
Roost Selection in Relation to a Patchy, Mosaic Management Burn by a Threatened Clutter-Adapted Bat


Forest Science, NSW Department of Primary Industries, Locked Bag 5022, Parramatta, NSW 2124, Australia
*
Author to whom correspondence should be addressed.
Forests 2022, 13(8), 1327; https://doi.org/10.3390/f13081327
Submission received: 19 July 2022
/
Revised: 12 August 2022
/
Accepted: 16 August 2022
/
Published: 19 August 2022
(This article belongs to the Special Issue Effects of Forest Management Practices on Bat Habitat and Community Structure)
Abstract
:Fire is a major disturbance for forests and its impacts can be complex, influenced by a range of factors including fire severity and frequency. Changes to global climate have increased the frequency and lengthened the window for wildfires. Anthropogenic fires are now commonly used to try to mitigate the risk and spread of wildfires or for ecological purposes, yet it is unclear how many flora and fauna species respond to this lower severity and more patchily distributed treatment. We assessed day-roost selection by a threatened narrow space bat, Nyctophilus corbeni, after a management burn left a mosaic of unburnt forest, low severity (ground scorch) burnt and higher severity (midstorey to crown scorch) burnt forest. Radio-tracking was used to identify day-roosts of 11 individuals (8 lactating females and 3 males) during the maternity season. Characteristics of day-roost trees (n = 42 trees) were similar for males and lactating females and were comparable to other day-roosts used elsewhere in the broader study area, with bats selecting moderately sized (23–24 cm dbhob) dead buloke (Allocasuarina luehmannii) with hollows. However, roost selection at the scale of the neighborhood varied by sex and was random for lactating females. Selection of the post-burn mosaic by males was non-random at all scales of assessment greater than a 100 m neighborhood, with bats selecting areas burnt by low-severity fire at these scales, but avoiding areas of higher severity fire. Locally, there were ~14 more hollow trees per ha surrounding roosts in areas burnt by low severity fire than in the unburnt forest, whereas forest burnt by higher severity fire had on average ~8 fewer hollow trees per hectare. Our study confirmed that dead buloke with hollows is a key resource for N. corbeni that should be prioritized for retention, particularly when identifying areas to offset habitat loss. Patchy management fires appear to be compatible with roosting habits of breeding N. corbeni, provided hollow resources are not impacted by fire. Management burns may promote roosting habitat for male bats, but areas of higher severity burns need to be minimized. Further research is needed to assess the use of the post-burn mosaic for nightly movements, including foraging. The effects of repeated burning on hollow availability and roost selection by N. corbeni should be investigated to identify suitable intervals between fires for this threatened species.
1. Introduction
Forests are dynamic ecosystems that are impacted by a range of disturbances, both natural and human-induced. Fire is a major disturbance to forests and many other ecosystems [1,2]. Naturally occurring fires (‘wildfires’ or ‘bushfires’) that are ignited by processes such as lightning strikes can affect extensive areas of forest, though the impacts of these fires can be complex and dependent on the interaction of multiple factors, such as fire severity and the existing fire regimes (e.g., time since last fire). A changing climate has resulted in an increase in global temperatures, which has led to more wildfires and longer wildfire seasons [3,4,5]. This is particularly relevant in Australia, the most bushfire-prone of all the continents [6].
To potentially mitigate the risk of devastating wildfires, anthropogenic fires (hereafter ‘management burns’) are used to reduce forest fuel loads or to create firebreaks that aim to limit the spread of wildfire. Management burns are also used for ecological purposes, such as to control weeds and to create a desired ground and understorey cover. As such, these fires tend to be lower severity and patchy in distribution. Despite the common use of fire for land management, it is unclear how many flora and fauna species respond to this treatment. This is particularly true for bats which are important components of forests and comprise almost 20% of the world’s extant mammal fauna, and almost 30% of the Australian mammal fauna [7,8].
Globally, a number of studies have assessed the responses of bats to both wildfire and anthropogenic burns (see [9,10,11]). Fire directly affects bats via mortality, exposure to toxic gases, and burns, and indirectly by altering roosting and foraging habitat and the availability of insect prey. Indirect effects may be positive, neutral, or negative. For example, the activity of some bat species/guilds increases after prescribed fire or wildfire, while that of other species declines or does not change (e.g., [12,13,14,15,16,17,18]).
Few studies have assessed roost selection by bats in relation to prescribed fire. In North America, an assessment of the response of bats to prescribed fire revealed potential benefits to bats via the creation of additional roosting habitat [19,20] or by enlarging existing or creating new canopy gaps [21]. In Australia, no studies have assessed roosting preferences or selection by bats following prescribed fires.
The south-eastern long-eared bat (Nyctophilus corbeni) is a narrow-space bat species listed nationally as a threatened species in Australia (Vulnerable, Environment Protection and Biodiversity Conservation Act 1999) and in New South Wales (Vulnerable, Biodiversity Conservation Act 2016). The species avoids roosting in commercially thinned stands where thinning is extensive but selects old regrowth, whereas logged and mechanically thinned (where machinery is used to ‘chop and roll’ along narrow alleys) stands are used in proportion to availability for roosting [22,23]. A study in the Mallee systems of Victoria revealed that N. corbeni roosted in hollows of dead stems, in areas with abundant hollows that were usually long unburnt [24], suggesting the species may be sensitive to fire. No maternity roosts were located in that study and it is unknown whether roosting behavior is similar in other parts of the distribution of N. corbeni.
In this study, we assessed roost selection by N. corbeni during the maternity season 20 months after a management burn that created a patchy mosaic of unburnt forest and forest burnt by low severity (ground scorch) and higher severity (midstorey to crown scorch) fire. We predicted that the species would avoid roosting in burnt areas if these areas were associated with a lower abundance of hollow trees, particularly buloke (Allocasuarina luehmannii), that the species prefers for roosting in the study area [22,23].
2. Materials and Methods
2.1. Study Area
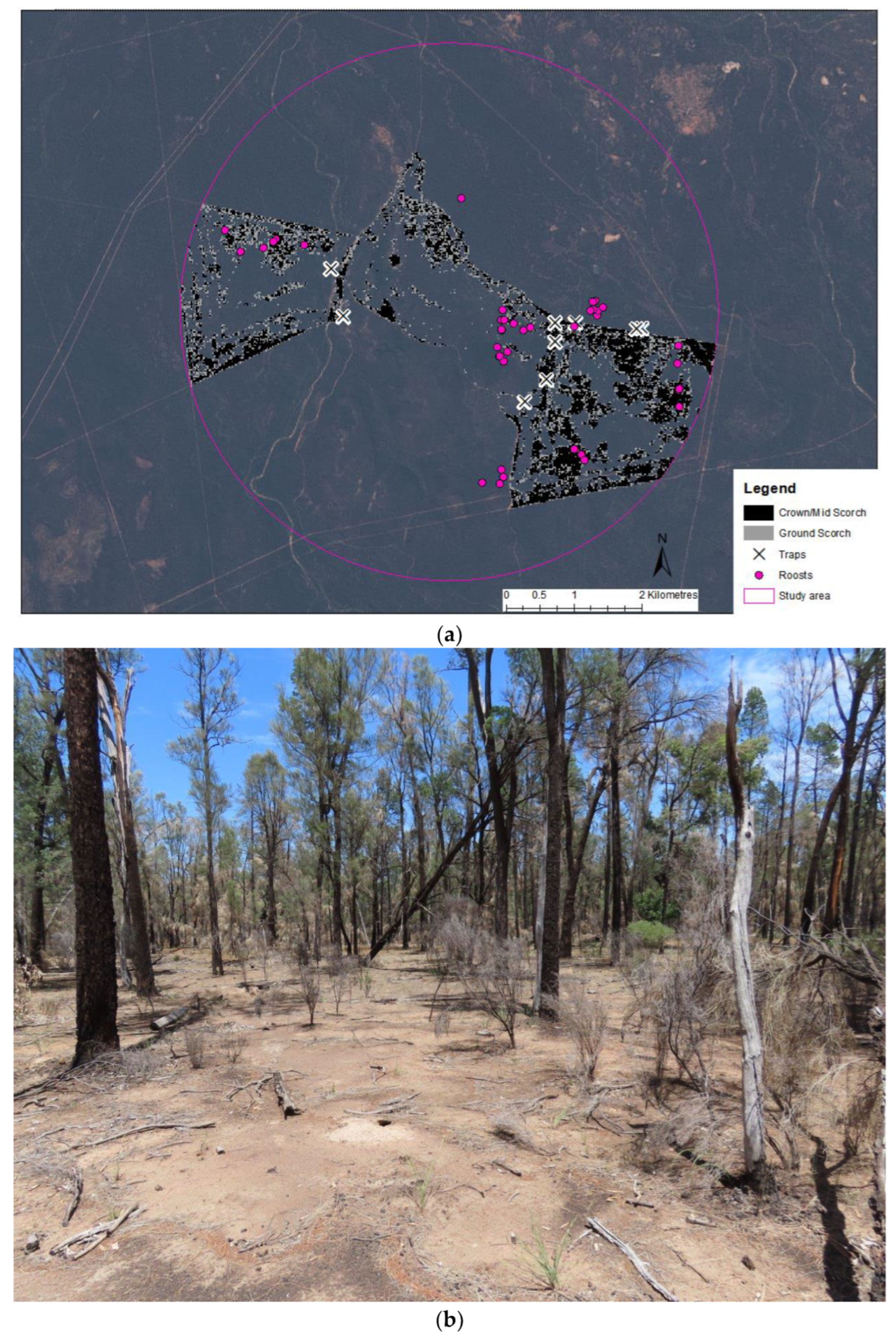
The study was carried out at the Australian Wildlife Conservancy fenced site in the Pilliga, NSW. The site contains 35,632 ha of vegetation that is dominated by cypress pine (Callitris glaucophylla) and buloke, with ironbarks (Eucalyptus crebra and E. sideroxylon) also present. The area is managed to exclude introduced herbivores and predators. In April 2018, a small management burn (577 ha) was undertaken to reduce the risk of wildfire and to promote a mosaic of vegetation structure (Figure 1a). The burn was variable in severity, with 224 ha mapped as ground scorching (Figure 1b) and 353 ha considered higher severity that burnt the midstorey and/or crown (Figure 1c).
