1. Introduction
A series of insects and pathogens have pulsed through the Eastern Deciduous Forest in the United States, causing widespread tree mortality. For example, the fungal pathogen
Cryphonectria parasitica (Murrill) Barr swept through the Appalachian Mountains, devastating American chestnut (
Castanea dentata (Marshall) Borkh.) and initiating what has become a flagship restoration effort [
1,
2,
3,
4,
5]. A series of mortality agents have had similar effects, including the butternut canker, dogwood anthracnose, gypsy moth and hemlock wooly adelgid, among many others [
6,
7,
8,
9]. The loss of canopy trees has the potential to interact with ecological invasion by exotic plants. Widespread tree mortality caused by invasive organisms has the potential to facilitate invasional meltdown [
10] by creating gaps in the forest canopy, which reduce interspecific competition and increase sunlight availability for invasive plants [
11]. Given the ever-increasing pace of global commerce and associated dispersal of species across historically impassable boundaries and distances, the likelihood of future pest and pathogen epidemics is only increasing. The ecology of temperate deciduous forests is shaped by multiple interacting disturbance factors [
12], including pests and pathogens [
13], and broad-scale tree mortality events are likely to be a fundamental challenge for ecological management, and impetus for restoration, in the 21st century and beyond.
The loss of overstory ash (
Fraxinus spp.) due to the emerald ash borer (EAB) in forests where exotic species are present has created the need for reforestation in many areas of the American Midwest. Originally from Asia, EAB is a highly effective mortality agent and has spread rapidly [
14,
15,
16]. Infestation of EAB within a particular stand of trees results in near total mortality of
Fraxinus and the consequent loss of forest canopy trees [
17]. Indeed, where
Fraxinus trees were removed from forest stands, 18.7% of the herbaceous cover was an invasive plant, whereas, where
Fraxinus trees were left intact, there was less than 1% coverage by invasive species [
18]. After
Fraxinus trees die and eventually fall to the ground, canopy gaps will be inevitable and will likely result in higher invasive coverage in infested forests. In southwestern Ohio, EAB has created mortality in forests where invasive plants are a significant concern. For example,
Lonicera maackii (Rupr.) Herder is a significant invasive species [
19] that co-occurs in most forested ecosystems where EAB has created canopy gaps. In southwestern Ohio, the Five Rivers MetroParks (FRMP) own and manage
ca. 6500 hectares of land, and their conservation group has developed a community-engaged reforestation program that seeks to address the loss of canopy trees and impede ecological invasion. As in much of the Midwest, many of the FRMP conservation areas are old farms that were left to regenerate, and several had overstories dominated by
