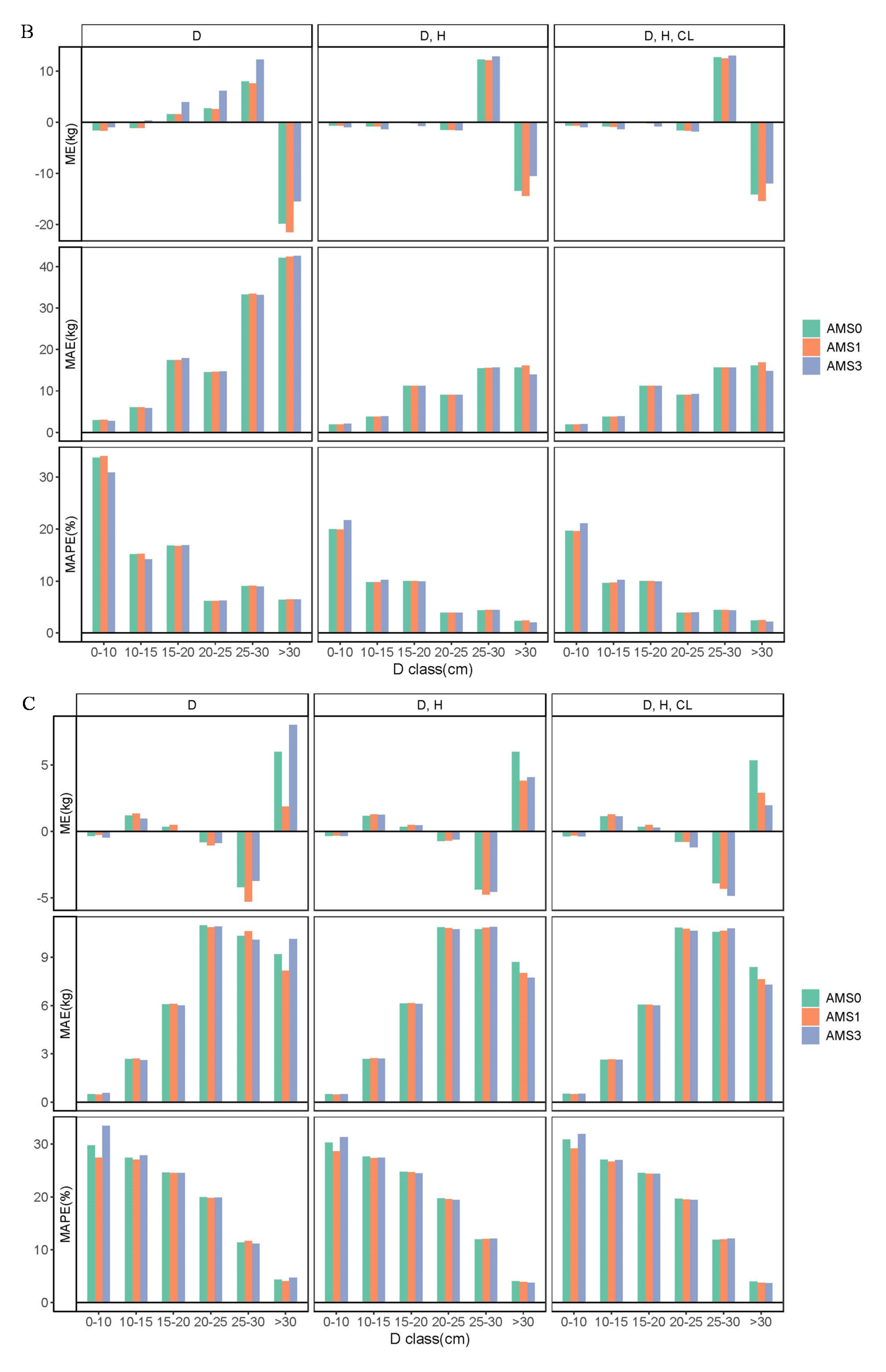
The three aggregative modeling approaches were constructed to develop biomass models. The AMS0 contains no parameter restriction, while the AMS1 and AMS3 contain one and three parameter restrictions, respectively, which guarantees that the sum of the component estimations will be exactly the same as the direct estimation of subtotals and total biomass. In this study, the focus was on demonstrating the differences between AMS0, AMS1, and AMS3 due to the parameter restrictions imposed on the model systems. Our results indicate that all the three aggregative modeling approaches performed well in the context of average prediction errors for components, subtotals, and total biomass estimations. Compared with AMS1 and AMS3, AMS0 was actually better to predict the component, crown, aboveground, and total biomass, although AMS1 only utilizes the total biomass model as a dependent equation, while AMS3 specifies all of the crown, aboveground, and total biomass. These results conform with the previous findings reported by Zhao et al. [
21] and Dong et al. [
24], which specifically stated that AMS0 is the most suitable approach to construct aggregated systems of biomass models.