2.2. Trapping and Radio-Tracking
Bats were trapped at nine locations spread among areas that were burnt over 29 harp trap-nights in November 2019 (maternity season). Traps were mostly located on tracks adjacent to burnt areas, with some traps also placed in vegetative clutter off-track. To improve trapping efficiency, an acoustic lure (Autobat—[25]) playing a mixture of Nyctophilus-specific and generic European bat social calls was used at a subset of sites. Traps were cleared once prior to midnight and in the morning and any captured bats were removed from traps, identified to species, sexed, and released from the point of capture or fitted with a radio-transmitter prior to release.
In all, 11 N. corbeni (8 lactating females and 3 males) were fitted with VHF transmitters (LB-2X, 0.27 g; Holohil, Carp, ON, Canada). The mass of transmitters represented 1.7% of the mean body mass of N. corbeni. Transmitters were glued between the shoulder blades of bats using Urobond (Urocare) and released 20–60 min later. Bats fitted with transmitters (hereafter, ‘tagged bats’) were radio-tracked for up to 10 days, with day roosts located by ‘homing in on the signal’. Limited nighttime tracking was undertaken to confirm transmitters were operational when bats could not be located during the day and to assist in locating potential roost areas. Fine-scale nightly movements of bats were the focus of a separate study using GPS tags.
All trapping, animal handling, and radio-tracking were carried out under Scientific License (SL100623) and Animal Ethics Research Authority 29/19–22 issued by DPI Forestry Corporation NSW Animal Ethics Committee.
2.3. Roost Tree Assessments
Once roost trees were located, an assessment was made to characterize the roost tree and neighboring trees in the local environment. For each roost tree, we recorded the tree species, whether it was living or dead, diameter at breast height over bark (dbhob, 1.3 m), canopy cover, senescence score (0–8, following [26]), a hollow score (0–3; 0 = no hollows, 1 = at least one hollow, 2 = two hollows, 3 = three or more hollows) and whether the tree had been burnt. If the bat was visible, we recorded the roost type (bark, fissure, hollow). If it was not possible to see the bat, the roost type was recorded as ‘unknown’. We repeated the process (excepting roost type) for the nearest neighboring tree (>10 cm dbhob) in each of four quadrants surrounding the roost tree. The density of hollow trees surrounding roosts in unburnt forest and forest burnt by low and higher severity fire was calculated using the point-quarter technique to estimate tree density and then multiplying this value by the proportion of hollow trees that comprised trees neighboring roost trees [27].
2.4. Landscape-Level Assessments of Roosts in Relation to Fire
Although N. corbeni was previously reported to make movements of up to 3.96 km (Law et al. 2016), a concurrent study on nightly movements using GPS tracking revealed that the species was capable of significantly greater movements outside of the maternity season (Unpublished data—authors). This contrasts with the minimum extent of local areas (3.0 ± 1.7 ha or within a radius of ~100 m) used by bats for roosting in the current study. Previous studies in the broader study area have also identified bats roosting within a small local area (mean distance between consecutive roosts = ~270 m for females and ~1200 m for males; [22]). To define the area over which to calculate the availability/extent of unburnt forest, low severity burnt and higher severity burnt forest, a series of different sized neighborhoods were generated and centered on each unique roost tree. Neighborhood sizes were 100 m (3 ha), 250 (20 ha) m, 500 m (79 ha), and 1 km (314 ha). We used GIS (ArcMap, ESRI) and a spatial layer that mapped the management burn (Unpublished data—AWC) to calculate the area of unburnt forest, low severity burnt and higher severity burnt forest within each neighborhood to provide a measure of availability.
The locations of all roost trees were then overlaid onto the mapped fire layer and assigned to one of three mapped categories (i.e., unburnt, ‘low severity’ fire, and ‘high severity’ fire) and confirmed based on descriptions of the severity of fire at the roost site. The proportion of points in each category was then tabulated.
2.5. Data Analyses
Chi-square goodness of fit tests were used to assess whether proportional use of different tree species and living and dead trees differed from their local availability. Since data collected for roost tree characteristics were not normally distributed, a series of Wilcoxon ranked sum tests was used to assess whether characteristics (dbhob, height, canopy cover, senescence score, and hollow score) of roost trees were significantly different from locally available trees. A canonical analysis of principal components (CAP) was used to identify the characteristics of trees (living and dead) selected for roosting by N. corbeni relative to locally available trees (living and dead). Generalized linear mixed models (GLMMs) were used to assess whether roost trees used by male and lactating female N. corbeni were located in each fire category as would be expected based on the availability of each category within the different scales of study area (i.e., 100 m, 250 m, 500 m, and 1 km neighborhoods). For each model, individual ID was included as a random effect and the interaction of fire category and use/availability as fixed effects. The response variable was the proportion of roost trees that fell in each fire severity category (unburnt, low severity, and higher severity burn) or the extent of each fire severity category within different sized neighborhoods of each roost tree. A Tweedie distribution with a log link was fitted since the data had a point mass at zero. Models were generated using the glmmTMB function in the glmmTMB package [28] in R [29].
3. Results
3.1. Bat Captures
In all, 393 bats representing 11 species were captured (Table 1). Of these, Vespadelus vulturnus, Nyctophilus gouldi and Chalinolobus gouldii were the most commonly trapped species. A total of 17 Nyctophilus corbeni (13 lactating females and 4 males) were captured.
3.2. Roost Tree Use and Characteristics
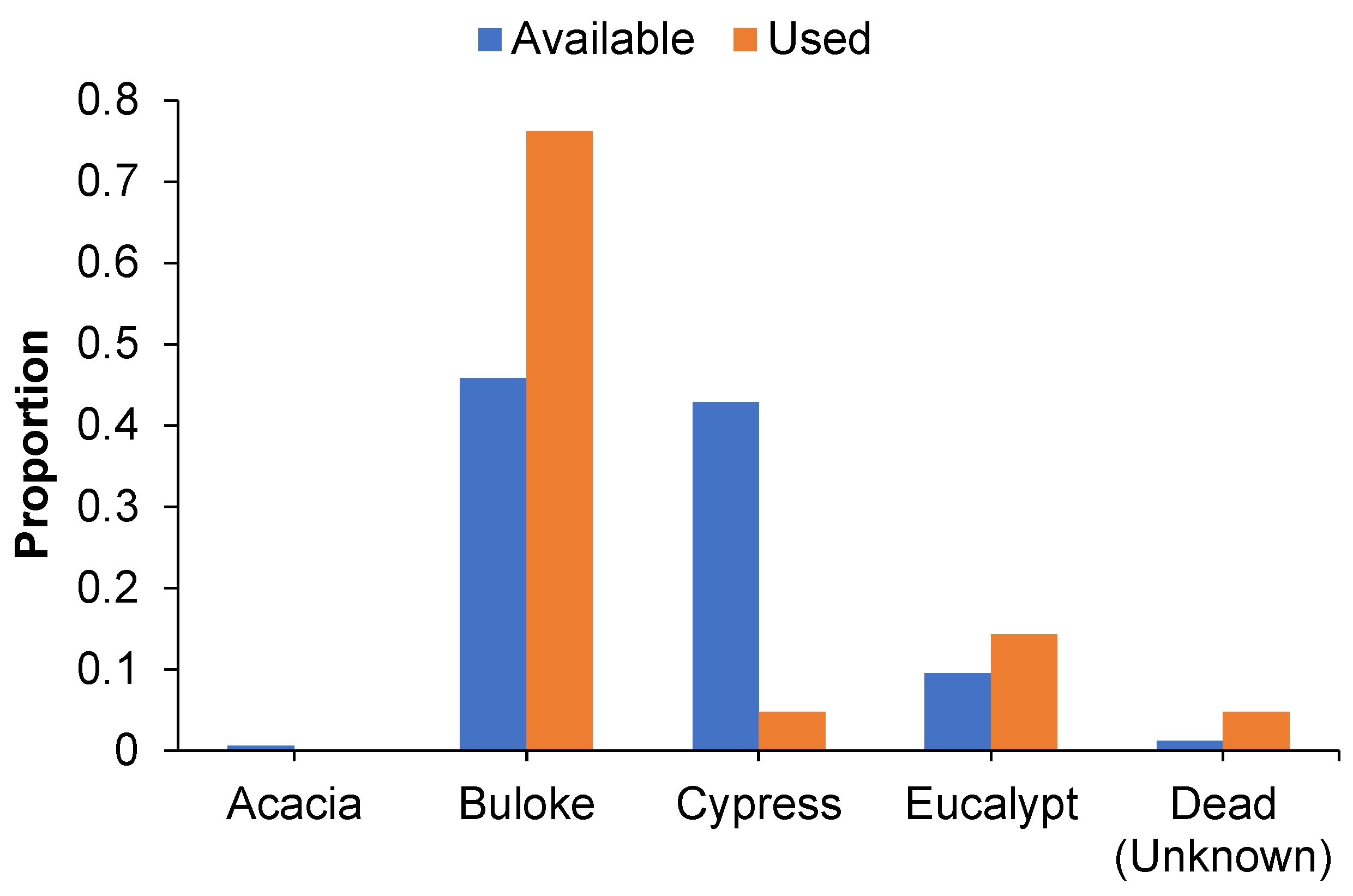
In all, 79 roost tree days were located representing 42 unique roost trees. Roost tree use differed significantly from the availability of species/groups in the local landscape (χ2 = 28.417, p < 0.001; Figure 2). Buloke was the most commonly used roost tree species, with use greater than expected based on the local availability of this species (508 stems > 10 cm dbhob per ha). Cypress pine was used less than expected based on availability in the local landscape (475 stems > 10 cm dbhob per ha) whereas eucalypts (106 stems > 10 cm dbhob per ha) and dead (unidentified) (13 stems > 10 cm dbhob per ha) were used in proportion to their availability. This pattern was similar for male and lactating female N. corbeni, with 67% and 83% of roost trees, respectively, represented by buloke. Cypress pine represented < 6% of roost trees irrespective of sex, whereas eucalypts made up 13%–17% of roost trees for females and males, respectively. Approximately 11% of male roost trees could not be allocated to a species as the trees were dead and could not be confidently identified to a species.
