
Variations in the Diversity and Biomass of Soil Bacteria and Fungi under Different Fire Disturbances in the Taiga Forests of Northeastern China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
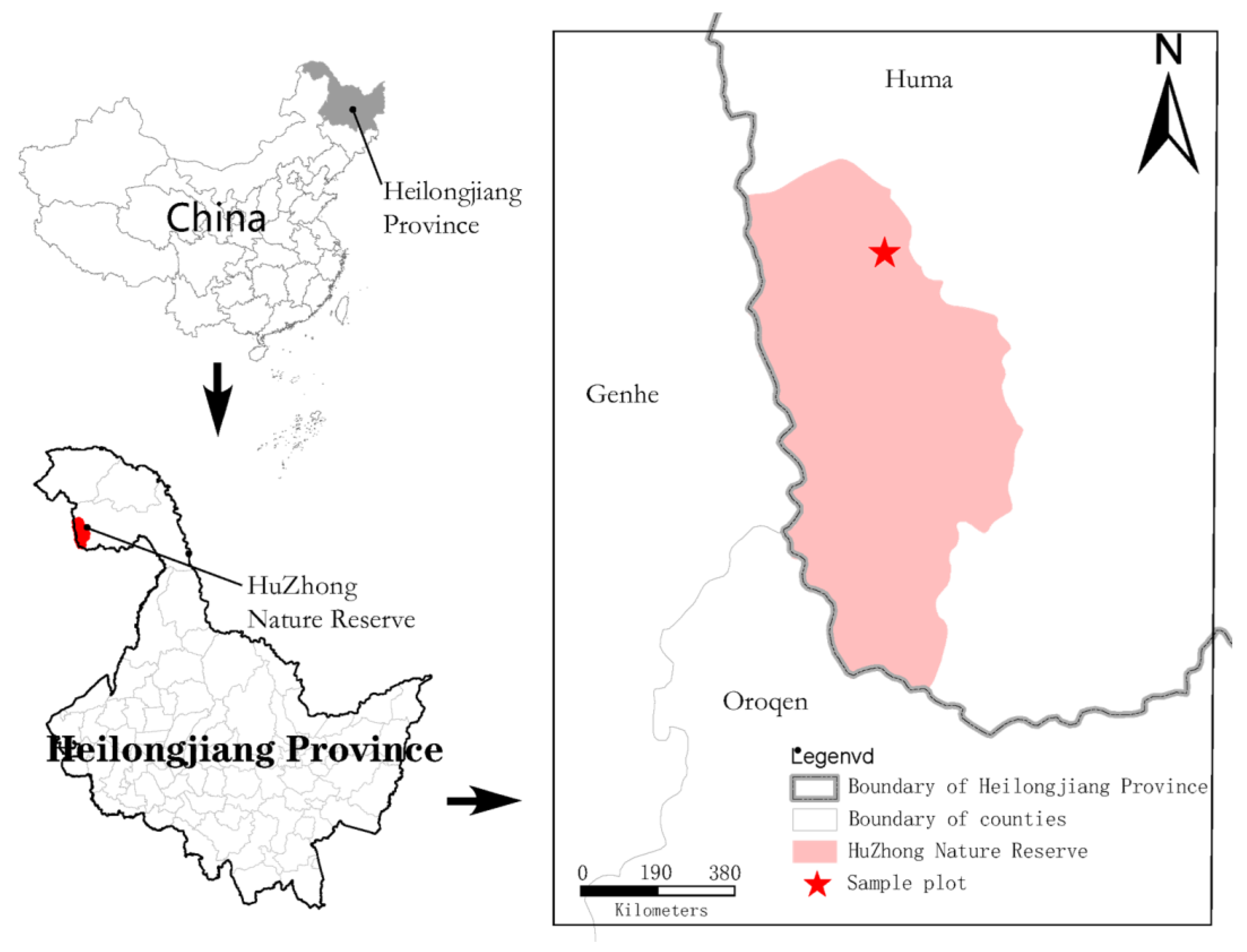
2.1. Sample Sites Description
2.2. Sample Plots
2.3. Soil Sample Collection and Experimental Methods
2.4. Determination of Soil Physicochemical Properties
2.5. Phospholipid Fatty Acid Detection
2.6. Data Analysis
3. Results
3.1. The Soil Physicochemical Parameters of Different Fire Severities
3.2. Differences in Microbial Diversity of the Soils in Fire Sites
3.3. Changes in the Content and Community Structure of Microbial PLFAs in Soils of the Fire Sites
3.4. Correlation Analysis of Factors Affecting Soil Microbial Communities in the Fire Sites
4. Discussion
4.1. Effects of Forest Fires on Soil Physicochemical Properties
4.2. Effects of Forest Fires on Soil Microbial Diversity
4.3. Effects of Forest Fires on the Microbial Composition of Soils
5. Conclusions
- (1)
- Soil microbial biomass carbon (MBC), moisture content (MC), and total nitrogen (TN) all increased significantly in the soil of the investigated fire sites compared to control soil, but the content of available potassium (AK) decreased significantly.