Fraxinus species with understories where invasive plants had established. The overall objective was to plant a diverse woody understory in areas where
Fraxinus has been lost before invasive species had the opportunity for population expansion into the open niche spaces.
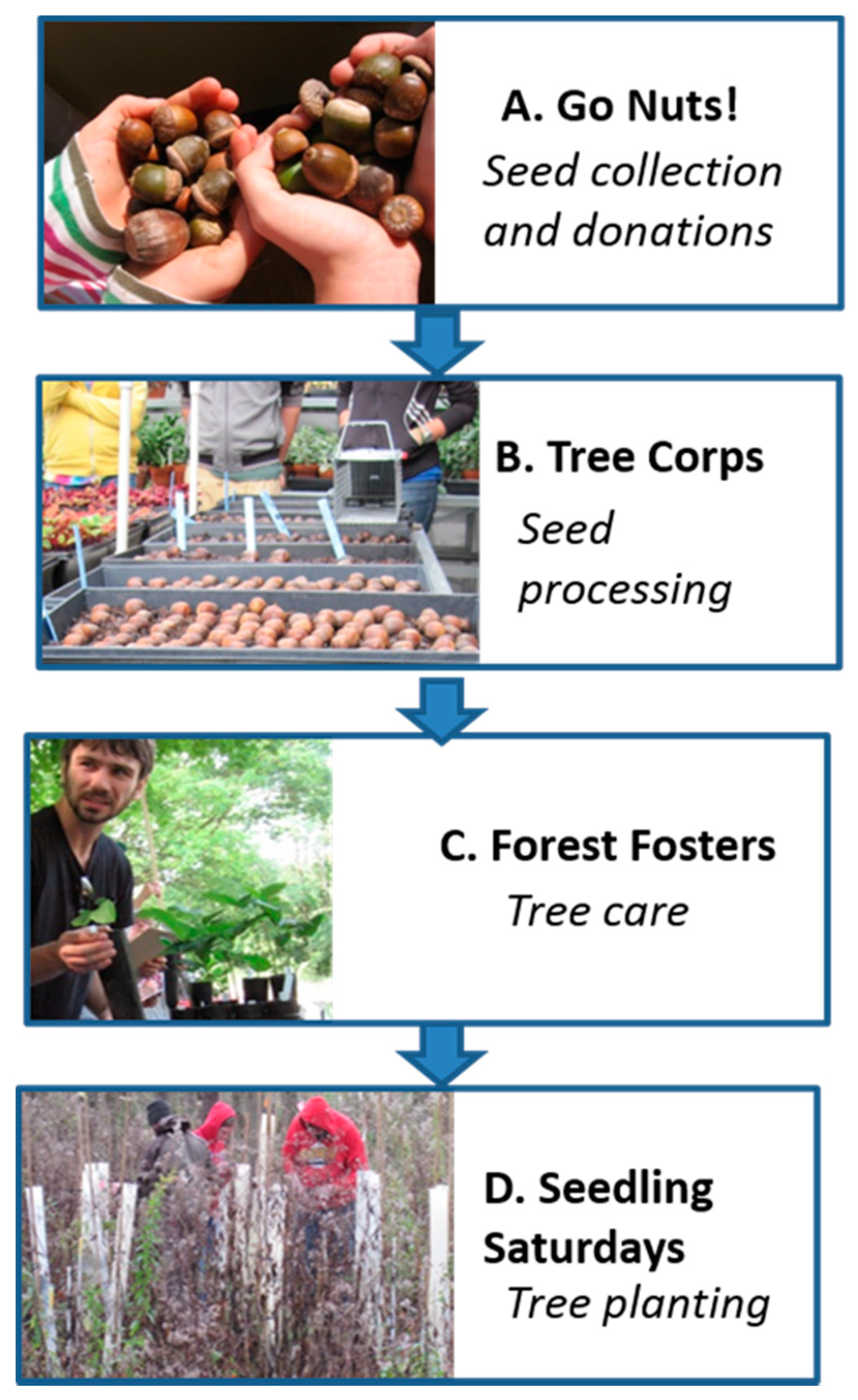
The FRMP developed a reforestation program with the potential to become a regional model for community-engaged reforestation (
Figure 1). In many of their properties, the initial phase of work is removal of an extensive shrub understory dominated by invasive species including
L. maackii. When the site preparation is complete, the next phase is developing a seedling pool for planting. This process starts with the “Go Nuts!” program, wherein citizens were encouraged to collect tree seeds and donate them to FRMP. Training materials were provided for the general public through FRMP publications, and the program has generally resulted in large quantities of seeds being donated. The next step is the “Tree Corps” program, wherein FRMP volunteers were trained in seed processing and stratification. The next phase in the process is the “Forest Fosters” program, in which individual families from the general public were encouraged to volunteer to take a flat of tree seedlings home with them over the summer and provide care for the seedlings. “Foster families” were provided training and were generally excited to take care of the seedlings, resulting in very low mortality. Finally, the program culminates in “Seedling Saturdays”, wherein volunteers were provided training in tree planting in areas where
Fraxinus has been lost. Volunteer tree planting groups were highly motivated, and, with minimal training and support, were effective in tree planting. This process was implemented in 2011 and is still being used in 2020 for many forest restoration projects. This process encourages community engagement at each step, from seed donations to seed processing, taking care of trees and finally planting them in target restoration sites alongside FRMP staff.
The “Go Nuts!” portion of the FRMP program attracts a wide variety of seeds from native species, as the interested public collects seeds on hikes, in cemeteries and back yards, and from street trees. In this experiment,
Quercus rubra L. (red oak),
Carya laciniosa (Michx. f.) G. Don (shell bark hickory) and
Juglans cinerea L. (butternut) were chosen as study species based on FRMP management objectives. Oak restoration is important because of ongoing, long-term dynamics across a large area of eastern North America [
12], and hickories are often found intermingled with oaks in these systems.