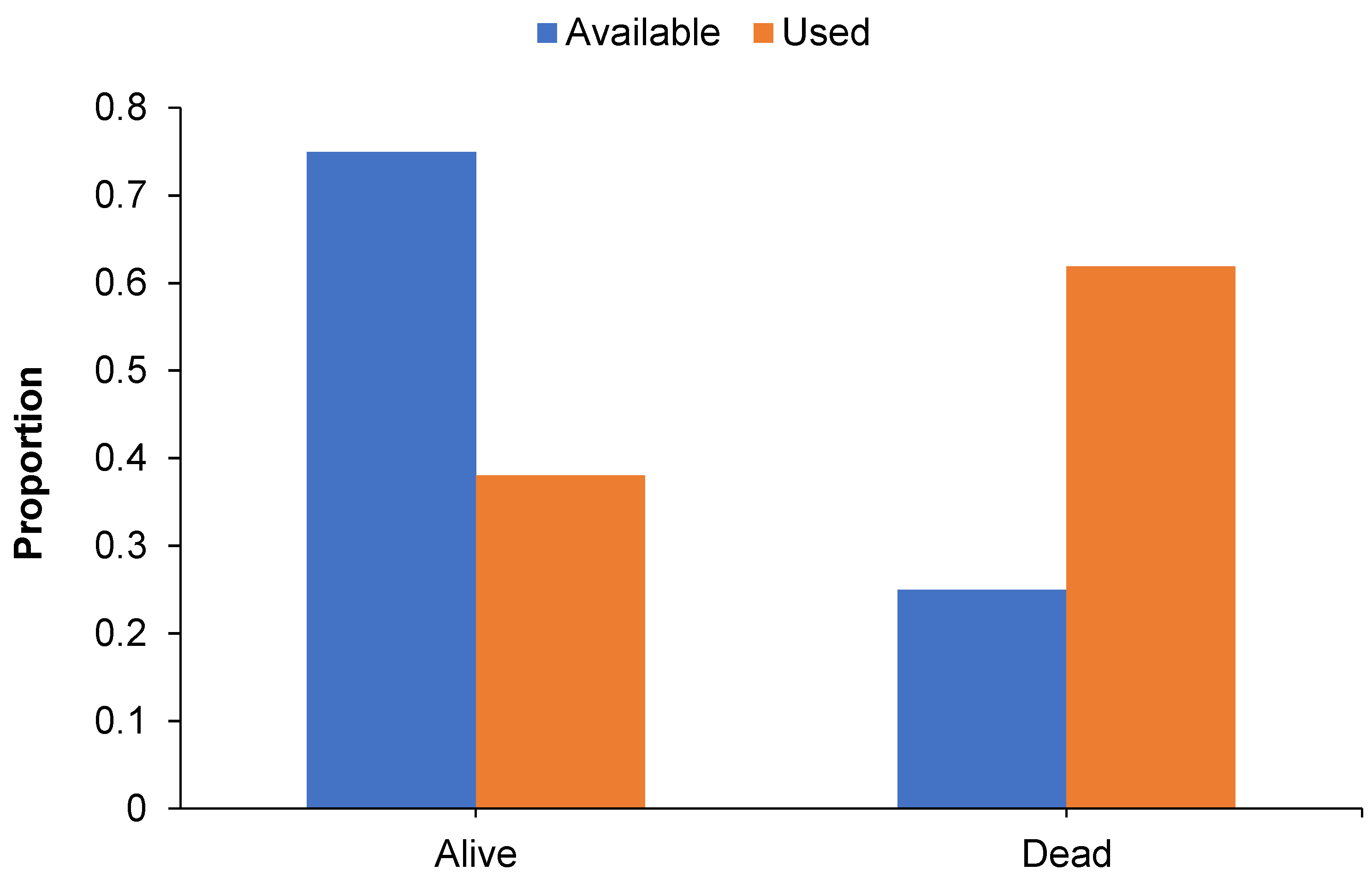
The use of living and dead trees for roosting differed significantly from local availability (χ2 = 30.508, p < 0.001; Figure 3). Roost trees were generally dead (62% of all roost trees), with the use of dead trees significantly greater than availability in the local landscape (270 stems > 10 cm dbhob per ha). Conversely, living trees (38% of all roost trees) were used less than expected based on availability in the local landscape (838 stems > 10 cm dbhob per ha). The use of dead trees for roosting by males (72% of roost trees) was greater than females (54%).
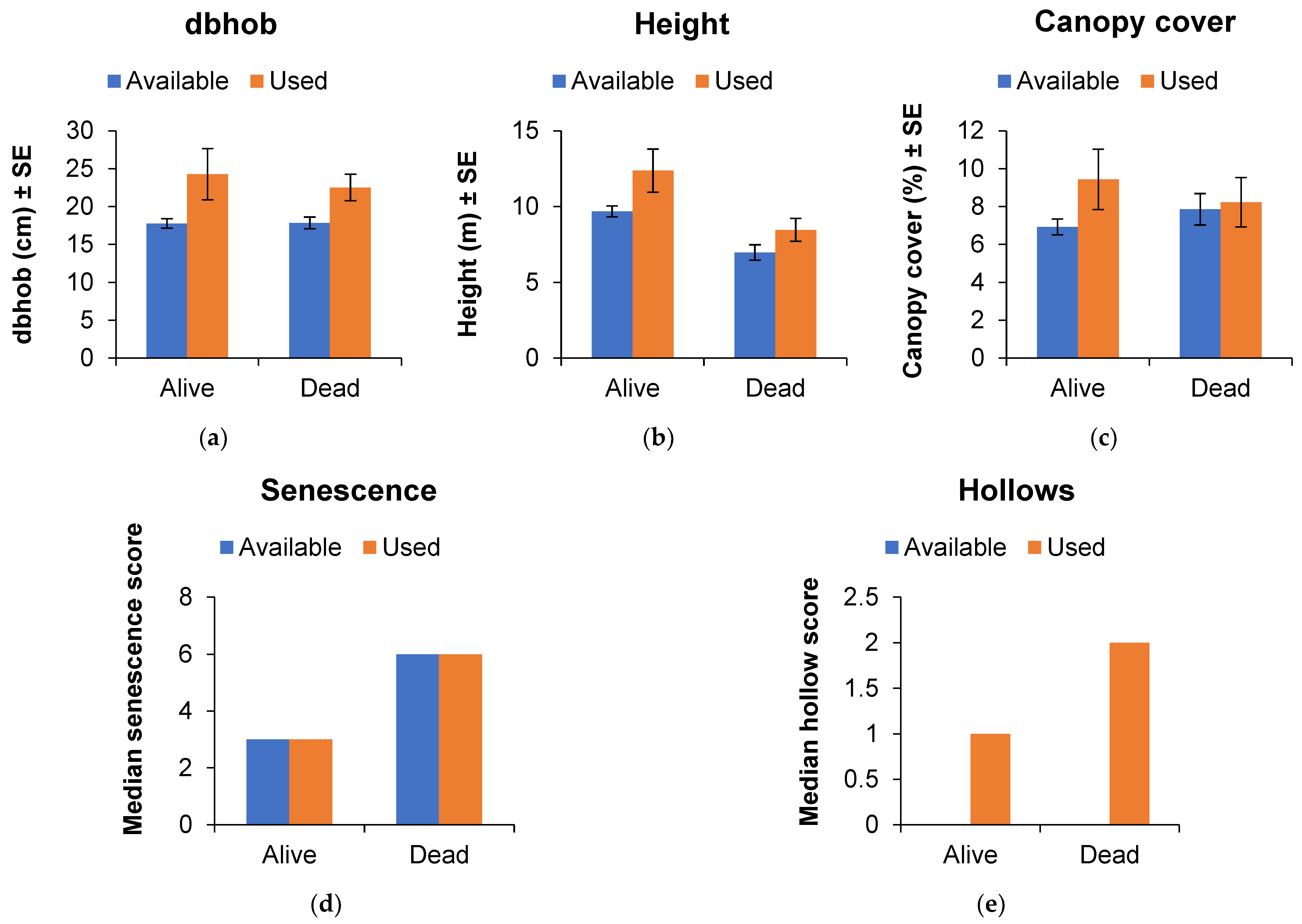
Most characteristics of roost trees differed significantly from locally available trees, with the exception of the senescence score (Figure 4a–e). The diameter of trees used for roosting was relatively small (~23 cm dbhob) but was significantly larger than locally available (~18 cm dbhob) trees irrespective of whether they were living (W = 107, p = 0.044, n = 16) or dead (W = 269, p = 0.017, n = 26). Similarly, roost trees were taller (8–12 m) than locally available trees (7–10 m) irrespective of whether they were living (W = 73.5, p = 0.007 n = 16) or dead (W = 258.5, p = 0.009, n = 26). Canopy cover over roost trees was low (8%–9%) but was greater than cover of locally available trees (7%–8%) irrespective of whether they were living (W = 50.5, p = 0.018, n = 16) or dead (W = 99, p = 0.025, n = 26). Roost trees contained at least 1–2 hollows on average whereas no hollows were recorded for locally available trees irrespective of whether they were living (W = 78, p = 0.002, n = 16) or dead (W = 231, p < 0.001, n = 26). Senescence score for dead roost trees did not differ from locally available dead trees (W = 6, p = 0.082, n = 26), as was the case for the living roost trees and locally available live trees (W = 52, p = 0.082, n = 16). Roost tree characteristics for male and female N. corbeni were similar (male and female: dbhob= 22 ± 3 cm and 24 ± 2 cm; height = 9 ± 1 m and 11 ± 1 m; canopy cover = 8.7 ± 1.7% and 8.7 ± 1.3%; senescence score median = 6 and 6; hollow score median = 1 and 1).
