
Regularly Planted Rather Than Natural Understory of Norway Spruce (Picea abies H. Karst.) Contributes to the Individual Stability of Canopy Silver Birch (Betula pendula Roth.)

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Trees
2.2. Static Tree-Pulling Tests
2.3. Data Processing and Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Forzieri, G.; Girardello, M.; Ceccherini, G.; Spinoni, J.; Feyen, L.; Hartmann, H.; Beck, P.S.A.; Camps-Valls, G.; Chirici, G.; Mauri, A.; et al. Emergent vulnerability to climate-driven disturbances in European forests. Nat. Commun. 2021, 12, 1081. [Google Scholar] [CrossRef] [PubMed]
- Oouchi, K.; Yoshimura, J.; Yoshimura, H.; Mizuta, R.; Kusunoki, S.; Noda, A. Tropical cyclone climatology in a global-warming climate as simulated in a 20 km-mesh global atmospheric model: Frequency and wind intensity analyses. J. Meteorol. Soc. Jpn. Ser. II 2006, 84, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Della-Marta, P.M.; Mathis, H.; Frei, C.; Lininger, M.A.; Kleinn, J.; Appenzeller, C. The return period of wind storms over Europe. Int. J. Climatol. 2009, 29, 437–459. [Google Scholar] [CrossRef]
- Griess, V.C.; Knoke, T. Growth performance, windthrow, and insects: Meta-analyses of parameters influencing performance of mixed-species stands in boreal and northern temperate biomes. Can. J. For. Res. 2011, 41, 1141–1159. [Google Scholar] [CrossRef]
- Valinger, E.; Fridman, J. Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in southern Sweden. For. Ecol. Manag. 2011, 262, 398–403. [Google Scholar] [CrossRef]
- Jactel, H.; Bauhus, J.; Boberg, J.; Bonal, D.; Castagneyrol, B.; Gardiner, B.; Gonzalez-Olabarria, J.R.; Koricheva, J.; Meurisse, N.; Brockerhoff, E.G. Tree diversity drives forest stand resistance to natural disturbances. Curr. For. Rep. 2017, 3, 223–243. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Mason, W.L. Are irregular stands more windfirm? For. Int. J. For. Res. 2002, 75, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Pukkala, T.; Laiho, O.; Lähde, E. Continuous cover management reduces wind damage. For. Ecol. Manag. 2016, 372, 120–127. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Blennow, K. Simulating wind disturbance impacts on forest landscapes: Tree-level heterogeneity matters. Environ. Model. Softw. 2014, 51, 1–11. [Google Scholar] [CrossRef]
- Díaz-Yáñez, O.; Mola-Yudego, B.; González-Olabarria, J.R.; Pukkala, T. How does forest composition and structure affect the stability against wind and snow? For. Ecol. Manag. 2017, 401, 215–222. [Google Scholar] [CrossRef]