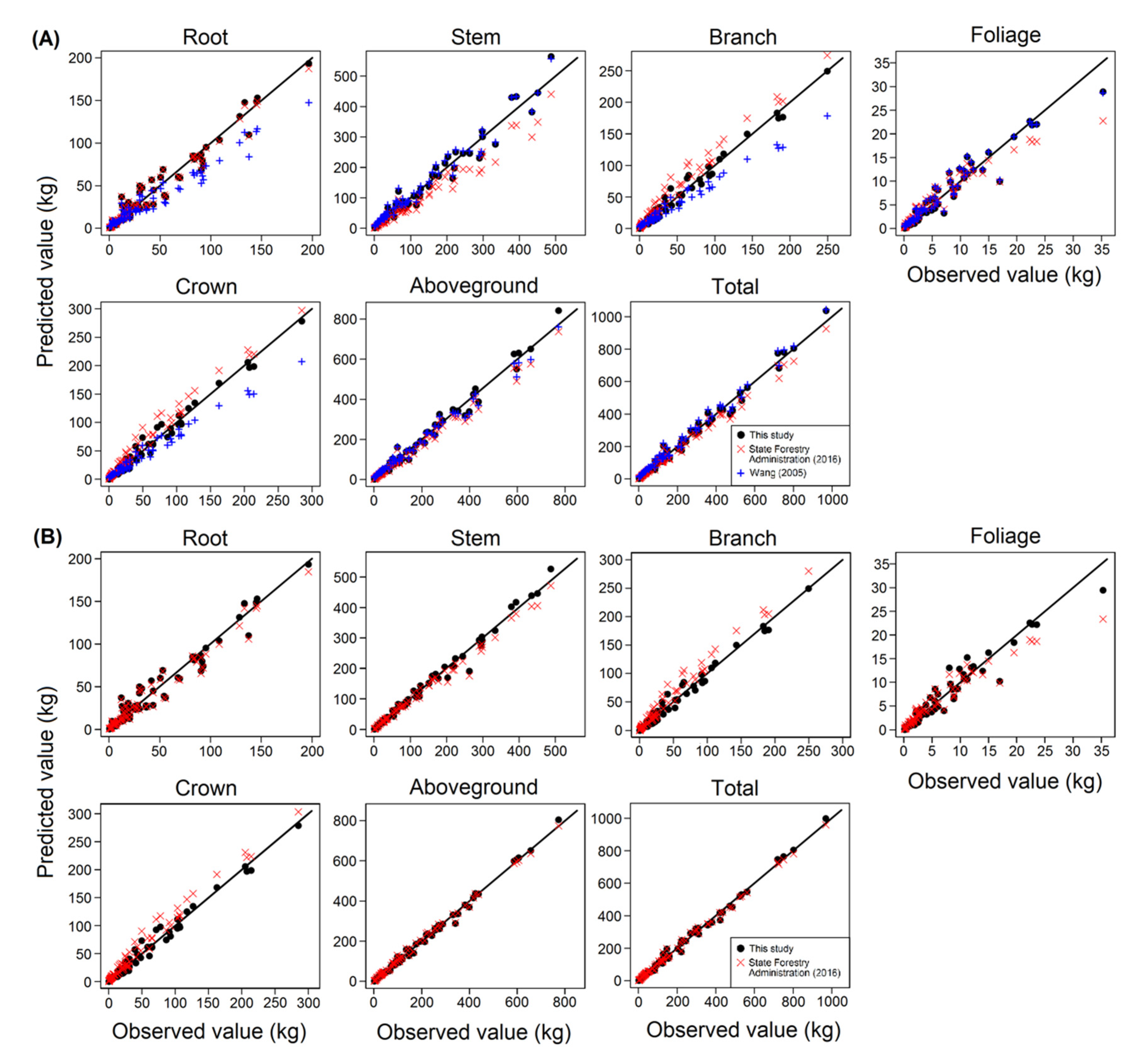
It was realized that a limited number of individual tree biomass equations exist for the particular tree species, especially in Northeast China. Wang [
36] developed biomass models for this species from the Maoershan Ecosystem Research Station in Heilongjiang Province, Northeastern China. The biomass models were also fitted by using a range of tree sizes, but unfortunately, the equations were lacking an additivity property for component, subtotal, and total biomass. Compared with the present research, Wang’s [
36] study produced similar predictions in estimating stem, aboveground, and total biomass for Mongolian oak, while there were some notable differences in estimating root, branch, and crown biomass. Moreover, the present study was compared with the previous equations developed by China State Forestry Administration [
42]. The State’s equations tend to give a distinguished underestimate of stem biomass and produce a remarkable overestimate prediction on branch and crown equations, chiefly for large-sized trees, whereas delivered similar estimations in estimating root, aboveground, and total biomass. The potential explanations of these discrepancies may be due to the differences in sampling sizes, sites, and the characteristic of stands where the trees were sampled. These issues may inevitably relate to the disparities in terms of climate variations, altitude, slope, and soil conditions as external factors; along with stand density, competition from the neighboring trees, and the morphological traits of tree roots as internal factors [
6,
61,
62].
5. Conclusions