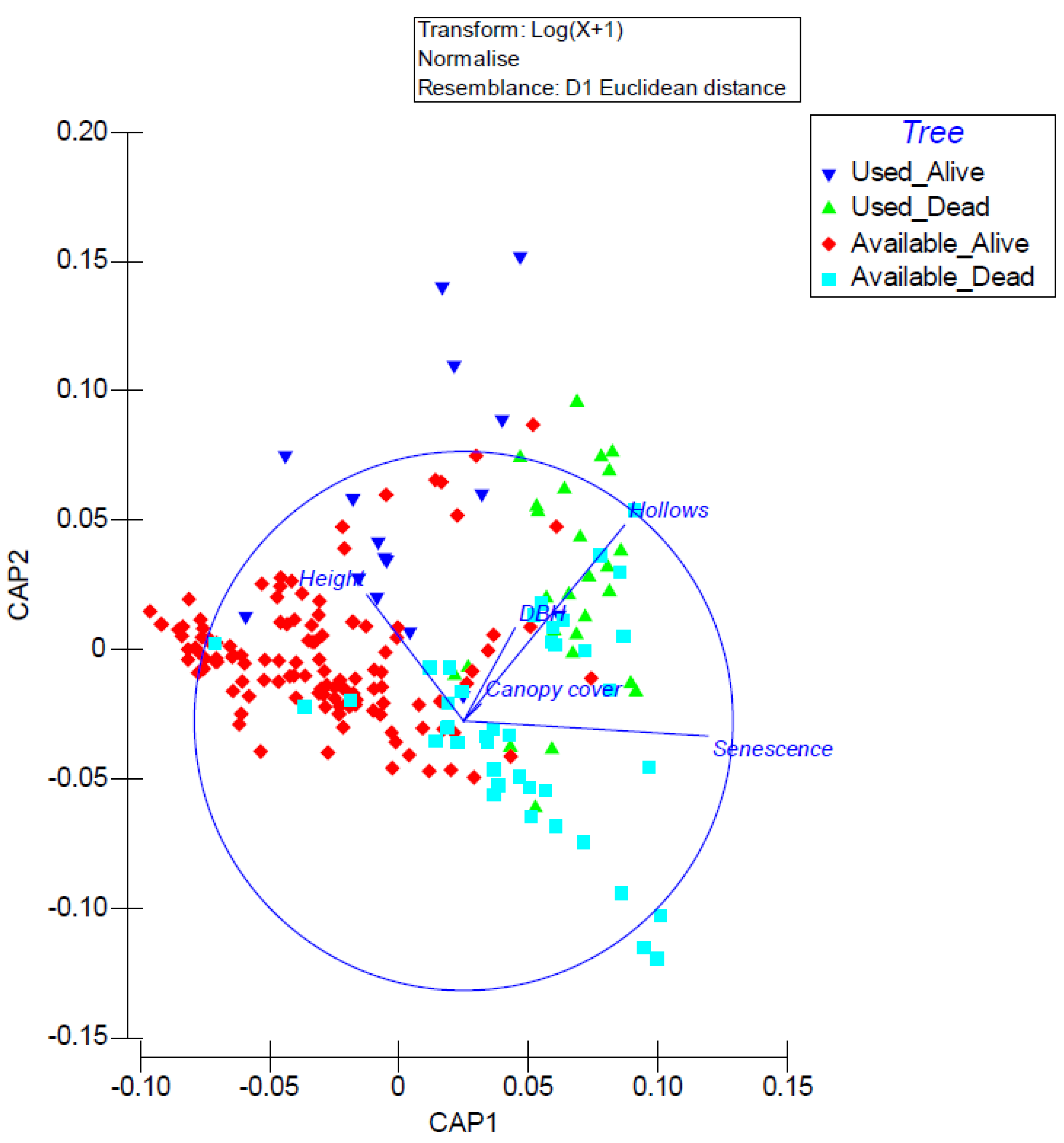
A canonical analysis of principal coordinates revealed that trees used for roosting by N. corbeni were positively associated with hollow abundance and this was particularly the case for dead trees with hollows (Figure 5). Living and dead trees not used for roosting tended to not have hollows and were smaller than trees used for roosting (though these were still relatively small; ~23 cm on average).
Hollows (59.5%) and under bark (21.4%) were the most common types of roosts used by N. corbeni, representing ~80% of all roosts. Use of fissures for roosting was limited (~2%) and all other roosts could not be confidently assigned to a hollow type category. Hollows represented 63% of roosts used by lactating females, whereas 17% of roosts were under bark and the remaining could not be verified (i.e., a bat may have been in a hollow/fissure or in a hollow/under bark). Most roosts used by males were hollows (55% of roosts) followed by under bark (28%) or in a fissure (6%). The remaining male roosts could not be assigned to a roost type with any confidence.
3.3. Use of the Post-Burn Mosaic for Roosting
In all, 42 unique roost trees were located. Of these, 26 trees (62%) were in unburnt and 16 in burnt parts of the landscape (Figure 1 and Figure 6). A single roost tree was lightly burnt.
