Ectomycorrhizal Fungi: Participation in Nutrient Turnover and Community Assembly Pattern in Forest Ecosystems

1
College of Resources and Environment, Fujian Agriculture and Forestry University, Fujian University Engineering Center of Soil Remediation, Fuzhou 350002, China
2
Key Laboratory of Environmental and Applied Microbiology, CAS, Environmental Microbiology Key Laboratory of Sichuan Province, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China
*
Author to whom correspondence should be addressed.
Forests 2020, 11(4), 453; https://doi.org/10.3390/f11040453
Submission received: 18 February 2020
/
Revised: 30 March 2020
/
Accepted: 15 April 2020
/
Published: 17 April 2020
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:Ectomycorrhizal fungi (EcMF) are involved in soil nutrient cycling in forest ecosystems. These fungi can promote the uptake of nutrients (e.g., nitrogen (N) and phosphorus (P)) and water by host plants, as well as facilitate host plant growth and resistance to stresses and diseases, thereby maintaining the aboveground primary productivity of forest ecosystems. Moreover, EcMF can acquire the carbon (C) sources needed for their growth from the host plants. The nutrient regulation mechanisms of EcMF mainly include the decay of soil organic matter via enzymatic degradation, nonenzymatic mechanism (Fenton chemistry), and priming effects, which in turn promote C and N cycling. At the same time, EcMF can secrete organic acids and phosphatases to improve the availability of soil P, or increase mycelium inputs to facilitate plant acquisition of P. The spatiotemporal distribution of EcMF is influenced by a combination of historical factors and contemporary environmental factors. The community of EcMF is associated with various factors, such as climate change, soil conditions, and host distribution. Under global climate change, investigating the relationships between the nutrient cycling functions of EcMF communities and their distribution patterns under various spatiotemporal scales is conducive to more accurate assessments of the ecological effects of EcMF on the sustainable development of forest.
1. Ectomycorrhizal Fungi and Forest Ecosystems
Mycorrhizal fungi are an important group of soil microorganisms that form symbiosis, namely, mycorrhiza, with 97% of known terrestrial plants [1]. According to the criteria for root morphological differentiation and host plant phylogeny, there are mainly four types of mycorrhiza: arbuscular mycorrhiza (AM), ectomycorrhiza (EcM), orchid mycorrhiza (OrM), and ericoid mycorrhiza (ErM) [2]. EcM is formed by fungi infecting the roots of approximately 2% of vascular plants (mainly including Pinaceae, Fagaceae, Dipterocarpaceae, and Myrtaceae) [2,3,4]; this type of mycorrhiza is structurally characterized by the presence of a mantle and a Hartig net, with intercellular hyphae that do not enter cells [5]. In total, approximately 7750 species of fungi are known to form EcM in the world, although a final estimate of ectomycorrhizal fungi (EcMF) species richness would likely be between 20,000 and 25,000 on the basis of estimates of knowns and unknowns in macromycete diversity [6]. These EcMF belong to more than 80 independently evolved lineages and also belong to more than 250 genera [7,8], mainly from Basidiomycota and Ascomycota [9].
EcMF are essential for plant nutrient uptake, seedling settlement, community restoration, and succession processes [10,11,12,13] (Figure 1a). Specifically, symbiosis between plant roots and EcMF are through a favorable exchange for photosynthetic product to enhance plant uptake of nitrogen (N) and phosphorus (P), thus playing a role in the carbon (C), N, and P cycles in forest ecosystems [5]. Moreover, seedlings exhibit enhanced resistance to pathogens due to the mycelial network, while their nutrient uptake is also promoted [11]. The mycelial networks formed by EcMF are also involved in water transport among plants [14]. Additionally, EcMF can relieve salt stress and heavy metal stress in host plants [15,16,17]. The involvement of EcMF in the material cycle and their nutritional relationships with plants are comprehensively regulated by host type, EcM species, and climatic and environmental conditions (temperature, rainfall, and N availability). Recently, based on a climate prediction model, Steidinger et al. [18] indicated that the global abundance of EcM-associated trees will decline by 10% by the end of 2070, and the maximum proportion of decline will occur in the boreal and temperate ecotone. In light of the decline in biodiversity that may occur under global climate change, studies of fungal composition and richness across habitats are crucial for understanding and predicting fungi–host–environment relationships and their influence on forest ecosystem function [19].
Numerous studies have investigated the spatiotemporal distribution patterns of EcMF at various scales (such as global, regional, and local scales). The results demonstrated that the variation of biogeographic patterns in EcMF communities was scale-dependent [20,21,22]. In addition, environmental selection (deterministic processes) and dispersal limitation (stochastic processes), as the main processes influencing biological community assembly in ecosystems [23,24,25,26] (Appendix A), could drive the community assembly of EcMF simultaneously [27,28].
2. Role of EcMF in Nutrient Turnover and Transport of Water in Forest Soils
2.1. EcMF Promote the C, N, and P Cycles through Involvement in Soil Organic Matter Turnover
Soil organic matter (SOM) refers to a mixture of C-containing organic compounds in soil, including soil microorganisms, soil animals, and their secretions, as well as plant residues and root exudates [33,39]. EcMF influence the dynamic changes in SOM mainly through two pathways: (1) EcMF alter organic matter turnover through enzymatic degradation, using oxidative and hydrolytic enzymes, as well as a nonenzymatic mechanism (Fenton chemistry) (Figure 1b) [29,40,41]. Enzymatic and nonenzymatic degradation involve two processes, N mineralization and C storage. N mineralization refers to EcMF oxidizing organic matter to varying degrees to acquire small molecules containing N while leaving abundant C substrates [39,42,43,44]. In the case of enzymatic reactions, EcMF produce extracellular enzymes, such as glycoside hydrolases, class II fungal peroxidases, glyoxal oxidases, and phenol oxidases, which take part in organic matter degradation and forage organic N [45,46,47] (Table 1). EcMF that are evolved from white-rot fungi have been shown to conduct enzymatic actions due to their retention of genes encoding lignocellulolytic enzymes [38,48,49]. For example, Hebeloma cylindrosporum Romagn. produces various enzymes, including glyoxal oxidases, peroxidases, cellobiohydrolase 6, cellobiohydrolase 7, and lytic polysaccharide monooxygenases; this species can oxidize polyphenols in organic matter using peroxidases [45]. Fenton chemistry produces hydroxyl radicals (˙OH) () [40,50]. ˙OH is involved in organic matter decay by EcMF through multiple pathways. ˙OH can directly liberate NH4+ from a variety of amino acids and peptides and can also oxidize organic matter, followed by degradation with proteases [40]. A study showed that the EcMF Paxillus involutus (Batsch) Fr. secreted the diarylcyclopentenone involutin during organic matter decay; in the presence of H2O2, involutin drove the Fenton reaction through Fe3+ reduction, similar to the mechanism of wood decay by brown-rot fungi [40]. EcMF that evolved from brown-rot fungi can release C from lignified tissue via the saprotrophic ancestor decay mechanism and thereby acquire organic N complexes from organic matter [40,50,51]. C storage refers to EcMF competing with free-living saprotrophs for N, which may limit the nutrients for saprotrophs and reduce their activity, thus delaying organic matter decomposition and increasing soil C accumulation; this process is also known as the Gadgil effect [35,52] (Figure 1b). Past studies showed that the removal of living roots and mycorrhizal fungi by a trenching method improved the decomposition rate of litter, due to the decreasing Gadgil effect [35,53]. However, there were also contradictory results suggesting that EcMF and free-living saprotrophs had different substrate preferences rather than competitive relationships; the isolation of roots and hyphae was not conducive to the growth of saprotrophs originally inhabiting the soil [54]. Moreover, as the C source of EcMF is mainly derived from the C supplied by their hosts, entry of more EcMF and extramatrical mycelium residues into soil may increase C storage in forests [55]; and (2) EcMF stimulate the decay of SOM through the priming effect of active hyphae [56] (Figure 1b). The priming effect refers to the secretion of labile C compounds by fine roots and mycorrhizal fungi to alleviate C limitations for free-living saprotrophs and simultaneously stimulate the mineralization of SOM (soil organic matter) [34,56,57]. Additionally, EcMF can stimulate organic matter decay through the priming effect under specific environmental conditions (e.g., elevated carbon dioxide (CO2) concentrations) [34,58]. Therefore, the priming effect promotes the growth of saprotrophic fungi by stimulating organic matter decay with EcMF, whereas the Gadgil effect inhibits the growth of saprotrophic fungi. These two effects may act simultaneously.