- Bauhus, J.; Forrester, D.I.; Gardiner, B.; Jactel, H.; Vallejo, R.; Pretzsch, H. Ecological stability of mixed-species forests. In Mixed Species Forests; Springer: Berlin, Germany, 2017; pp. 337–382. [Google Scholar] [CrossRef]
- Gardiner, B.; Blennow, K.; Carnus, J.-M.; Fleischer, P.; Ingemarson, F.; Landmann, G.; Lindner, M.; Marzano, M.; Nicoll, B.; Orazio, C.; et al. Destructive Storms in European Forests: Past and Forthcoming Impacts; European Forests Institute: Joensuu, Finland, 2010. [Google Scholar]
- Nicoll, B.C.; Ray, D. Adaptive growth of tree root systems in response to wind action and site conditions. Tree Physiol. 1996, 16, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Peltola, H.; Kellomäki, S.; Hassinen, A.; Granander, M. Mechanical stability of Scots pine, Norway spruce and birch: An analysis of tree-pulling experiments in Finland. For. Ecol. Manag. 2000, 135, 143–153. [Google Scholar] [CrossRef]
- Krišāns, O.; Matisons, R.; Kitenberga, M.; Donis, J.; Rust, S.; Elferts, D.; Jansons, Ā. Wind resistance of Eastern Baltic silver birch (Betula pendula roth.) suggests its suitability for periodically waterlogged sites. Forests 2021, 12, 21. [Google Scholar] [CrossRef]
- Gardiner, B. Wind damage to forests and trees: A review with an emphasis on planted and managed forests. J. For. Res. 2021, 26, 248–266. [Google Scholar] [CrossRef]
- Dhôte, J.-F. Implication of forest diversity in resistance to strong winds. In Forest Diversity and Function; Springer: Berlin/Heidelberg, Germany, 2005; pp. 291–307. [Google Scholar] [CrossRef]
- Bergeron, C.; Ruel, J.C.; Élie, J.G.; Mitchell, S.J. Root anchorage and stem strength of black spruce (Picea mariana) trees in regular and irregular stands. For. Int. J. For. Res. 2009, 82, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.G.; Gong, M. Mechanical stimulation-induced cross-adaptation in plants: An overview. J. Plant Biol. 2011, 54, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Hynynen, J.; Niemistö, P.; Viherä-Aarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in Northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]
- Baders, E.; Senhofa, S.; Purina, L.; Jansons, A. Baltic forestry natural succession of Norway spruce stands in hemiboreal forests: Case study in Slitere National Park, Latvia. Balt. For. 2017, 23, 522–528. [Google Scholar]
- Bāders, E.; Lūkins, M.; Zariņš, J.; Krišāns, O.; Jansons, Ā.; Jansons, J. Recent land cover changes in Latvia. Res. Rural Dev. 2018, 1, 34–39. [Google Scholar] [CrossRef]
- Latvian State Forest Research Institute “Silava”. National Forest Inventory of Latvia. Available online: http://www.silava.lv/petijumi/nacionlais-mea-monitorings.aspx (accessed on 7 February 2022).