Quercus rubra and
C. laciniosa have wildlife value and are common in overstory forests of the region and, therefore, hold good potential as restoration target species for the future forest canopy.
Juglans cinerea was chosen because it loosely approximates the ecology of the common, closely related,
J. nigra, and it is interesting from a restoration standpoint because of the loss of this species due to butternut canker [
7] All of these species have been cultivated from seeds for many decades, creating a wealth of knowledge on which seed characteristics lead to greatest germination and seedling success.
Carya species, with larger nuts, tend to have larger seedlings [
20], but seed size has not been linked to germination rates. The USDA Woody Plant Seed Manual recommendations do not list specific seed sizes nor seed stratification media with any of the selected species [
21]. The main considerations when selecting a stratification medium are finding an optimal amount of moisture retention, such that the seeds will not desiccate or rot but will remain viable after being stratified over winter. Thus, vermiculite is an appealing media due to its high water retention, whereas sand may provide a more aerated environment for the seeds, preventing them from becoming too moist. In an effort to streamline volunteer efforts, FRMP aimed to identify if there is a seed size screening that would promote overall germination and to determine if stratification media plays a role in facilitating germination.
Our goal in this experiment was to leverage ongoing efforts (
Figure 1) to answer practical questions related to tree seed handling using techniques that approximate those of large-scale, low-technology restoration projects. We tested the efficacy of sand or vermiculite storage media against storage with no media in this experiment and asked: (
Q1) does stratification medium influence total germination or time to germination? Seeds collected from the field vary greatly in size, and another question addressed in this experiment was how size related to germination success. If seed size was a strong determinate on seedling germination and survival, then seed processing could include a size-based selection process to eliminate those seeds unlikely to create seedlings, thereby saving space and time during the project. Therefore, we asked: (
Q2) does seed size influence overall germination or time to germination? This project was focused on answering practical questions relevant to ongoing activities and was directly integrated into restoration practice.
2. Materials and Methods
Seeds for this study were collected by volunteers within 100 miles of Dayton, OH, USA during the months of August, September and October 2011 through the “Go Nuts!” program (
Figure 1). In total, the study began with 1376 seeds, 468
C. laciniosa, 584
Q. rubra and 324
J. cinerea. Due to handling loss and the obvious emergence of insect larvae, and based on screening efforts described below, 88 seeds were lost before the project began (65
C. laciniosa, 14
Q. rubra, and 9
J. cinerea), leaving a total of 1288 seeds in the study.
Seeds were batched together into species-specific plastic bags and cleaned of husks, caps and shells. Seeds were then placed into a bucket of water for 24 hours to test for soundness. Those which floated were discarded, and the rest were laid out on screens to surface dry. Once dry, any seeds that showed insect holes, splits, or cracks were discarded. Each seed’s length and width were measured, and each seed was assigned an individual number and placed into an individual plastic bag. Seeds were randomly assigned stratification media treatments within their individual bags: moist vermiculite, moist sand, or no media. Seeds were then stratified in the dark at 1.1 °C degrees for 3 months.
Seeds were removed from refrigeration on 6 March 2012. Each seed was checked for germination. If a seed had germinated, it was planted in a cone-shaped container (6.4 cm diameter, depth of 25.4 cm). If it had not germinated, it was placed in a flat of soil, tagged and covered with about 2.5 cm of soil. In total, 319
Q. rubra, 5
J. cinerea and no
C. laciniosa had germinated prior to removal from refrigeration. The flats were placed in a warm greenhouse (ca. 21–26 °C) with natural light and watered once a day. Seeds were uncovered and checked for germination once a week for 7 weeks. Upon germination, the seeds were planted into the cone-shaped containers. Planted seeds were placed in a cooler greenhouse (ca. 7 °C) under automatic irrigation with a dilute fertilizer solution of (10-4-3: NPK) 4 days a week. Flats were placed so that each stem had equal competition and were sorted occasionally for removal of dead stems. Flats were moved outside in May 2012 in preparation for “Seeding Saturdays” activity (
Figure 1).
