
The Potential of Paulownia fortunei L. for the Phytoremediation of Pb
by

Lu Du
1,†,
Hang Yang
1,†,
Juan Xie
1,
Liangze Han
1,
Zhiyi Liu
1,
Zhiming Liu
1,2,
Yonghua Chen
1,* and
Rongkui Su
1 1
College of Environmental Science and Engineering, Central South University of Forestry and Technology, Changsha 410004, China
2
Department of Biology, Eastern New Mexico University, Portales, NM 88130, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Forests 2023, 14(6), 1245; https://doi.org/10.3390/f14061245
Submission received: 29 March 2023
/
Revised: 8 May 2023
/
Accepted: 10 May 2023
/
Published: 15 June 2023
(This article belongs to the Special Issue Soil Contamination in Forest Ecosystem)
Abstract
:Pb endangers forest ecological health; phytoremediation is an effective Pb remediation technology. Woody plants with Pb tolerance provided a mechanism for the phytoremediation of Pb. Paulownia fortunei (L.), a fast-growing woody plant, has a good tolerance to Pb. However, its tolerance mechanism is unclear. The results in this study revealed that P. fortunei seedlings can withstand 400 mg·L−1 Pb stress. The quantification of Pb in different P. fortunei tissues showed an increasing trend of accumulation in root > leaf > stem; the transport coefficient and enrichment coefficient decreased with an increase in Pb concentration. The tolerance of P. fortunei to Pb may be related to cell partition and immobilization by the cell wall. Microstructural analysis performed using scanning electron microscopy showed that the absorbed Pb is mainly distributed in cell wall components, and when the concentration of Pb increases, it can be transferred to soluble parts and organelles. The Fourier transform infrared spectrometry results showed that excess hydroxyl groups occurred under Pb stress in the outer epidermis cell walls of roots and leaves adsorbing heavy metals. When the concentration of Pb was over 400 mg·L−1, the growth of P. fortunei was inhibited, the root cell wall was deformed, the plasmolysis occurred in the cauline cell, and the internal leaf capsule was ruptured. Furthermore, antioxidant enzyme activity was significantly reduced. Therefore, P. fortunei can transfer the underground part of Pb to the aboveground part up to the concentration of 400 mg·L−1. This study provides a theoretical basis and technical reference for fully utilizing woody plant resources to restore the ecological environment of forests.
1. Introduction
Lead (Pb) is a highly toxic heavy metal pollutant that has long-term persistence and can accumulate to high concentrations in soil. Its toxic effects can have a devastating impact on single organisms and the entire ecosystem [1]. It is considered to be one of the most hazardous heavy metal pollutants in the environment [2]. Even low doses of Pb exposure can result in severe mental environmental impairment and increase the likelihood of depression and anxiety [3,4]. Pb is hazardous to plants, disrupting their defense mechanisms, as well as photosynthesis [5]. Peroxidase (POD), superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX) can significantly improve the redox homeostasis of cells under Pb stress [6]. The toxicity of ROS produced by excessive Pb can disturb the redox balance of plants [7]. Forests play a vital role in national ecological security, with several key functions, such as maintaining soil health, protecting biodiversity, regulating the climate, and promoting sustainable development of the ecosystem [8]. Compared with agricultural soil, Pb has caused a low risk to the human food chain; thus, heavy metal pollution in forest soil receives less attention. Recent studies showed that the migration rate of heavy metal in forest soil is higher than that in agricultural land under the same conditions, which may not only increase the risk of heavy metals in the groundwater and surface water, but also increase the damage to the forest ecosystem [9]. Therefore, there is an urgent need for effective and appropriate remediation tools to remediate heavy metal contamination [10].
Phytoremediation is a good way to restore the environment. The benefits of phytoremediation are its low cost, high efficiency, environmental protection, and ecological friendliness [11]. The phytoremediation of heavy metal-contaminated soil includes plant extraction, plant degradation, nodule filtration, and plant stabilization [12]. Previous studies on heavy metal phytoremediation have mainly focused on super-cumulative plants for boosting plant extraction; the study by Mohtadi, Ghaderian indicated that calamine populations of Silene vulgaris and Noccaea caerulescens were hyper-tolerant to Pb, could survive in environments with a relatively high Pb concentration, and could effectively extract Pb [13,14]. Hyperaccumulation plants are mostly low-biomass annual herbaceous plants, which are dwarfed and have underdeveloped primary root systems, making them difficult to be used in large-scale restoration projects and limiting their application in phytoremediation [15]. Compared with herbaceous plants, many woody plants have developed root systems, are rich in biomass and easy to harvest, can grow rapidly in poor soil environments, have some tolerance and accumulation capacity [16], and have become alternatives to hyperaccumulation plants [17].
