
Localization of TWISTED NEEDLES Locus on Linkage Map of Japanese Cedar (Cryptomeria japonica D. Don)
by
 ,
,
Yoshinari Moriguchi
1,*,
Ryunosuke Saito
1,
Saneyoshi Ueno
2,
Yoichi Hasegawa
2,
Hiroyuki Kakui
1 and
Asako Matsumoto
2 1
Graduate School of Science and Technology, Niigata University, 8050 Ikarashi 2-Nocho, Nishi-ku, Niigata 950-2181, Japan
2
Department of Forest Molecular Genetics and Biotechnology, Forestry and Forest Products Research Institute, Forest Research and Management Organization, 1 Matsunosato, Tsukuba 305-8687, Japan
*
Author to whom correspondence should be addressed.
Forests 2022, 13(9), 1524; https://doi.org/10.3390/f13091524
Submission received: 14 August 2022
/
Revised: 6 September 2022
/
Accepted: 16 September 2022
/
Published: 19 September 2022
(This article belongs to the Special Issue Application of Biotechnology Techniques on Tree Species—Series II)
Abstract
:Sugi (Cryptomeria japonica D. Don) is an important forestry species in Japan. C. japonica ‘Spiralis’ is a mutant with twisted needles. The mutant is called Yore-sugi in Japan and is known as ‘Rasen’ in other countries. The twisted trait is regulated by a dominant gene called TWISTED NEEDLES, and it can be identified by observing the needles within 1 to 2 months after germination. The TWISTED NEEDLES gene may be useful for improving the efficiency of genome editing technology in C. japonica. In this study, we attempted to identify the linkage group of the TWISTED NEEDLES locus and investigate markers that sandwich this locus using the mapping family. First, we identified the linkage group containing the TWISTED NEEDLES locus based on the distortion from the expected segregation ratio using 32 mutant individuals of the mapping family. The segregation distortion showed that the TWISTED NEEDLES locus was located on the 11th linkage group (LG11). Next, a linkage map of LG11 was constructed based on genotype data from the single-nucleotide polymorphism (SNP) markers and double digested restriction site-associated DNA sequencing (ddRAD-seq) using 123 individuals of the MMY-1 family. On this map, six markers were located at the same position as the TWISTED NEEDLES locus. To investigate markers sandwiching the TWISTED NEEDLES locus, a partial linkage map around the TWISTED NEEDLES locus was constructed using 643 individuals of the MMY-1 family. The TWISTED NEEDLES locus was located in the 0.6 cM region between gSNP01822 and the other five markers (Contig_4705-179, Contig_4518-93, Contig_4398-118, gSNP04056, and Contig_4970-113).
1. Introduction
Generally, conifers have a large genome (>10 Gb) and contain many repetitive sequences, which makes it difficult to perform genomic studies on them. However, due to the rapid progress of sequencing technology in recent years, genome sequence studies have been carried out on some coniferous trees, e.g., Picea glauca [1], Pinus lambertiana [2], Pseudotsuga menziesii [3], Abies alba [4], and Sequoiadendron giganteum [5].
Sugi (Cryptomeria japonica D. Don) is an allogamous, diploid (2n = 22), wind-pollinated conifer species with high importance for commercial forestry in Japan. Since 1957, a government-funded tree improvement program, currently covering an area of 4.5 million hectares (44% of all Japanese artificial forests), has been conducted in Japan [6,7]. Many studies, such as expressed sequence tag (EST) collection [8,9,10,11,12,13], DNA marker development [14,15,16,17,18,19], linkage map construction [20,21,22,23], marker-assisted selection [24], experimental pyramiding breeding [25], plant regeneration through somatic embryogenesis [26,27,28,29,30,31,32], genetic transformation [26,33,34,35], and genome editing [36], have been performed to accelerate the molecular breeding of C. japonica.
Because tree species such as C. japonica have a long generation time and require space for growing seedlings, a highly efficient transformation system is required for their genetic engineering, including genome editing. Therefore, it is important to develop a simple method to accurately evaluate the efficiency of genome editing. Nanasato et al. [36] first introduced the green fluorescent protein (GFP) gene into embryonic callus (using calli derived from immature seeds for transformation) and produced calli with GFP fluorescence in C. japonica. Then, they knocked out the GFP activity using CRISPR/Cas9 and evaluated the genome editing efficiency by observing GFP signals. Next, they disrupted the endogenous magnesium chelatase subunit I (CjChlI) gene, which is required for chlorophyll biosynthesis, by genome editing. As a result, several lines showed albino or pale-green phenotypes, indicating successful genome editing in C. japonica [36].
C. japonica ‘Spiralis’ [37] is a mutant that shows helical growth of needles, i.e., it has twisted needles (Figure 1). The mutant is called Yore-sugi in Japan [38]. In other countries, it is known as ‘Rasen’ [39]. The twisted needle trait is regulated by a dominant gene named TWISTED NEEDLES [38] and several studies of its heritability and isozyme pattern have been conducted due to its interesting phenotype [40,41]. Many plants with helical growth phenotypes are known such as Arabidopsis thaliana and Oryza sativa. In many cases, the helical growth phenotype is based on defects in cell expansion symmetry, which are caused by the microtubule system, cell wall functions, auxin transport, and more [39]. In C. japonica, the trait can be observed within 1 to 2 months from germination. Therefore, we considered that the efficiency of genome editing could be evaluated quickly and accurately if we could knock out the TWISTED NEEDLES gene and observe the twisted trait. It is difficult to evaluate the efficiency of genome editing accurately using CjChlI because it is an essential gene for plant growth, and genome-edited plantlets show strong growth inhibition [36]. However, the twisted trait should enable us to accurately evaluate genome editing efficiency because it does not affect plant growth. Furthermore, the TWISTED NEEDLES gene does not have to be introduced in advance, as opposed to GFP, because it is an endogenous gene in C. japonica. By targeting TWISTED NEEDLES, the efficiency of genome editing can be evaluated by observing the phenotype of plantlets. Moreover, the dominant twisted trait can be distinguished accurately between wild-type and heterozygous genotypes, whereas the heterozygous condition of CjChlI may be overlooked by phenotypic observation. Nanasato et al. [36] reported that one heterozygous line of CjChlI produced a green phenotype that appeared the same as the wild-type phenotype [36].