The capacity of EcMF for SOM decay is influenced by both organic matter characteristics and environmental factors: (1) The extent to which EcMF modify SOM depends on organic matter composition. For example, complexation of N-containing organic molecules with phenol molecules in organic matter is likely to delay decay. Physicochemical binding of organic matter to minerals and the formation of aggregates could also affect the process of organic matter decay [59,60,61]; (2) the capacity of EcMF to decay SOM could be positively correlated with the proportion of host C allocating to EcMF [57,62,63]. Because photosynthesis in host plants exhibits strong seasonal variability, the capacity of EcMF to decay SOM may also vary with seasons; (3) the capacity of EcMF to decay SOM is related to the availability of soil N, and could be inhibited following an increase in soil N availability (e.g., fertilizer application and atmospheric N deposition) [38,64,65]. It was found that the class II peroxidase activity of EcMF belonging to the genus Cortinarius was reduced in a boreal forest supplied with inorganic N [49]; (4) the capacity of EcMF to decay SOM is also associated with their functional genes. As EcMF have evolved independently from saprotrophic ancestors more than 85 times [7], distinct lineages retained various functional genes of saprotrophs during their evolutionary process, leading to differences in the mode and capacity of organic matter decay [39]; and (5) EcMF are contributors to organic matter after their death, and the different biochemical components of various EcMF species influence organic matter decay [62]. For example, chitin is modified easily, possibly providing an important source of soil C and N; by contrast, melanin is not modified easily, reducing the overall tissue decomposition ability [33]. Moreover, the different morphologies and biochemical composition of dead hyphae can influence their rate of decay [66]. In short, SOM decay is subjected to simultaneous regulation by the abovementioned five factors. The relative importance of each factor varies with the interaction of organic matter with the mineral surface, the seasonal distribution of photosynthetic products from the host, the composition of microbial communities, and the type of ecosystem [67].
In forest ecosystems, P is usually derived from organic compounds, and organic phosphate (Po) is mainly present in the form of phospholipids, which can be absorbed and used by plants only after degradation and leaching. EcMF can mobilize Po and mineral P in soil and hence affect P uptake by plants via the following two pathways. First, EcMF release organic acids, phosphomonoesterases (alkaline and acid phosphatases), phosphodiesterases, and phytases that act on recalcitrant Po in soil [32,68,69,70], thereby increasing soil available P content (Figure 1b). Specifically, low-molecular-weight organic acids liberated by EcMF, including oxalate, citrate, and malate, can increase the P availability from insoluble P sources [71]. Additionally, EcMF can promote the mineralization of Po by cleaving the ester bonds with extracellular phosphatases, which in turn liberates phosphate for plant uptake [72]. Under relatively high N availability, EcMF secrete more phosphatases, and soil phosphatase activity is increased, thus enhancing soil P uptake and transport ability [73]. Second, EcMF significantly increase the volume of soil occupied by fine roots through the production of massive hyphae, which broadens the range of plant P uptake [5]. Additionally, EcMF transfer inorganic phosphate (Pi) from the soil to colonizing roots at the symbiotic interface via their expressed P transporters, such as the H+:Pi transporter [36,74]. Moreover, other mechanisms such as Pi:Na+, low-affinity Pi and Po transporters may exist in EcMF simultaneously [36] (Figure 1c). However, these mechanisms by which EcMF transfer available P sources and Pi to plant cells have not been fully understood [36]. Additionally, the type and morphology of EcMF hyphae influence P uptake by host plants. Bödeker et al. [75] showed that a high diversity of EcMF facilitated P uptake by Fagus sylvatica L. and that the taxa with abundant extramatrical mycelium appeared to be more beneficial for P acquisition than were the taxa of the contact exploration type.
2.2. EcMF Are Involved in Other Nutrient Element (K, Ca, Mg, etc.) Cycles
Apart from their involvement in N and P acquisition, EcMF also promote the acquisition of other elements, such as potassium (K), magnesium (Mg), and calcium (Ca) by host plants [76,77]. EcMF can accelerate the weathering of minerals and rocks through acidification, complexation, and physical fragmentation [78], which liberates available metal ions into the soil (Figure 1d). In the acidification process, organic acids secreted by EcMF reduce pH in the rhizosphere soil and increase the exchange capacity of soluble and exchangeable cations (e.g., K+, Ca2+, and Mg2+), thereby liberating these mineral elements [37]. Low-molecular-weight organic acids produced by EcMF can also form a complex with metal cations (e.g., K+, Ca2+, and Mg2+) and thus accelerate mineral weathering [78]. Furthermore, the extramatrical mycelium of EcMF can enter soil minerals and rock cracks, leading to physical fragmentation [79].
2.3. EcMF Are Involved in Water Transport
Furthermore, mycelial networks formed by EcMF take part in water transport to regulate plant water requirements [80] (Figure 1a). Mycelial networks can transfer water among plants of the same and different species [14], which not only ensures water for seedling growth [81] but also provides water to the host to cope with drought stress [82]. Evidence indicates that under drought conditions, EcM can increase water uptake and regulate aquaporins through enormous mycelial networks, thus improving plant drought tolerance [82,83,84]. However, the relative contribution of the hyphal pathway to plant water uptake compared to root water uptake is still unclear [85]. Moreover, the water transport capacity of EcMF varies depending on hyphal morphology and exploration type. For example, Thelephora terrestris Ehrh., which forms hydrophobic hyphae, can promote water transfer among plants, whereas Suillus granulatus (L.) Roussel lacks the capacity for water transport [81].
3. Spatio-Temporal Distribution Patterns and the Driving Factors of EcMF Communities
3.1. Spatial Distribution Patterns and Driving Factors of EcMF Communities
Significant variations in EcMF richness and community composition exist in forest ecosystems across climatic zones. A global meta-analysis revealed that EcMF richness followed a unimodal pattern along the latitudinal gradient from tropical rainforest to subarctic tundra [21]. Specifically, the richness was highest in temperate and boreal forests and was relatively low in tropical lowland environments and subarctic regions [21] (more details are provided in Figure 5 in [21]). This pattern is inconsistent with the phenomenon observed in most macroorganisms, in that richness is negatively correlated with latitudinal increases [21]. On a global scale, /tomentella-thelephora and /russula-lactarius are the most abundant of EcMF lineages, and they can establish a mutualistic symbiosis with multiple host plants in various ecosystems [9]. Based on variations in community composition along the latitudinal gradient, the globe can be roughly divided into three EcMF distribution zones: northern temperate regions, Austral regions (including Australia, New Zealand, and southern South America), and tropical regions [9,86]. Among them, EcMF taxa in northern temperate regions are the richest, and many lineages, such as /leucangium, /geopora, and /hydnotrya, are unique to northern temperate regions; /austropaxillus is a unique lineage found in Austral regions; and Guyanagarika is a unique genus found in tropical regions [9,86,87] (more details are presented in Figure 2 in [9]).
The richness of EcMF is mainly influenced by climate factors on a global scale [21]. EcMF richness exhibits a unimodal pattern with latitude because greater environmental selection occurs at the two ends of the temperature gradient, which in turn results in relatively low phylogenetic diversity of EcMF in lowland tropical and subarctic regions [21,88,89]. EcMF richness increases with decreasing precipitation; perhaps as precipitation decreases, more species of EcMF are beneficial for the host to resist drought stress [21]. Host plants also significantly influence the phylogenetic diversity of EcMF. Most hosts of EcMF in Asia, Africa, and Oceania were found to belong to the same family: 83% to Dipterocarpaceae in Asia, 58% to Fabaceae in Africa, and 64% to Myrtaceae in Oceania; however, the family distribution of several host plants in the neotropical region was relatively even [4] (Table 2, more details are shown in Table 1 and Figure 3 in [4]). Therefore, the diversity of host plants is also involved in shaping the global distribution pattern of EcMF. It was shown that the host plants of EcMF had low diversity and abundance in tropical forest regions dominated by plants forming symbiosis with arbuscular mycorrhizal fungi [9,90,91]. By contrast, extremely high EcMF richness was found in tropical forest regions dominated by host plants of EcMF [92,93]. The distribution of EcMF is also regulated by other microorganisms that are in a symbiotic relationship with the host. For example, global and regional studies on the EcMF community associated with Alnus plants revealed the presence of N-fixing Frankia in Alnus plant roots [94,95]. Hence, the community structure of EcMF associated with Alnus plants might be subjected to selective limitation by the symbiotic regulation system of Frankia, and taxa adapted to the high-N environment were therefore selected [94,95,96]. Additionally, geographic isolation, spatial heterogeneity of soil physiochemical properties, nutrient requirements of host plants, and C allocation of host plants to EcMF all affect the symbiosis between a plant and EcMF [70].
Unlike its global trend, the richness of EcMF communities in Chinese forest ecosystems increase from tropical to cold temperate zone, possibly because of Pinaceae, which can form a symbiotic relationship with EcMF, is widely distributed in the cold temperate zone [20]. A study conducted in a secondary forest ecosystem showed that the phylogeny of host plant and geographic distance played an important role in shifting the EcMF community associated with Betulaceae plants [95]. As EcMF have certain host preferences and the specific characteristics of the tree species affect the habitat properties, the choice of tree species in afforestation influences mycorrhizal colonization [97,98,99]. At a local scale, the key factor driving the community assembly of EcMF was the intensity of forest management, whereas the key factors at a large scale (>1000 km) were soil pH and root resource traits (including the root C/N ratio and the mineral nutrition ratio between the root and soil) [27].