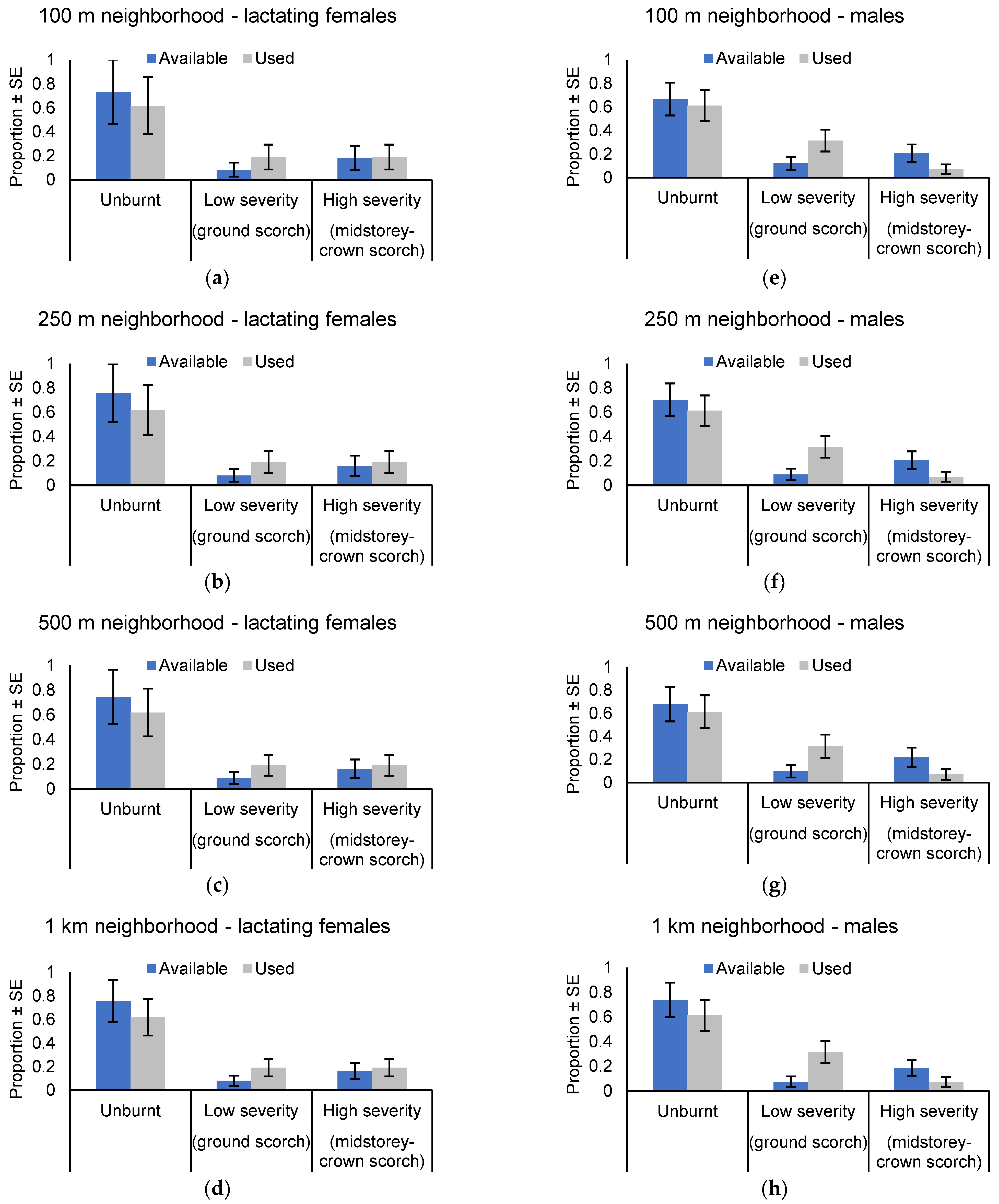
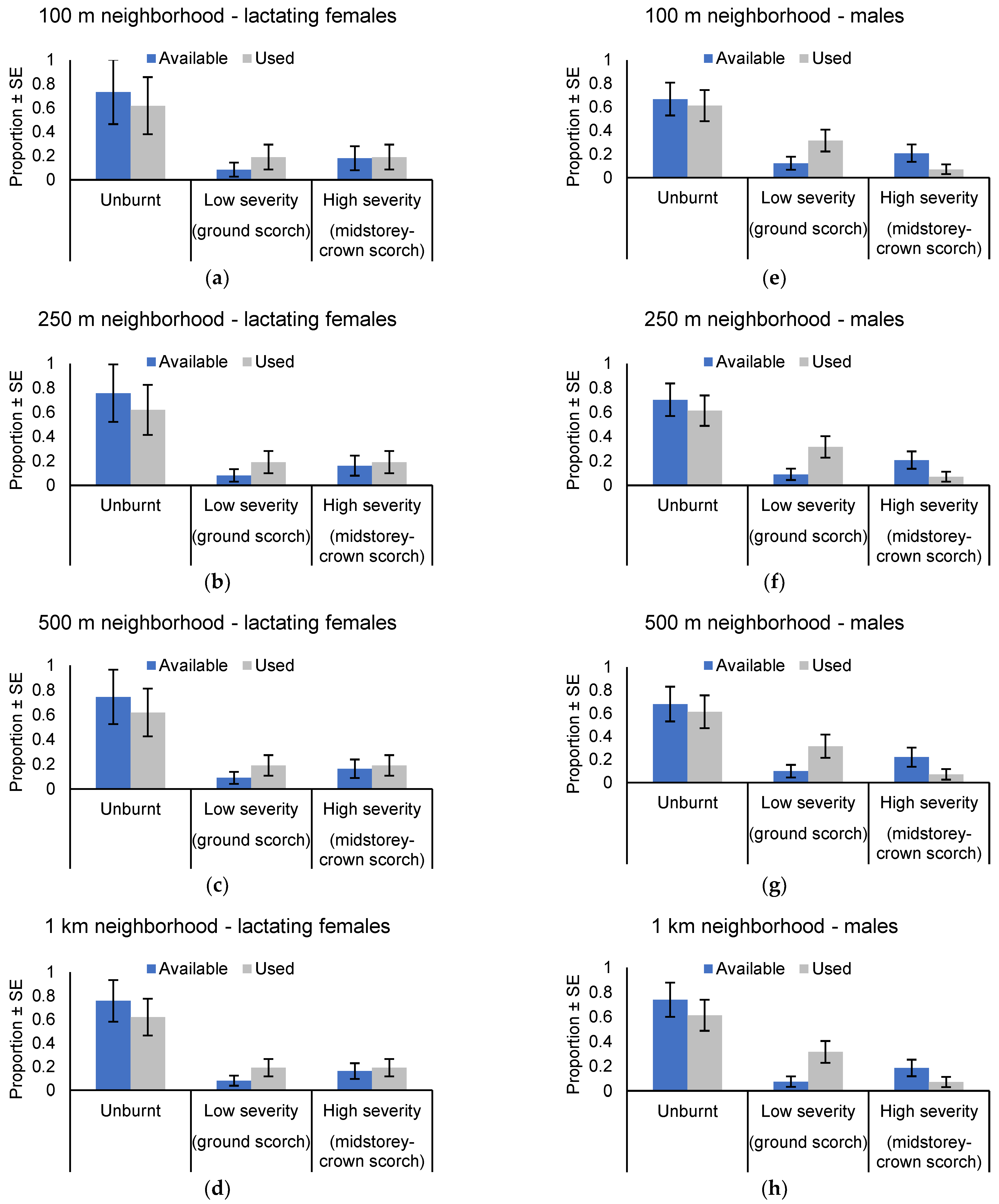
Roost selection varied by sex. In all, 24 unique roost trees were located for lactating females. Of these, 16 were located in unburnt forest (67%) while four (17%) trees each were located in areas burnt by low severity and higher severity fire. Although use of areas burnt by low severity fire was ~2 times greater than availability, the use of the post-burn mosaic for roosting was proportional to availability at all scales of assessment (100 m neighborhood: χ2 = 0.908, p = 0.635; 250 m neighborhood: χ2 = 1.336, p = 0.513; 500 m neighborhood: χ2 = 1.310, p = 0.520; 1 km neighborhood χ2 = 2.087, p = 0.352: Figure 7a–d).
Eighteen unique roost trees were located for male N. corbeni. Of these, 10 were located in unburnt forest (56%), whereas six (33%) and two (11%) trees were located in areas burnt by low severity and higher severity fire, respectively. The use of the post-burn mosaic for roosting was not random for scales of assessment >100 m neighborhoods (100 m neighborhood: χ2 = 5.619, p = 0.060; 250 m neighborhood: χ2 = 7.340, p = 0.025; 500 m neighborhood: χ2 = 5.685, p = 0.050; 1 km neighborhood χ2 = 7.619, p = 0.022: Figure 7e–h). Nyctophilus corbeni males selected areas of low severity fire for roosting at these scales, with the use of this part of the landscape ~3–5 times greater than expected based on availability. Male bats avoided roosting in areas burnt by higher severity fire, whereas the use of unburnt forest was proportional to availability.
The density of hollow-bearing trees in areas burnt by low severity fire (64 ± 2 stems per ha) was greater than in unburnt (49 ± 1 stems per ha) and higher severity fire-affected areas (42 ± 2 stems per ha), indicating a higher density of hollow trees in areas of low severity fire.
4. Discussion
This study assessed roost selection of N. corbeni in relation to a patchy, mosaic management burn during the maternity season, albeit with low sample sizes (n = 11 individuals). Nonetheless, characteristics of roost trees were similar between sexes and to those previously recorded for the species elsewhere in the broader study area [22,23], with bats selecting for larger (>diameter and taller) dead buloke with greater canopy cover and hollows than locally available trees. Roost selection in the post-management burn mosaic varied by sex and was random for lactating females, but non-random for males at all scales greater than the 100 m neighborhoods, with bats selecting areas burnt by low-severity fire at these scales, but avoiding areas of higher severity fire. The extent of roost networks detected in this and other studies [22,23] suggest that a more localized scale of assessment of roost availability is appropriate for N. corbeni. Together, these results suggest that a single patchy management burn is compatible with roosting habits of breeding N. corbeni, provided hollow resources are not impacted by fire. Management burns may promote roosting habitat for male bats, but areas of higher severity burns need to be minimized.
Maternity roost tree characteristics identified for N. corbeni in this study were consistent with those described for the species in previous studies in the Pilliga [22,23]. The use of buloke for roosting was disproportionately greater than its availability in the local landscape, as was the case for use of dead trees with hollows. Retention of clumps of larger buloke (>23 cm dbhob) within the distribution of N. corbeni should be a priority, particularly when identifying areas to offset habitat loss and during timber harvesting operations. Previous research has also identified that areas with a high density of dead buloke are particularly important for roosting N. corbeni [22,23].
Fire is considered a key threat for N. corbeni, with bushfires, fuel reduction burns and frequent burning regimes thought to have a detrimental impact on the species [30]. The location of roost sites in the post-burn mosaic (19 months post-burn) allowed us to assess whether N. corbeni avoided roosting in burnt areas in preference to unburnt patches as has been found for non-reproductive bats in Victorian Mallee [24]. Accounting for the availability of unburnt and fire-affected (both, low severity fire and higher severity fire) areas at different scales of assessment in the study area, roost selection varied by sex. Although areas with low severity fire were used for roosting ~2 times greater than expected based on availability, the selection of the post-burn mosaic was random for lactating females. Use of these areas by male N. corbeni was ~3–5 times greater than expected based on the proportional availability of low severity fire affected areas when assessed beyond the 100 m neighborhood, though a similar trend was also evident at the 100 m neighborhood. Although few field studies have been undertaken, other bat species have also been recorded roosting in burnt forest more often than in unburnt forest following a management burn, with bats using trees and stags that contained more microhabitats for roosting [20]. Maternity colonies of the same species (northern myotis, Myotis septentrionalis) in management burnt forests were associated with larger overall canopy gaps than roost trees within unburnt forest [21]. It is unclear why bats in our study were selecting (males) or showed a preference for (lactating females) forest that had experienced low severity fire for roosting. One possible reason is that low severity fire and associated heat can cause the bark to peel from buloke, creating more roosting opportunities and a network of roosts that can be used by bats. However, only a single roost was located under bark in areas burnt by low severity fire. Locally, the density of hollow trees was higher surrounding roosts in areas burnt by low severity fire and this may explain a preference for roosting in these areas. It is unknown whether this disparity in local hollow tree density occurred prior to the management burn or was created by the fire. Furthermore, areas of forest burnt by higher severity fire had on average ~8 fewer hollow trees per hectare than unburnt forest, although we acknowledge that we lack pre-fire data on hollow tree density. There was evidence of collapse for hollow trees in these areas and while previous assessment of the rates of collapse following low-intensity management burns revealed that 26% of hollow eucalypt trees collapsed [31], it is unknown how fire affects collapse rates for dead buloke with hollows.