We developed three aggregative systems of biomass equation for Mongolian oak occupying five mountainous regions in Northeast China: one based on D only, one on the combination of D and H, and one on the combination of D, H, and CL. We found that the inclusion of H into the aggregative systems improved the performances and model fit significantly for the stem, aboveground, and total biomass models, while adding CL into the aggregative model system only gave a slight improvement for the model fitting of the foliage and crown biomass models. All of the new systems developed here guarantee the additivity property between the components, subtotals, and total biomass, and provide more accurate estimations than previous research works. There was not available any single function or structural system that was perfect for estimating biomass for all of the tree components, crown, aboveground, or total biomass. Based on numerous fit statistics, the overall performances of the three aggregative model systems can be ranked in the following order: AMS0 > AMS1 > AMS3.
The carbon concentrations were found to be significantly different among tree components and growing regions in our natural secondary forest sites. Different procedures for quantifying the carbon stocks of Mongolian oak individual trees were evaluated. The results indicate that considering carbon concentration differences among tree tissues and growing regions appeared to be the best approach. Contrarily, the usage of a 0.50 generic carbon concentration value will give a relatively high bias in quantifying carbon stocks and evidently appeared as the worst method. The tree species used in this research is widespread in northeastern China. The aggregative biomass models and the carbon concentration analyses presented in this study might become an essential reference for researchers, forest practitioners, and governments, particularly for the Chinese National Forest Inventory. However, the newly developed model systems are not recommended to be used in estimating the biomass of trees outside the scope of the data, both geographically and in terms of tree size.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}