As another advantage, the twisted trait is expected to be useful as a positive selection marker to determine whether a vector is present in plantlets based on phenotype observation by introducing the TWISTED NEEDLES gene (Tw, twisted trait; see Section 2) into a genome editing vector. In the T1 generation of the genome editing, plantlets having the vector could be selected easily by observing the twisted trait. It is important to remove unnecessary vectors after genome editing for future applications such as field plantation. Therefore, it should be possible to select genome-edited plantlets without external vector sequences by selecting non-twisted plantlets from the T2 (or later) generation. Thus, the identification of the TWISTED NEEDLES gene is important for both accelerating the investigation of helical needle growth mechanisms in conifers and also contributing to molecular biological techniques, such as improvement of genome editing technology, in C. japonica.
In this study, we attempted to localize the TWISTED NEEDLES locus in C. japonica. We identified a linkage group of the TWISTED NEEDLES locus using genomic microsatellite (simple sequence repeat, SSR) and EST-SSR markers, constructed a linkage map of LG11 (on which the TWISTED NEEDLES locus mapped) based on genotype data from double digested restriction site-associated DNA sequencing (ddRAD-seq) and the previously reported single-nucleotide polymorphism (SNP) markers [20] using 123 individuals of a mapping family, and finally constructed a partial linkage map around the TWISTED NEEDLES locus using 643 individuals of a mapping family. This study provides basic information for future identification of the TWISTED NEEDLES gene of C. japonica.
2. Materials and Methods
2.1. Mapping Family and Evaluation of Twisted Trait
In this study, we represented the dominant and recessive alleles for the TWISTED NEEDLES locus as Tw (twisted needle trait) and tw (normal trait (wild type)), respectively. To construct a linkage map, we used the MMY-1 family (Figure 2). The family was derived from a backcross between ‘Midori 5’ [tw/tw] and F1 plant M5Y-1 [Tw/tw]. The M5Y-1 plant [Tw/tw] was an F1 plant between ‘Midori 5’ [tw/tw] and ‘Houyore’ [Tw/tw]. ‘Midori 5’ had normal needles, and ‘Houyore’ and M5Y-1 had twisted needles. Strobili production was promoted by spraying the trees with gibberellin-3 (100 ppm) in July 2016, and the artificial crossing was performed during March 2017. In autumn 2017, seeds were collected from the mother tree, ‘Midori 5’. The seeds were sown on sterilized paper in plastic Petri dishes in a controlled-environment growth chamber (BIOTRON; Nippon Medical and Chemical Instruments, Japan) at 25 °C under fluorescent light (15,000 lx, 16 h light/8 h dark photoperiod), and germinated seeds with ≥2 mm roots were transplanted in a 128-cell plug tray filled with spagmoss (Sphagnum subnitens; Besgrow, Christchurch, New Zealand). Seedlings were grown at 25 °C during the winter season and transferred outside in April 2018. The needle phenotype (twisted or normal) of seedlings was evaluated visually at ≥2 months after germination.
2.2. DNA Extraction
Genomic DNA was extracted from needle tissues using a modified hexadecyltrime-thylammonium bromide (CTAB) method [42]. The concentration of DNA samples was determined using agarose gel electrophoresis. First, electrophoresis using a 2% agarose gel was performed. For this process, 1 µL of genomic DNA or λDNA (100 ng/µL) as a standard was mixed with 1 µL of loading buffer, 7 µL of sterile water, and 1 µL of fluorescent dye (UltraPower DNA Stain; Gellex International, Tokyo, Japan) and incubated for 3 min at room temperature. Electrophoresis was carried out in TAE buffer for 15 min at a constant voltage of 100 V. The DNA bands were visualized with FAS-IV (NIPPON Genetics, Tokyo, Japan), and electrophoresis images were acquired. DNA concentrations for ddRAD-seq were quantified using a Qubit 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA); those for other markers were estimated by electrophoresis using λDNA.
2.3. Identification of Linkage Group Including TWISTED NEEDLES Locus
We used only 32 twisted-needle individuals of the MMY-1 family to identify the linkage group containing the TWISTED NEEDLES locus. If a marker is linked to the TWISTED NEEDLES locus, the genotypic segregation ratio of the marker and the TWISTED NEEDLES locus should be significantly distorted from the expected outcome. For this analysis, a total of 22 genomic SSR or EST-SSR markers were used (see Section 3). The KAPA2G Fast PCR Kit (Kapa Biosystems, Wilmington, MA, USA), Go-Taq polymerase (Promega, Fitchburg, WI, USA), and Multiplex PCR master mix (Qiagen, Venlo, The Netherlands) were used for the PCR amplifications of genomic SSR and EST-SSR markers. PCR amplifications were carried out using the Takara PCR Thermal Cycler (Takara, Tokyo, Japan). For the KAPA2G Fast PCR Kit, a reaction mixture with a total volume of 10 μL was used. It consisted of 5 ng of genomic DNA, 1× PCR Kapa2G buffer with 1.5 mM MgCl2, 0.2 µL of 25 mM MgCl2, 0.2 µL of 10 mM each dNTP mix, 0.8 µL of 5 µM fluorescently labeled forward primers, 0.8 µL of 5 µM reverse primers, 5 ng template DNA, and 0.2 U KAPA2G Fast PCR polymerase. The PCR conditions were 15 min at 95 °C, then 40 cycles of 15 s at 95 °C, 15 s at 60 °C, and 1 min at 72 °C, followed by 1 min at 72 °C. For the Go-Taq polymerase, a reaction mixture with a total volume of 10 μL was used. The mixture consisted of 5 ng of genomic DNA, 1× PCR buffer, 0.8 µL of 25 mM MgCl2, 1.0 µL of 2 mM each dNTP mix, 0.8 µL of 5 µM fluorescently labeled forward primers, 0.8 µL of 5 µM reverse primers, and 0.5 U Go-Taq polymerase. The PCR conditions were 5 min at 94 °C, then 35 cycles of 30 s at 94 °C, 30 s at 60 °C, and 30 s at 72 °C, followed by 5 min at 72 °C. For the Multiplex PCR master mix, a reaction mixture with a total volume of 6 μL was used. It consisted of 5 ng of genomic DNA, consisting of 1× Multiplex PCR master mix (Qiagen) and 2.0 µL of primer mix (equal concentrations of fluorescently labeled forward primer and reverse primer). The PCR conditions were 15 min at 95 °C, then 30 cycles of 30 s at 94 °C, 45 s at 60 °C, and 1 min at 72 °C, followed by 30 min at 60 °C. The PCR products and the DNA size marker (600LIZ; Thermo Fisher Scientific) were separated by capillary electrophoresis on an ABI 3130 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). The DNA fragments were analyzed using GeneMarker software (ver. 2.4.0; SoftGenetics, State College, PA, USA). The segregation independence of each SSR or EST-SSR marker was investigated using the chi squared test.