The patchy nature of the management burn in our study, both spatially and with respect to severity would likely not have the same impact on roosting as more severe and homogeneous fires. In a separate study assessing the nightly movements of N. corbeni using GPS transmitters, three roosts used by a single female N. corbeni outside of the maternity season were located in dead standing ironbarks or cypress pine in an area of Cumbil/Euligal/Coomore Creek State Forest that was extensively (>90,000 ha) burnt by an intense wildfire 12 years earlier (Unpublished data—Authors). A second female N. corbeni also roosted in this area, though its roost tree could not be located. This suggests that these extensively burnt areas can be used for roosting by N. corbeni if sufficient hollow resources are present post-fire. Further work is needed to assess whether the species selects or avoids roosting in areas burnt by wildfire, particularly following multiple fires that may reduce hollow resources significantly [32,33], though fire frequency can be positively associated with the abundance of hollow-bearing trees in unlogged dry sclerophyll forests [34].
5. Conclusions
Our study confirmed that dead buloke with hollows are a key resource for N. corbeni that should be prioritized for retention, particularly when identifying areas to offset habitat loss or during timber harvesting. Our study, albeit with a low sample size, suggests that the use of a single patchy management burn appears to be compatible with the roosting habits of breeding N. corbeni, provided hollow resources are not impacted by fire. Research is needed to assess the use of the post-burn mosaic for nightly movements, including foraging, and the impact of low-intensity fire on the density of buloke with hollows. The effects of repeated burning on hollow availability and roost selection by N. corbeni should be investigated to identify suitable intervals between fires.
Author Contributions
Conceptualization, L.G. and B.L.; methodology, L.G. and B.L.; formal analysis, L.G.; investigation, L.G., B.L., T.B., I.K. and C.O.; writing—original draft preparation, L.G.; writing—B.L., funding acquisition, L.G. and B.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Whitehaven Coal offsets for research on N. corbeni via NSW Department of Planning and Environment, Biodiversity and Conservation Division.
Institutional Review Board Statement
The animal study protocol was approved by the DPI Forestry Corporation NSW Animal Ethics Committee (Animal Ethics Research Authority 29/19–22 approved on 29 August 2019).
Informed Consent Statement
Not applicable.
Data Availability Statement
Given the data indicate locations of roost sites for a threatened species, data will not be made publicly available. However, data may be made available upon reasonable request to the corresponding author.
Acknowledgments
We thank Australian Wildlife Conservancy staff, particularly Wayne Sparrow and Greg Holland, for providing site access and Rod Kavanagh for providing fire severity spatial data. We also thank Todd Soderquist (NSW DPE) for coordinating funding and Patrick Tap (Forestry Corporation NSW) for providing comments on a draft manuscript. The manuscript was considerably improved by comments from two anonymous reviewers.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Krawchuk, M.A.; Moritz, M.A.; Parisien, M.A.; van Dorn, J.; Hayhoe, K. Global pyrogeography: The current and future distribution of wildfire. PLoS ONE 2009, 4, e5102. [Google Scholar] [CrossRef]
- Bradstock, R.A. A biogeographic model of fire regimes in Australia: Current and future implications. Glob. Ecol. Biogeogr. 2010, 19, 145–158. [Google Scholar] [CrossRef]
- Bowman, D.M.; Williamson, G.J.; Abatzoglou, J.T.; Kolden, C.A.; Cochrane, M.A.; Smith, A. Human exposure and sensitivity to globally extreme wildfire events. Nat. Ecol. Evol. 2017, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pyne, S.J. Burning Bush: A Fire History of Australia; Allen & Unwin: Sydney, Australia, 1991; 520p. [Google Scholar]
- Milne, D.J.; Armstrong, M.; Fisher, A.; Flores, T.; Pavey, C.R. Structure and environmental relationships of insectivorous bat assemblages in tropical Australian savannas. Austral Ecol. 2005, 30, 906–919. [Google Scholar] [CrossRef] [PubMed]
- Frick, W.F.; Kingston, T.; Flanders, J. A review of the major threats and challenges to global bat conservation. Ann. N. Y. Acad. Sci. 2020, 1469, 5–25. [Google Scholar] [CrossRef]
- Perry, R.W. A review of fire effects on bats and bat habitat in the eastern oak region. In Proceedings of the 4th Fire in Eastern Oak Forests Conference, Springfield, MO, USA, 17–19 May 2011. [Google Scholar]
- Loeb, S.C. Qualitative synthesis of temperate bat responses to silvicultural treatments—where do we go from here? J. Mammal. 2020, 101, 1513–1525. [Google Scholar] [CrossRef]
- Loeb, S.C.; Blakey, R.V. Bats and fire: A global review. Fire Ecol. 2021, 17, 1–8. [Google Scholar] [CrossRef]
- Inkster-Draper, T.E.; Sheaves, M.; Johnson, C.N.; Robson, S.K. Prescribed fire in eucalypt woodlands: Immediate effects on a microbat community of northern Australia. Wildl. Res. 2013, 40, 70–76. [Google Scholar] [CrossRef]
- Austin, L.V.; Silvis, A.; Muthersbaugh, M.S.; Powers, K.E.; Ford, W.M. Bat activity following repeated prescribed fire in the central Appalachians, USA. Fire Ecol. 2018, 14, 1–11. [Google Scholar] [CrossRef]
- Austin, L.V.; Silvis, A.; Ford, W.M.; Muthersbaugh, M.; Powers, K.E. Bat activity following restoration prescribed burning in the central Appalachian upland and riparian habitats. Nat. Areas J. 2018, 38, 183–195. [Google Scholar] [CrossRef]
- Law, B.; Doty, A.; Chidel, M.; Brassil, T. Bat activity before and after a severe wildfire in Pilliga forests: Resilience influenced by fire extent and landscape mobility? Austral Ecol. 2018, 43, 706–718. [Google Scholar] [CrossRef]
- Law, B.; Kathuria, A.; Chidel, M.; Brassil, T. Long-term effects of repeated fuel-reduction burning and logging on bats in south-eastern Australia. Austral Ecol. 2019, 44, 1013–1024. [Google Scholar] [CrossRef]
- Ancillotto, L.; Bosso, L.; Conti, P.; Russo, D. Resilient responses by bats to a severe wildfire: Conservation implications. Anim. Conserv. 2021, 24, 470–481. [Google Scholar] [CrossRef]
- Broken-Brow, J.; Hitch, A.T.; Armstrong, K.N.; Leung, L.K. Effect of fire on insectivorous bat activity in northern Australia: Does fire intensity matter on a local scale? Aust. J. Zool. 2020, 67, 260–268. [Google Scholar] [CrossRef]
- Boyles, J.G.; Aubrey, D.P. Managing forests with prescribed fire: Implications for a cavity-dwelling bat species. For. Ecol. Manag. 2006, 222, 108–115. [Google Scholar] [CrossRef]
- Lacki, M.J.; Cox, D.R.; Dodd, L.E.; Dickinson, M.B. Response of northern bats (Myotis septentrionalis) to prescribed fires in eastern Kentucky forests. J. Mammal. 2009, 90, 1165–1175. [Google Scholar] [CrossRef]
- Johnson, J.B.; Edwards, J.W.; Ford, W.M.; Gates, J.E. Roost tree selection by northern myotis (Myotis septentrionalis) maternity colonies following prescribed fire in a Central Appalachian Mountains hardwood forest. For. Ecol. Manag. 2009, 258, 233–242. [Google Scholar] [CrossRef]
- Law, B.; Gonsalves, L.; Chidel, M.; Brassil, T. Subtle use of a disturbance mosaic by the south-eastern long-eared bat (Nyctophilus corbeni): An extinction-prone, narrow-space bat. Wildl. Res. 2016, 43, 153–168. [Google Scholar] [CrossRef]
- Law, B.; Gonsalves, L.; Brassil, T.; Hill, D. Does thinning homogenous and dense regrowth benefit bats? Radio-tracking, ultrasonic detection and trapping. Diversity 2018, 10, 45. [Google Scholar] [CrossRef]
- Lumsden, L.; Nelson, J.; Lindeman, M. Ecological Research on the Eastern Long-Eared Bat Nyctophilus timoriensis (South-Eastern Form). A Report to the Mallee Catchment Management Authority; Arthur Rylah Institute for Environmental Research, Department of Sustainability and Environment: Melbourne, Australian, 2018. [Google Scholar]
- Hill, D.A.; Armstrong, K.N.; Barden, P.A. Preliminary assessment suggests that acoustic lures can increase capture rates of Australian echolocating bats. Aust. Mammal. 2014, 37, 104–106. [Google Scholar] [CrossRef]
- Gibbons, P.; Lindenmayer, D.B.; Barry, S.C.; Tanton, M.T. Hollow formation in eucalypts from temperate forests in southeastern Australia. Pac. Conserv. Biol. 2000, 6, 218–228. [Google Scholar] [CrossRef]
- Pollard, J.H. On distance estimators of density in randomly distributed forests. Biometrics 1971, 4, 991–1002. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 97, 378–400. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Duncan, A.M.; Baker, G.B.; Montgomery, N. The Action Plan for Australian Bats; Biodiversity Group Environment Australia: Canberra, Australia, 1999. [Google Scholar]
- Parnaby, H.; Lunney, D.; Shannon, I.; Fleming, M. Collapse rates of hollow-bearing trees following low intensity prescription burns in the Pilliga forests, New South Wales. Pac. Conserv. Biol. 2010, 16, 209–220. [Google Scholar] [CrossRef]
- Salmona, J.; Dixon, K.M.; Banks, S.C. The effects of fire history on hollow-bearing tree abundance in montane and subalpine eucalypt forests in southeastern Australia. For. Ecol. Manag. 2018, 428, 93–103. [Google Scholar] [CrossRef]
- Radford, I.J.; Oliveira, S.L.; Byrne, B.; Woolley, L.A. Tree hollow densities reduced by frequent late dry-season wildfires in threatened Gouldian finch (Erythrura gouldiae) breeding habitat. Wildl. Res. 2021, 48, 511–520. [Google Scholar] [CrossRef]
- McLean, C.M.; Bradstock, R.; Price, O.; Kavanagh, R.P. Tree hollows and forest stand structure in Australian warm temperate Eucalyptus forests are adversely affected by logging more than wildfire. For. Ecol. Manag. 2015, 341, 37–44. [Google Scholar] [CrossRef]
Figure 1.