The differences in EcMF communities at the regional and local scales are due to the difference in combination of climatic factors, host characteristics, and soil characteristics at different sites. In summary, at the regional and global scales, the community assembly of EcMF is mainly influenced by environmental factors.
3.2. Temporal Evolution Patterns and Community Assembly Mechanisms
Historical factors and contemporary environmental heterogeneity are important factors that influence EcMF community composition. Phylogeographical studies have enabled the identification of the origin of fungi and reproduction of their historical migration routes [100]. Many EcMF lineages might have originated in high latitudes and subsequently migrated to tropical regions [21,101]. Studies have found that three EcMF lineages with a worldwide distribution (/russula-lactarius, /clavulina, and /inocybe) all originated in the tropics [4,102,103]. Additionally, Li et al. [104] indicated that Strobilomyces might have originated in Africa during the early Eocene and then spread to Southeast Asia through the boreotropical forest; during the Oligocene, Strobilomyces was dispersed from Southeast Asia northward and southward to East Asia and Australasia, respectively; and during the Miocene, Strobilomyces migrated across the Bering land bridge, from East Asia to North America, and then was dispersed from the Asian continent to Japan through the East China Sea basin. At the same time, the climatic changes from the middle Miocene to the present might have driven the separation and differentiation between the regions [104]. Dispersion has played a crucial role in the formation of the current distribution pattern of Strobilomyces, and during its migration process across different continents, there was a shift in host plants from Dipterocarpaceae to Pinaceae and Fagaceae in East Asia and the northern temperate regions and then to Myrtaceae and Nothofagaceae in Oceania [86,104]. The EcMF communities of a circumpolar distribution possibly migrated during the glacier cycle, and long-distance dispersal may have played a major role in EcMF community assembly during the phylogeographical history of high-latitude areas in the northern hemisphere [105].
Stand development age and seasonal change can also influence the community composition and assembly mechanisms of EcMF. During different successional stages of subtropical secondary forests, the abundances of the /cortinarius and /elaphomyces lineages were found to decrease significantly from 10- to 40-year-old to 41- to 80-year-old forests, whereas the abundance of /tomentella-thelephora exhibited an inverse trend [106]. The EcMF community in old forests (>80 years) was influenced by environmental selection and dispersal limitation; however, in young (10–40 years) to mature forests (41–80 years), the community was influenced by environmental selection only, possibly because the process from young to mature forest was controlled by various environmental factors [106].
Additionally, the amount of EcMF extramatrical mycelium varied significantly with the season. Simulation experiment conducted in Mediterranean coniferous forests showed that the reduction in soil moisture and the increase in soil temperature resulted in a significant increase in extramatrical mycelium of Lactarius vinosus (Quél.) Bataille during winter–spring, and a significant decrease during summer [107]. The increase in temperature would lengthen the growth cycle of L. vinosus in winter, and the reduction of soil moisture would prevent the growth of L. vinosus in summer, suggesting that the cycle shifts of soil mycelium was driven by climatic factors [107].
In summary, the development and distribution of EcMF are influenced by different environmental conditions and historical evolution processes, including both natural variation and human interference, which lead to differences in the spatiotemporal evolutional mechanism of EcMF communities. Deterministic and stochastic processes often act on the community assembly of EcMF simultaneously, but this effect has strong scale dependence. On a relatively large spatial scale, environmental selection among the deterministic processes has the most pronounced influence on the present community composition of EcMF, followed by stochastic processes, albeit with the influence of stochastic factors differing in time and space; during the phylogeographical history of EcMF, stochastic factors play more important roles [108,109].
3.3. The Impact of Environmental Change on EcMF
EcMF communities are affected by various environmental factors, such as the concentration of atmospheric CO2 and atmospheric temperature, soil nutrient availability, and soil water. Studies have suggested that with an increase in the concentration of atmospheric CO2, the EcMF community structure might change, and the abundance of species with hyphae of the long-distance exploration type increase [19,110]. A plausible reason is that species with hyphae of the long-distance exploration type require more C than species of the short-distance exploration type to maintain larger structures [19]. It was previously thought that as the CO2 concentration has increased, N limitation might be enhanced in temperate and boreal forests, while P limitation would become more severe in tropical forests [19,111]. With an increasing concentration of atmospheric CO2, the host might preferentially make an association with species forming hyphae of the long-distance exploration type, thereby enhancing N and P acquisition [19]. However, it was shown that the host allocated more C to EcMF following a CO2 concentration elevation, which could increase N immobilization and thereby lead to N limitation of the host [112].
The richness of EcMF communities decreases with increasing atmospheric temperature [113,114]. Analysis of the EcMF temperature niche position on the Japanese archipelago revealed that the richness and diversity of the EcMF community are highest in cold regions; this further indicates that climate warming might limit the distribution of EcMF only adapted to the cold climate and thus reduce EcMF diversity [113]. However, there exist considerable differences in the temperature sensitivity and temperature adaptability among various EcMF taxa. Compared with other taxa, the genus Cortinarius exhibited a greater adaptability to temperature increases, while its richness did not change significantly with increasing temperature [114].
Different taxa of EcMF exhibit certain preferences for N forms [19]. The taxa with hydrophobic extramatrical mycelium of the middle- and long-distance exploration types are more inclined to mobilize difficult-to-decompose N [115]; by contrast, the taxa with smooth hyphae of the contact, short-, and middle-distance exploration types are more suitable for the acquisition of labile N due to their low proteolytic abilities [115,116]. EcMF are highly sensitive to the addition of inorganic N, and their peroxidase activity decreases substantially with increasing concentrations of inorganic N [49,117]. When the concentration of inorganic N in the environment is relatively high, EcMF reduces organic N decay. Under long-term N deposition, the richness of EcMF taxa may decline, and the community structure could change, with a shift from taxa that are beneficial for N uptake in a low-N environment to taxa that have a high overall nutrient use efficiency and, eventually, to taxa with the ability to uptake P in an N-rich environment [116,118].
Soil moisture affects the root colonization and community structure of EcMF [75]. In soil containing excessive water, hypoxia leads to an inhibition of mycorrhization [119]. In an arid and seasonally water-deficient environment, some EcMF taxa prone to melanization of the cell wall (e.g., Cenococcum geophilum Fr.) may become dominant, as the melanization of cell walls enhances drought tolerance in EcMF [120,121,122]. Moreover, the exploration type of extramatrical mycelium of EcMF might change under water stress [121], from primarily contact exploration like Lactarius tabidus Fr. to mainly short-distance exploration like C. geophilum and Genea hispidula Berk. ex Tul. & C. Tul. and middle-distance rhizomorph exploration like Tomentellopsis submollis (Svrček) Hjortstam [75].
Variation in the ability to use different N-containing compounds, melanin deposition in the cell wall, and a shift in the hyphal exploration type are important characteristics of EcMF under the fluctuations in environmental conditions [19]. Changes in these characteristics could ultimately influence ecosystem functions [19]. Studies on the distribution of EcMF and its participation in nutrient cycling in a specific ecosystem contribute to the prediction of ecosystem function under global climate change [39].
4. Conclusions
In this paper, we summarized the nutrient acquisition strategy and community assembly model of EcMF as well as the driver factors in the spatiotemporal distribution of EcMF communities at different scales based on current studies. The nutrient cycling functions of EcMF communities are different among ecosystems, and the community assembly model of EcMF communities are driven by different factors at various scales, suggesting ecosystem and scale need to be combined simultaneously in predicting forest ecosystem cycling, using the ecological function of EcMF under global climate change.
5. Research Prospects
Based on the existing information, research prospects are proposed from the following perspectives:
(1) Existing studies on the involvement of EcMF in nutrient cycling have mainly focused on organic matter decay and the C and N cycles. However, most past studies were conducted in EcMF peroxidase activity based on pure cultures or on a simulated environment. As organic matter composition is more complicated in natural conditions, the capacity of EcMF for organic matter decay needs to be further investigated. Compared with arbuscular mycorrhizal fungi, EcMF are still insufficiently studied with respect to their mechanisms and contribution in the transport and uptake of P and other nutrients. Future studies should combine field in situ experiments and laboratory simulations to further explore the mechanisms of EcMF in forest ecosystems and quantify their contribution to nutrient cycling.
(2) Combining microbial ecological theory with community assembly pattern at various spatiotemporal scales, a distribution model of EcMF can be constructed to predict the change in EcMF communities across broader spatiotemporal scales.
(3) Many studies have been conducted to investigate the distribution patterns and assembly mechanisms of EcMF across global and regional scales. However, past studies have regional limitations, such as a preference for temperate and cold regions, while research on the African continent and in East Asia is still limited [86]. Additionally, EcMF studies in tropical regions are mainly concentrated in lowland tropical rainforests and tropical montane forests, while tropical savannas and dry forests have been less investigated [4]. The dominant hosts rarely overlap across different regions, and there may be many undiscovered plant species [4]. Therefore, EcMF richness in the tropics may be underestimated.