2.4. Construction of a Linkage Map Using ddRAD-Seq
The 123 F1 individuals from the MMY-1 family (74 twisted-needle individuals and 49 normal-needle individuals), which were not selected intentionally (derived from plastic Petri dish No.1 and No.2), were used to construct a linkage map of LG11, on which the TWISTED NEEDLES locus is located (see Section 3).
First, high-throughput double digest RAD-seq [43] was performed. Genomic DNA was digested with PstI and SphI and ligated with adapters (Fluidigm, San Francisco, CA, USA). After PCR amplification using adapter-specific primers, equal amounts of PCR products from each sample were mixed to construct a RAD library. The library obtained was size-selected using the Blue Pippin Prep tool (Sage Science, Beverly, MA, USA), and the quantity was assessed using real-time PCR (LightCycler480, Roche Molecular Systems, Mannheim, Germany). After adjusting the concentration of the library, sequencing was performed with paired-end (2 × 150 bp) reads on an Illumina HiSeqX system (Illumina, San Diego, CA, USA) by the Macrogen Japan Corporation (Kyoto, Japan). Second, genotyping using the BioMark 48.48 Dynamic Array (Fluidigm) was performed for the six SNP markers previously mapped on LG11 [12], where the TWISTED NEEDLES locus was located (see Section 3). The assays were carried out following the protocol provided by the manufacturer. The data obtained were analyzed using Fluidigm SNP Genotyping Analysis software v4.1.2 (Fluidigm, San Francisco, CA, USA).
For markers derived from same contig in ddRAD-seq and showing the same genotypes in all individuals in mapping family, we used only one, excluding the others. Linkage analysis was performed using JoinMap v4.1 (Kyazma, Wageningen, Netherlands) with a back-cross population design, the regression mapping algorithm, and two rounds of map calculation [44]. Markers were initially assigned to tentative linkage groups using logarithm-of-odds ratio (LOD) thresholds of 3.0 to 15.0, with increments of 1.0; a LOD threshold of 12.0 was used when groups of markers were defined. The map distance was calculated using the Kosambi mapping function [45]. The default settings were used for the recombination frequency threshold and ripple value. The linkage group number (1–11) was defined following the genetic map published by Moriguchi et al. [20]. Images of the linkage groups were drawn using MapChart ver. 2.0 [46].
2.5. Identification of Markers Which Sandwich the TWISTED NEEDLES Locus
To investigate markers sandwiching the TWISTED NEEDLES locus, the 643 F1 individuals from the MMY-1 family (378 twisted-needle individuals and 265 normal-needle individuals), including the 123 F1 individuals described in Section 2.4, were used to construct a partial linkage map around the TWISTED NEEDLES locus. To do so, we designed SNPType primer sets using the online tool D3 Assay Design (Fluidigm). Of the six markers mapped to the same position as the TWISTED NEEDLES locus on the linkage map (see Section 3), five markers (Contig_4705-179, Contig_4518-93, Contig_4398-118, gSNP04056, and Contig_4970-113) could be designed primers for BioMark 48.48 Dynamic Array (Fluidigm). In addition to them, the gSNP01822 marker for which primers for BioMark 48.48 Dynamic Array (Fluidigm) had been previously designed and closely linked to the TWISTED NEEDLES locus were used for analysis. Genotyping and linkage analyses were performed as described in Section 2.3.
3. Results and Discussion
In this study, we used a total of 22 genomic SSR or EST-SSR markers to identify the linkage group of the TWISTED NEEDLES locus. They were selected from 49 markers scattered on all linkage groups previously reported [17,18,19,21] (three to seven loci per linkage group, Table S1). The expected segregation ratios of these are 1:1:1:1 or 1:1. The former is obtained in the locus when both parents are heterozygous, i.e., the case in which there is no common allele between parents (ab × cd type) and the case in which one allele is common between parents (ef × eg type). The latter is obtained when one parent is homozygous and the other is heterozygous, i.e., the case in which the female parent is homozygous, and the male parent is heterozygous (nn × np type) and the case in which the female parent is heterozygous, and the male parent is homozygous (lm × ll type). Among these, the lm × ll type markers cannot be used for the localization because the genotype of all F1 individuals having the twisted-needle Tw is homozygous. Thus, we excluded them from the analysis. The chi-square test results for independence for the 22 markers revealed that one locus, Cjgssr124_S, on LG11 showed high segregation distortion (χ2 = 28.13, p < 0.0001; Table 1). Therefore, the TWISTED NEEDLES locus was located on LG11.
As a result of constructing a linkage map using 123 individuals of the MMY-1 family, a total of 106 markers and the TWISTED NEEDLES locus were mapped on LG11 (Figure 3). All markers were derived from ddRAD-seq, except for the six markers previously reported (gSNP01822, gSNP04056, gSNP01363, gSNP00119, gSNP00725, and gSNP02051) [20]. The total map length of LG11 was 110.8 cM (1.05 cM/marker). The order of the six SNP markers was the same as that of the map reported in Moriguchi et al. [20]. In the linkage map constructed in this study, six markers (one from the previously reported SNP marker (gSNP04056) and the remaining five derived from ddRAD-seq (Contig_4518-93, Con-tig_4970-113, Contig_4705-179, Contig_4398-118, Contig_1472-16)) were mapped at the same position as the TWISTED NEEDLES locus.