- (2)
- The species diversity of soil microorganisms and the contents of soil Ba, G+, G− and TPLFAs were restored to pre-fire levels after a moderate fire.
- (3)
- Soil MC, TN, SOC, AP and AN being the main influencing factors of the soil microbial community composition.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- German, D.P.; Chacon, S.S.; Allison, S.D. Substrate concentration and enzyme allocation can affect rates of microbial decomposition. Ecology 2011, 92, 1471–1480. [Google Scholar] [CrossRef]
- Hu, H.Q.; Luo, S.S.; Luo, B.Z.; Wei, S.J.; Wang, Z.S.; Wu, Z.P. Effects of forest fire disturbance on soil organic carbon and its components of Cunninghamia lanceolata forest in Guangdong Province, southern China. J. Beijing For. Univ. 2019, 41, 108–118. [Google Scholar] [CrossRef]
- Silva, L.C.R.; Hoffmann, W.A.; Rossatto, D.R.; Haridasan, M.; Franco, A.C.; Horwath, W.R. Can savannas become forests? A coupled analysis of nutrient stocks and fire thresholds in central Brazil. Plant Soil 2013, 373, 829–842. [Google Scholar] [CrossRef]
- Lamont, B.B. Historical links between climate and fire on species dispersion and trait evolution. Plant Ecol. 2022, 223, 711–732. [Google Scholar] [CrossRef]
- Ivanova, G.A.; Ivanov, V.A.; Kovaleva, N.M.; Conard, S.G.; Zhila, S.V.; Tarasov, P.A. Succession of vegetation after a high-intensity fire in a pine forest with lichens. Contemp. Probl. Ecol. 2017, 10, 52–61. [Google Scholar] [CrossRef]
- Cannon, J.B.; Gannon, B.M.; Feinstein, J.A.; Padley, E.A.; Metz, L.J. Simulating spatial complexity in dry conifer forest restoration: Implications for conservation prioritization and scenario evaluation. Landsc. Ecol. 2020, 35, 2301–2319. [Google Scholar] [CrossRef]
- Thammanu, S.; Han, H.; Ekanayake, E.M.B.P.; Jung, Y.; Chung, J. The Impact on Ecosystem Services and the Satisfaction Therewith of Community Forest Management in Northern Thailand. Sustainability 2021, 13, 13474. [Google Scholar] [CrossRef]
- Xiang, X.; Shi, Y.; Yang, J.; Kong, J.J.; Lin, X.J.; Zhang, H.Y.; Zeng, J.; Chu, H.Y. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci. Rep. 2014, 4, 3829. [Google Scholar] [CrossRef]
- Bai, A.Q.; Fu, B.J.; Qu, L.Y.; Huang, Y.Y.; Wang, L.H. The characteristics of soil microbial communities at burned forest sites for the Great Xingan Mountains. Acta Ecol. Sin. 2012, 32, 4762–4771. [Google Scholar] [CrossRef]
- Perez-Valera, E.; Goberna, M.; Faust, K.; Raes, J.; Garcia, C.; Verdu, M. Fire modifies the phylogenetic structure of soil bacterial co-occurrence networks. Environ. Microbiol. 2017, 19, 317–327. [Google Scholar] [CrossRef]
- Lou, H.; Cai, H.Y.; Fu, R.; Guo, C.; Fan, B.Z.; Hu, H.Q.; Zhang, J.; Sun, L. Effects of wildfire disturbance on forest soil microbes and colonization of ericoid mycorrhizal fungi in northern China. Environ. Res. 2023, 231, 116220. [Google Scholar] [CrossRef]
- Qiang, W.; He, L.; Zhang, Y.; Liu, B.; Pang, X. Aboveground vegetation and soil physicochemical properties jointly drive the shift of soil microbial community during subalpine secondary succession in southwest China. Catena 2021, 202, 105251. [Google Scholar] [CrossRef]