The research of plant tolerance to heavy metals is the key problem of phytoremediation by woody plants. Excessive absorption of Pb induces ROS production in plants, which disturbs the redox balance of plants [7]. According to the research conducted by Kong and Li [18], high levels of Pb hinders chlorophyll synthesis in Rhodiola rosea, resulting in a reduction in both the plant’s chlorophyll content and its photosynthetic rate. Therios [19] studied the effect of Pb on photosynthesis and the antioxidant activity in Citrus aurantium L. leaves, and found that Pb toxicity inhibits antioxidant activity, interferes with mineral nutrition and water balance within the plant, and disrupts photosynthetic mechanisms. However, Pb does not always exert a harmful effect [18]. The germination vigor and speed of the seeds were marginally higher than those of the control group at the135 mg·L−1 Pb concentration level, suggesting that a specific concentration of Pb could stimulate the germination of Rabdosia rubescens seeds. Kong Si-Xin and others treated the seeds of Rabdosia rubescens with various concentrations of Pb solution. Yang [20] stressed three varieties of eucalyptus with different concentrations of Pb. The obtained results found that in the presence of Pb treatment, a variety grew well without any toxic symptoms of leaf necrosis and growth retardation, supporting a species-dependent response to heavy metal stress. Recently, the detection of Pb in forest soils [21,22] has become a growing concern for domestic and international researchers.
Paulownia fortunei L. is a heavy metal-tolerant woody plant and has a certain remediation effect on Zn [23]. However, little is known about the response mechanism of P. fortunei to Pb stress. Consequently, this study explored the physiological and biochemical response of P. fortunei seedlings under Pb stress; the subcellular distribution and chemical morphology of Pb in the roots, stems, and leaves; alterations in the cell microstructure; and the heavy metal microdistribution and functional group composition.
2. Materials and Methods
2.1. Experimental Design
The experiment was conducted in the forest nursery of the Central South University of Science and Technology. P. fortunei (L.) with similar growth characteristics were selected and transplanted into fine sand soaked in 2% nitric acid overnight and cleaned, watered with 500 mL 1/2 Hoagland nutrient solution every 2 days, incubated for 1 week, and treated with different concentrations of Pb in the form of Pb (NO3)2. The plants were irrigated once every 2 days with a nutrient solution containing different Pb concentrations (50, 100, 200, 400, 600, or 800 mg·L−1). An equivalent nutrient solution without heavy metals was used for the control group. After 30 days of stress, plants were harvested for measurement. Each of the 7 treatments consisted of 6 biological replicates (i.e., 42 plants in total).
2.2. Experiment Method
2.2.1. Plant Growth Parameters
The biomass was measured by dividing the plants into three parts: roots, stems, and leaves. Samples were washed with tap water and dried in a clean envelope. Thus, samples were oven dried at 75 °C and used for dry biomass determination and Pb measurements. Plant height was measured from the root neck to the top of the stem using a tape. Root architecture was determined using a WinRHIZO PRO root scanner (Regent Instrument, Québec, QC, Canada), measuring the root length, root surface area, average root diameter, and root tip number.
2.2.2. Plant Physiological and Biochemical Measures
The pigment concentrations (i.e., chlorophyll a, b, and carotenoids) in plant leaves were determined using a UV-5100 UV-visible spectrophotometer, following an extraction using 95% Ethanol, for 3~5 min at 25 °C in dark [24].
For the experiment, three P. fortunei (L.) trees were randomly selected: three normal growth and leaf positions were selected for leaves on the same plant for the measurement; five parallel leaves were used for each treatment. The content of malondialdehyde (MDA) and the activity of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) in the leaves was measured following the methods reported in [25].
2.2.3. Pb Content
The concentration of Pb in the plants and soil was measured with a Flame Atomic Absorption Spectrophotometer (FAAS, AA-7002, Thermo Fisher Scientific, Waltham, MA, USA). The Pb content in plants was determined using the “HNO3–HClO4 wet method” according to Mohtadi et al. [14], and the soil Pb content was determined using the “aqua regia-perchloric acid hot plate method” according to Deng et al. [26]. The bioconcentration factor (BCF) was calculated as the ratio of the Pb concentration in plants to that in the soil; the transport factor (TF) was calculated in terms of the concentration of Pb in the aboveground part of plants compared to that in the underground part of plants; and the transfer quantity factor (TQ) was calculated as the total content of heavy metal in the shoot divided by that in the whole plant.