All data are publicly available [
22]. Statistical analyses were conducted in R version 3.6.2 [
23]. We applied logistic regressions to determine the impact of stratification media on germination rates of seeds from each of the three species using the function
glm. Further, we used linear models to assess the time to seed germination as influenced by germination medium and seed size for each species using the
lm function, using one-way ANOVA and linear regression respectively. We also assessed the effect of seed size on germination success using one-way ANOVA.
3. Results
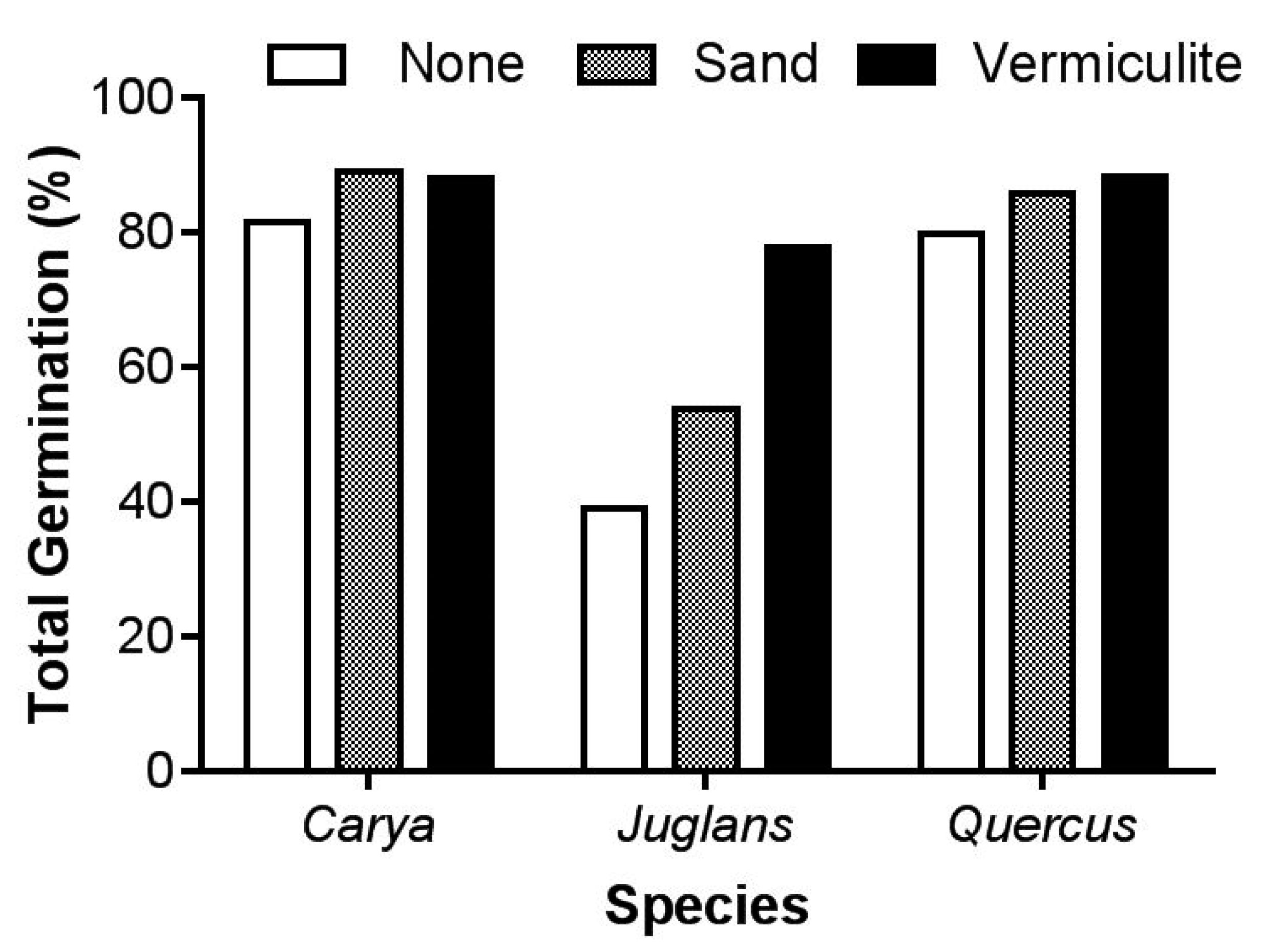
The germination response of seeds to storage substrate was species-specific (
Figure 2). Germination varied by stratification media for
J. cinerea, such that seeds stratified in vermiculite had the highest amount of germination (77.7%), followed by sand (53.7%) and with the lowest germination when stratified without media (38.9%; z
3,315 = −2.53,
P = 0.011). Germination ranged between 80 and 90% for
C. laciniosa, and there was no effect of substrate on total germination (P > 0.05). For
Q. rubra, germination ranged between 79.6% for no substrate to 88.3% for vermiculite, with no statistically significant differences between substrate types (
P > 0.05).