- Dooley, S.R.; Treseder, K.K. The Effect of Fire on Microbial Biomass: A Meta-Analysis of Field Studies. Biogeochemistry 2012, 109, 49–61. [Google Scholar] [CrossRef]
- Cui, F.X. Effects of Fire Disturbance on Soil Microbial Diversity and Greenhouse Gas Emissions in Xing’an Larch Forest; Northeast Forestry University: Harbin, China, 2022. [Google Scholar]
- Zhong, C.; Guo, M.; Zhou, F.F.; Li, J.N.; Yu, F.B.; Guo, F.T.; Li, W.S. Forest succession trajectories after fires in valleys and on slopes in the Greater Khingan Mountains, China. J. For. Res. 2023, 34, 623–640. [Google Scholar] [CrossRef]
- Li, X.; Wang, G.P.; Li, Y.H.; Han, D.X.; Cong, J.X.; Gao, C.Y. Aerobic and anaerobic burning alter trace metal availability in peat soils: Evidence from laboratory experiments. Eur. J. Soil Sci. 2023, 74, e13385. [Google Scholar] [CrossRef]
- Guo, X.Y.; Zhang, H.Y.; Wang, Y.Q.; Zhao, J.J.; Zhang, Z.X. The driving factors and their interactions of fire occurrence in Greater Khingan Mountains, China. J. Mt. Sci. 2020, 17, 2674–2690. [Google Scholar] [CrossRef]
- Yang, J.W.; Zhang, Q.L.; Hao, S. Effects of fire disturbance on Larix gmelinii growth-climate relationship. Ecol. Indic. 2022, 143, 109377. [Google Scholar] [CrossRef]
- Frank, A.A.; Reinhardt, E.D. Modeling ignition and burning rate of large woody natural fuels. Int. J. Wildland Fire 1995, 5, 81–91. [Google Scholar] [CrossRef]
- Shang, W.; Wu, X.D.; Zhao, L.; Yue, G.Y.; Zhao, Y.H.; Qiao, Y.P.; Li, Y.Q. Seasonal variations in labile soil organic matter fractions in permafrost soils with different vegetation types in the Central Qinghai-Tibet Plateau. Catena 2016, 137, 670–678. [Google Scholar] [CrossRef]
- Kim, H.M.; Jung, J.Y.; Yergeau, E.; Hwang, C.Y.; Hinzman, L.; Nam, S.; Hong, S.G.; Kim, O.S.; Chun, J.; Lee, Y.K. Bacterial community structure and soil properties of a subarctic tundra soil in Council, Alaska. FEMS Microbiol. Ecol. 2014, 89, 465–475. [Google Scholar] [CrossRef]
- Wear, E.K.; Wilbanks, E.G.; Nelson, C.E.; Carlson, C.A. Primer selection impacts specific population abundances but not community dynamics in a monthly time-series 16S rRNA gene amplicon analysis of coastal marine bacterioplankton. Environ. Microbiol. 2018, 20, 2709–2726. [Google Scholar] [CrossRef] [PubMed]
- Ade, L.J.; Hu, L.; Zi, H.B.; Wang, C.T.; Lerdau, M.; Dong, S.K. Effect of snowpack on the soil bacteria of alpine meadows in the Qinghai-Tibetan Plateau of China. Catena 2018, 164, 13–22. [Google Scholar] [CrossRef]
- Hu, W.G.; Zhang, Q.; Li, D.Y.; Cheng, G.; Mu, J.; Wu, Q.B.; Niu, F.; An, L.Z.; Feng, H.Y. Diversity and community structure of fungi through a permafrost core profile from the Qinghai-Tibet Plateau of China. J. Basic Microbiol. 2015, 54, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Drenovsky, R.E.; Elliott, G.N.; Graham, K.J.; Scow, K.M. Comparison of phospholipid fatty acid (PLFA) and total soil fatty acid methylesters (TSFAME)for characterizing soil microbial communities. Soil Biol. Biochem. 2004, 36, 1793–1800. [Google Scholar] [CrossRef]