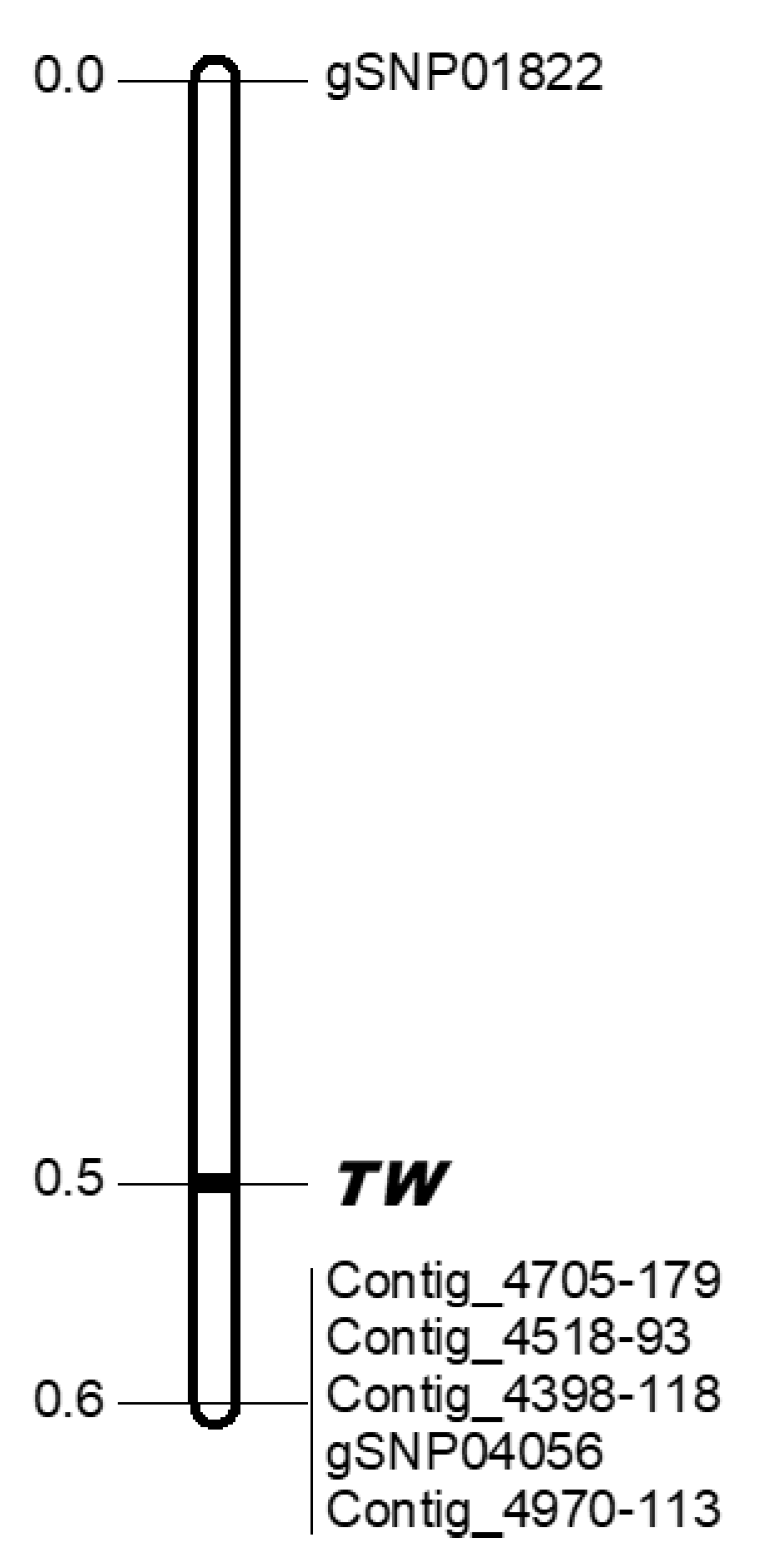
To investigate the markers that sandwiched the TWISTED NEEDLES locus, a partial linkage map around the TWISTED NEEDLES locus was constructed using 643 individuals of the MMY-1 family. As the result of this analysis, the TWISTED NEEDLES locus was lo-cated in the 0.6 cM region between gSNP01822 and the other five markers (Contig_4705-179, Contig_4518-93, Contig_4398-118, gSNP04056, and Contig_4970-113) (Figure 3). A BLASTN homology search of these markers against the genome sequence of Sequoia sempervirens [5], a coniferous species in the same family (Cupressaceae) as C. japonica, identified homologous positions on chromosome 8 with coordinates ranging from 564.1 Mb to 643.2 Mb for Contig_4518-93 and gSNP01822, respectively (Table S2). It corresponds to approximately 79 Mbp in S. sempervirens. Five out of the six SNP markers around the Tw locus aligned to the only available Cupressaceae genome sequence, S. sempervirens, suggesting that some of the regions were conserved and the Tw locus may be narrowed down to the region (even in the case of C. japonica, the 0.6 cM region may correspond to a similar physical distance).
Of the 643 individuals of the MMY-1 family, 378 individuals had twisted needles and the remaining 265 individuals had normal needles. This was significantly distorted from the 1:1 separation ratio expected (χ2 = 19.54, p < 0.001). The five markers mapped at the same position as the TWISTED NEEDLES locus were also distorted (Figure 4). The clustering of markers with distorted segregation has also been reported in some crop species [47,48,49]. Mukai et al. [50] and Iwata et al. [16] pointed out that clusters of markers with distorted segregation may be due to linkage with deleterious or lethal alleles. Because the number of individuals with normal needles was fewer than the number with twisted needles, the cause of the distortion of the needle trait observed in the MMY-1 population is thought to be linkage between the tw allele and deleterious/lethal alleles derived from ‘Midori 5’.
