
MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia

 ,
,  ,
, 
Abstract
:1. Introduction
- Predictive distribution modelling of Pinus gerardiana under current climatic/environmental conditions.
- Forecasting of potential distribution variations under proposed future climate change scenarios;
- The identification of the most influential environmental factors, and;
- The identification of the possible impacts of future ditributional variations of the species on the associated local communities.
2. Materials and Methods
2.1. Chilgoza Pine and Study Area
2.2. Species Occurrence Data
2.3. Predictors Data
2.4. Variables Selection
2.5. Model Optimization, Calibration, Validation and Prediction Reclassification
3. Results
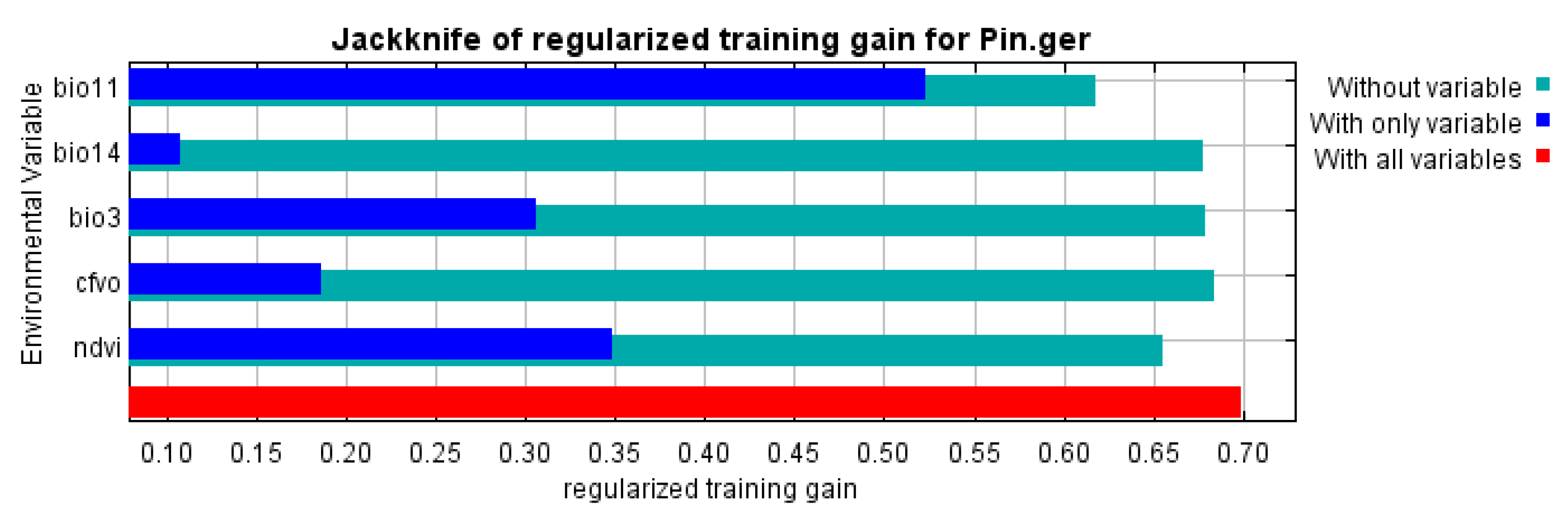
3.1. Model Performance and Variables Importance
3.2. Variables Response Curves
3.3. Present Distribution and Extent of Occurrence
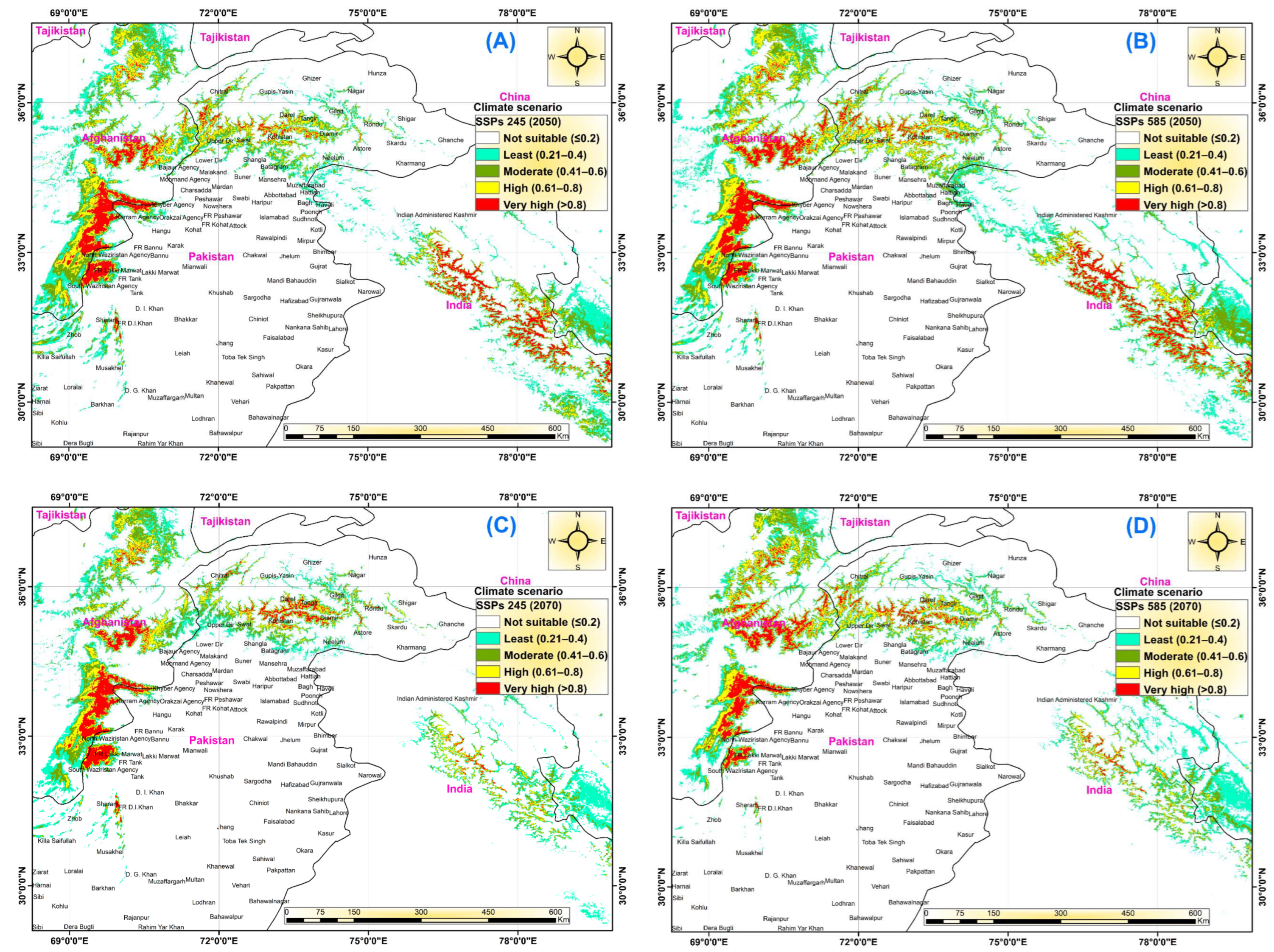
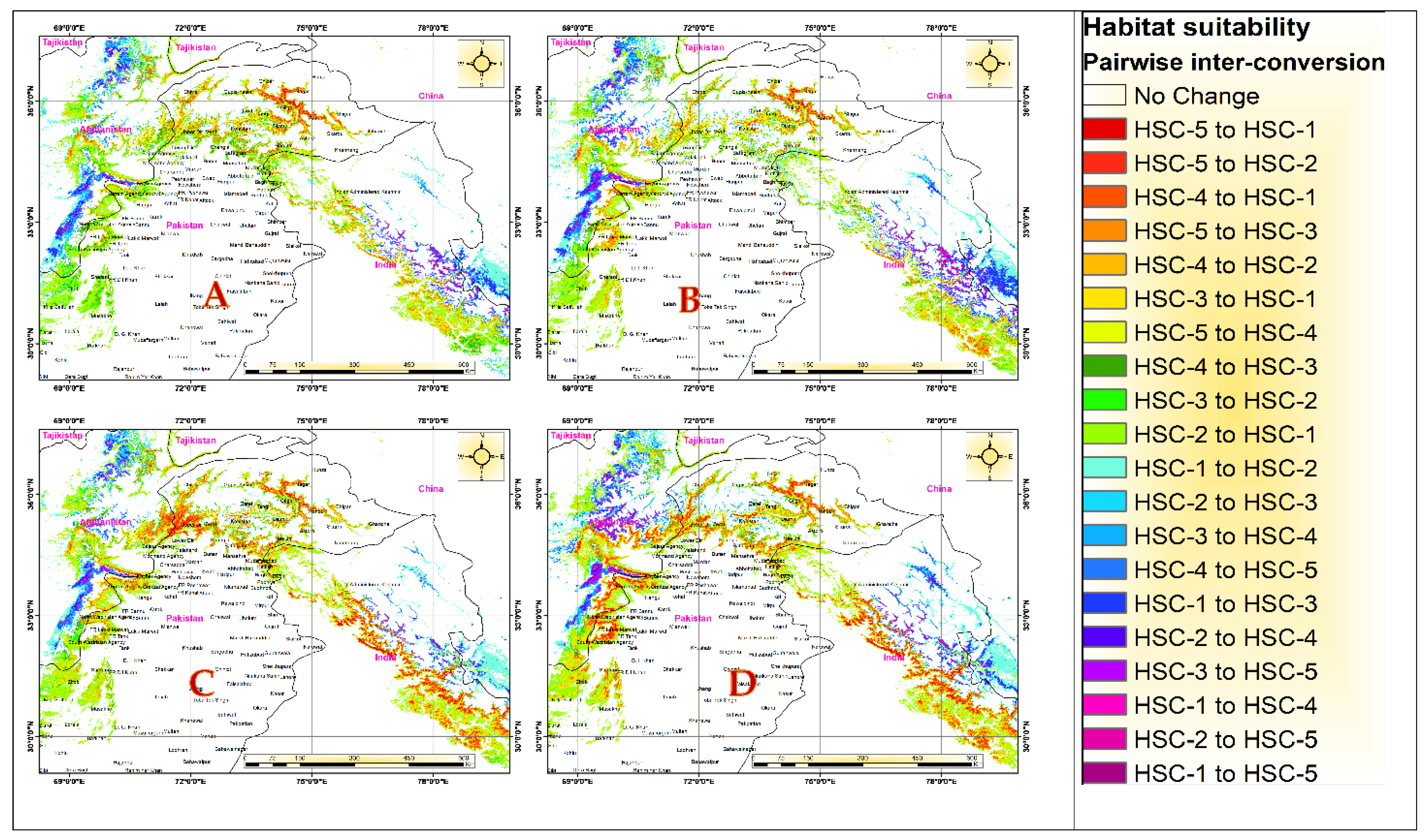
3.4. Future Predicted Distributions
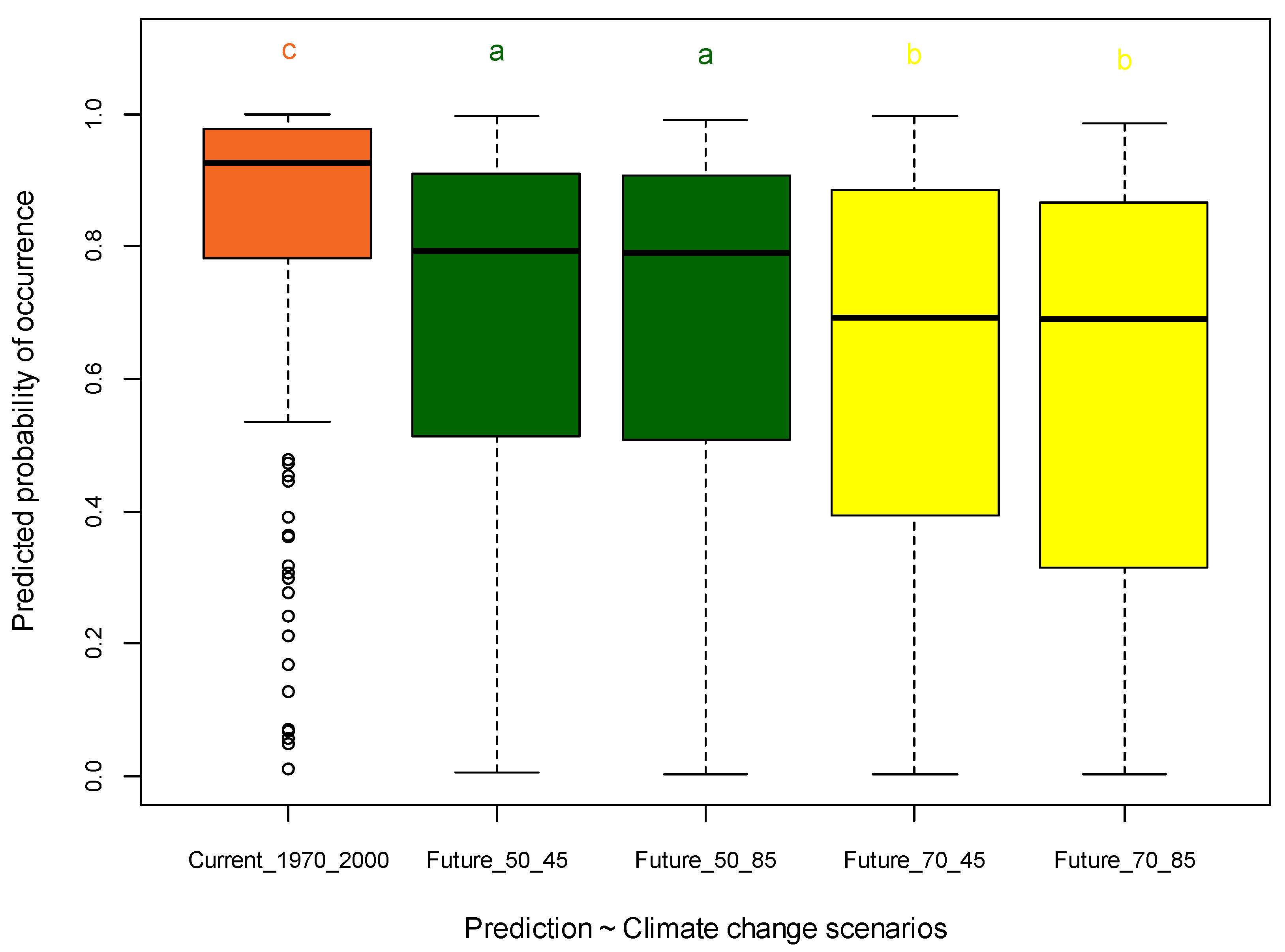
3.5. Significance Testing of MaxEnt Predictions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rebelo, H.; Jones, G. Ground validation of presence-only modelling with rare species: A case study on Barbastella barbastellus (Chiroptera: Vespertilionidae). J. Appl. Ecol. 2010, 47, 410–420. [Google Scholar] [CrossRef]
- Di Febbraro, M.; Martinoli, A.; Russo, D.; Preatoni, D.; Bertolino, S. Modelling the effects of climate change on the risk of invasion by alien squirrels. Hystrix 2016, 27, 1–8. [Google Scholar] [CrossRef]
- Bosso, L.; Rebelo, H.; Garonna, A.P.; Russo, D. Modelling geographic distribution and detecting conservation gaps in Italy for the threatened beetle Rosalia alpina. J. Nat. Conserv. 2013, 21, 72–80. [Google Scholar] [CrossRef]
- Busby, J.R. A biogeoclimatic analysis of Nothofagus cunninghamii (Hook.) Oerst. in southeastern Australia. Aust. J. Ecol. 1986, 11, 1–7. [Google Scholar] [CrossRef]
- Razgour, O.; Hanmer, J.; Jones, G. Using multi-scale modelling to predict habitat suitability for species of conservation concern: The grey long-eared bat as a case study. Biol. Conserv. 2011, 144, 2922–2930. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.D.; Roberts, D.; Reid, N. Applying species distribution modelling to identify areas of high conservation value for endangered species: A case study using Margaritifera margaritifera (L.). Biol. Conserv. 2011, 144, 821–829. [Google Scholar] [CrossRef]
- Elith, J.; Franklin, J. Species Distribution Modelling. Encyclopedia of Biodiversity; Levin, S., Ed.; Academic Press: Waltham, MA, USA, 2013; pp. 692–705. [Google Scholar]
- Farren, A.; Prodöhl, P.A.; Laming, P.; Reid, N. Distribution of the common lizard (Zootoca vivipara) and landscape favourability for the species in Northern Ireland. Amphibia-Reptilia 2010, 31, 387–394. [Google Scholar] [CrossRef] [Green Version]
- Olsson, O.; Rogers, D.J. Predicting the distribution of a suitable habitat for the white stork in Southern Sweden: Identifying priority areas for reintroduction and habitat restoration. Anim. Conserv. 2009, 12, 62–70. [Google Scholar] [CrossRef]
- Gibson, L.A.; Wilson, B.A.; Cahill, D.M.; Hill, J. Spatial prediction of rufous bristlebird habitat in a coastal heathland: A GIS-based approach. J. Appl. Ecol. 2004, 41, 213–223. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Brambilla, M.; Caprio, E.; Assandri, G.; Scridel, D.; Bassi, E.; Bionda, R.; Chamberlain, D. A spatially explicit definition of conservation priorities according to population resistance and resilience, species importance and level of threat in a changing climate. Divers. Distrib. 2017, 23, 727–738. [Google Scholar] [CrossRef]
- Pearson, R. Species Distribution Modeling for Conservation Educators and Practitioners. Synthesis. Am. Mus. Nat. Hist. 2007, 50, 54–89. [Google Scholar]
- Jaynes, E.T. Information theory and statistical mechanics. Phys. Rev. 1957, 106, 620–630. [Google Scholar] [CrossRef]