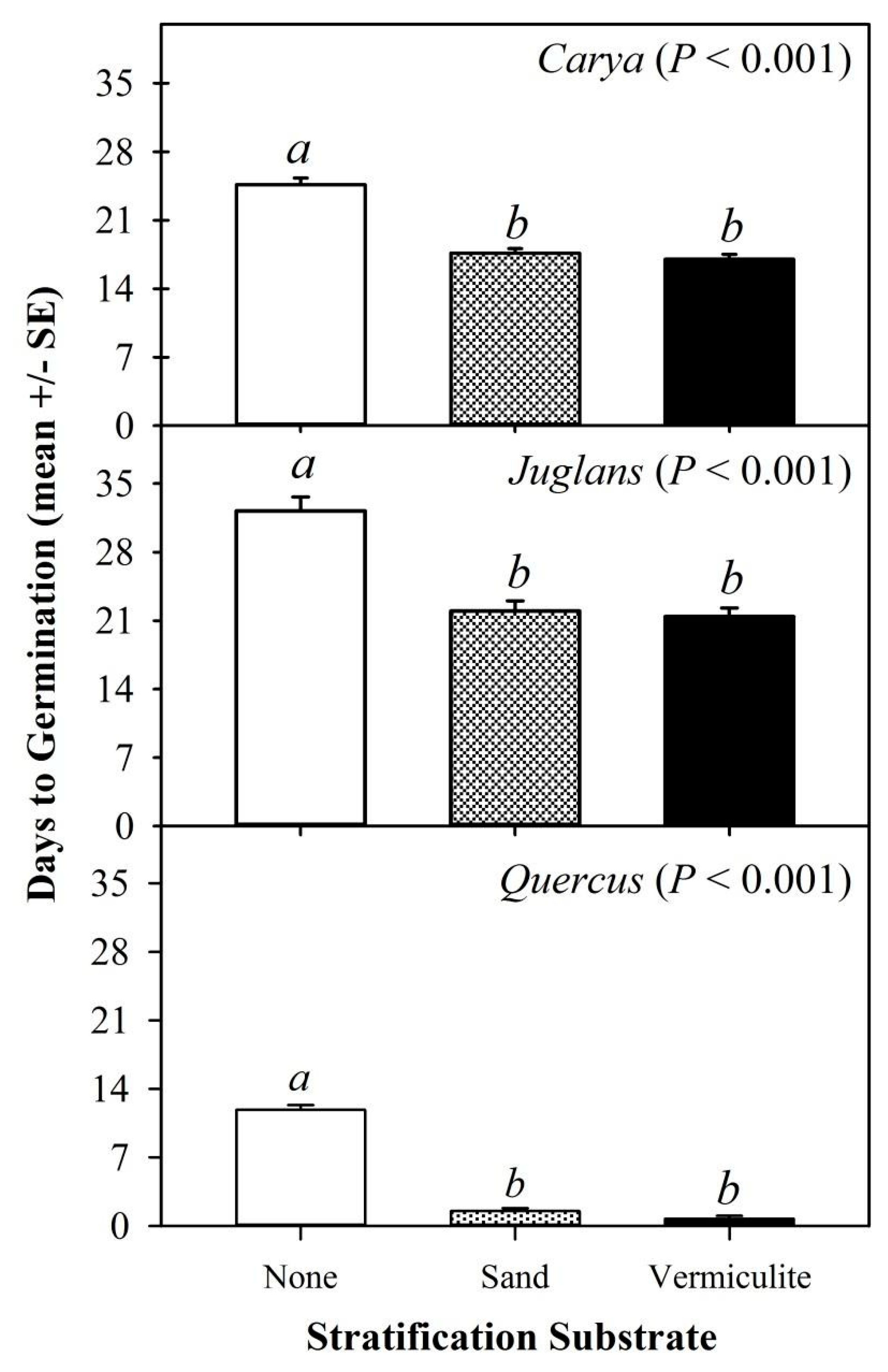
Germination timing, which we are defining as the number of days to germination after the seeds were removed from stratification conditions in the refrigerator, exhibited a similar response to storage media across species (
Figure 3). For each species, there was an overall treatment effect such that seeds stratified without substrate exhibited a longer time to germination than those which were stratified with either vermiculite or sand (F
8,1002 = 344.5,
P < 0.001).