2.2.4. Subcellular Distribution and Chemical Forms of Pb
Fresh roots, stems, and leaves were washed with distilled water and homogenized with quartz sand and extraction buffer (250 mM sucrose, 1.0 mM dithioerythritol, and 50 mM Tris-HCl adjusted to pH 7.5). Subcellular fractions of Pb in roots, stems, and leaves were separated by centrifugation as follows: the cell wall fraction was separated to the bottom at 2000× g for 1 min; the supernatant solution was further centrifuged at 10,000× g for 30 min to obtain an organelle fraction in precipitate. The soluble fraction remained in the supernatant solution. All procedures were performed at 4 °C. The chemical forms of Pb were extracted following the method reported in [27]. Pb chemical forms were extracted through the following steps: (1) inorganic forms were extracted using 80% ethanol; (2) organic forms were extracted using H2O; (3) pectates and protein forms were extracted using 0.6 mol·L−1 NaCl; (4) undissolved phosphate forms were extracted using 2% HAc; (5) oxalate forms were extracted using 0.6 mol·L−1 HCl; and finally, (6) the residual Pb form was left. The contents of Pb in different subcellular fractions were quantified by AAS.
2.2.5. Plant Microstructure and Functional Group Composition
Microstructures and functional groups in plants were processed following Han’s method [28]: briefly, 2 mm × 2 mm × 1 mm samples of fresh roots, stems, and leaves were stored in 2.5% glutaraldehyde solution at 4 °C until the microstructural analysis was performed using a Sigma 500 emission scanning electron microscope (SEM, ZEISS, Oberkochen, Baden-Wurtberg Germany). Roots, stems, and leaves were heated at 105 °C for 30 min and dried at 75 °C for 12h, then ground into powder through a 200-mesh sieve. The composition of functional groups of each sample was entrusted to Shanghai Huiming Testing Company for determination using a Fourier-Transform Infrared Spectrometer (FTIR, Nicolet-460, Nicolet, Madison, Wisconsin, USA) within the range 400–4000 cm−1.
2.3. Data Analysis
SPSS 22.0 statistical software (IBM, Armonk, New York, USA) was used for significance testing and correlation analysis. Significance tests were performed using the independent-samples t-test (Student’s t-test) and one-way analysis of variance (ANOVA, UK). When the variance of the one-way ANOVA was not homogeneous, Tukey’s multiple comparison method was used, and p < 0.05 was considered as a significant difference. Origin 2017 software was used to draw the graphs.
3. Results
3.1. Influence of Pb Stress on the Growth of P. fortunei Seedlings
The plant height, total biomass, root length, root surface area, and root tip number of P. fortunei increased significantly (p < 0.05) with increasing Pb concentrations (Table 1). Compared with the Pb-free group, the values of those indexes in the 400 mg·L−1 stress group increased by 50%, 346%, 32%, 106%, 633%, and 89%. However, when the Pb concentration was higher than 400 mg·L−1, the above values decreased significantly (p < 0.05). Moreover, the root diameter of P. fortunei increased in the face of Pb stress.
3.2. Pb Transport and Accumulation in P. fortunei Seedlings
Pb concentrations in the roots, stems, and leaves of P. fortunei increased with increasing Pb concentrations: the Pb content varied from 210.7 to 1600.2 mg·kg−1 in the root, from 6.3 to 386.7 mg·kg−1 in the stems, and from 12.6 to 196.5 mg·kg−1 in the leaves (Table 2). Pb was mainly fixed in the roots. The enrichment coefficient of Pb generally decreased with the increase in Pb concentration (p < 0.05), from 0.18 at 50 mg·L−1 to 0.06 at 800 mg·L−1, and the enrichment coefficients of Pb were all less than 0.2, indicating a weak enrichment ability of P. fortunei for Pb. Moreover, the transfer coefficient of Pb in each group increased and then decreased with the increase in Pb concentration; the maximum transfer coefficient was 0.42 at 600 mg·L−1.
3.3. Changes in Pb Contents in the Subcellular Components of P. fortunei Seedlings
According to the content of subcellular heavy metals in organs of P. fortunei seedlings (Table 3), the contents of subcellular Pb in the roots, stems, and leaves increased with the increase in Pb concentration in the solution. In this study, most Pb was stored in the cell wall component (F1), followed by the soluble component (F3), and the content of heavy metals in the organelle and nuclear component (F2) was the least. These results indicate that P. fortunei reduced the toxicity of Pb to itself by storing Pb in the cell wall and soluble components. The changes in Pb concentrations in the subcellular components of P. fortunei roots, stems, and leaves were also different. With the increase in Pb concentration in the solution, the Pb concentration in the cell wall fractions of the roots, stems, and leaves also increased, but its ratio to the total content decreased. In contrast, when the content of Pb exceeded a certain concentration, the proportion of nuclear, organelle components and soluble components in the roots, stems, and leaves increased. The results indicated that soluble components were the main site for chelating Pb.