- Gailis, A.; Karklina, A.; Purvinš, A.; Matisons, R.; Zeltinš, P.; Jansons, A. Effect of breeding on income at first commercial thinning in silver birch plantations. Forests 2020, 11, 327. [Google Scholar] [CrossRef] [Green Version]
- Belda, M.; Holtanová, E.; Halenka, T.; Kalvová, J. Climate classification revisited: From Köppen to Trewartha. Clim. Res. 2014, 59, 1–13. [Google Scholar] [CrossRef] [Green Version]
- LEGMC Latvian Environment, Geology and Meteorology Centre. Climate in Latvia. 2020. Available online: https://klimats.meteo.lv/klimats/latvijas_klimats/ (accessed on 21 March 2022).
- Jaagus, J.; Briede, A.; Rimkus, E.; Remm, K. Precipitation pattern in the Baltic countries under the influence of large-scale atmospheric circulation and local. Int. J. Climatol. 2010, 30, 705–720. [Google Scholar] [CrossRef] [Green Version]
- Karagali, I.; Hahmann, A.N.; Badger, M.; Hasager, C.; Mann, J. New European wind atlas offshore. J. Phys. Conf. Ser. 2018, 1037, 052007. [Google Scholar] [CrossRef]
- Krišāns, O.; Čakša, L.; Matisons, R.; Rust, S.; Elferts, D.; Seipulis, A.; Jansons, Ā. A static pulling test is a suitable method for comparison of the loading resistance of silver birch (Betula pendula roth.) between urban and peri-urban forests. Forests 2022, 13, 127. [Google Scholar] [CrossRef]
- Hui, G.; Zhang, G.; Zhao, Z.; Yang, A. Methods of forest structure research: A review. Curr. For. Rep. 2019, 5, 142–154. [Google Scholar] [CrossRef]
- Detter, A.; Richter, K.; Rust, C.; Rust, S. Aktuelle Untersuchungen zum Primärversagen von grünem Holz-Current studies on primary failure in green wood. In Proceedings of the Conference Deutsche Baumpflegetage, Augsburg, Germany, 5–7 May 2015; pp. 156–167. [Google Scholar]
- Detter, A.; Rust, S.; Rust, C.; Maybaum, G. Determining strength limits for standing tree stems from bending tests. In Proceedings of the 18th International Nondestructive Testing and Evaluation of Wood Symposium, Madison, WI, USA, 24–27 September 2013; Ross, R.J., Wang, X., Eds.; U.S. Department of Agriculture, ForestService, Forest Products Laboratory: Madison, WI, USA, 2013; p. 226. [Google Scholar]
- Detter, A.; Van Wassenaer, P.; Rust, S. Stability recovery in London Plane trees eight years after primary anchorage failure. Arboric. Urban For. 2019, 45, 279–288. [Google Scholar] [CrossRef]
- Krišāns, O.; Matisons, R.; Vuguls, J.; Rust, S.; Elferts, D.; Seipulis, A.; Saleniece, R.; Jansons, Ā. Silver birch (Betula pendula Roth.) on dry mineral rather than on deep peat soils is more dependent on frozen conditions in terms of wind damage in the Eastern Baltic region. Plants 2022, 11, 1174. [Google Scholar] [CrossRef]