- Jain, D.K.; Providenti, M.; Tanner, C.; Cord, I.; Stroes-Gascoyne, S. Characterization of microbial communities in deep groundwater from granitic rock. Can. J. Microb. 1997, 43, 272–283. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Potthoff, M. Microbial react ion in activity, biomass, and community structure after long term continuous mixing of a grassland soil. Soil Biol. Biochem. 2005, 37, 1249–1258. [Google Scholar] [CrossRef]
- Sakamoto, K.; Iijima, T.; Higuchi, R. Use of specific phospholipid fatty acids for identifying and quantifying the external hyphae of the arbuscular mycorrhizal fungus Gigaspora rosea. Soil Biol. Biochem. 2004, 36, 1827–1834. [Google Scholar] [CrossRef]
- Wang, M.; Weng, X.; Zhang, R.; Yang, L.; Liu, Y.; Sui, X. The Diversity and Composition of Soil Microbial Community Differ in Three Typical Wetland Types of the Sanjiang Plain, Northeastern China. Sustainability 2022, 14, 14394. [Google Scholar] [CrossRef]
- Covington, W.W.; Sackett, S.S. Soil mineral nitrogen changes following prescribed burning in ponderosa pine. For. Ecol. Manag. 1992, 54, 175–191. [Google Scholar] [CrossRef]
- Ellingson, L.J.; Kauffman, J.B.; Cummings, D.L.; Sanford, R.L.; Jaramillo, V.J. Soil N dynamics associated with deforestation, biomass burning, and pasture conversion in a Mexican tropical dry forest. For. Ecol. Manag. 2000, 137, 41–51. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Scharenbroch, B.C.; Nix, B.; Jacobs, K.A.; Bowles, M.L. Two decades of low-severity prescribed fire increases soil nutrient availability in a Midwestern, USA oak (Quercus) forest. Geoderma 2012, 183–184, 80–91. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Zhang, L.J.; Ma, B.; Tang, C.X.; Yu, H.D.; Lv, X.F.; Rodrigues, J.L.M.; Dahlgren, R.A.; Xu, J.M. Habitat heterogeneity induced by pyrogenic organic matter in wildfire-perturbed soils mediates bacterial community assembly processes. ISME J. 2021, 15, 1943–1955. [Google Scholar] [CrossRef]
- Dicen, G.P.; Rallos, R.V.; Labides, J.L.R.; Navarrete, I.A. Vulnerability of soil organic matter to microbial decomposition as a consequence of burning. Biogeochemistry 2020, 150, 123–137. [Google Scholar] [CrossRef]
- Mcintosh, P.D.; Laffan, M.D.; Hewitt, A.E. The role of fire and nutrient loss in the genesis of the forest soils of Tasmania and southern New Zealand. For. Ecol. Manag. 2005, 220, 185–215. [Google Scholar] [CrossRef]
- Yue, X.; Tun, X.J.; Cui, D.J.; Guo, X.D.; Song, X.Y.; Liu, L.; Wang, L.L.; Jiang, C.; Xu, D.W.; Li, S.M. Effects of Different Fire Intensities on Forest Soil Nutrients. Shandong Agric. Sci. 2018, 50, 72–76. [Google Scholar] [CrossRef]
- Liu, J.; Qiu, L.P.; Wang, X.; Wei, X.R.; Gao, H.L.; Zhang, Y.J.; Cheng, J.M. Effects of wildfire and topography on soil nutrients in a semiarid restored grassland. Plant Soil 2018, 428, 123–136. [Google Scholar] [CrossRef]
- Heydari, M.; Omidipour, R.; Abedi, M.; Baskin, C. Effects of fire disturbance on alpha and beta diversity and on beta diversity components of soil seed banks and aboveground vegetation. Plant Ecol. Evol. 2017, 150, 247–256. [Google Scholar] [CrossRef]