As mentioned previously, using the twisted trait of C. japonica ‘Spiralis’ is expected to improve the efficiency of genome editing. This will be achieved by identification of the gene of TWISTED NEEDLES, which will be promoted by clarification of the genome sequence in C. japonica, as in other foreign conifers, in the future. In conifers, although a variety of pine (Pinus strobus) with the twisted needle trait, ‘Contorta’, is known [39], the mechanics behind helical needle growth are not understood. The future study of the TWISTED NEEDLES gene in C. japonica will also promote the investigation of helical needle growth mechanisms in conifers.
Supplementary Materials
The following are available online at: https://www.mdpi.com/article/10.3390/f13091524/s1. Table S1: Summary of SSR and EST-SSR markers used in this study, Table S2: Blast homology search against sequoia genome reference sequence for markers tightly linked to TW locus in C. japonica. References [51,52] are cited in Table S1.
Author Contributions
Conceived and designed the experiments, Y.M.; performed the experiments and analyzed the data, R.S., S.U., A.M. and Y.H.; methodology, Y.M. and S.U.; funding acquisition, S.U.; writing—original draft, Y.M., S.U., R.S. and H.K.; writing—review and editing, Y.M., R.S., H.K., Y.H. and S.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Research grant of the Forestry and Forest Products Research Institute (#201906) to S.U.
Data Availability Statement
The datasets used and/or analyzed in this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors would like to thank Y. Komatsu for laboratory work. We also thank M. Nishiguchi for providing information about genome editing in C. japonica.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Warren, R.L.; Keeling, C.I.; Yuen, M.M.; Raymond, A.; Taylor, G.A.; Vandervalk, B.P.; Mohamadi, H.; Paulino, D.; Chiu, R.; Jackman, S.D.; et al. Improved white spruce (Picea glauca) genome assemblies and annotation of large gene families of conifer terpenoid and phenolic defense metabolism. Plant J. 2016, 83, 189–212. [Google Scholar] [CrossRef] [PubMed]
- Stevens, K.A.; Wegrzyn, J.L.; Zimin, A.; Puiu, D.; Crepeau, M.; Cardeno, C.; Paul, R.; Conzalez-Ibeas, D.; Koriabine, M.; Holtz-Morris, A.E.; et al. Sequence of the sugar pine megagenome. Genetics 2016, 204, 1613–1626. [Google Scholar] [CrossRef]
- Neal, D.B.; McGuire, P.E.; Wheeler, N.C.; Stevens, K.A.; Crepeau, M.W.; Cardeno, C.; Zimin, A.V.; Puiu, D.; Pertea, G.M.; Sezen, U.U.; et al. The Douglas-Fir genome sequence reveals specialization of the photosynthetic apparatus in Pinaceae. G3 (Bethesda) 2017, 7, 3157–3167. [Google Scholar] [CrossRef] [PubMed]
- Mosca, E.; Cruz, F.; Gómez-Garrido, J.; Bianco, L.; Rellstab, C.; Brodbeck, S.; Csilléry, K.; Fady, B.; Fladung, M.; Fussi, B.; et al. A reference genome sequence for the European silver fir (Abies alba Mill.): A community-generated genomic resource. G3 (Bethesda) 2019, 9, 2039–2049. [Google Scholar] [CrossRef]
- Scott, A.D.; Zimin, A.V.; Puiu, D.; Workman, R.; Britton, M.; Zaman, S.; Caballero, M.; Read, A.C.; Bogdanove, A.J.; Burns, M.; et al. A reference genome sequence for giant sequoia. G3 (Bethesda) 2020, 10, 3907–3919. [Google Scholar] [CrossRef] [PubMed]
- Forestry Agency. Statistical Handbook of Forest and Forestry; Forestry Agency, Ministry of Agriculture, Forestry and Fisheries: Tokyo, Japan, 2014; pp. 8–9. (In Japanese) [Google Scholar]
- Tsumura, Y. Cryptomeria. In Wild Crop Relatives: Genomics and Breeding Resources, Forest Trees; Kole, C., Ed.; Springer: Berlin, Germany, 2011; pp. 49–64. [Google Scholar]
- Ujino-Ihara, T.; Yoshimura, K.; Ugawa, Y.; Yoshimaru, H.; Nagasaka, K.; Tsumura, Y. Expression analysis of ESTs derived from the inner bark of Cryptomeria japonica. Plant Mol. Biol. 2000, 43, 451–457. [Google Scholar] [CrossRef]
- Ujino-Ihara, T.; Taguchi, Y.; Yoshimura, K.; Tsumura, Y. Analysis of expressed sequence tags derived from developing seed and pollen cones of Cryptomeria japonica. Plant Biol. 2003, 5, 600–607. [Google Scholar] [CrossRef]
- Ujino-Ihara, T.; Kanamori, H.; Yamane, H.; Taguchi, Y.; Namiki, N.; Mukai, Y.; Yoshimura, K.; Tsumura, Y. Comparative analysis of expressed sequence tags of conifers and angiosperms reveals sequences specifically conserved in conifers. Plant Mol. Biol. 2005, 59, 895–907. [Google Scholar] [CrossRef]
- Futamura, N.; Ujino-Ihara, T.; Nishiguchi, M.; Kanamori, H.; Yoshimura, K.; Sakaguchi, M.; Shinohara, K. Analysis of expressed sequence tags from Cryptomeria japonica pollen reveals novel pollen-specific transcripts. Tree Physiol. 2006, 26, 1517–1528. [Google Scholar] [CrossRef]
- Futamura, N.; Totoki, Y.; Toyoda, A.; Igasaki, T.; Nanjo, T.; Seki, M.; Sakaki, Y.; Mari, A.; Shinozaki, K.; Shinohara, K. Characterization of expressed sequence tags from a full-length enriched cDNA library of Cryptomeria japonica male strobili. BMC Genom. 2008, 9, 383. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Nishiguchi, M.; Futamura, N.; Nanjo, T. Expressed sequence tags from Cryptomeria japonica sapwood during the drying process. Tree Physiol. 2007, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, Y.; Suyama, Y.; Yoshimura, K.; Shirato, N.; Mukai, Y. Sequence-tagged-sites (STSs) of cDNA clones in Cryptomeria japonica and their evaluation as molecular markers in conifers. Theor. Appl. Genet. 1997, 94, 764–772. [Google Scholar] [CrossRef]
- Nikaido, A.M.; Ujino, T.; Iwata, H.; Yoshimura, K.; Yoshimura, H.; Suyama, Y.; Murai, M.; Nagasaka, K.; Tsumura, Y. AFLP and CAPS linkage maps of Cryptomeria japonica. Theor. Appl. Genet. 2000, 100, 825–831. [Google Scholar] [CrossRef]