- Core Team, R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Bartoń, K. MuMIn: Multi-Model Inference. Available online: http://cran.rproject.org/package=MuMIn (accessed on 19 March 2022).
- Csilléry, K.; Kunstler, G.; Courbaud, B.; Allard, D.; Lassègues, P.; Haslinger, K.; Gardiner, B. Coupled effects of wind-storms and drought on tree mortality across 115 forest stands from the Western Alps and the Jura mountains. Glob. Chang. Biol. 2017, 23, 5092–5107. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Peet, R.K.; Christensen, N.L. Competition and tree death. Bioscience 1987, 37, 586–595. [Google Scholar] [CrossRef]
- Forrester, D.I. Linking forest growth with stand structure: Tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For. Ecol. Manag. 2019, 447, 139–157. [Google Scholar] [CrossRef]
- Samariks, V.; Īstenais, N.; Seipulis, A.; Miezīte, O.; Krišāns, O.; Jansons, Ā. Root-soil plate characteristics of silver birch on wet and dry mineral soils in Latvia. Forests 2021, 12, 20. [Google Scholar] [CrossRef]
- Shanin, V.; Grabarnik, P.; Shashkov, M.; Ivanova, N.; Bykhovets, S.; Frolov, P.; Stamenov, M. Crown asymmetry and niche segregation as an adaptation of trees to competition for light: Conclusions from simulation experiments in mixed boreal stands. Math. Comput. For. Nat. Resour. Sci. 2020, 12, 26–49. [Google Scholar]
- Flepp, G.; Robyr, R.; Scotti, R.; Giadrossich, F.; Conedera, M.; Vacchiano, G.; Fischer, C.; Ammann, P.; May, D.; Schwarz, M. Temporal dynamics of root reinforcement in European spruce forests. Forests 2021, 12, 815. [Google Scholar] [CrossRef]
- Külla, T.; Lõhmus, K. Influence of cultivation method on root grafting in Norway spruce (Picea abies (L.) Karst.). In The Supporting Roots of Trees and Woody Plants: Form, Function and Physiology; Stokes, A., Ed.; Springer: Dordrecht, The Netherlands, 2000; pp. 109–118. ISBN 978-94-017-3469-1. [Google Scholar]
- Graham, B.F.; Bormann, F.H. Natural root grafts. Bot. Rev. 1966, 32, 255–292. [Google Scholar] [CrossRef]
- Quine, C.P.; Gardiner, B.A.; Moore, J. Wind disturbance in forests: The process of wind created gaps, tree overturning, and stem breakage. In Plant Disturbance Ecology: The Process and the Response; Johnson, E.A., Miyanishi, K., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 117–184. ISBN 9780128188132. [Google Scholar]
- Skomarkova, M.V.; Vaganov, E.A.; Mund, M.; Knohl, A.; Linke, P.; Boerner, A.; Schulze, E.D. Inter-annual and seasonal variability of radial growth, wood density and carbon isotope ratios in tree rings of beech (Fagus sylvatica) growing in Germany and Italy. Trees 2006, 20, 571–586. [Google Scholar] [CrossRef]