- Wang, R.; Hu, Y.X.; Khan, A.; Du, L.L.; Wang, Y.; Hou, F.B.; Guo, S.L. Soil prokaryotic community structure and co-occurrence patterns on the fragmented Chinese Loess Plateau: Effects of topographic units of a soil eroding catena. Catena 2021, 198, 105035. [Google Scholar] [CrossRef]
- Weng, X.H.; Sui, X.; Liu, Y.N.; Yang, L.B.; Zhang, R.T. Effect of nitrogen addition on the carbon metabolism of soil microorganisms in a Calamagrostis angustifolia wetland of the Sanjiang Plain, northeastern China. Ann. Microbiol. 2022, 72, 18. [Google Scholar] [CrossRef]
- Ma, Y.; Zu, L.; Long, F.; Yang, X.; Wang, S.; Zhang, Q.; He, Y.; Chen, D.; Sui, M.; Zhang, G.; et al. Promotion of Soil Microbial Community Restoration in the Mu Us Desert (China) by Aerial Seeding. Sustainability 2022, 14, 15241. [Google Scholar] [CrossRef]
- Monleon, V.J.; Cromack, J.K.; Landsberg, J.D. Short-and long term effects of prescribed underburning on nitrogen availability in ponderosa pine stands in central Oregon. Can. J. For. Res. 1997, 27, 369–378. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Artz, R.R.E.; Reid, E.; Anderson, I.C.; Campbell, C.D.; Cairney, J.W.G. Long term repeated prescribed burning increases evenness in the basidiomycete laccase gene pool in forest soils. FEMS Microbiol. Ecol. 2009, 67, 397–410. [Google Scholar] [CrossRef]
- Otsuka, S.; Sudiana, I.; Komori, A.; Isobe, K.; Deguchi, S.; Nishiyama, M.; Shimizu, H.; Senoo, K. Community Structure of Soil Bacteria in a Tropical Rainforest Several Years After Fire. Microbes Environ. 2008, 23, 49–56. [Google Scholar] [CrossRef]
- Peng, S.L.; Ge, Z.W.; Liu, G.C.; Mao, L.F. Environmental drivers of soil microbial activity and diversity along an elevational gradient. J. Mt. Sci. 2022, 19, 1336–1347. [Google Scholar] [CrossRef]
- Liu, R.; Liu, Y.; Gao, Y.; Zhao, F.; Wang, J. The Nitrogen Cycling Key Functional Genes and Related Microbial Bacterial Community α−Diversity Is Determined by Crop Rotation Plans in the Loess Plateau. Agronomy 2023, 13, 1769. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef]
- Hebel, C.L.; Smith, J.E.; Cromack, K. Invasive plant species and soil microbial response to wildfire burn severity in the Cascade Range of Oregon. Appl. Soil Ecol. 2009, 42, 150–159. [Google Scholar] [CrossRef]
- Abreu, C.I.; Friedman, J.; Woltz, V.L.A.; Gore, J. Mortality causes universal changes in microbial community composition. Nat. Commun. 2019, 10, 2120. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liang, C.; Wang, Y.Q.; Cheng, H.; An, S.S.; Chang, S.X. Soil extracellular enzyme stoichiometry reflects the shift from p- to n-limitation of microorganisms with grassland restoration. Soil Biol. Biochem. 2020, 149, 107928. [Google Scholar] [CrossRef]
- Li, B.Y. Effects of Different Fire Intensities on Soil Organic Carbon and Soil Nutrients in Pinus Tabulaeformis Forests in Northern China; Beijing Forestry University: Beijing, China, 2018. [Google Scholar]
- Liang, Q. The Ecological Effects of Forest Fire on Soil and Seed Germination and the Study of Pyrophyte Species in Karst Mountainous Areas of Bijie; Southwest University: Chongqing, China, 2017. [Google Scholar]