3.4. Changes in the Chemical Morphology of Pb in P. fortunei Seedlings
In the roots and stems, Pb phosphate was found to be the main chemical form of Pb. In the leaves, lead oxalate was the dominant chemical form. With the increase of the Pb concentration, the percentage of pectin- and protein-bound Pb, water-insoluble Pb phosphate, and Pb oxalate decreased (Figure 1); the Pb phosphate decreased the most, from 52%, 48%, and 32% to 37%, 37%, and 19% in the roots, stems, and leaves, respectively. The percentages of Pb inorganic matter and Pb organic matter increased, among which the Pb inorganic matter state increased the most, from 8%, 5%, and 6% to 18%, 12%, and 14% in the roots, stems and leaves, respectively. There was no significant pattern of change in the residue state.
The percentage of Pb inorganic, Pb organic, and pectin-protein-bound Pb in P. fortunei showed an increasing trend with an increasing Pb concentration: oxalic acid Pb and residual Pb both decreased with an increasing Pb concentration; phosphate Pb also increased with an increasing Pb concentration, but the largest percentage was at 600 mg·L−1 (Figure 1). The results showed (Table S1) that as the concentration of Pb stress increased, the percentage of the Pb organic state in the tissues of P. fortunei increased significantly, and the difference between Pb treatment groups was significant (p < 0.05).
3.5. Physiological and Biochemical Effects of Pb Stress on P. fortunei Seedlings
With the increase in Pb concentration, the values of total chlorophyll, chlorophyll a, chlorophyll b, carotenoids, and chlorophyll a/b in leaves showed a trend of increasing first and then decreasing (Table 4). The total chlorophyll content reached the maximum value when the added Pb concentration was 400 mg·L−1, which was significantly increased by 45% compared with the control group (p < 0.05). When the added Pb concentration was increased to 800 mg·L−1, the total chlorophyll content reached the lowest value, which was significantly decreased by 57% compared with the control group (p < 0.05). The results indicated that low concentrations of Pb promoted chlorophyll synthesis, but the chlorophyll content decreased significantly with an increase in Pb. The chlorophyll a/b decreased rapidly when the concentration of Pb was higher than 400 mg·L−1.
The activities of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) showed a trend first of increasing and then decreasing with the increase in Pb concentration in the solution (Figure 2). When the Pb concentration was 400 mg·L−1, the SOD, CAT, and POD activities reached their maximum values, which were significantly increased by 24%, 57%, and 97%, respectively, compared with the control group (p < 0.05). The content of MDA in leaves increased significantly with the increase in Pb concentration (p < 0.05), and the content of MDA in the Pb 800 treatment group was 378 nmol·mg−1, 1.72 times greater than that of the control group.
3.6. Characterization of Roots, Stems, and Leaves of P. fortunei Seedlings under Pb Stress
Plants showed normal growth in the control group, with an intact cell wall and membrane structures of the root, stem, and leaves, osmiophilic granules, and thylakoids evenly distributed around starch granules, and no obvious damage in organelles, as highlighted by electron microscopy images (Figure 3A,D,H) of the control group. When the concentration was 400 mg·L−1, P. fortunei roots (Figure 3B), stems (Figure 3E), and leaves (Figure 3I) were destroyed, black particles attached to the cell wall and organelle, and cracks began to appear between cells. When the concentration was 800 mg/L, the cell contours in P. fortunei roots (Figure 3C), stems (Figure 3F), and leaves (Figure 3G) were hard to identify, and cells were enriched with a large number of black particles and were seriously wrinkled or even deformed. The thylakoid in the leaves was ruptured, and extravasation phenomena appeared.