(4) The relative contribution of contemporary environmental factors and historical evolutionary factors to the spatial variation in EcMF communities is still largely controversial. Some studies indicated that the biotic and abiotic factors included in the experiments only explained part of the community variation; future studies need to explore the role of environmental variables and stochastic processes more comprehensively. The variation of EcMF community in a chronosequence could be revealed by the community characteristics at different succession stages in spatial position. In the future, a soil biological sample bank containing the samples collected and treated by standardizing procedures can be established to study the evolution of EcMF communities over a larger spatial scale and chronosequence. Furthermore, most controlled experiments on EcMF responses to environmental change only involve the change in one or two environmental factors. However, due to the interactions between environmental factors, multivariate experimental methods may be more useful to demonstrate EcMF responses to global climate change.
Author Contributions
Y.K. and X.L. designed and revised the manuscript; Y.L. analyzed and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Sichuan Science and Technology Program (No. 2018SZDZX0030) and the West Light Foundation of Chinese Academy of Sciences (No. Y8C2041100).
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
| Deterministic processes | The role of environmental selection and the cooperation and antagonism among species determine community assembly. |
| Stochastic processes | Unpredictable disturbance, random extinction, and ecological drift are important for community assembly. |
| Dispersal limitation | Movement of organisms to or establishment of organisms in a new location is restricted. |
References
- Guo, L. Progress of microbial species diversity research in China. Biodiversity Sci. 2012, 20, 572–580. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Marcel, G.A.; van der Heijden, M.G.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Corrales, A.; Henkel, T.W.; Smith, M.E. Ectomycorrhizal associations in the tropics–biogeography, diversity patterns and ecosystem roles. New Phytol. 2018, 220, 1076–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008; pp. 189–385. [Google Scholar]
- Rinaldi, A.C.; Comandini, O.; Kuyper, T.W. Ectomycorrhizal fungal diversity: Separating the wheat from the chaff. Fungal Divers. 2008, 33, 1–45. [Google Scholar]
- Tedersoo, L.; Smith, M.E. Lineages of ectomycorrhizal fungi revisited: Foraging strategies and novel lineages revealed by sequences from belowground. Fungal Biol. Rev. 2013, 27, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Geml, J.Z. Altitudinal Gradients in Mycorrhizal Symbiose. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer International: Cham, Switzerland, 2017; Volume 230, pp. 107–125. [Google Scholar]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef] [PubMed]
- Teste, F.P.; Kardol, P.; Turner, B.L.; Wardle, D.A.; Zemunik, G.; Renton, M.; Laliberté, E. Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands. Science 2017, 355, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E.; Lambers, H.; Burgess, T.I.; Wright, S.J. Phosphorus limitation, soil-borne pathogens and the coexistence of plant species in hyperdiverse forests and shrublands. New Phytol. 2015, 206, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Veselá, P.V.M.; Hofmannová, K.; Edwards-Jonášová, M.; Cudlín, P. Ectomycorrhizal Community on Norway Spruce Seedlings Following Bark Beetle Infestation. Forests 2019, 10, 740. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, R.E.; Chapin, F.S., 3rd; Hollingsworth, T.N.; Taylor, D.L. The potential for mycobiont sharing between shrubs and seedlings to facilitate tree establishment after wildfire at Alaska arctic treeline. Mol. Ecol. 2017, 26, 3826–3838. [Google Scholar] [CrossRef] [PubMed]
- Egerton-Warburton, L.M.; Querejeta, J.I.; Allen, M.F. Common mycorrhizal networks provide a potential pathway for the transfer of hydraulically lifted water between plants. J. Exp. Bot. 2007, 58, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Zwiazek, J.J.; Equiza, M.A.; Karst, J.; Senorans, J.; Wartenbe, M.; Calvo-Polanco, M. Role of urban ectomycorrhizal fungi in improving the tolerance of lodgepole pine (Pinus contorta) seedlings to salt stress. Mycorrhiza 2019, 29, 303–312. [Google Scholar] [CrossRef]
- Guerrero-Galan, C.; Calvo-Polanco, M.; Zimmermann, S.D. Ectomycorrhizal symbiosis helps plants to challenge salt stress conditions. Mycorrhiza 2019, 29, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.B.; Wu, C.H.; Zhang, C.; Li, H.; Lipka, U.; Polle, A. The role of ectomycorrhizas in heavy metal stress tolerance of host plants. Environ. Exp. Bot. 2014, 108, 47–62. [Google Scholar] [CrossRef]
- Steidinger, B.S.; Crowther, T.W.; Liang, J.; Van Nuland, M.E.; Werner, G.D.A.; Reich, P.B.; Nabuurs, G.J.; de-Miguel, S.; Zhou, M.; Picard, N.; et al. Climatic controls of decomposition drive the global biogeography of forest-tree symbioses. Nature 2019, 569, 404–408. [Google Scholar] [CrossRef] [Green Version]
- Koide, R.T.; Fernandez, C.; Malcolm, G. Determining place and process: Functional traits of ectomycorrhizal fungi that affect both community structure and ecosystem function. New Phytol. 2014, 201, 433–439. [Google Scholar] [CrossRef]
- Hu, Y.; Veresoglou, S.D.; Tedersoo, L.; Xu, T.; Ge, T.; Liu, L.; Chen, Y.; Hao, Z.; Su, Y.; Rillig, M.C.; et al. Contrasting latitudinal diversity and co-occurrence patterns of soil fungi and plants in forest ecosystems. Soil Biol. Biochem. 2019, 131, 100–110. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Toots, M.; Diedhiou, A.G.; Henkel, T.W.; Kjøller, R.; Morris, M.H.; Nara, K.; Nouhra, E.; Peay, K.G. Towards global patterns in the diversity and community structure of ectomycorrhizal fungi. Mol. Ecol. 2012, 21, 4160–4170. [Google Scholar] [CrossRef]
- Bahram, M.; Peay, K.G.; Tedersoo, L. Local-scale biogeography and spatiotemporal variability in communities of mycorrhizal fungi. New Phytol. 2015, 205, 1454–1463. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Nelson, M.; Helgason, T.; Dytham, C.; Fitter, A.H. Relative roles of niche and neutral processes in structuring a soil microbial community. ISME J. 2010, 4, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langenheder, S.; Szekely, A.J. Species sorting and neutral processes are both important during the initial assembly of bacterial communities. ISME J. 2011, 5, 1086–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottenie, K. Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Hollingsworth, T.N.; McFarland, J.W.; Lennon, N.J.; Nusbaum, C.; Ruess, R.W. A first comprehensive census of fungi in soil reveals both hyperdiversity and fine-scale niche partitioning. Ecol. Monogr. 2014, 84, 3–20. [Google Scholar] [CrossRef]
- Schroter, K.; Wemheuer, B.; Pena, R.; Schoning, I.; Ehbrecht, M.; Schall, P.; Ammer, C.; Daniel, R.; Polle, A. Assembly processes of trophic guilds in the root mycobiome of temperate forests. Mol. Ecol. 2019, 28, 348–364. [Google Scholar] [CrossRef]
- Vincenot, L.; Selosse, M.-A. Population Biology and Ecology of Ectomycorrhizal Fungi. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer International: Cham, Switzerland, 2017; Volume 230, pp. 39–60. [Google Scholar]