- Voelker, S.L.; Lachenbruch, B.; Meinzer, F.C.; Strauss, S.H. Reduced wood stiffness and strength, and altered stem form, in young antisense 4CL transgenic poplars with reduced lignin contents. New Phytol. 2011, 189, 1096–1109. [Google Scholar] [CrossRef]
- Waghorn, M.J.; Mason, E.G.; Watt, M.S. Influence of initial stand density and genotype on longitudinal variation in modulus of elasticity for 17-year-old Pinus radiata. For. Ecol. Manag. 2007, 1–3, 67–72. [Google Scholar] [CrossRef]
- Šilinskas, B.; Varnagiryte-Kabašinskiene, I.; Aleinikovas, M.; Beniušiene, L.; Aleinikoviene, J.; Škema, M. Scots pine and Norway spruce wood properties at sites with different stand densities. Forests 2020, 11, 587. [Google Scholar] [CrossRef]
- Christensen-Dalsgaard, K.K.; Ennos, A.R. Effects of drought acclimation on the mechanical properties of Ochroma pyramidale, Betula pendula and Acacia karroo tree seedling stems. Forestry 2012, 85, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Koponen, T.; Karppinen, T.; Hæggström, E.; Saranpää, P.; Serimaa, R. The stiffness modulus in Norway spruce as a function of year ring. Holzforschung 2005, 59, 451–455. [Google Scholar] [CrossRef]
- Pollet, C.; Henin, J.M.; Hébert, J.; Jourez, B. Effect of growth rate on the physical and mechanical properties of Douglas-fir in western Europe. Can. J. For. Res. 2017, 47, 1056–1065. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S.; Väisänen, H.; Ikonen, V.P. A mechanistic model for assessing the risk of wind and snow damage to single trees and stands of Scots pine, Norway spruce, and birch. Can. J. For. Res. 1999, 29, 647–661. [Google Scholar] [CrossRef]
- Luostarinen, K.; Möttönen, V. Radial variation in the anatomy of Betula pendula wood from different growing sites. Balt. For. 2010, 16, 209–216. [Google Scholar]
- Dunham, R.A.; Cameron, A.D.; Petty, J.A. The effect of growth rate on the strength properties of sawn beams of silver birch (Betula pendula Roth). Scand. J. For. Res. 1999, 14, 18–26. [Google Scholar] [CrossRef]
- Lachowicz, H.; Sajdak, M.; Paschalis-Jakubowicz, P.; Cichy, W.; Wojtan, R.; Witczak, M. The influence of location, tree age and forest habitat type on basic fuel properties of the wood of the silver birch (Betula pendula Roth.) in Poland. BioEnergy Res. 2018, 11, 638–651. [Google Scholar] [CrossRef] [Green Version]
- Yousefpour, R.; Hanewinkel, M.; Le Moguédec, G. Evaluating the suitability of management strategies of pure Norway spruce forests in the Black Forest area of Southwest Germany for adaptation to or mitigation of climate change. Environ. Manag. 2010, 45, 387–402. [Google Scholar] [CrossRef]
- Čermák, P.; Kolář, T.; Žid, T.; Trnka, M.; Rybníček, M. Norway spruce responses to drought forcing in areas affected by forest decline. For. Syst. 2019, 28, 13. [Google Scholar] [CrossRef] [Green Version]