- Prayogo, C.; Jones, J.E.; Baeyens, J.; Bending, G.D. lmpact of biochar on mineralisation of C and N from soil and willow litter and its relationship with microbial community biomass and structure. Biol. Fertil. Soils 2014, 50, 695–702. [Google Scholar] [CrossRef]
- Sun, J.; Wang, B.C.; Xu, G. Effects of wheat strawbiochar on carbon mineralization and guidance for largescale soil quality improvement in the coastal wetland. Ecol. Eng. 2014, 62, 43–47. [Google Scholar] [CrossRef]
- Fanin, N.; Haettenschwiler, S.; Fromin, N. Litter fingerprint on microbial biomass, activity, and community structure in the underlying soil. Plant Soil 2014, 379, 79–91. [Google Scholar] [CrossRef]
- Chen, L.Y.; Liu, L.; Qin, S.Q.; Yang, G.B.; Fang, K.; Zhu, B.; Kuzyakov, Y.; Chen, P.D.; Xu, Y.P.; Yang, Y.H. Regulation of priming effect by soil organic matter stability over a broad geographic scale. Nat. Commun. 2019, 10, 5112. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Newcombe, G.; Campbell, J.; Griffith, D.; Baynes, M.; Launchbaugh, K.; Pendleton, R. Revisiting the Life Cycle of Dung Fungi, Including Sordaria fimicola. PLoS ONE 2016, 11, e0147425. [Google Scholar] [CrossRef]
- Ott, A.; Spencer-Phillips, P.T.N.; Willey, N.; Johnston, M.A. Topology: A novel method to describe branching patterns in Peronospora viciae colonies. Mycol. Res. 2003, 107, 1123–1131. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Severity | Grade Index (k·Wm−1) | Burned Dead Wood Ratio | Delineation Criteria |
|---|---|---|---|
| L (Light fire) | 350–750 | <30% | Understory shrub was partially burned (less than 50%) with tree trunk blackening up to 2 m in height |
| M (Moderate fire) | 750–3500 | 30%–70% | The litter layer and half-rot layer was burned, with unchanged color below the half-rot layer. |
| H (Heavy fire) | >3500 | >70% | All undergrowth was burned and tree trunks are blackened to a height of 5 m or more. |
| Microbial Group | Detected PLFA Biomarker |
|---|---|
| Unspecified bacteria (BA) | 12:0, 13:0, 14:0, 15:0, 16:0, 18:0, 22:0, 24:0 |
| Gram-positive bacteria (G+) | 11:0 iso, 11:0 anteiso, 12:0 iso, 12:0 anteiso, 13:0 iso, 13:0 anteiso, 14:1 iso w7c, 14:0 iso, 14:0 anteiso, 15:1 iso w9c, 15:1 iso w6c, 15:1 anteiso w9c, 15:0 iso, 15:0 anteiso, 16:0 iso, 16:0 anteiso, 17:1 iso w9c, 17:0 iso, 17:0 anteiso, 18:0 iso, 19:0 iso, 19:0 anteiso, 20:0 iso, 22:0 is0 |
| Gram-negative bacteria (G−) | 13:1 w5c, 13:1 w4c, 13:1 w3c, 12:0 2OH, 14:1 w7c, 14:1 w5c, 15:1 w9c, 15:1 w8c, 15:1 w7c, 15:1 w6c, 15:1 w5c, 14:02OH, 16:1 w9c, 16:1 w7c, 16:1 w6c, 16:1 w4c, 16:1 w3c, 17:1 w9c, 17:1 w8c, 17:1 w7c, 17:1 w6c, 17:0 cyclo w7c, 17:1 w5c, 17:1 w4c, 17:1 w3c, 16:0 2OH, 18:1 w8c, 18:1 w7c, 18:1 w6c, 18:1 w5c, 18:1 w3c, 19:1 w9c, 19:1 w8c, 19:1 w7c, 19:1 w6c, 19:0cyclo w9c, 19:0 cyclo w7c, 19:0 cyclo w6c, 20:1 w9c, 20:1 w8c, 20:1 w6c, 20:1 w4c, 20:0 cyclo w6c, 21:1 w9c, 21:1 w8c, 21:1 w6c, 21:1 w5c, 21:1 w4c, 21:1w3c, 22:1w9c, 22:1w8c, 22:1 w6c, 22:1 w5c, 22:1 w3c, 22:0 cyclo w6c, 24:1 w9c, 24:1 w7c |
| Unspecified Fungi (FU) | 18:1ω9c, 23:0 |
| Sample | MBC mg/kg | MC | pH | AK mg/kg | TN g/kg | SOC g/kg | AP mg/kg | AN mg/kg |
|---|---|---|---|---|---|---|---|---|