- Frey, S.D. Mycorrhizal fungi as mediators of soil organic matter dynamics. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 237–259. [Google Scholar] [CrossRef]
- Deslippe, J.R.; Hartmann, M.; Grayston, S.J.; Simard, S.W.; Mohn, W.W. Stable isotope probing implicates a species of Cortinarius in carbon transfer through ectomycorrhizal fungal mycelial networks in A rctic tundra. New Phytol. 2016, 210, 383–390. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Critchley, C.; Ng, H.; Bledsoe, C. Reciprocal N (15NH4+ or 15NO3−) transfer between nonN2-fixing Eucalyptus maculata and N2-fixing Casuarina cunninghamiana linked by the ectomycorrhizal fungus Pisolithus sp. New Phytol. 2004, 163, 629–640. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef]
- Fernandez, C.W.; Langley, J.A.; Chapman, S.; McCormack, M.L.; Koide, R.T. The decomposition of ectomycorrhizal fungal necromass. Soil Biol. Biochem. 2016, 93, 38–49. [Google Scholar] [CrossRef]
- Brzostek, E.R.; Dragoni, D.; Brown, Z.A.; Phillips, R.P. Mycorrhizal type determines the magnitude and direction of root-induced changes in decomposition in a temperate forest. New Phytol. 2015, 206, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.W.; Kennedy, P.G. Revisiting the ‘Gadgil effect’: Do interguild fungal interactions control carbon cycling in forest soils? New Phytol. 2016, 209, 1382–1394. [Google Scholar] [CrossRef] [PubMed]
- Plassard, C.; Becquer, A.; Garcia, K. Phosphorus transport in Mycorrhiza: How far are we? Trends Plant Sci. 2019, 24, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Landeweert, R.; Hoffland, E.; Finlay, R.D.; Kuyper, T.W.; van Breemen, N. Linking plants to rocks: Ectomycorrhizal fungi mobilize nutrients from minerals. Trends Ecol. Evol. 2001, 16, 248–254. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi – potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Zak, D.R.; Pellitier, P.T.; Argiroff, W.; Castillo, B.; James, T.Y.; Nave, L.E.; Averill, C.; Beidler, K.V.; Bhatnagar, J.; Blesh, J.; et al. Exploring the role of ectomycorrhizal fungi in soil carbon dynamics. New Phytol. 2019, 223, 33–39. [Google Scholar] [CrossRef]
- Op De Beeck, M.; Troein, C.; Peterson, C.; Persson, P.; Tunlid, A. Fenton reaction facilitates organic nitrogen acquisition by an ectomycorrhizal fungus. New Phytol. 2018, 218, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Nicolás, C.; Martin-Bertelsen, T.; Floudas, D.; Bentzer, J.; Smits, M.; Johansson, T.; Troein, C.; Persson, P.; Tunlid, A. The soil organic matter decomposition mechanisms in ectomycorrhizal fungi are tuned for liberating soil organic nitrogen. ISME J. 2019, 13, 977–988. [Google Scholar] [CrossRef] [Green Version]
- Orwin, K.H.; Kirschbaum, M.U.; St John, M.G.; Dickie, I.A. Organic nutrient uptake by mycorrhizal fungi enhances ecosystem carbon storage: A model-based assessment. Ecol. Lett. 2011, 14, 493–502. [Google Scholar] [CrossRef]
- Averill, C.; Turner, B.L.; Finzi, A.C. Mycorrhiza-mediated competition between plants and decomposers drives soil carbon storage. Nature 2014, 505, 543. [Google Scholar] [CrossRef]
- Phillips, R.P.; Brzostek, E.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon-nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Kohler, A.; Kuo, A.; Nagy, L.G.; Morin, E.; Barry, K.W.; Buscot, F.; Canbäck, B.; Choi, C.; Cichocki, N.; Clum, A. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 2015, 47, 410. [Google Scholar] [CrossRef] [PubMed]
- Courty, P.-E.; Pouysegur, R.; Buée, M.; Garbaye, J. Laccase and phosphatase activities of the dominant ectomycorrhizal types in a lowland oak forest. Soil Biol. Biochem. 2006, 38, 1219–1222. [Google Scholar] [CrossRef]
- Tedersoo, L.; Naadel, T.; Bahram, M.; Pritsch, K.; Buegger, F.; Leal, M.; Kõljalg, U.; Põldmaa, K. Enzymatic activities and stable isotope patterns of ectomycorrhizal fungi in relation to phylogeny and exploration types in an afrotropical rain forest. New Phytol. 2012, 195, 832–843. [Google Scholar] [CrossRef]
- Pellitier, P.T.; Zak, D.R. Ectomycorrhizal fungi and the enzymatic liberation of nitrogen from soil organic matter: Why evolutionary history matters. New Phytol. 2018, 217, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Bödeker, I.T.M.; Clemmensen, K.E.; de Boer, W.; Martin, F.; Olson, A.; Lindahl, B.D. Ectomycorrhizal Cortinarius species participate in enzymatic oxidation of humus in northern forest ecosystems. New Phytol. 2014, 203, 245–256. [Google Scholar] [CrossRef]
- Rineau, F.; Roth, D.; Shah, F.; Smits, M.; Johansson, T.; Canbäck, B.; Olsen, P.B.; Persson, P.; Grell, M.N.; Lindquist, E.; et al. The ectomycorrhizal fungus Paxillus involutus converts organic matter in plant litter using a trimmed brown-rot mechanism involving Fenton chemistry. Environ. Microbiol. 2012, 14, 1477–1487. [Google Scholar] [CrossRef] [Green Version]
- Shah, F.; Schwenk, D.; Nicolas, C.; Persson, P.; Hoffmeister, D.; Tunlid, A. Involutin Is an Fe3+ reductant secreted by the ectomycorrhizal fungus Paxillus involutus during fenton-based decomposition of organic matter. Appl. Environ. Microbiol. 2015, 81, 8427–8433. [Google Scholar] [CrossRef] [Green Version]
- Averill, C.; Hawkes, C.V. Ectomycorrhizal fungi slow soil carbon cycling. Ecol. Lett. 2016, 19, 937–947. [Google Scholar] [CrossRef]
- Sterkenburg, E.; Clemmensen, K.E.; Ekblad, A.; Finlay, R.D.; Lindahl, B.D. Contrasting effects of ectomycorrhizal fungi on early and late stage decomposition in a boreal forest. ISME J. 2018, 12, 2187–2197. [Google Scholar] [CrossRef] [Green Version]
- Sietio, O.-M.; Santalahti, M.; Putkinen, A.; Adamczyk, S.; Sun, H.; Heinonsalo, J. Restriction of plant roots in boreal forest organic soils affects the microbial community but does not change the dominance from ectomycorrhizal to saprotrophic fungi. FEMS Microbiol. Ecol. 2019, 95, fiz133. [Google Scholar] [CrossRef] [PubMed]
- Godbold, D.L.; Hoosbeek, M.R.; Lukac, M.; Cotrufo, M.F.; Janssens, I.A.; Ceulemans, R.; Polle, A.; Velthorst, E.J.; Scarascia-Mugnozza, G.; De Angelis, P.; et al. Mycorrhizal hyphal turnover as a dominant process for carbon input into soil organic matter. Plant Soil 2006, 281, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Sulman, B.N.; Brzostek, E.R.; Medici, C.; Shevliakova, E.; Menge, D.N.; Phillips, R.P. Feedbacks between plant N demand and rhizosphere priming depend on type of mycorrhizal association. Ecol. Lett. 2017, 20, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Jackson, O.; Quilliam, R.S.; Stott, A.; Grant, H.; Subke, J.A. Rhizosphere carbon supply accelerates soil organic matter decomposition in the presence of fresh organic substrates. Plant Soil 2019, 440, 473–490. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.P.; Meier, I.C.; Bernhardt, E.S.; Grandy, A.S.; Wickings, K.; Finzi, A.C. Roots and fungi accelerate carbon and nitrogen cycling in forests exposed to elevated CO2. Ecol. Lett. 2012, 15, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Torn, M.S.; Trumbore, S.E.; Chadwick, O.A.; Vitousek, P.M.; Hendricks, D.M. Mineral control of soil organic carbon storage and turnover. Nature 1997, 389, 170–173. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Deckmyn, G.; Meyer, A.; Smits, M.; Ekblad, A.; Grebenc, T.; Komarov, A.; Kraigher, H. Simulating ectomycorrhizal fungi and their role in carbon and nitrogen cycling in forest ecosystems. Can. J. For. Res. 2014, 44, 535–553. [Google Scholar] [CrossRef]