- Shannon, C.A. Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicating impact of climate change on medicinal asclepiads of Pakistan using Maxent modeling. Acta Oecologica 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Papes, M.G. Modelling ecological niches from low numbers of occurrences: Assessment of the conservation status of poorly known viverrids (Mammalia, Carnivora) across two continents. Divers. Distrib. 2007, 13, 890–902. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modelling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of Maxent for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Fuller, D.O.; Ahumada, M.L.; Quiñones, M.L.; Herrera, S.; Beier, J.C. Nearpresent and future distribution of Anopheles albimanus in Mesoamerica and the Caribbean Basin modeled with climate and topographic data. Int. J. Health Geogr. 2012, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, J.N.; Loyola, R.D. Applying niche-based models to predict endangered-hylid potential distributions: Are neotropical protected areas effective enough? Trop. Sci. 2008, 1, 417–445. [Google Scholar] [CrossRef]
- Chen, G.J.; Peterson, A.T. A new technique for predicting distribution of terrestrial vertebrates using inferential modelling. Zool. Res. 2000, 21, 231–237. [Google Scholar]
- Morrison, M.L.; Hall, L.S. Standard terminology: Toward a common language to advance ecological understanding and application. In Predicting Species Occurrences: Issues of Accuracy and Scale; Scott, J.M., Heglund, P.J., Morrison, M.L., Haufler, J.B., Raphael, M.G., Wall, W.A., Samson, F.B., Eds.; Island Press: Washington, DC, USA, 2002; pp. 43–52. [Google Scholar]
- Negga, H. Predictive Modelling of Amphibian Distribution Using Ecological Survey Data: A Case Study of Central Portuga. Master’s Thesis, International Institute for Geo-Information Science and Earth Observation, Enschede, The Netherlands, 2007. [Google Scholar]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Ellis, C.J. Ancient woodland indicators signal the climate change risk for dispersal-limited species. Ecol. Indic. 2015, 53, 106–114. [Google Scholar] [CrossRef]
- Khan, A.M.; Qureshi, R.; Qaseem, M.F.; Munir, M.; Ilyas, M.; Saqib, Z. Floristic checklist of district Kotli, Azad Jammu & Kashmir. Pak. J. Bot. 2015, 47, 1957–1968. [Google Scholar]
- Khan, A.M.; Khalid, N. Climate Change is Driving Greening Trend Whereas Tree Cover is Declining in The Western Himalaya of Pakistan. J. Bioresour. Manag. 2021, 8, 1202.0200. [Google Scholar] [CrossRef]
- Majeed, M.; Tariq, A.; Anwar, M.M.; Khan, A.M.; Arshad, F.; Mumtaz, F.; Shaukat, S. Monitoring of land use–land cover change and potential causal factors of climate change in Jhelum district, Punjab, Pakistan, through GIS and multi-temporal satellite data. Land 2021, 10, 1026. [Google Scholar] [CrossRef]
- Khan, A.M.; Qureshi, R.; Arshad, M.; Mirza, S.N. Climatic and flowering phenological relationships of western Himalayan flora of Muzaffarabad district, Azad Jammu and Kashmir, Pakistan. Pak. J. Bot. 2018, 50, 1093–1112. [Google Scholar]
- Slee, B.; Feliciano, D. Challenges in the design of indicators for assessing the impact of the Scotland Rural Development Programme 2007–2013 on climate change mitigation. Ecol. Indic. 2015, 59, 94–103. [Google Scholar] [CrossRef]
- Ali, F.; Khan, N.; Ali, K.; Khan, I. Influence of environmental variables on the distribution of woody species in Muslim graveyards of Malakand Division, Hindukush Range Mountains of Pakistan. Pak. J. Bot. 2017, 49, 2357–2366. [Google Scholar]
- Critchfield, W.B.; Little, E.L. Geographic Distribution of the Pines of the World; Department of Agriculture, Forest Service: Washington, DC, USA, 1966. [Google Scholar]
- Amjad, M.S.; Qaeem, M.F.; Ahmad, I.; Khan, S.U.; Chaudhari, S.K.; Malik, N.Z.; Shaheen, H.; Khan, A.M. Descriptive study of plant resources in the context of the ethnomedicinal relevance of indigenous flora: A case study from Toli Peer National Park, Azad Jammu and Kashmir, Pakistan. PLoS ONE 2017, 12, e0171896. [Google Scholar] [CrossRef]