| CK | 149.97 ± 33.56 b | 0.04 ± 0.004 g | 6.79 ± 0.03 a | 213.05 ± 19.19 a | 1.69 ± 0.01 d | 97.05 ± 3.41 e | 11.74 ± 0.69 e | 96.14 ± 11.33 b |
| 00L | 1092.24 ± 267.27 a | 0.47 ± 0.013 b | 6.11 ± 0.02 bc | 9.33 ± 0.87 d | 3.28 ± 0.52 c | 193.22 ± 2.42 c | 6.3 ± 0.69 f | 253.89 ± 20.4 a |
| 00M | 1143.35 ± 94.89 a | 0.6 ± 0.054 a | 6.7 ± 0.21 a | 10.04 ± 1.4 d | 4.84 ± 0.06 a | 304.86 ± 0.84 a | 22.6 ± 1.18 c | 300.73 ± 74.27 a |
| 00H | 849.63 ± 94.06 a | 0.42 ± 0.014 c | 6.36 ± 0.41 abc | 24.94 ± 1.46 d | 4.28 ± 0.14 b | 235.56 ± 4.7 b | 11.15 ± 1.16 e | 285.21 ± 54.59 a |
| 10L | 268.37 ± 23.67 b | 0.16 ± 0.002 e | 6.64 ± 0.02 ab | 140.07 ± 5.2 b | 1.95 ± 0.06 d | 122.98 ± 5.96 d | 46.9 ± 0.83 a | 89.37 ± 12.3 b |
| 10M | 847.07 ± 369.5 a | 0.12 ± 0.006 f | 6.53 ± 0.05 ab | 80.43 ± 6.38 c | 1.71 ± 0.06 d | 93.16 ± 6.49 e | 18.21 ± 0.66 d | 82.13 ± 24.22 b |
| 10H | 349.99 ± 12.5 b | 0.27 ± 0.03 d | 5.84 ± 0.4 c | 80.97 ± 11.17 c | 2.96 ± 0.43 c | 76.06 ± 3.24 f | 27.48 ± 1.57 b | 148.16 ± 17.29 b |
| Sample | Shannon | Simpson | Menhinick | Margalef |
|---|---|---|---|---|
| CK | 3.41 ± 0.04 ab | 0.95 ± 0.00 a | 27.7 ± 0.35 d | 40.47 ± 0.35 d |
| 00L | 3.45 ± 0.04 a | 0.95 ± 0.00 a | 36.24 ± 2.04 a | 49.65 ± 2.37 a |
| 00M | 3.47 ± 0.01 a | 0.95 ± 0.00 a | 28.3 ± 0.6 d | 41.08 ± 0.61 d |
| 00H | 3.47 ± 0.03 a | 0.95 ± 0.00 a | 33.4 ± 1.22 b | 46.45 ± 1.33 b |
| 10L | 3.22 ± 0.02 c | 0.93 ± 0.00 b | 25.12 ± 0.8 e | 37.93 ± 0.78 e |
| 10M | 3.18 ± 0.01 c | 0.93 ± 0.00 b | 27.17 ± 0.12 d | 39.95 ± 0.12 d |
| 10H | 3.37 ± 0.06 b | 0.95 ± 0.00 a | 30.8 ± 0.39 c | 43.65 ± 0.4 c |
| Index | MBC | MC | pH | AK | TN | SOC | AP | AN |
|---|---|---|---|---|---|---|---|---|
| Shannon | 0.30 | 0.64 ** | −0.20 | −0.39 | 0.71 *** | 0.62 ** | −0.58 ** | 0.76 *** |
| Simpson | 0.20 | 0.50 * | −0.34 | −0.30 | 0.58 ** | 0.39 | −0.64 ** | 0.63 ** |
| Menhinick | 0.50 * | 0.55 ** | −0.54 * | −0.61 ** | 0.50 * | 0.32 | −0.67 *** | 0.57 ** |
| Margalef | 0.50 * | 0.55 * | −0.53 * | −0.61 ** | 0.49 * | 0.32 | −0.66 ** | 0.56 ** |
| Soil Factor | r2 | p |
|---|---|---|
| MBC | 0.2754 | 0.055 |
| MC | 0.6615 | 0.001 *** |
| pH | 0.138 | 0.248 |
| AK | 0.3967 | 0.016 * |
| TN | 0.6837 | 0.001 *** |
| SOC | 0.5454 | 0.003 ** |
| AP | 0.8098 | 0.001 *** |
| AN | 0.7259 | 0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Z.; Wu, S.; Du, J.; Pan, H.; Lu, X.; Liu, Y.; Yang, L. Variations in the Diversity and Biomass of Soil Bacteria and Fungi under Different Fire Disturbances in the Taiga Forests of Northeastern China. Forests 2023, 14, 2063. https://doi.org/10.3390/f14102063
Cheng Z, Wu S, Du J, Pan H, Lu X, Liu Y, Yang L. Variations in the Diversity and Biomass of Soil Bacteria and Fungi under Different Fire Disturbances in the Taiga Forests of Northeastern China. Forests. 2023; 14(10):2063. https://doi.org/10.3390/f14102063
Chicago/Turabian StyleCheng, Zhichao, Song Wu, Jun Du, Hong Pan, Xinming Lu, Yongzhi Liu, and Libin Yang. 2023. "Variations in the Diversity and Biomass of Soil Bacteria and Fungi under Different Fire Disturbances in the Taiga Forests of Northeastern China" Forests 14, no. 10: 2063. https://doi.org/10.3390/f14102063