- Terrer, C.; Vicca, S.; Hungate, B.A.; Phillips, R.P.; Prentice, I.C. Mycorrhizal association as a primary control of the CO2 fertilization effect. Science 2016, 353, 72–74. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, S.G.; Taylor, B.N.; Cooper, E.R.; Beidler, K.V.; Strand, A.E.; McCormack, M.L.; Zhang, S.Y. Long-term dynamics of mycorrhizal root tips in a loblolly pine forest grown with free-air CO2 enrichment and soil N fertilization for 6 years. Glob. Chang. Biol. 2014, 20, 1313–1326. [Google Scholar] [CrossRef]
- Averill, C.; Dietze, M.C.; Bhatnagar, J.M. Continental-scale nitrogen pollution is shifting forest mycorrhizal associations and soil carbon stocks. Glob. Chang. Biol. 2018, 24, 4544–4553. [Google Scholar] [CrossRef] [PubMed]
- Certano, A.K.; Fernandez, C.W.; Heckman, K.A.; Kennedy, P.G. The afterlife effects of fungal morphology: Contrasting decomposition rates between diffuse and rhizomorphic necromass. Soil Biol. Biochem. 2018, 126, 76–81. [Google Scholar] [CrossRef]
- Keller, A.B.; Phillips, R.P. Leaf litter decay rates differ between mycorrhizal groups in temperate, but not tropical, forests. New Phytol. 2019, 222, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Jansa, J.; Finlay, R.; Wallander, H.; Smith, F.A.; Smith, S.E. Role of Mycorrhizal Symbioses in Phosphorus Cycling. In Phosphorus in Action: Biological Processes in Soil Phosphorus Cycling; Bünemann, E., Oberson, A., Frossard, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 26, pp. 137–168. [Google Scholar]
- Liu, X.B.; Burslem, D.; Taylor, J.D.; Taylor, A.F.S.; Khoo, E.; Majalap-Lee, N.; Helgason, T.; Johnson, D. Partitioning of soil phosphorus among arbuscular and ectomycorrhizal trees in tropical and subtropical forests. Ecol. Lett. 2018, 21, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Ruess, R.W.; Swanson, M.M.; Kielland, K.; McFarland, J.W.; Olson, K.D.; Taylor, D.L. Phosphorus Mobilizing Enzymes of Alnus-Associated Ectomycorrhizal Fungi in an Alaskan Boreal Floodplain. Forests 2019, 10, 554. [Google Scholar] [CrossRef] [Green Version]
- Hodge, A. Accessibility of Inorganic and Organic Nutrients for Mycorrhizas. In Mycorrhizal Mediation of Soil; Johnson, N.C., Gehring, C., Jansa, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 129–148. [Google Scholar]
- Treseder, K.K.; Lennon, J.T. Fungal traits that drive ecosystem dynamics on land. Microbiol. Mol. Biol. Rev. 2015, 79, 243–262. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, T.; Kataoka, R.; Futai, K. Plant growth and nutrition in pine (Pinus thunbergii) seedlings and dehydrogenase and phosphatase activity of ectomycorrhizal root tips inoculated with seven individual ectomycorrhizal fungal species at high and low nitrogen conditions. Soil Biol. Biochem. 2008, 40, 1235–1243. [Google Scholar] [CrossRef]
- Wang, J.; Li, T.; Wu, X.; Zhao, Z. Molecular cloning and functional analysis of a H+-dependent phosphate transporter gene from the ectomycorrhizal fungus Boletus edulis in southwest China. Fungal Biol. 2014, 118, 453–461. [Google Scholar] [CrossRef]
- Kohler, J.; Yang, N.; Pena, R.; Raghavan, V.; Polle, A.; Meier, I.C. Ectomycorrhizal fungal diversity increases phosphorus uptake efficiency of European beech. New Phytol. 2018, 220, 1200–1210. [Google Scholar] [CrossRef]
- Neba, G.A.; Newbery, D.M.; Chuyong, G.B. Limitation of seedling growth by potassium and magnesium supply for two ectomycorrhizal tree species of a Central African rain forest and its implication for their recruitment. Ecol. Evol. 2016, 6, 125–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmalenberger, A.; Duran, A.L.; Bray, A.W.; Bridge, J.; Bonneville, S.; Benning, L.G.; Romero-Gonzalez, M.E.; Leake, J.R.; Banwart, S.A. Oxalate secretion by ectomycorrhizal Paxillus involutus is mineral-specific and controls calcium weathering from minerals. Sci. Rep. 2015, 5, 12187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Jia, L.M. Research progress of mineral weathering by ectomycorrhizal fungi. Chin. J. Ecol. 2014, 33, 3447–3454. [Google Scholar] [CrossRef]
- Bonneville, S.; Smits, M.M.; Brown, A.; Harrington, J.; Leake, J.R.; Brydson, R.; Benning, L.G. Plant-driven fungal weathering: Early stages of mineral alteration at the nanometer scale. Geology 2009, 37, 615–618. [Google Scholar] [CrossRef]
- Marjanović, Ž.; Nehls, U. Ectomycorrhiza and water transport. In Mycorrhiza: State of the Art, Genetics and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure and Systematics; Varma, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 149–159. [Google Scholar]
- Prieto, I.; Roldán, A.; Huygens, D.; del Mar Alguacil, M.; Navarro-Cano, J.A.; Querejeta, J.I. Species-specific roles of ectomycorrhizal fungi in facilitating interplant transfer of hydraulically redistributed water between Pinus halepensis saplings and seedlings. Plant Soil 2016, 406, 15–27. [Google Scholar] [CrossRef]
- Lehto, T.; Zwiazek, J.J. Ectomycorrhizas and water relations of trees: A review. Mycorrhiza 2011, 21, 71–90. [Google Scholar] [CrossRef]
- Long, D.; Liu, J.; Han, Q.; Wang, X.; Huang, J. Ectomycorrhizal fungal communities associated with Populus simonii and Pinus tabuliformis in the hilly-gully region of the Loess Plateau, China. Sci. Rep. 2016, 6, 24336. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Kemppainen, M.; El Kayal, W.; Lee, S.H.; Pardo, A.G.; Cooke, J.E.; Zwiazek, J.J. Overexpression of Laccaria bicolor aquaporin JQ585595 alters root water transport properties in ectomycorrhizal white spruce (Picea glauca) seedlings. New Phytol. 2015, 205, 757–770. [Google Scholar] [CrossRef]
- Johnson, D. Water, water everywhere … but how does it affect the functional diversity of ectomycorrhizal fungi? New Phytol. 2018, 220, 950–951. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Yang, Z.L. Ectomycorrhizal symbioses: Diversity of mycobionts and molecular mechanisms that entail the development of ectomycorrhizae (in Chinese). Sci. Sin. Vitae 2019, 49, 436–444. [Google Scholar] [CrossRef]
- Sanchez-Garcia, M.; Henkel, T.W.; Aime, M.C.; Smith, M.E.; Matheny, P.B. Guyanagarika, a new ectomycorrhizal genus of Agaricales from the Neotropics. Fungal Biol. 2016, 120, 1540–1553. [Google Scholar] [CrossRef] [Green Version]
- Bjorbaekmo, M.F.M.; Carlsen, T.; Brysting, A.; Vralstad, T.; Hoiland, K.; Ugland, K.I.; Geml, J.; Schumacher, T.; Kauserud, H. High diversity of root associated fungi in both alpine and arctic Dryas octopetala. BMC Plant Biol. 2010, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Nara, K. General latitudinal gradient of biodiversity is reversed in ectomycorrhizal fungi. New Phytol. 2010, 185, 351–354. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Bahram, M.; Kõljalg, U.; Courty, P.-E.; Diédhiou, A.G.; Kjøller, R.; Põlme, S.; Ryberg, M.; Veldre, V.; Tedersoo, L.; Wurzburger, N. The distance decay of similarity in communities of ectomycorrhizal fungi in different ecosystems and scales. J. Ecol. 2013, 101, 1335–1344. [Google Scholar] [CrossRef]
- Henkel, T.W.; Aime, M.C.; Chin, M.M.L.; Miller, S.L.; Vilgalys, R.; Smith, M.E. Ectomycorrhizal fungal sporocarp diversity and discovery of new taxa in Dicymbe monodominant forests of the Guiana Shield. Biodivers. Conserv. 2012, 21, 2195–2220. [Google Scholar] [CrossRef]