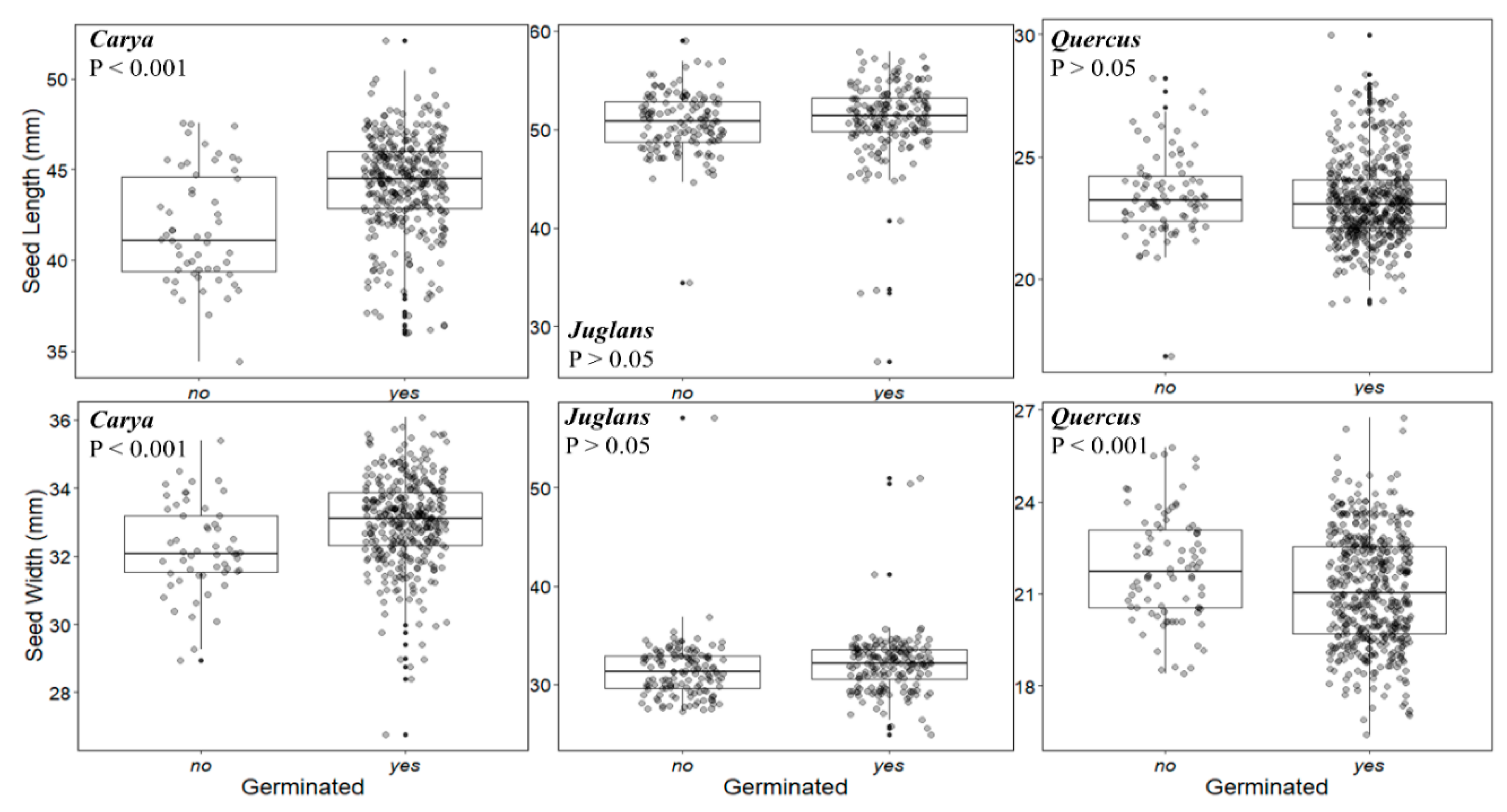
The effect of seed height and width on germination success was species-specific (
Figure 4). For
C. laciniosa, there was a statistically detectable difference between seed size for seeds that died and those that germinated, such that longer seeds (F
1,401 = 40.08,
P < 0.001) and wider seeds (F
1,401 = 17.63,
P < 0.001) both had higher germination success.