(a) Satellite image of the AWC site in central Pilliga with areas affected by the management burn mapped. Pink line indicates the maximum foraging range (3.96 km radius from the mid-point of the burn extent) previously recorded for the species within the study area [22]. The location of roost trees used by N. corbeni and trap locations are also displayed; (b) Image showing a typical area of forest burnt by low severity fire; (c) Image showing a typical area of forest burnt by higher severity fire. Note the collapsed tree post-fire in the background and burnt-out fallen log in foreground.
Figure 1.
(a) Satellite image of the AWC site in central Pilliga with areas affected by the management burn mapped. Pink line indicates the maximum foraging range (3.96 km radius from the mid-point of the burn extent) previously recorded for the species within the study area [22]. The location of roost trees used by N. corbeni and trap locations are also displayed; (b) Image showing a typical area of forest burnt by low severity fire; (c) Image showing a typical area of forest burnt by higher severity fire. Note the collapsed tree post-fire in the background and burnt-out fallen log in foreground.
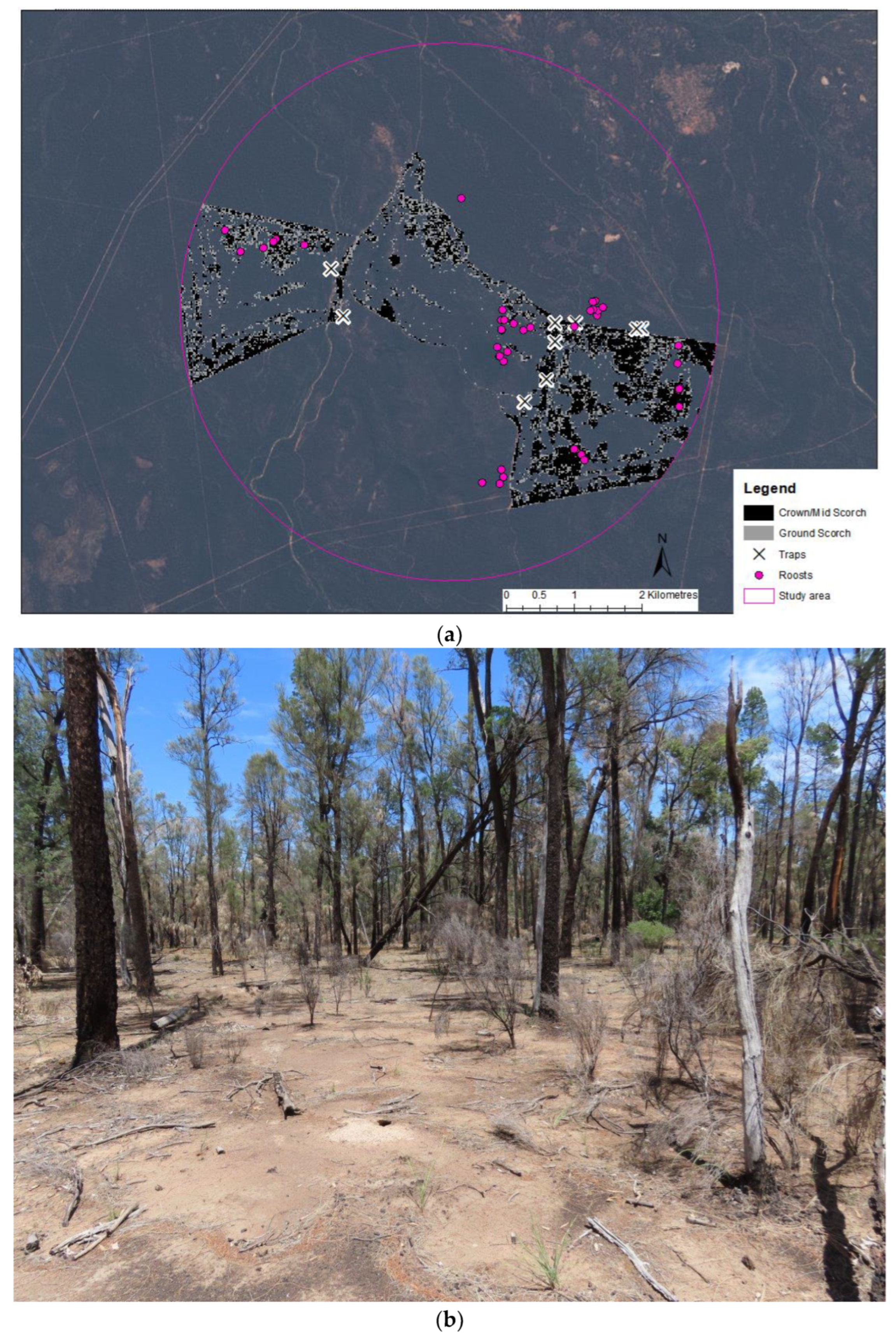

Figure 2.
Column graph illustrating the proportion of roost (used) and locally available trees (>10 cm dbhob) represented by five broad species/groups in the AWC site, Central Pilliga.
Figure 2.
Column graph illustrating the proportion of roost (used) and locally available trees (>10 cm dbhob) represented by five broad species/groups in the AWC site, Central Pilliga.
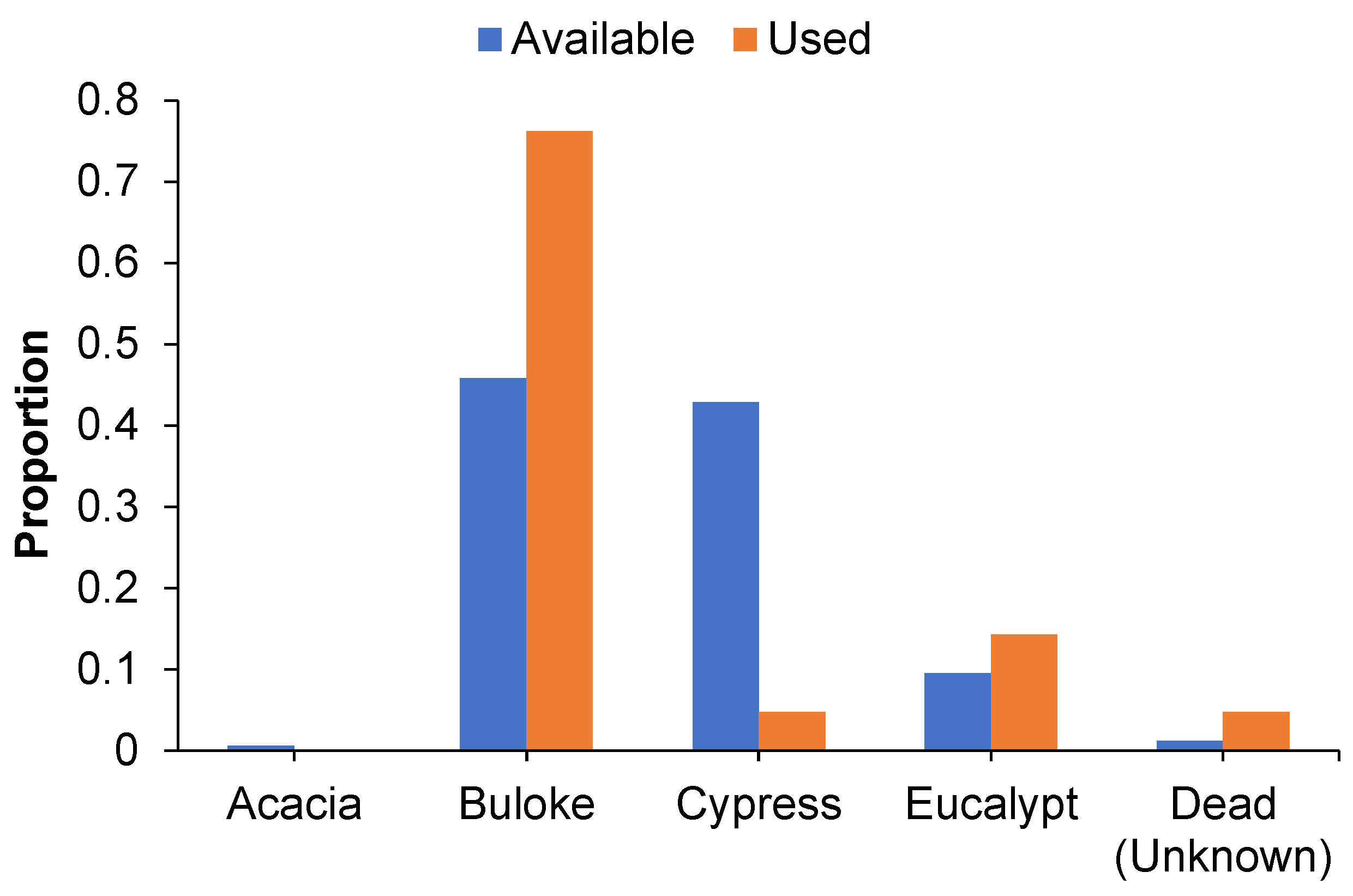
Figure 3.