- Smith, M.E.; Henkel, T.W.; Aime, M.C.; Fremier, A.K.; Vilgalys, R. Ectomycorrhizal fungal diversity and community structure on three co-occurring leguminous canopy tree species in a Neotropical rainforest. New Phytol. 2011, 192, 699–712. [Google Scholar] [CrossRef]
- Polme, S.; Bahram, M.; Yamanaka, T.; Nara, K.; Dai, Y.C.; Grebenc, T.; Kraigher, H.; Toivonen, M.; Wang, P.H.; Matsuda, Y.; et al. Biogeography of ectomycorrhizal fungi associated with alders (Alnus spp.) in relation to biotic and abiotic variables at the global scale. New Phytol. 2013, 198, 1239–1249. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.L.; Gao, C.; Chen, L.; Ji, N.N.; Wu, B.W.; Li, X.C.; Lu, P.P.; Zheng, Y.; Guo, L.D. Host plant phylogeny and geographic distance strongly structure Betulaceae-associated ectomycorrhizal fungal communities in Chinese secondary forest ecosystems. FEMS Microbiol. Ecol. 2019, 95, 15. [Google Scholar] [CrossRef]
- Tedersoo, L.; Suvi, T.; Jairus, T.; Ostonen, I.; Polme, S. Revisiting ectomycorrhizal fungi of the genus Alnus: Differential host specificity, diversity and determinants of the fungal community. New Phytol. 2009, 182, 727–735. [Google Scholar] [CrossRef]
- Polme, S.; Bahram, M.; Jacquemyn, H.; Kennedy, P.; Kohout, P.; Moora, M.; Oja, J.; Opik, M.; Pecoraro, L.; Tedersoo, L. Host preference and network properties in biotrophic plant-fungal associations. New Phytol. 2018, 217, 1230–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahnmann, B.; Mašínová, T.; Halvorsen, R.; Davey, M.L.; Sedlák, P.; Tomšovský, M.; Baldrian, P. Effects of oak, beech and spruce on the distribution and community structure of fungi in litter and soils across a temperate forest. Soil Biol. Biochem. 2018, 119, 162–173. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Peay, K.G.; Kennedy, P.G.; Talbot, J.M. Dimensions of biodiversity in the Earth mycobiome. Nat. Rev. Microbiol. 2016, 14, 434–447. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Ryberg, M.; Otsing, E.; Koljalg, U.; Abarenkov, K. Global biogeography of the ectomycorrhizal /sebacina lineage (Fungi, Sebacinales) as revealed from comparative phylogenetic analyses. Mol. Ecol. 2014, 23, 4168–4183. [Google Scholar] [CrossRef]
- Matheny, P.B.; Aime, M.C.; Bougher, N.L.; Buyck, B.; Desjardin, D.E.; Horak, E.; Kropp, B.R.; Lodge, D.J.; Soytong, K.; Trappe, J.M.; et al. Out of the Palaeotropics? Historical biogeography and diversification of the cosmopolitan ectomycorrhizal mushroom family Inocybaceae. J. Biogeogr. 2009, 36, 577–592. [Google Scholar] [CrossRef]
- Kennedy, P.G.; Matheny, P.B.; Ryberg, K.M.; Henkel, T.W.; Uehling, J.K.; Smith, M.E. Scaling up: Examining the macroecology of ectomycorrhizal fungi. Mol. Ecol. 2012, 21, 4151–4154. [Google Scholar] [CrossRef]
- Han, L.H.; Feng, B.; Wu, G.; Halling, R.E.; Buyck, B.; Yorou, N.S.; Ebika, S.T.N.; Yang, Z.L. African origin and global distribution patterns: Evidence inferred from phylogenetic and biogeographical analyses of ectomycorrhizal fungal genus Strobilomyces. J. Biogeogr. 2018, 45, 201–212. [Google Scholar] [CrossRef]
- Geml, J.; Timling, I.; Robinson, C.H.; Lennon, N.; Nusbaum, H.C.; Brochmann, C.; Noordeloos, M.E.; Taylor, D.L. An arctic community of symbiotic fungi assembled by long-distance dispersers: Phylogenetic diversity of ectomycorrhizal basidiomycetes in Svalbard based on soil and sporocarp DNA. J. Biogeogr. 2012, 39, 74–88. [Google Scholar] [CrossRef]
- Gao, C.; Zhang, Y.; Shi, N.-N.; Zheng, Y.; Chen, L.; Wubet, T.; Bruelheide, H.; Both, S.; Buscot, F.; Ding, Q.; et al. Community assembly of ectomycorrhizal fungi along a subtropical secondary forest succession. New Phytol. 2015, 205, 771–785. [Google Scholar] [CrossRef]
- Castano, C.; Alday, J.G.; Parlade, J.; Pera, J.; de Aragon, J.M.; Bonet, J.A. Seasonal dynamics of the ectomycorrhizal fungus Lactarius vinosus are altered by changes in soil moisture and temperature. Soil Biol. Biochem. 2017, 115, 253–260. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Konopka, A.E.; Fredrickson, J.K. Stochastic and deterministic assembly processes in subsurface microbial communities. ISME J. 2012, 6, 1653–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.Z.; Wang, J.T. Mechanisms of community organization and spatiotemporal patterns of soil microbial communities. Acta Ecol. Sin. 2015, 35, 6575–6583. [Google Scholar] [CrossRef]
- Fransson, P. Elevated CO2 impacts ectomycorrhiza-mediated forest soil carbon flow: Fungal biomass production, respiration and exudation. Fungal Ecol. 2012, 5, 85–98. [Google Scholar] [CrossRef]
- Lloyd, J.; Farquhar, G.D. The CO2 Dependence of Photosynthesis, Plant Growth Responses to Elevated Atmospheric CO2 Concentrations and Their Interaction with Soil Nutrient Status. I. General Principles and Forest Ecosystems. Funct. Ecol. 1996, 10, 4–32. [Google Scholar] [CrossRef]
- Nasholm, T.; Hogberg, P.; Franklin, O.; Metcalfe, D.; Keel, S.G.; Campbell, C.; Hurry, V.; Linder, S.; Hogberg, M.N. Are ectomycorrhizal fungi alleviating or aggravating nitrogen limitation of tree growth in boreal forests? New Phytol. 2013, 198, 214–221. [Google Scholar] [CrossRef]
- Miyamoto, Y.; Terashima, Y.; Nara, K. Temperature niche position and breadth of ectomycorrhizal fungi: Reduced diversity under warming predicted by a nested community structure. Glob. Chang. Biol. 2018, 24, 5724–5737. [Google Scholar] [CrossRef]
- Morgado, L.N.; Semenova, T.A.; Welker, J.M.; Walker, M.D.; Smets, E.; Geml, J. Summer temperature increase has distinct effects on the ectomycorrhizal fungal communities of moist tussock and dry tundra in Arctic Alaska. Glob. Chang. Biol. 2015, 21, 959–972. [Google Scholar] [CrossRef] [Green Version]
- Geml, J.; Morgado, L.N.; Semenova-Nelsen, T.A.; Schilthuizen, M. Changes in richness and community composition of ectomycorrhizal fungi among altitudinal vegetation types on Mount Kinabalu in Borneo. New Phytol. 2017, 215, 454–468. [Google Scholar] [CrossRef] [Green Version]
- Lilleskov, E.A.; Fahey, T.J.; Horton, T.R.; Lovett, G.M. Belowground ectomycorrhizal fungal community change over a nitrogen deposition gradient in Alaska. Ecology 2002, 83, 104–115. [Google Scholar] [CrossRef]
- Corrales, A.; Turner, B.L.; Tedersoo, L.; Anslan, S.; Dalling, J.W. Nitrogen addition alters ectomycorrhizal fungal communities and soil enzyme activities in a tropical montane forest. Fungal Ecol. 2017, 27, 14–23. [Google Scholar] [CrossRef]
- Lilleskov, E.A.; Hobbie, E.A.; Horton, T.R. Conservation of ectomycorrhizal fungi: Exploring the linkages between functional and taxonomic responses to anthropogenic N deposition. Fungal Ecol. 2011, 4, 174–183. [Google Scholar] [CrossRef]
- Stenström, E. The effects of flooding on the formation of ectomycorrhizae in Pinus sylvestris seedlings. Plant Soil 1991, 131, 247–250. [Google Scholar] [CrossRef]
- Pigott, C.D. Survival of mycorrhiza formed by Cenococcum geophilum FR. in dry soils. New Phytol. 1982, 92, 513–517. [Google Scholar] [CrossRef]
- Reis, F.; Valdiviesso, T.; Varela, C.; Tavares, R.M.; Baptista, P.; Lino-Neto, T. Ectomycorrhizal fungal diversity and community structure associated with cork oak in different landscapes. Mycorrhiza 2018, 28, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, C.W.; Koide, R.T. The function of melanin in the ectomycorrhizal fungus Cenococcum geophilum under water stress. Fungal Ecol. 2013, 6, 479–486. [Google Scholar] [CrossRef]
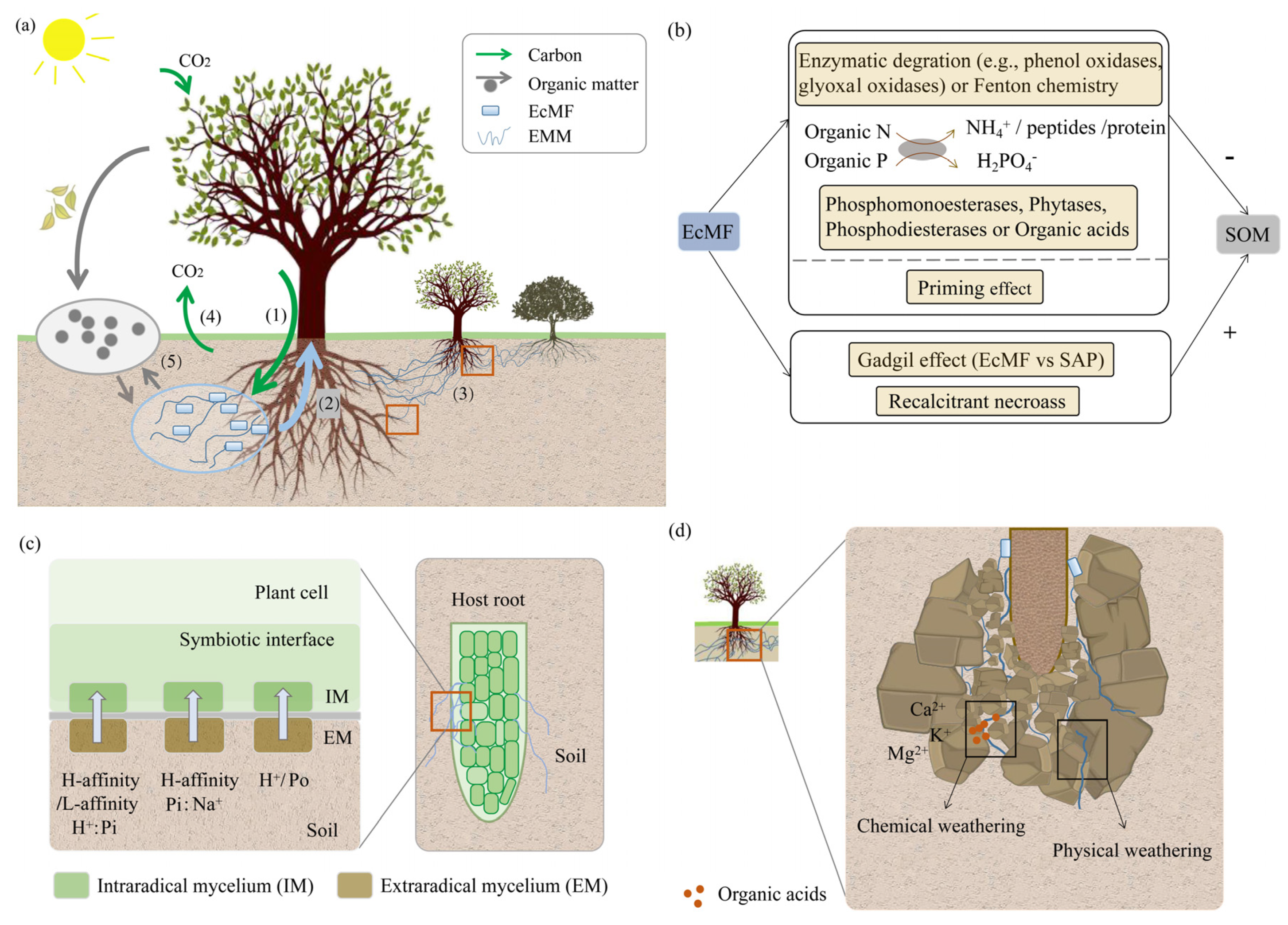
Figure 1.