- Malik, A.R.; Shamet, G.S.; Butola, J.S. Natural regeneration status of chilgoza pine (Pinus gerardiana wall.) in Himachal Pradesh, India: An endangered pine of high edible value. Appl. Ecol. Env. Res. 2012, 10, 365–373. [Google Scholar] [CrossRef]
- Dogra, P. Gymnosperms of India: II. Chilgoza Pine (Pinus gerardiana wall); National Botanic Gardens: Lucknow, India, 1964. [Google Scholar]
- Krugman, S.L.; Jenkinson, J.L. Pinus L. Pine. In Seeds of Woody Plants in the United States, Agricultural Handbook; Department of Agriculture: Washington, DC, USA, 1974; pp. 598–638. [Google Scholar]
- Farjon, A. Pines: Drawings and Descriptions of the Genus Pinus; EJ Brill: Leiden, The Netherlands, 1984. [Google Scholar]
- Farjon, A. World Checklist and Bibliography of Conifers; Royal Botanical Gardens Kew: Richmond, UK, 1998. [Google Scholar]
- Ilyas, M.; Qureshi, R.; Akhtar, N.; Ziaul-Haq, M.K.A.; Khan, A.M. Floristic diversity and vegetation structure of the remnant subtropical broad leaved forests from Kabal valley, Swat, Pakistan. Pak. J. Bot. 2018, 50, 217–230. [Google Scholar]
- Khan, H.M.A.; Zaman, M.; Hyder, A.A.S.; Nafees, M.K.M.A.; Begum, G.R.F.; Hussain, S.A.; Ali, M. Diameter size class distributions of Pinus gerardiana Wall. ex D. Don from Gohar Abad Valley District Diamer, Gilgit-Baltistan, Pakistan. J. Biodivers. Environ. Sci. 2015, 6, 50–56. [Google Scholar]
- Ahmed, M.; Ashfaq, M.; Amjad, M.; Saeed, M. Vegetation structure and dynamics of Pinus gerardiana forests in Balouchistan, Pakistan. J. Veg. Sci. 1991, 2, 119–124. [Google Scholar] [CrossRef]
- Mossadegh, A. World Forest Geography; Tehran University Press: Tehran, Iran, 2005. [Google Scholar]
- Ahmed, M.; Husain, T.; Sheikh, A.H.; Hussain, S.S.; Siddiqui, M.F. Phytosociology and structure of Himalayan forests from different climatic zones of Pakistan. Pak. J. Bot. 2006, 38, 361. [Google Scholar]
- Rafi, M. Vegetation Types of Balochistan Province. Pak. Govt. Print. Press. Punjab. Lahore Pakistan 1965, 116, 1–8. [Google Scholar]
- Chaudhri, I.I. Distribution of gymnosperms in West Pakistan. Vegetation 1963, 11, 372–382. [Google Scholar] [CrossRef]
- Tariq, A.; Shu, H.; Siddiqui, S.; Munir, I.; Sharifi, A.; Li, Q.; Lu, L. Spatio-temporal analysis of forest fire events in the Margalla Hills, Islamabad, Pakistan using socio-economic and environmental variable data with machine learning methods. J. For. Res. 2021, 13, 12. [Google Scholar] [CrossRef]
- Urooj, R.; Jabeen, A. Present status of Pinus gerardiana Wall. in Pakistan: A review. Middle East J. Bus. 2015, 10, 8897. [Google Scholar] [CrossRef]
- Saeed, M.; Thanos, C.A. The effect of seed coat removal on seed germination of Pinus gerardiana Wallich ex D. Don. chilgoza pine. J. Appl. Emerg. Sci. 2006, 1, 174–177. [Google Scholar]
- Khan, A.M.; Qureshi, R.; Saqib, Z.; Habib, T.; Ilyas, M.; Maqsood, M.; Kosar, R.; Akram, M.; Rahim, B.Z. A novel study of the interrelationship of seasonality, satellite data and weed compositional changes of the agro-ecological system of Gujrat, Pakistan. Appl. Ecol. Environ. Res. 2018, 16, 2995–3018. [Google Scholar] [CrossRef]
- Ali, F.; Khan, N.; Ahmad, A.; Khan, A.A. Structure and biomass carbon of Olea ferruginea forests in the foot hills of Malakand division, Hindukush Range Mountains of Pakistan. Acta Ecol. Sin. 2019, 39, 261–266. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. World Clim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Khan, A.M.; Qureshi, R.; Saqib, Z. Multivariate analyses of the vegetation of the western Himalayan forests of Muzaffarabad district, Azad Jammu and Kashmir, Pakistan. Ecol. Indic. 2019, 104, 723–736. [Google Scholar] [CrossRef]