- Krišāns, O.; Puriņa, L.; Mesters, D.; Kāpostiņš, R.; Rieksts-Riekstiņš, J.; Jansons, Ā. Intra-annual radial growth of European beech-a case study in north easternmost stand in Europe. For. Stud. Metsanduslikud Uurim. 2016, 65, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Cawley, K.M.; Campbell, J.; Zwilling, M.; Jaffé, R. Evaluation of forest disturbance legacy effects on dissolved organic matter characteristics in streams at the Hubbard Brook Experimental Forest, New Hampshire. Aquat. Sci. 2014, 76, 611–622. [Google Scholar] [CrossRef]
- Piri, T. Early development of root rot in young Norway spruce planted on sites infected by Heterobasidion in southern Finland. Can. J. For. Res. 2003, 33, 604–611. [Google Scholar] [CrossRef]
- Piri, T.; Korhonen, K.; Sairanen, A. Occurrence of heterobasidion annosum in pure and mixed spruce stands in Southern Finland. Scand. J. For. Res. 1990, 5, 113–125. [Google Scholar] [CrossRef]
- Baders, E.; Jansons, A.; Matisons, R.; Elferts, D.; Desaine, I. Landscape diversity for reduced risk of insect damage: A case study of spruce bud scale in Latvia. Forests 2018, 9, 545. [Google Scholar] [CrossRef] [Green Version]
- Krišāns, O.; Saleniece, R.; Rust, S.; Elferts, D.; Kapostins, R.; Jansons, A.; Matisons, R. Effect of bark-stripping on mechanical stability of Norway Spruce. Forests 2020, 11, 357. [Google Scholar] [CrossRef] [Green Version]
- Krisans, O.; Matisons, R.; Rust, S.; Burnevica, N.; Bruna, L.; Elferts, D.; Kalvane, L.; Jansons, A. Presence of root rot reduces stability of Norway spruce (Picea abies): Results of static pulling tests in Latvia. Forests 2020, 11, 416. [Google Scholar] [CrossRef] [Green Version]


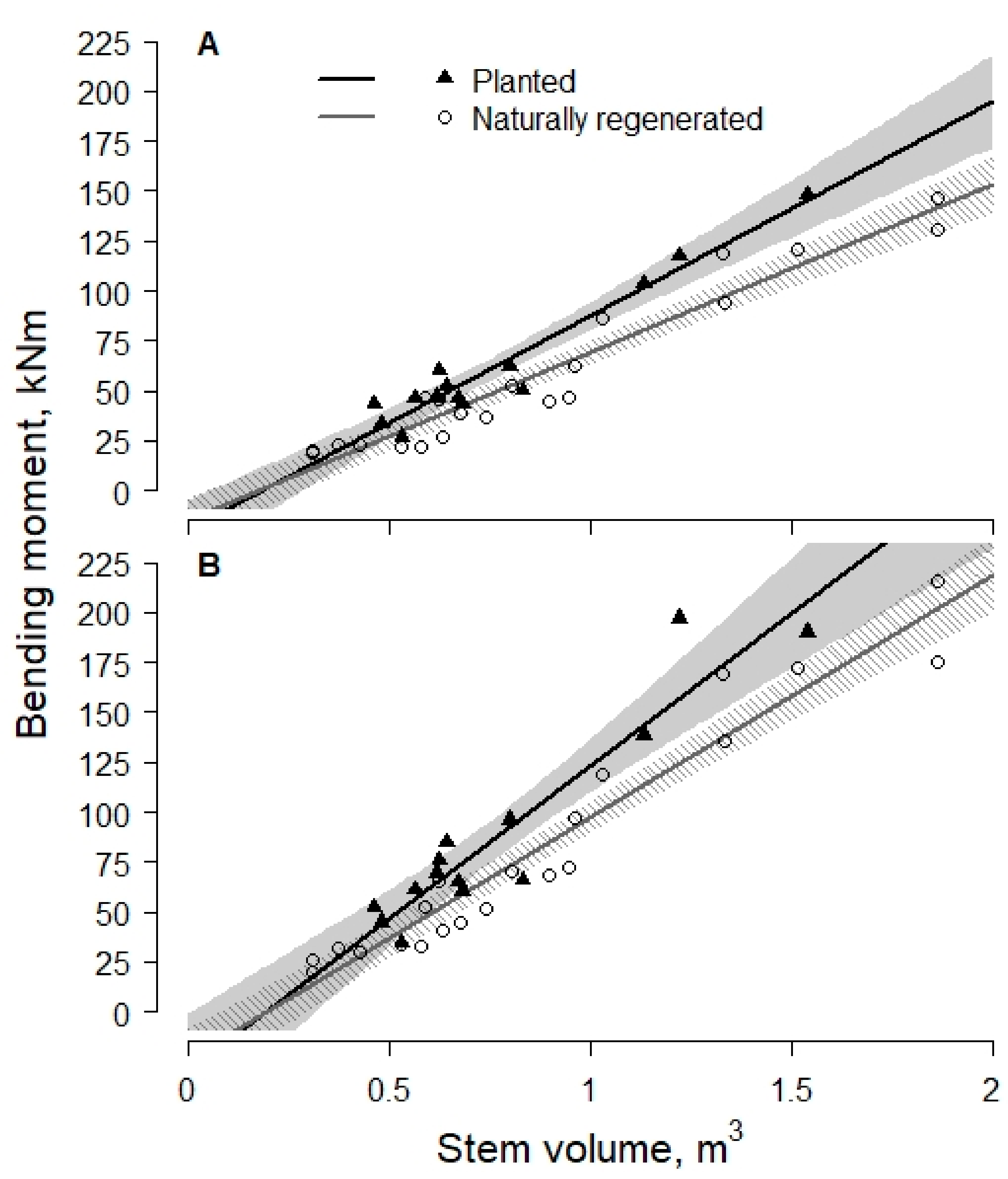{kind=link}
| Site No. | Species Composition (%) | Tree n | DBH (cm) | H (m) | H/DBH | Vstem (m3) | Vroots (m3) | G (m2 ha−1) | θg (%) | ρSoil (kg m−3) |
|---|---|---|---|---|---|---|---|---|---|---|
| Planted | ||||||||||
| 1 | B (64.8) S (34.6) OT (0.6) | 7 | 27.0 ± 3.7 | 29.2 ± 1.1 | 1.10 ± 0.10 | 0.78 ± 0.27 | 1.05 ± 0.30 | 43.3 ± 2.9 | 10.5 ± 1.6 | 1290 ± 43 |
| 2 | B (50.3) S (49.6) OT (0.1) | 7 | 26.5 ± 3.2 | 29.8 ± 1.7 | 1.14 ± 0.11 | 0.76 ± 0.21 | 0.89 ± 0.46 | 49.2 ± 12.1 | 8.8 ± 3.4 | 1275 ± 53 |
| Naturally Regenerated | ||||||||||
| 3 | B (70.2) S (26.8) OT (3.0) | 9 | 25.3 ± 1.7 | 28.8 ± 1.2 | 1.15 ± 0.08 | 0.66 ± 0.09 | 0.94 ± 0.35 | 38.7 ± 3.1 | 7.2 ± 1.4 | 1213 ± 37 |
| 4 | B (65.4) S (34.1) OT (0.5) | 4 | 21.7 ± 2.8 | 22.9 ± 1.9 | 1.06 ± 0.09 | 0.39 ± 0.13 | 1.23 ± 0.58 | 41.1 ± 9.8 | 13.6 | 1306 |