Juglans cinerea size was not related to the size of the seed, either width or length (
P > 0.05). The seed length of
Q. rubra did not predict the germination rate (
P > 0.05), but the seeds with smaller widths were more likely to germinate than wider seeds (F
1,568 = 40.08,
P < 0.001).
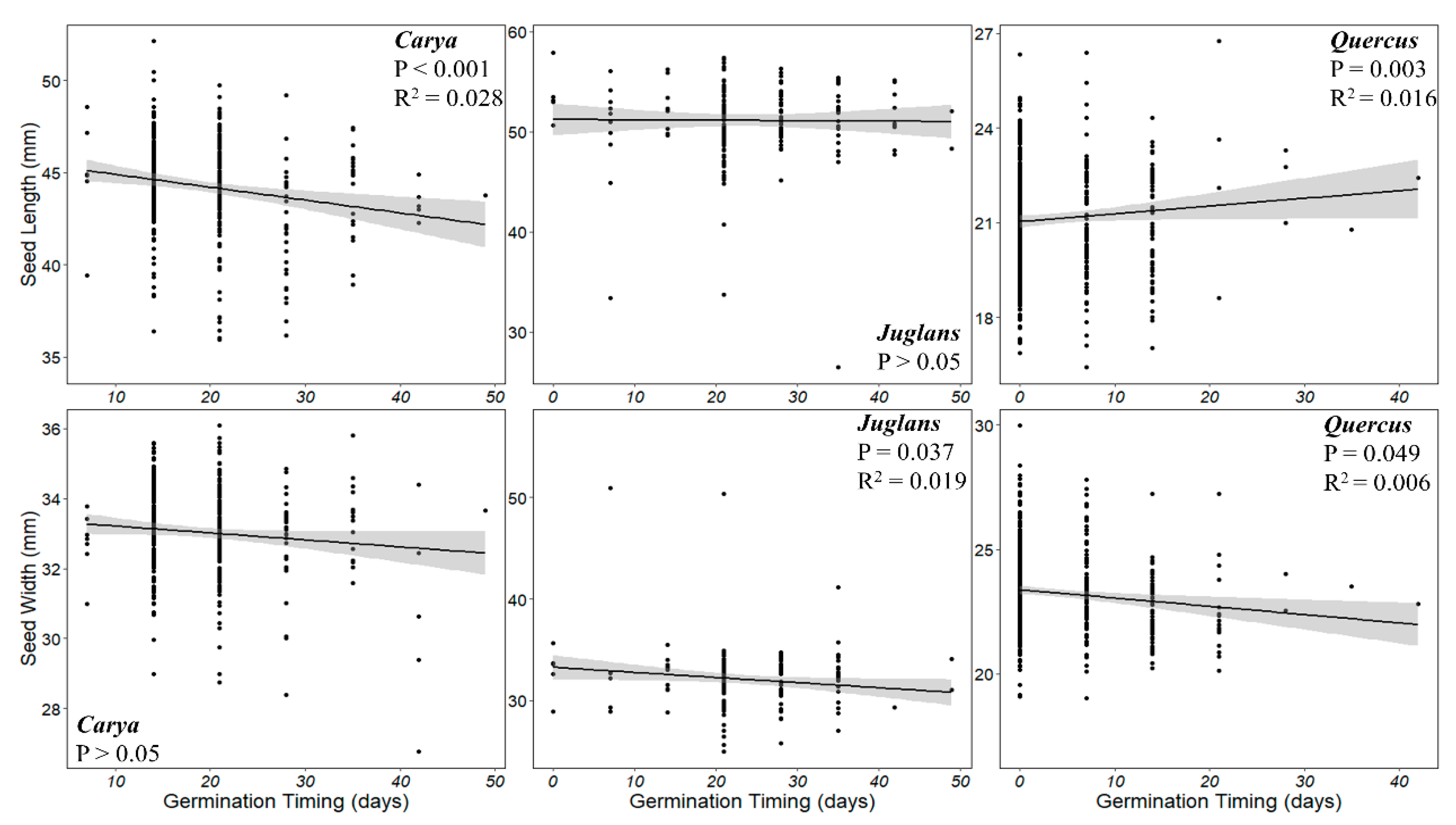
The relationship between germination timing and seed size was also species-specific (
Figure 5). Germination occurred earlier for long
C. laciniosa seeds (F
1,346 = 11.2,
P < 0.001), but timing was not dependent on the width of the seeds (
P > 0.05). Meanwhile, wider
J. cinerea seeds typically germinated faster than smaller seeds, but this trend is weak (F
1,177 = 4.376,
P = 0.037).