- Khan, A.M.; Qureshi, R.; Saqib, Z.; Munir, M.; Shaheen, H.; Habib, T.; Dar, M.E.U.I.; Fatimah, H.; Afza, R.; Hussain, M. A first ever detailed ecological exploration of the western Himalayan forests of Sudhan Gali and Ganga summit, Azad Jammu and Kashmir, Pakistan. Appl. Ecol. Environ. Res. 2019, 17, 15477–15505. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Gao, T.; Xu, Q.; Liu, Y.; Zhao, J.; Shi, J. Predicting the Potential Geographic Distribution of Sirex nitobei in China under Climate Change Using Maximum Entropy Model. Forests 2021, 12, 151. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Optimized maxent model predictions of climate change impacts on the suitable distribution of cunninghamia lanceolata in China. Forests 2020, 11, 302. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; p. 83. [Google Scholar] [CrossRef] [Green Version]
- Radosavljevic, A.; Anderson, R.P.; Araújo, M. Making better Maxentmodels of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Manel, S.; Ceri Williams, H.; Ormerod, S.J. Evaluating presence-absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Van Proosdij, A.S.J.; Sosef, M.S.M.; Wieringa, J.J.; Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 2016, 39, 542–552. [Google Scholar] [CrossRef]
- Akhlaq, R.; Amjad, M.S.; Qaseem, M.F.; Fatima, S.; Chaudhari, S.K.; Khan, A.M. Species diversity and vegetation structure from different climatic zones of tehsil Harighel, Bagh, Azad Kasmir, Pakistan analysed through multivariate techniques. Appl. Ecol. Environ. Res. 2018, 16, 5193–5211. [Google Scholar] [CrossRef]
- Marshall, A.G.; Hudson, D.; Wheeler, M.C.; Hendon, H.H.; Alves, O. Simulation and prediction of the Southern Annular Mode and its influence on Australian intra-seasonal climate in POAMA. Clim. Dyn. 2012, 38, 2483–2502. [Google Scholar] [CrossRef]
- Bosso, L.; Luchi, N.; Maresi, G.; Cristinzio, G.; Smeraldo, S.; Russo, D. Predicting current and future disease outbreaks of Diplodia sapinea shoot blight in Italy: Species distribution models as a tool for forest management planning. For. Ecol. Manag. 2017, 400, 655–664. [Google Scholar] [CrossRef]
- Liu, J.; Milne, R.I.; Mo¨ller, M.; Zhu, G.F.; Ye, L.J.; Luo, Y.H.; Yang, J.B.; Wambulwa, M.C.; Wang, C.N.; Li, D.Z.; et al. Integrating a comprehensive DNA barcode reference library with a global map of yews (Taxus L.) for forensic identification. Mol. Ecol. Resour. 2018, 18, 1115–1131. [Google Scholar] [CrossRef]
- Shalizi, M.N.; Khurram, S.; Groninger, J.W.; Ruffner, C.M.; Burney, O.T. Indigenous knowledge and stand characteristics of a threatened tree species in a highly insecure area: Chilgoza pine in Afghanistan. For. Ecol. Manag. 2018, 413, 1–8. [Google Scholar] [CrossRef]
- Rahimzadeh, A. Socio-economic and Environmental Implications of the Decline of Chilgoza Pine Nuts of Kinnaur, Western Himalaya. Conserv. Soc. 2020, 18, 315–326. [Google Scholar] [CrossRef]
- Akbar, M.; Ahmed, M.; Alamdar, H.; Zafar, M.U.; Khan, M. Quantitative forests description from Skardu, Gilgit and Astore Districts of Gilgit- Baltistan, Pakistan. Fuuast J. Biol. 2011, 1, 149–160. [Google Scholar]
- Aziz, M.A.; Adnan, M.; Hussain, S.K.; Hashem, A.; Alqarawi, A.A.; Abd_Allah, E.F. Comparative regeneration status of Pinus gerardiana in two forest-use types of Sulaiman mountain range near Pak-Afghan border region: Historical, current and future perspectives. Pak. J. Bot. 2017, 49, 227–236. [Google Scholar]
- Peltier, R.; Dauffy, V. The Chilgoza of Kinnaur. Influence of the Pinus gerardiana edible seed market chain organization on forest regeneration in the Indian Himalayas. Fruits 2009, 64, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Shamet, G.S.; Chaturvedi, O.P.; Avasthe, R.K.; Singh, C. Ecology of chilgoza pine (Pinus gerardiana Wall) in dry temperate forests of North West Himalaya. Ecol. Environ. Conserv. 2013, 19, 1063–1066. [Google Scholar]
- Parmesan, C.; Hanley, M.E. Plants and climate change: Complexities and surprises. Ann. Bot. 2015, 116, 849–864. [Google Scholar] [CrossRef]