- Iwata, H.; Ujino-Ihara, T.; Yoshimura, K.; Nagasaka, K.; Mukai, Y.; Tsumura, Y. Cleaved amplified polymorphic sequence markers in sugi, Cryptomeria japonica D. Don, and their locations on a linkage map. Theor. Appl. Genet. 2001, 103, 881–895. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Iwata, H.; Ihara, T.; Yoshimura, K.; Taira, H.; Tsumura, Y. Development and characterization of microsatellite markers for Cryptomeria japonica D. Don. Theor. Appl. Genet. 2003, 106, 751–758. [Google Scholar] [CrossRef]
- Tani, N.; Takahashi, T.; Ujino-Ihara, T.; Iwata, H.; Yoshimura, K.; Tsumura, Y. Development and characteristics of microsatellite markers for sugi (Cryptomeria japonica D. Don) from microsatellite enriched libraries. Ann. For. Sci. 2004, 61, 569–575. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Ueno, S.; Ujino-Ihara, T.; Futamura, N.; Matsumoto, A.; Shinohara, K.; Tsumura, Y. Characterization of EST-SSRs from Cryptomeria japonica. Conserv. Genet. Resour. 2009, 1, 373–376. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Ujino-Ihara, T.; Uchiyama, K.; Futamura, N.; Saito, M.; Ueno, S.; Matsumoto, A.; Tani, N.; Taira, H.; Shinohara, K.; et al. The construction of a high-density linkage map for identifying SNP markers that are tightly linked to a nuclear-recessive major gene for male sterility in Cryptomeria japonica D. Don. BMC Genom. 2012, 13, 95. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Ueno, S.; Higuchi, Y.; Miyajima, D.; Itoo, S.; Futamura, N.; Tsumura, Y. Establishment of a microsatellite panel covering the sugi (Cryptomeria japonica) genome, and its application for localization of a male sterile gene (ms2). Mol. Breed. 2014, 33, 315–325. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Uchiyama, K.; Ueno, S.; Ujino-Ihara, T.; Matsumoto, A.; Iwai, J.; Miyajima, D.; Saito, M.; Sato, M.; Tsumura, Y. A high-density linkage map with 2560 markers and its application for the localization of the male-sterile genes ms3 and ms4 in Cryptomeria japonica D. Don. Tree Genet. Genomes 2016, 12, 57. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Ueno, S.; Matsumoto, A.; Ujino-Ihara, T.; Uchiyama, K.; Totsuka, S.; Iwai, J.; Hakamata, T.; Moriguchi, Y. Fine mapping of the male-sterile genes (MS1, MS2, MS3, and MS4) and development of SNP markers for marker-assisted selection in Japanese cedar (Cryptomeria japonica D. Don). PLoS ONE 2018, 13, e0206695. [Google Scholar] [CrossRef] [PubMed]
- Moriguchi, Y.; Ueno, S.; Hasegawa, Y.; Tadama, T.; Watanabe, M.; Saito, R.; Hirayama, S.; Iwai, J.; Konno, Y. Marker-assisted selection of trees with MALE STERILITY 1 in Cryptomeria japonica D. Don. Forests 2020, 11, 734. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Totsuka, S.; Iwai, J.; Matsumoto, A.; Ueno, S.; Tsumura, Y. Pyramiding of male-sterile genes in Cryptomeria japonica D. Don with the aid of closely linked markers. Tree Genet. Genomes 2017, 13, 61. [Google Scholar] [CrossRef]
- Maruyama, E.; Tanaka, T.; Hosoi, Y.; Ishii, K.; Morohoshi, N. Embryogenic cell culture, protoplast regeneration, cryopreservation, biolistic gene transfer and plant regeneration in Japanese cedar (Cryptomeria japonica D. Don). Plant Biotechnol. 2000, 17, 281–296. [Google Scholar] [CrossRef]
- Maruyama, E.T.; Hosoi, Y.; Futamura, N.; Saito, M. Initiation of embryogenic cultures from immature seeds of pollen-free sugi (Cryptomeria japonica). Kanto Shinrin Kenkyu 2014, 65, 107–110, (In Japanese with English abstract). [Google Scholar]
- Taniguchi, T.; Konagaya, K.; Nanasato, Y. Somatic embryogenesis in artificially pollinated seed families of 2nd generation plus trees and cryopreservation of embryogenic tissue in Cryptomeria japonica D. Don (sugi). Plant Biotechnol. 2020, 37, 239–245. [Google Scholar] [CrossRef]
- Igasaki, T.; Sato, T.; Akashi, N.; Mohri, T.; Maruyama, E.; Kinoshita, I.; Walter, C.; Shinohara, K. Somatic embryogenesis and plant regeneration from immature zygotic embryos of Cryptomeria japonica D. Don. Plant Cell Rep. 2003, 22, 239–243. [Google Scholar] [CrossRef]
- Igasaki, T.; Akashi, N.; Ujino-Ihara, T.; Matsubayashi, Y.; Sakagami, Y.; Shinohara, K. Phytosulfokine stimulates somatic embryogenesis in Cryptomeria japonica. Plant Cell Physiol. 2003, 44, 1412–1416. [Google Scholar] [CrossRef]
- Nakagawa, R.; Ogita, S.; Kubo, T.; Funada, R. Effect of polyamines and L-ornithine on the development of proembryogenic masses of Cryptomeria japonica. Plant Cell Tissue Organ. Cult. 2006, 85, 229–234. [Google Scholar] [CrossRef]
- Maruyama, E.; Hosoi, Y. Polyethylene glycol enhance somatic embryo production in Japanese cedar (Cryptomeria japonica D. Don). Propag. Ornam. Plants 2007, 7, 57–61. [Google Scholar]
- Taniguchi, T.; Ohmiya, Y.; Kurita, M.; Tsubomura, M.; Kondo, T. Regeneration of transgenic Cryptomeria japonica D. Don after Agrobacterium tumefaciens-mediated transformation of embryogenic tissue. Plant Cell Rep. 2008, 27, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Konagaya, K.; Kurita, M.; Taniguchi, T. High-efficiency Agrobacterium-mediated transformation of Cryptomeria japonica D. Don by co-cultivation on filter paper wicks followed by meropenem treatment to eliminate Agrobacterium. Plant Biotechnol. 2013, 30, 523–528. [Google Scholar] [CrossRef] [Green Version]
- Konagaya, K.; Nagasato, Y.; Taniguchi, T. A protocol for Agrobacterium-mediated transformation of Japanese cedar, Sugi (Cryptomeria japonica D. Don) using embryogenic tissue explants. Plant Biotechnol. 2020, 37, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Nanasato, Y.; Mikami, M.; Futamura, N.; Endo, M.; Nishiguchi, M.; Ohmiya, Y.; Konagaya, K.; Taniguchi, T. CRISPR/Cas9-mediated targeted mutagenesis in Japanese cedar (Cryptomeria japonica D. Don). Sci. Rep. 2021, 11, 16186. [Google Scholar] [CrossRef]
- Yonekura, K.; Kajita, T.; BG Plants. YList. Available online: http://bean.bio.chibau.jp/bgplants/ylist_main.html (accessed on 11 November 2021).