Column graph illustrating the proportion of roost (used) and locally available trees (>10 cm dbhob) represented by living and dead trees in the AWC site, Central Pilliga.
Figure 3.
Column graph illustrating the proportion of roost (used) and locally available trees (>10 cm dbhob) represented by living and dead trees in the AWC site, Central Pilliga.

Figure 4.
Column graphs illustrating the characteristics of roost (used) and locally available trees (>10 cm dbhob) represented by living and dead trees in the AWC site, Central Pilliga: (a) diameter at breast height over bark, (b) height, (c) canopy cover, (d) senescence and (e) hollow index.
Figure 4.
Column graphs illustrating the characteristics of roost (used) and locally available trees (>10 cm dbhob) represented by living and dead trees in the AWC site, Central Pilliga: (a) diameter at breast height over bark, (b) height, (c) canopy cover, (d) senescence and (e) hollow index.

Figure 5.
Canonical analysis of principal coordinates illustrating the importance of tree characteristics of N. corbeni roost and available trees. Green triangle = dead tree roosts; blue inverted triangle = living tree roosts; blue square = dead tree available for roosting; red diamond = living tree available for roosting.
Figure 5.
Canonical analysis of principal coordinates illustrating the importance of tree characteristics of N. corbeni roost and available trees. Green triangle = dead tree roosts; blue inverted triangle = living tree roosts; blue square = dead tree available for roosting; red diamond = living tree available for roosting.
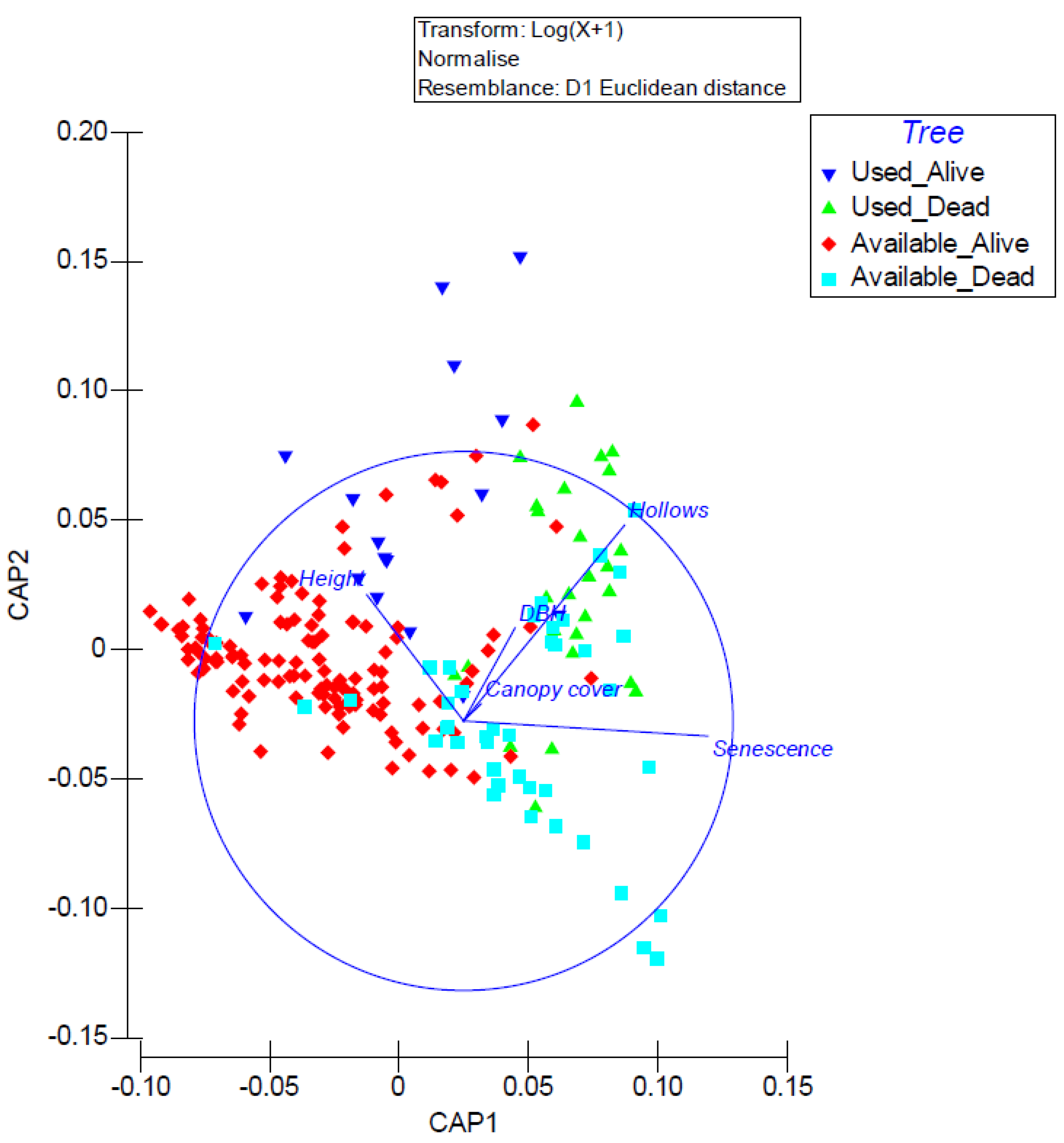
Figure 6.
Images of a selection of roost trees used by N. corbeni. Red arrow indicates location of bat.
Figure 6.
Images of a selection of roost trees used by N. corbeni. Red arrow indicates location of bat.

Figure 7.
Column graphs illustrating the distribution of roost trees among the post-burn mosaic of unburnt, low severity burn, and higher severity burn. Data represent modeled estimates of means after accounting for random variation associated with individual bats. Also shown are the relative proportions available within (a) 100 m neighborhood, (b) 250 m neighborhood, (c) 500 m neighborhood and (d) 1 km neighborhood of each female roost tree, and (e) 100 m neighborhood, (f) 250 m neighborhood, (g) 500 m neighborhood and (h) 1 km neighborhood of each male roost tree.
Figure 7.
Column graphs illustrating the distribution of roost trees among the post-burn mosaic of unburnt, low severity burn, and higher severity burn. Data represent modeled estimates of means after accounting for random variation associated with individual bats. Also shown are the relative proportions available within (a) 100 m neighborhood, (b) 250 m neighborhood, (c) 500 m neighborhood and (d) 1 km neighborhood of each female roost tree, and (e) 100 m neighborhood, (f) 250 m neighborhood, (g) 500 m neighborhood and (h) 1 km neighborhood of each male roost tree.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bat species captured across all sites.
| Species | No. of Captures |
|---|---|
| Chalinolobus gouldii | 58 |
| Chalinolobus picatus | 3 |
| Nyctophilus corbeni | 17 |
| Nyctophilus geoffroyi | 23 |
| Nyctophilus gouldi | 56 |
| Ozimops planiceps | 1 |
| Ozimops ridei | 2 |
| Scotorepens balstoni | 27 |
| Scotorepens greyii | 31 |
| Scotorepens sp. | 59 |
| Vespadelus vulturnus | 115 |
| Unidentified * | 1 |
* Escaped from trap.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gonsalves, L.; Law, B.; Brassil, T.; Kerr, I.; O’Loughlin, C. Roost Selection in Relation to a Patchy, Mosaic Management Burn by a Threatened Clutter-Adapted Bat. Forests 2022, 13, 1327. https://doi.org/10.3390/f13081327
AMA Style
Gonsalves L, Law B, Brassil T, Kerr I, O’Loughlin C. Roost Selection in Relation to a Patchy, Mosaic Management Burn by a Threatened Clutter-Adapted Bat. Forests. 2022; 13(8):1327. https://doi.org/10.3390/f13081327
Chicago/Turabian StyleGonsalves, Leroy, Brad Law, Traecey Brassil, Isobel Kerr, and Christopher O’Loughlin. 2022. "Roost Selection in Relation to a Patchy, Mosaic Management Burn by a Threatened Clutter-Adapted Bat" Forests 13, no. 8: 1327. https://doi.org/10.3390/f13081327
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.