- Ashraf, U.; Ali, H.; Chaudry, M.N.; Ashraf, I.; Batool, A.; Saqib, Z. Predicting the potential distribution of Olea ferruginea in Pakistan incorporating climate change by using Maxent model. Sustainability 2016, 8, 722. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 10. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Tariq, A.; Shu, H.; Gagnon, A.S.; Li, Q.; Mumtaz, F.; Hysa, A.; Siddique, M.A.; Munir, I. Assessing Burned Areas in Wildfires and Prescribed Fires with Spectral Indices and SAR Images in the Margalla Hills of Pakistan. Forests 2021, 12, 18. [Google Scholar] [CrossRef]
- Koc, D.E.; Svenning, J.C.; Meral, A.V.C.I. Climate change impacts on the potential distribution of Taxus baccata L. in the Eastern Mediterranean and the Bolkar Mountains (Turkey) from last glacial maximum to the future. Eurasian J. For. Sci. 2018, 6, 69–82. [Google Scholar]
- Li, P.; Zhu, W.; Xie, Z.; Qiao, K. Integration of multiple climate models to predict range shifts and identify management priorities of the endangered Taxus wallichiana in the Himalaya–Hengduan Mountain region. J. For. Res. 2020, 31, 2255–2272. [Google Scholar] [CrossRef] [Green Version]
- Khan, W.; Hussain, M.; Ali, K.; Ali, M.; Nisar, M. Distribution and phenotypic variation in Juglans regia L. growing in Hindu Kush ranges of Pakistan. Acta Ecol. Sin. 2020, 40, 363–372. [Google Scholar] [CrossRef]
- Awan, M.N.; Saqib, Z.; Buner, F.; Lee, D.C.; Pervez, A. Using ensemble modeling to predict breeding habitat of the red-listed Western Tragopan (Tragopan melanocephalus) in the Western Himalayas of Pakistan. Glob. Ecol. Conserv. 2021, 31, e01864. [Google Scholar] [CrossRef]
- Khalid, N.; Khan, A.M.; Qureshi, R.; Saqib, Z.; Zahid, N.; Bussmann, R.W. Exploration of important environmental determinants of flowering phenology in the western Himalayan forests of Dhirkot, Azad Jammu and Kashmir, Pakistan. Appl. Ecol. Environ. Res. 2022, 20, 301–337. [Google Scholar] [CrossRef]
- Thuiller, W. BIOMOD—Optimizing predictions of species distributions and projecting potential future shifts under global change. Glob. Chang. Biol. 2003, 9, 1353–1362. [Google Scholar] [CrossRef]
- Hamann, A.; Wang, T. Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecology 2006, 87, 2773–2786. [Google Scholar] [CrossRef] [Green Version]
- Fordham, D.A.; Akçakaya, H.R.; Araújo, M.B.; Keith, D.A.; Brook, B.W. Tools for integrating range change, extinction risk and climate change information into conservation management. Ecography 2013, 36, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Tariq, A.; Shu, H.; Siddiqui, S.; Mousa, B.G.; Munir, I.; Nasri, A.; Waqas, H.; Lu, L.; Baqa, M.F. Forest fire monitoring using spatial-statistical and Geo-spatial analysis of factors determining forest fire in Margalla Hills, Islamabad, Pakistan. Geomatics, Nat. Hazards Risk 2021, 12, 1212–1233. [Google Scholar] [CrossRef]
- Rodriguez, L.; Martínez, B.; Tuya, F. Atlantic corals under climate change: Modelling distribution shifts to predict richness, phylogenetic structure and trait-diversity changes. Biodivers. Conserv. 2019, 28, 3873–3890. [Google Scholar] [CrossRef]
- Dunk, J.R.; Woodbridge, B.; Schumaker, N.; Glenn, E.M.; White, B.; LaPlante, D.W.; Thrailkill, J. Conservation planning for species recovery under the Endangered Species Act: A case study with the Northern Spotted Owl. PLoS ONE 2019, 14, e0210643. [Google Scholar] [CrossRef] [PubMed]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.R. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Kaky, E.; Gilbert, F. Using species distribution models to assess the importance of Egypt's protected areas for the conservation of medicinal plants. J. Arid Environ. 2016, 135, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; Fois, M.; Dakhil, M.A.; Bacchetta, G.; El-Sherbeny, G.A. Predicting the potential current and future distribution of the endangered endemic vascular plant Primula Boveana Decne. ex Duby in Egypt. Plants 2020, 9, 957. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Kaky, E.; Gilbert, F. Allowing for human socioeconomic impacts in the conservation of plants under climate change. Plant Biosyst. 2019, 154, 295–305. [Google Scholar] [CrossRef]