- Ohba, K.; Maeda, T.; Fukuhara, N. Inheritance of twisted-leaf sugi, Cryptomeria japonica D. DON and linkage between the twisted-leaf gene and two recessive genes, albino and green (midori sugi). J. Jpn. For. Soc. 1974, 56, 276–281, (In Japanese with English abstract). [Google Scholar]
- Buschmann, H.; Borchers, A. Handedness in plant cell expansion: A mutant perspective on helical growth. New Phytol. 2020, 225, 53–69. [Google Scholar] [CrossRef]
- Kuromaru, M. Time serial changes of isozyme patterns of primary leaves on sugi (Cryptomeria japonica D. DON) seedlings. J. Jpn. For. Soc. 1983, 65, 73–81, (In Japanese with English abstract). [Google Scholar]
- Kuromaru, M.; Kawasaki, H.; Ohba, K. Genetic analysis of the peroxidase isozyme of seedlings derived from a twisted-leaf sugi (Cryptomeria japonica D. Don). J. Jpn. For. Soc. 1983, 65, 253–257, (In Japanese with English abstract). [Google Scholar]
- Hasegawa, Y.; Ueno, S.; Fu-Jin, W.; Matsumoto, A.; Ujino-Ihara, T.; Uchiyama, K.; Moriguchi, Y.; Kasahara, M.; Fujino, T.; Shigenobu, S.; et al. Development of diagnostic PCR and LAMP markers for MALE STERILITY 1 (MS1) in Cryptomeria japonica D Don. BMC Res. Note 2020, 13, 457. [Google Scholar] [CrossRef]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double digest RADseq: An inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS ONE 2012, 7, e37135. [Google Scholar]
- Van Ooijen, J.W.; Voorrips, R.E. JoinMap: Version 3.0: Software for the Calculation of Genetic Linkage Maps; University and Research Center: Wageningen, The Netherlands, 2001. [Google Scholar]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eugen. 1944, 12, 172–175. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Heun, M.; Kennedy, A.E.; Anderson, J.A.; Lapitan, N.L.V.; Sorrells, M.E.; Tanksley, S.D. Construction of a restriction fragment length polymorphism map for barley (Hordeum vulgare). Genome 1991, 34, 437–447. [Google Scholar] [CrossRef]
- Kiss, G.B.; Csanadi, G.; Kalman, K.; Kalo, P.; Okresz, L. Construction of a basic genetic map for alfalfa using RFLP, RAPD, isozyme and morphological markers. Mol. Gen. Gent. 1993, 238, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Nodari, R.O.; Tsai, S.M.; Gilbertson, R.L.; Gepts, P. Towards an integrated linkage map of common bean 2. Development of an RFLP-based linkage map. Theor. Appl. Genet. 1993, 85, 513–520. [Google Scholar] [CrossRef]
- Mukai, Y.; Suyama, Y.; Tsumura, Y.; Kawahara, T.; Yoshimaru, H.; Kondo, T.; Tomaru, N.; Kuramoto, N.; Mukai, M. A linkage map for sugi (Cryptomeria japonica) based on RFLP, RAPD, and isozyme loci. Theor. Appl. Genet. 1995, 90, 835–840. [Google Scholar] [CrossRef]
- Guan, L.; Shiraishi, S. Tetranucleotide microsatellite markers in Cryptomeria japonica. Conserv. Genet. Resour. 2011, 3, 283–285. [Google Scholar] [CrossRef]
- Ueno, S.; Moriguchi, Y.; Uchiyama, K.; Ujino-Ihara, T.; Futamura, N.; Sakurai, T.; Shinohara, K.; Tsumura, Y. A second generation framework for the analysis of microsatellites in expressed sequence tags and the development of EST-SSR markers for a conifer, Cryptomeria japonica. BMC Genom. 2012, 13, 136. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Plants and needles of and normal individual (A,C) and Yore-sugi (B,D). Scale bar = 5 cm.

Figure 2.
The MMY-1 family used for linkage analysis in this study.