| 5 | B (72.5) S (3.6) P (6.4) O (4.5) A (4.5) OT (8.5) | 8 | 34.0 ± 3.1 | 33.0 ± 1.2 | 0.99 ± 0.10 | 1.36 ± 0.26 | 1.99 ± 0.48 | 33.4 ± 2.6 | 9.2 ± 0.9 | 1260 ± 43 |
| Site No. | BBMPF (kNm∙m−3) | BBMSF (kNm∙m−3) | MOE (GPa) |
|---|---|---|---|
| Planted | |||
| 1 | 74.5 ± 12.7 | 100.0 ± 16.8 | 16.2 ± 3.5 |
| 2 | 83.6 ± 8.5 | 119.9 ± 16.8 | 19.0 ± 2.7 |
| Naturally Regenerated | |||
| 3 | 52.7 ± 7.5 | 73.7 ± 9.7 | 12.0 ± 1.6 |
| 4 | 67.2 ± 8.3 | 81.4 ± 10.8 | 9.7 ± 1.6 |
| 5 | 73.4 ± 8.7 | 105.6 ± 11.0 | 16.1 ± 2.4 |
| BBMPF | BBMSF | MOE | Vroots | |||||
|---|---|---|---|---|---|---|---|---|
| Predictors | χ2 | p-Value | χ2 | p-Value | χ2 | p-Value | χ2 | p-Value |
| (Intercept) | 3.34 | 0.07 | 2.96 | 0.09 | 27.26 | <0.001 | 3.66 | 0.06 |
| Vstem | 157.69 | <0.001 | 128.78 | <0.001 | 8.24 | <0.01 | 6.45 | <0.05 |
| Stand type | 0.53 | 0.47 | 0.51 | 0.48 | 8.84 | <0.01 | 3.23 | 0.07 |
| Vstem by stand type | 5.39 | <0.01 | 4.98 | <0.05 | 2.55 | 0.11 | 2.07 | 0.15 |
| Random Effects | ||||||||
| σ2 | 95.77 | 224.21 | 12.45 | 0.27 | ||||
| τ00 | 30.03 | 79.48 | 0.10 | 0.08 | ||||
| ICC | 0.24 | 0.26 | 0.01 | 0.23 | ||||
| nsite | 5 | 5 | 5 | 4 | ||||
| Observations | 35 | 35 | 34 | 27 | ||||
| Marginal R2 | 0.91 | 0.90 | 0.40 | 0.44 | ||||
| Conditional R2 | 0.93 | 0.92 | 0.41 | 0.57 | ||||
| Site No. | Distance (m) | z-Score | p-Value | Spatial Distribution |
|---|---|---|---|---|
| Planted | ||||
| 1 | 1.6 | 0.519 | 0.603 | Random |
| 2 | 1.7 | 5.647 | <0.001 | Dispersed |
| Naturally | ||||
| 3 | 3.9 | 5.977 | <0.001 | Dispersed |
| 4 | 3.7 | 1.352 | 0.176 | Random |
| 5 | 4.5 | 9.122 | <0.001 | Dispersed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krišāns, O.; Matisons, R.; Vuguls, J.; Bāders, E.; Rust, S.; Elferts, D.; Saleniece, R.; Jansons, Ā. Regularly Planted Rather Than Natural Understory of Norway Spruce (Picea abies H. Karst.) Contributes to the Individual Stability of Canopy Silver Birch (Betula pendula Roth.). Forests 2022, 13, 942. https://doi.org/10.3390/f13060942
Krišāns O, Matisons R, Vuguls J, Bāders E, Rust S, Elferts D, Saleniece R, Jansons Ā. Regularly Planted Rather Than Natural Understory of Norway Spruce (Picea abies H. Karst.) Contributes to the Individual Stability of Canopy Silver Birch (Betula pendula Roth.). Forests. 2022; 13(6):942. https://doi.org/10.3390/f13060942
Chicago/Turabian StyleKrišāns, Oskars, Roberts Matisons, Jānis Vuguls, Endijs Bāders, Steffen Rust, Didzis Elferts, Renāte Saleniece, and Āris Jansons. 2022. "Regularly Planted Rather Than Natural Understory of Norway Spruce (Picea abies H. Karst.) Contributes to the Individual Stability of Canopy Silver Birch (Betula pendula Roth.)" Forests 13, no. 6: 942. https://doi.org/10.3390/f13060942