- Majeed, M.; Bhatti, K.H.; Pieroni, A.; Sõukand, R.; Bussmann, R.W.; Khan, A.M.; Chaudhari, S.K.; Aziz, M.A.; Amjad, M.S. Gathered Wild Food Plants among Diverse Religious Groups in Jhelum District, Punjab, Pakistan. Foods 2021, 10, 594. [Google Scholar] [CrossRef]
- Liang, E.; Leuschner, C.; Dulamsuren, C.; Wagner, B.; Hauck, M. Global warming-related tree growth decline and mortality on the north-eastern Tibetan plateau. Clim. Chang. 2016, 134, 163–176. [Google Scholar] [CrossRef]
- Liang, E.Y.; Wang, Y.F.; Piao, S.L.; Lu, X.M.; Camarero, J.J.; Zhu, H.F.; Zhu, L.P.; Ellison, A.M.; Ciais, P.; Pen˜uelas, J. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2016, 113, 4380–4385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Duveneck, M.J.; Gustafson, E.J.; Serra-Diaz, J.M.; Thompson, J.R. How disturbance, competition, and dispersal interact to prevent tree range boundaries from keeping pace with climate change. Glob. Chang. Biol. 2018, 24, e335–e351. [Google Scholar] [CrossRef]
- Wang, D.J.; Wei, H.Y.; Zhang, X.H.; Fang, Y.Q.; Gu, W. Habitat suitability modeling based on remote sensing to realize time synchronization of species and environmental variables. J. Plant Ecol. 2021, 14, 241–256. [Google Scholar] [CrossRef]
- Wang, W.J.; He, H.S.; Thompson, F.R.; Spetich, M.A.; Fraser, J.S. Effects of species biological traits and environmental heterogeneity on simulated tree species distribution shifts under climate change. Sci. Total Environ. 2018, 634, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Poudel, R.C.; Moller, M.; Liu, J.; Gao, L.M.; Baral, S.R.; Li, D.Z. Low genetic diversity and high inbreeding of the endangered yews in Central Himalaya: Implications for conservation of their highly fragmented populations. Divers. Distrib. 2014, 20, 1270–1284. [Google Scholar] [CrossRef]
- Tariq, A.; Riaz, I.; Ahmad, Z.; Amin, M.; Kausar, R.; Andleeb, S.; Farooqi, M.A.; Rafiq, M. Land surface temperature relation with normalized satellite indices for the estimation of spatio-temporal trends in temperature among various land use land cover classes of an arid Potohar region using Landsat data. Environ. Earth Sci. 2020, 79, 40. [Google Scholar] [CrossRef]
- Tariq, A.; Shu, H. CA-Markov chain analysis of seasonal land surface temperature and land use landcover change using optical multi-temporal satellite data of Faisalabad, Pakistan. Remote Sens. 2020, 12, 3402. [Google Scholar] [CrossRef]
- Tariq, A.; Shu, H.; Siddiqui, S.; Imran, M.; Farhan, M. Monitoring land use and land cover changes using geospatial techniques, a case study of Fateh Jang, Attock, Pakistan. Geogr. Environ. Sustain. 2021, 14, 41–52. [Google Scholar] [CrossRef]
- Baqa, M.F.; Chen, F.; Lu, L.; Qureshi, S.; Tariq, A.; Wang, S.; Jing, L.; Hamza, S.; Li, Q. Monitoring and modeling the patterns and trends of urban growth using urban sprawl matrix and CA-Markov model: A case study of Karachi, Pakistan. Land 2021, 10, 700. [Google Scholar] [CrossRef]
- Hu, P.; Sharifi, A.; Tahir, M.N.; Tariq, A.; Zhang, L.; Mumtaz, F.; Shah, S.H.I.A. Evaluation of Vegetation Indices and Phenological Metrics Using Time-Series MODIS Data for Monitoring Vegetation Change in Punjab, Pakistan. Water 2021, 13, 2550. [Google Scholar] [CrossRef]
- Sharifi, A.; Mahdipour, H.; Moradi, E.; Tariq, A. Agricultural Field Extraction with Deep Learning Algorithm and Satellite Imagery. J. Indian Soc. Remote Sens. 2022, 50, 417–423. [Google Scholar] [CrossRef]
- Benito, B.M.; Cayuela, L.; Albuquerque, F.S. The impact of modelling choices in the predictive performance of richness maps derived from species-distribution models: Guidelines to build better diversity models. Methods Ecol. Evol. 2013, 4, 327–335. [Google Scholar] [CrossRef]
- Distler, T.; Schuetz, J.G.; Velásquez-Tibatá, J.; Langham, G.M. Stacked species distribution models and macroecological models provide congruent projections of avian species richness under climate change. J. Biogeogr. 2015, 42, 976–988. [Google Scholar] [CrossRef]