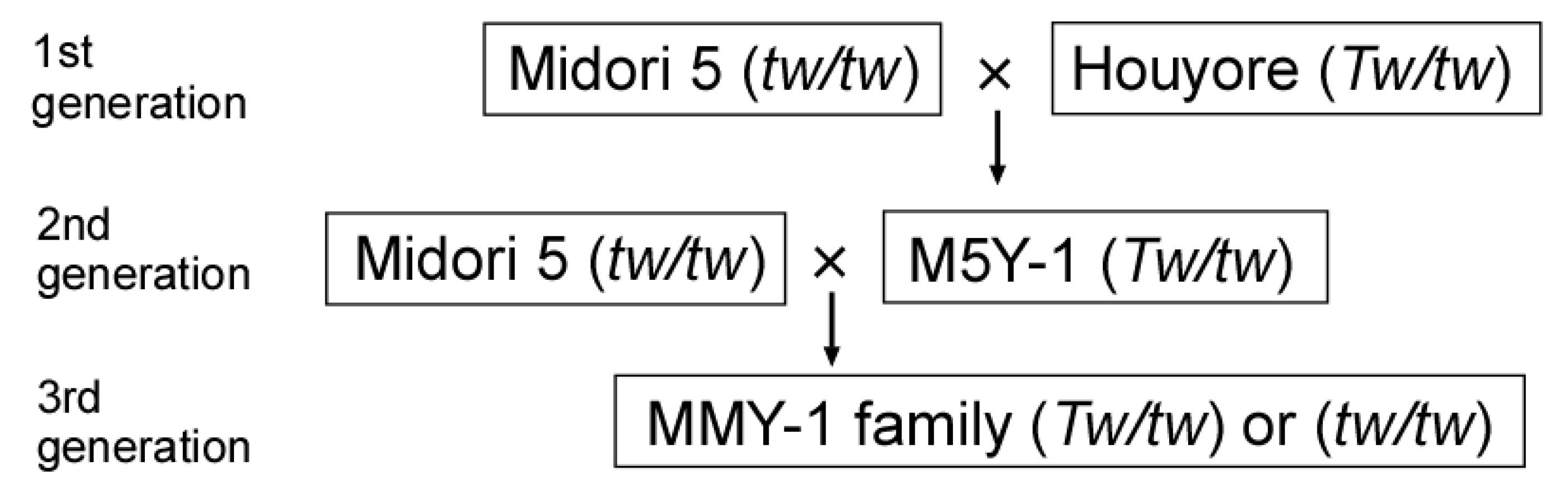
Figure 3.
The linkage map of LG11 constructed in this study. Marker names are indicated to the right of the linkage groups. Centimorgan distances (Kosambi) are indicated to the left of each linkage group. The TWISTED NEEDLES (TW) locus is indicated in bold.
Figure 3.
The linkage map of LG11 constructed in this study. Marker names are indicated to the right of the linkage groups. Centimorgan distances (Kosambi) are indicated to the left of each linkage group. The TWISTED NEEDLES (TW) locus is indicated in bold.

Figure 4.
The partial linkage map around the TWISTED NEEDLES (TW) locus obtained using 643 F1 individuals. Marker names are indicated to the right of the linkage groups. Centimorgan distances (Kosambi) are indicated to the left of each linkage group. The TW locus is indicated in bold.
Figure 4.
The partial linkage map around the TWISTED NEEDLES (TW) locus obtained using 643 F1 individuals. Marker names are indicated to the right of the linkage groups. Centimorgan distances (Kosambi) are indicated to the left of each linkage group. The TW locus is indicated in bold.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Linkage association between genomic microsatellite or EST-SSR markers and the TWISTED NEEDLES locus in C. japonica.
Table 1.
Linkage association between genomic microsatellite or EST-SSR markers and the TWISTED NEEDLES locus in C. japonica.
| Linkage Group | Marker Name | χ2 | P | Number of Analyzed Individuals |
|---|---|---|---|---|
| LG1 | HS4_c16648_ES | 0.35 | 0.56 | 32 |
| CS2169_S | 3.13 | 0.08 | 32 | |
| Cjgssr175_S | 0.50 | 0.48 | 32 | |
| LG2 | BY893784_ES | 1.13 | 0.29 | 32 |
| LG3 | S4049_S | 2.42 | 0.12 | 32 |
| Cjgssr77_S | 0.00 | 0.96 | 32 | |
| LG4 | CJS0333_S | 2.42 | 0.12 | 32 |
| Cjgssr121_S | 0.50 | 0.48 | 32 | |
| Cjgssr123_S | 0.07 | 0.80 | 32 | |
| LG5 | CS0038FC_S | 3.13 | 0.08 | 32 |
| Cjgssr181_S | 0.03 | 0.85 | 32 | |
| Cjgssr125_S | 0.50 | 0.48 | 32 | |
| LG6 | BY898881_ES | 0.00 | 1.00 | 32 |
| HS4_rep_c13952_ES | 0.13 | 0.72 | 32 | |
| Cjs1817FC_S | 0.00 | 1.00 | 32 | |
| LG7 | Cjgssr13_S | 0.00 | 0.96 | 32 |
| LG8 | CS1200FC_S | 0.45 | 0.50 | 32 |
| HS4_rep_c17715_ES | 0.00 | 1.00 | 32 | |
| LG9 | S4050_S | 0.14 | 0.71 | 32 |
| LG10 | CJS0201_S | 0.00 | 1.00 | 32 |
| HS4_rep_c39488_ES | 0.27 | 0.61 | 32 | |
| LG11 | Cjgssr124_S | 28.13 | <0.0001 | 32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moriguchi, Y.; Saito, R.; Ueno, S.; Hasegawa, Y.; Kakui, H.; Matsumoto, A. Localization of TWISTED NEEDLES Locus on Linkage Map of Japanese Cedar (Cryptomeria japonica D. Don). Forests 2022, 13, 1524. https://doi.org/10.3390/f13091524
AMA Style
Moriguchi Y, Saito R, Ueno S, Hasegawa Y, Kakui H, Matsumoto A. Localization of TWISTED NEEDLES Locus on Linkage Map of Japanese Cedar (Cryptomeria japonica D. Don). Forests. 2022; 13(9):1524. https://doi.org/10.3390/f13091524
Chicago/Turabian StyleMoriguchi, Yoshinari, Ryunosuke Saito, Saneyoshi Ueno, Yoichi Hasegawa, Hiroyuki Kakui, and Asako Matsumoto. 2022. "Localization of TWISTED NEEDLES Locus on Linkage Map of Japanese Cedar (Cryptomeria japonica D. Don)" Forests 13, no. 9: 1524. https://doi.org/10.3390/f13091524
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.