Juglans cinerea seed lengths did not predict the timing of germination (P > 0.05).
Quercus rubra seeds that were shorter germinated faster (F
1,482 = 8.63,
P = 0.003) and those which were wider germinated faster (F
1,482 = 3.886,
P = 0.049). Overall,
R2 values are low for statistically significant relationships, suggesting that seed size is not a strong determinate of germination timing.
4. Discussion
The response of seeds to germination media was species-specific. We found that
C. laciniosa and
Q. rubra germination success was not influenced by stratification media. Both
C. laciniosa and
Q. rubra germinated 1–2 weeks sooner when they were stratified with media, either sand or vermiculite, than when they were stratified without media. This suggests that seed handlers can stratify
C. laciniosa and
Q. rubra seeds without media without influencing overall germination success, but media may be useful to decrease time to germination. This may be due to the overall high germination success of these species, such that germination rates ranged between 80% and 90% for these species regardless of media.
Juglans cinerea germinated at a higher rate when stratified with vermiculite compared to sand or without media. Vermiculite provides a higher water content than either sand or a lack of media, and congeneric
J. regia had high germination rates when stratified in water alone [
24]. This suggests that
Juglans spp. may rely on high water content during stratification for increased germination success. Further,
J. cinerea germinated faster when stratified with either media type than without media. Thus,
J. cinerea seeds will have the highest success and fastest germination when stratified with a moist substrate, such as vermiculite.
The effect of seed size on germination success and timing was species-specific.
C. laciniosa germinated more often when seeds were both longer and wider, and longer seeds took less time to germinate. In contrast,
Q. rubra seeds germinated at a higher rate when seeds had a smaller width, but the length of seeds did not predict germination rate.
Quercus rubra seeds that had shorter length germinated faster than longer seeds, while higher seed width led to slower germination. Congeneric
Q. ilex germinated at a higher rate and faster when seeds were larger [
25], whereas
Q. robur did not vary in germination rate by seed size [
26], suggesting that findings may be species- or context-dependent. Finally,
J. cinerea germination success was not dependent on size, but germination timing was faster with wider seeds. Overall, these relationships are highly variable, and there is no clear threshold of seed size contributing to higher germination success or faster germination for any of the species. Implementing a seed size screening could lead to slightly higher germination success, especially for
C. laciniosa and
Q. rubra, but this would risk imposition of selection bias that could lead to genetic bottlenecks and lower genetic diversity of trees planted within restoration efforts.
In this study, we leveraged a community-engaged restoration project lead by local land managers in Southwestern Ohio, whose restoration practices have been successfully ongoing for 10 years (
Figure 1). We assessed seeds donated by the community in order to determine the efficiency and efficacy of FRMP’s cold stratification processes and seed-screening practices. We used the unconventional measurement of seed length and width to determine seed size. We used these parameters because they are easily measured by land managers and could be used to implement a quick screening of donated seeds. Because we found little indication that seed size, when measured by length or width, predicts germination success or timing, we advise that seeds are not screened by their sizes prior to germination. Such screening would induce bias and not be indicative of success. Further, seed origin and collection practices can alter germination success and timing [
27,
28], suggesting that community-engaged restoration projects may benefit from more strict collection policies when acquiring donations to ensure adequate genetic diversity and high success of restoration.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