- Alatawi, A.S.; Gilbert, F.; Reader, T. Modelling terrestrial reptile species richness, distributions and habitat suitability in Saudi Arabia. J. Arid Environ. 2020, 178, 104–153. [Google Scholar] [CrossRef]
- Gilani, H.; Goheer, M.A.; Ahmad, H.; Hussain, K. Under predicted climate change: Distribution and ecological niche modelling of six native tree species in Gilgit-Baltistan, Pakistan. Ecol. Indic. 2020, 111, 106049. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Name of Variable & Description | Database | Resolution | Unit |
|---|---|---|---|---|
| Bio1 | Annual Mean Temperature | WorldClim | 30 arc s | °C |
| Bio2 | Mean Diurnal Range | WorldClim | 30 arc s | °C |
| Bio3 | Isothermality (Bio2/Bio7) (×100) | WorldClim | 30 arc s | Percent |
| Bio4 | Temperature Seasonality (sd ×100) | WorldClim | 30 arc s | °C |
| Bio5 | Max. Temperature of Warmest Month | WorldClim | 30 arc s | °C |
| Bio6 | Min. Temperature of Coldest Month | WorldClim | 30 arc s | °C |
| Bio7 | Temperature Annual Range | WorldClim | 30 arc s | °C |
| Bio8 | Mean Temperature of Wettest Quarter | WorldClim | 30 arc s | °C |
| Bio9 | Mean Temperature of Driest Quarter | WorldClim | 30 arc s | °C |
| Bio10 | Mean Temperature of Warmest Quarter | WorldClim | 30 arc s | °C |
| Bio11 | Mean Temperature of Coldest Quarter | WorldClim | 30 arc s | °C |
| Bio12 | Annual Precipitation | WorldClim | 30 arc s | Mm |
| Bio13 | Precipitation of Wettest Month | WorldClim | 30 arc s | Mm |
| Bio14 | Precipitation of Driest Month | WorldClim | 30 arc s | Mm |
| Bio15 | Precipitation Seasonality (CV) | WorldClim | 30 arc s | Percent |
| Bio16 | Precipitation of Wettest Quarter | WorldClim | 30 arc s | Mm |
| Bio17 | Precipitation of Driest Quarter | WorldClim | 30 arc s | Mm |
| Bio18 | Precipitation of Warmest Quarter | WorldClim | 30 arc s | Mm |
| Bio19 | Precipitation of Coldest Quarter | WorldClim | 30 arc s | Mm |
| Bdod | Bulk Density | SoilGrids | 30 arc s | cg/cm3 |
| Cec | Cations Exchange Capacity (pH: 7) | SoilGrids | 30 arc s | mmol(c)/kg |
| Cfvo | Volumetric fraction of coarse fragments (>2 mm) | SoilGrids | 30 arc s | cm3/dm3 |
| Clay | Clay Contents | SoilGrids | 30 arc s | g/kg |
| Nitrogen | Total Nitrogen | SoilGrids | 30 arc s | cg/kg |
| Ocd | Organic Carbon Density | SoilGrids | 30 arc s | hg/dm3 |
| Phh2o | Soil pH × 10 | SoilGrids | 30 arc s | Nil |
| Sand | Sand Contents | SoilGrids | 30 arc s | g/kg |
| Silt | Silt Contents | SoilGrids | 30 arc s | g/kg |
| Soc | Soil Organic Carbon | SoilGrids | 30 arc s | dg/kg |
| NDVI | NDVI (MODIS/006/MOD13A2) | NASA LP DAAC | 30 arc s | Nil |
| Elevation | Elevation | SRTM DEM Global | 30 arc s | meter |
| Alf | Global ALOS Landforms | Global Science Partners | 30 arc s | Nil |
| Chin | Continuous Heat-Insolation Load Index (Global ALOS CHILI) | Global Science Partners | 30 arc s | Nil |
| Hillshade | Hillshade | SRTM DEM Global | 30 arc s | Degree |
| Northness | Cos-Transformed-Aspect (0–1) | Derived | 30 arc s | Nil |
| Slope | Slope | SRTM DEM Global | 30 arc s | Degree |
| Tdiv | Topographic Diversity | Cons. Science Partners | 30 arc s | Nil |
| Climatic Scenario | AUC | AUC Ratios | TSS | Kappa | p-AUC-ROC |
|---|---|---|---|---|---|
| Current climate | 0.957 | 1.68 | 0.939 | 0.951 | 0.84 |
| SSPs–245 (2050s) | 0.951 | 1.64 | 0.926 | 0.933 | 0.82 |
| SSPs–585 (2050s) | 0.953 | 1.66 | 0.933 | 0.945 | 0.83 |
| SSPs–245 (2070s) | 0.954 | 1.66 | 0.932 | 0.947 | 0.83 |
| SSPs–585 (2070s) | 0.956 | 1.64 | 0.936 | 0.952 | 0.82 |
| Variable | Code | Percent Contribution |
|---|---|---|
| Normalized Difference Vegetation Index | NDVI | 40.2 |
| Mean temperature of coldest quarter | Bio11 | 37.5 |
| Isothermality (Bio2/Bio7) (×100) | Bio3 | 11.7 |
| Precipitation of driest month | Bio14 | 6.3 |
| Volumetric fraction of coarse fragments | CFVO | 4.3 |
| Climate Scenario | Country | HSC-1 (0–0.2) | HSC-2 (0.2–0.4) | HSC-3 (0.4–0.6) | HSC-4 (0.6–0.8) | HSC-5 (0.8–1) | Total Suitable Area |
|---|---|---|---|---|---|---|---|
| Current climate | Afghanistan | 605,974 | 16,441 | 12,848 | 9125 | 8472 | 46,886 |