The role of ectomycorrhizal fungi (EcMF) in nutrient cycling: (a) Carbon (C) flow through EcMF promotes the turnover of soil organic matter (SOM) and cycling of nutrients [29]. (1) Plant hosts transfer photosynthate to EcMF primarily as simple sugars [5]. (2) EcMF increase the effective absorptive surface area of roots enabling the transfer of nutrients, water, and other soil resources to hosts. (3) Hyphae can form inter- and intraspecific host mycelial networks for the bi-directional exchange of nutrients and carbohydrates [30,31]. (4) Root–mycorrhizal systems are involved in the release of carbon dioxide (CO2) by soil respiration [32]. (5) The turnover of EcMF and fine roots are important sources of labile C and nutrients for microbial processes [29], and the residues of EcMF and the extramatrical mycelium are incorporated into organic matter [33]. (b) EcMF participate in the decay of organic matter through enzymatic degradation and Fenton chemistry [29], the priming effect mediated by root and EcMF exudates [34] and the Gadgil effect caused by reduced saprotrophs activity based on the competitive interactions between mycorrhizal fungi and saprotrophs [35]. (c) EcMF transport phosphorus (P) from the soil to the symbiotic interface through phosphate transporters [36]. (d) EcMF accelerate weathering of minerals and rocks through physicochemical action [37]. EMM: Ectomycorrhizal mycelium. SAP: Saprotroph. IM: Intraradical mycelium. EM: Extraradical mycelium. N: Nitrogen. Pi: Inorganic phosphate. Po: Organic phosphate. H-affinity H+:Pi: High-affinity H+:Pi transporters. L-affinity H+:Pi: Low-affinity H+:Pi transporters. H-affinity Pi:Na+: High-affinity Pi:Na+ transporters. H+/Po: organic phosphate transporters. Fenton chemistry: This is a nonenzymatic reaction in which hydrogen peroxide (H2O2) oxidizes Fe2+ to form hydroxyl radicals [38]. Priming effect: Mycorrhizal fungi and fine roots exude simple organic compounds to increase microbial activity [29]. Gadgil effect: Competition between mycorrhizal fungi and saprotrophs may decrease the decomposition rate of SOM [35]. Necromass: Organic matter derived from or composed of dead microorganisms [29].
Figure 1.
The role of ectomycorrhizal fungi (EcMF) in nutrient cycling: (a) Carbon (C) flow through EcMF promotes the turnover of soil organic matter (SOM) and cycling of nutrients [29]. (1) Plant hosts transfer photosynthate to EcMF primarily as simple sugars [5]. (2) EcMF increase the effective absorptive surface area of roots enabling the transfer of nutrients, water, and other soil resources to hosts. (3) Hyphae can form inter- and intraspecific host mycelial networks for the bi-directional exchange of nutrients and carbohydrates [30,31]. (4) Root–mycorrhizal systems are involved in the release of carbon dioxide (CO2) by soil respiration [32]. (5) The turnover of EcMF and fine roots are important sources of labile C and nutrients for microbial processes [29], and the residues of EcMF and the extramatrical mycelium are incorporated into organic matter [33]. (b) EcMF participate in the decay of organic matter through enzymatic degradation and Fenton chemistry [29], the priming effect mediated by root and EcMF exudates [34] and the Gadgil effect caused by reduced saprotrophs activity based on the competitive interactions between mycorrhizal fungi and saprotrophs [35]. (c) EcMF transport phosphorus (P) from the soil to the symbiotic interface through phosphate transporters [36]. (d) EcMF accelerate weathering of minerals and rocks through physicochemical action [37]. EMM: Ectomycorrhizal mycelium. SAP: Saprotroph. IM: Intraradical mycelium. EM: Extraradical mycelium. N: Nitrogen. Pi: Inorganic phosphate. Po: Organic phosphate. H-affinity H+:Pi: High-affinity H+:Pi transporters. L-affinity H+:Pi: Low-affinity H+:Pi transporters. H-affinity Pi:Na+: High-affinity Pi:Na+ transporters. H+/Po: organic phosphate transporters. Fenton chemistry: This is a nonenzymatic reaction in which hydrogen peroxide (H2O2) oxidizes Fe2+ to form hydroxyl radicals [38]. Priming effect: Mycorrhizal fungi and fine roots exude simple organic compounds to increase microbial activity [29]. Gadgil effect: Competition between mycorrhizal fungi and saprotrophs may decrease the decomposition rate of SOM [35]. Necromass: Organic matter derived from or composed of dead microorganisms [29].


{kind=link}
Table 1.
Degradation of soil organic matter (SOM) via extracellular enzymes produced by ectomycorrhizal fungi [29,33].
| SOM Sources | SOM Fractions | Enzymes | Enzymatic Attack |
|---|---|---|---|
| Plant material | Cellulose Hemicellulose | Cellulases | Hydrolytic |
| Hemicellulases | |||
| Lignin | Peroxidases, Phenol oxidases | Oxidative | |
| Microbial necromass | Chitin | Chitinases | Hydrolytic |
| Melanin | Peroxidases, Phenol oxidases | Oxidative |
Table 2.
Affiliation of host plant to families at different geographic regions (adopted from [4]).
Table 2.
Affiliation of host plant to families at different geographic regions (adopted from [4]).
| Plant Family | Asia | Africa | Oceania | Neotropical |
|---|---|---|---|---|
| Achatocarpaceae | - | - | - | 1.67% |
| Asteropeiaceae | - | 0.83% | - | - |
| Betulaceae | - | - | - | 1.67% |
| Cistaceae | - | - | - | 1.67% |
| Dipterocarpaceae | 83.05% | 9.17% | - | 1.67% |
| Fabaceae | 1.69% | 58.33% | 22.73% | 11.67% |
| Fagaceae | 5.08% | - | - | 18.33% |
| Gnetaceae | - | 2.50% | 2.27% | 3.33% |
| Goodeniaceae | - | - | 2.27% | - |
| Juglandaceae | 1.69% | - | - | 3.33% |
| Myrtaceae | 5.08% | - | 63.64% | - |
| Nothofagaceae | - | - | 6.82% | - |
| Nyctaginaceae | - | 1.67% | 2.27% | 23.33% |
| Phyllanthaceae | - | 16.67% | - | - |
| Pinaceae | 3.39% | - | - | 18.33% |
| Polygonaceae | - | - | - | 13.33% |
| Salicaceae | - | - | - | 1.67% |
| Sarcolaenaceae | - | 10.83% | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, Y.; Li, X.; Kou, Y. Ectomycorrhizal Fungi: Participation in Nutrient Turnover and Community Assembly Pattern in Forest Ecosystems. Forests 2020, 11, 453. https://doi.org/10.3390/f11040453
AMA Style
Liu Y, Li X, Kou Y. Ectomycorrhizal Fungi: Participation in Nutrient Turnover and Community Assembly Pattern in Forest Ecosystems. Forests. 2020; 11(4):453. https://doi.org/10.3390/f11040453
Chicago/Turabian StyleLiu, Yanjiao, Xiangzhen Li, and Yongping Kou. 2020. "Ectomycorrhizal Fungi: Participation in Nutrient Turnover and Community Assembly Pattern in Forest Ecosystems" Forests 11, no. 4: 453. https://doi.org/10.3390/f11040453
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.