| SSPs 245 (2050s) | Afghanistan | 607,043 | 17,812 | 11,665 | 9172 | 7167 | 45,817 |
| Rate of change (%) | 0 | 8 | −10 | 1 | −17 | −2 | |
| SSPs 585 (2050s) | Afghanistan | 605,706 | 17,961 | 11,363 | 9778 | 8052 | 47,154 |
| Rate of change (%) | 0 | 9 | −12 | 7 | −5 | 1 | |
| SSPs 245 (2070s) | Afghanistan | 613,207 | 16,555 | 8844 | 7972 | 6282 | 39,653 |
| Rate of change (%) | 1 | 1 | −37 | −14 | −30 | −17 | |
| SSPs 585 (2070s) | Afghanistan | 616,072 | 13,836 | 7824 | 8699 | 6428 | 36,788 |
| Rate of change (%) | 2 | −17 | −50 | −5 | −28 | −24 | |
| Current climate | China | 9,697,953 | 7266 | 1294 | 273 | 7 | 8840 |
| SSPs 245 (2050s) | China | 9,689,273 | 7313 | 6296 | 3348 | 564 | 17,521 |
| Rate of change (%) | 0 | 1 | 158 | 251 | 437 | 68 | |
| SSPs 585 (2050s) | China | 9,687,491 | 7404 | 7024 | 4559 | 315 | 19,302 |
| Rate of change (%) | 0 | 2 | 169 | 282 | 379 | 78 | |
| SSPs 245 (2070s) | China | 9,691,178 | 7800 | 6465 | 1246 | 104 | 15,615 |
| Rate of change (%) | 0 | 7 | 161 | 152 | 268 | 57 | |
| SSPs 585 (2070s) | China | 9,687,158 | 10,862 | 7420 | 1324 | 29 | 19,635 |
| Rate of change (%) | 0 | 40 | 175 | 158 | 141 | 80 | |
| Current climate | India | 3,249,627 | 9114 | 9664 | 8060 | 6564 | 33,402 |
| SSPs 245 (2050s) | India | 3,255,598 | 9870 | 7051 | 4341 | 6170 | 27,432 |
| Rate of change (%) | 0 | 8 | −32 | −62 | −6 | −20 | |
| SSPs 585 (2050s) | India | 3,252,309 | 10,250 | 8975 | 4979 | 6516 | 30,720 |
| Rate of change (%) | 0 | 12 | −7 | −48 | −1 | −8 | |
| SSPs 245 (2070s) | India | 3,266,559 | 8455 | 4105 | 2589 | 1322 | 16,471 |
| Rate of change (%) | 1 | −8 | −86 | −114 | −160 | −71 | |
| SSPs 585 (2070s) | India | 3,263,276 | 9709 | 5384 | 3400 | 1260 | 19,754 |
| Rate of change (%) | 0 | 6 | −59 | −86 | −165 | −53 | |
| Current climate | Pakistan | 832,259 | 17,384 | 12,230 | 8696 | 11,344 | 49,654 |
| SSPs 245 (2050s) | Pakistan | 852,249 | 11,259 | 8552 | 5733 | 4120 | 29,664 |
| Rate of change (%) | 2 | −43 | −36 | −42 | −101 | −52 | |
| SSPs 585 (2050s) | Pakistan | 855,573 | 8569 | 7307 | 6466 | 3998 | 26,340 |
| Rate of change (%) | 3 | −71 | −52 | −30 | −104 | −63 | |
| SSPs 245 (2070s) | Pakistan | 859,874 | 8953 | 5360 | 4056 | 3669 | 22,039 |
| Rate of change (%) | 3 | −66 | −82 | −76 | −113 | −81 | |
| SSPs 585 (2070s) | Pakistan | 864,221 | 6171 | 4510 | 4456 | 2556 | 17,692 |
| Rate of change (%) | 4 | −104 | −100 | −67 | −149 | −103 | |
| Current climate | Total | 14,385,814 | 50,204 | 36,037 | 26,154 | 26,387 | 138,782 |
| SSPs 245 (2050s) | Total | 14,404,163 | 46,255 | 33,564 | 22,594 | 18,021 | 120,433 |
| Rate of change (%) | 0 | −8 | −7 | −15 | −38 | −14 | |
| SSPs 585 (2050s) | Total | 14,401,080 | 44,184 | 34,670 | 25,782 | 18,881 | 123,516 |
| Rate of change (%) | 0 | −13 | −4 | −1 | −33 | −12 | |
| SSPs 245 (2070s) | Total | 14,430,817 | 41,764 | 24,774 | 15,864 | 11,377 | 93,779 |
| Rate of change (%) | 0 | −18 | −37 | −50 | −84 | −39 | |
| SSPs 585 (2070s) | Total | 14,430,727 | 40,579 | 25,138 | 17,879 | 10,274 | 93,869 |
| Rate of change (%) | 0 | −21 | −36 | −38 | −94 | −39 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.M.; Li, Q.; Saqib, Z.; Khan, N.; Habib, T.; Khalid, N.; Majeed, M.; Tariq, A. MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia. Forests 2022, 13, 715. https://doi.org/10.3390/f13050715
Khan AM, Li Q, Saqib Z, Khan N, Habib T, Khalid N, Majeed M, Tariq A. MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia. Forests. 2022; 13(5):715. https://doi.org/10.3390/f13050715
Chicago/Turabian StyleKhan, Arshad Mahmood, Qingting Li, Zafeer Saqib, Nasrullah Khan, Tariq Habib, Nadia Khalid, Muhammad Majeed, and Aqil Tariq. 2022. "MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia" Forests 13, no. 5: 715. https://doi.org/10.3390/f13050715
APA StyleKhan, A. M., Li, Q., Saqib, Z., Khan, N., Habib, T., Khalid, N., Majeed, M., & Tariq, A. (2022). MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia. Forests, 13(5), 715. https://doi.org/10.3390/f13050715