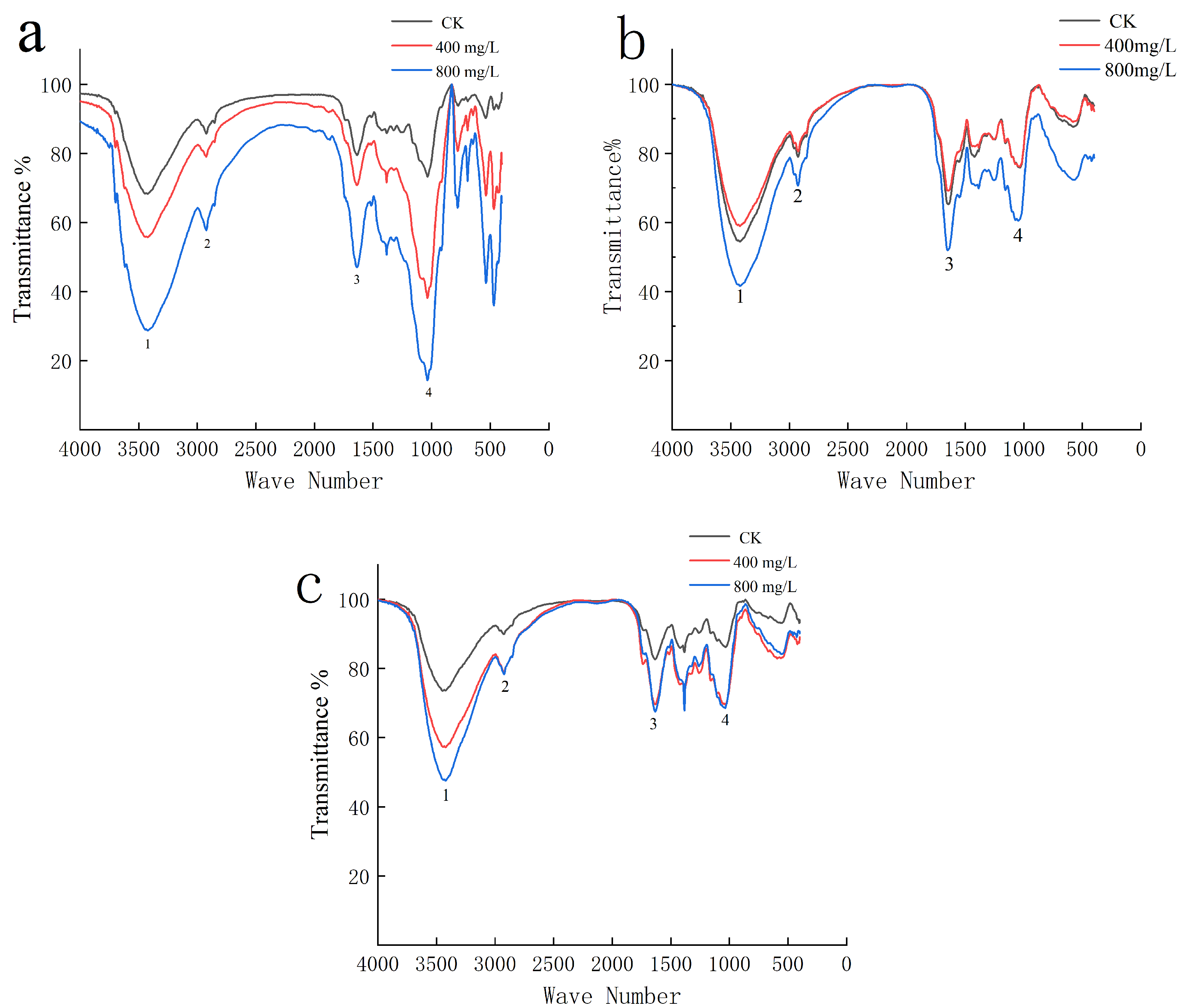
The functional group composition changes in the roots, stems, and leaves of P. fortunei seedlings exposed to Pb treatments were examined by FTIR (Figure 4). Compared with the control group, substantial changes in the transmittance rate at distances of 3420 cm−1, 2920 cm−1, 1630 cm−1, and 1040 cm−1 were found.
Absorption peaks in the 3420 cm−1 range were caused by the free hydroxyl group of carbohydrates, such as sugar, cellulose, and hemicellulose; the absorption peaks of the treated group roots and leaves were higher. Furthermore, the absorption peaks of roots and leaves treated with 800 mg·L−1 Pb were even higher than those observed in the group treated with 400 mg·L−1 Pb. The excess hydroxyl could not be eliminated and accumulated in the P. fortunei, resulting in serious membrane lipid peroxidation. The stem experienced less Pb stress than the roots and leaves. As a result, the stem treated with 400 mg·L−1 Pb exhibited a lower absorption peak than the control group, proving that the low Pb content in the stem had no impact on the binding of free hydroxides to heavy metals. The absorption peak of the 800 mg·L−1 Pb treatment group was higher than that of the control group, indicating that the high concentration of Pb stress prevented free hydroxyl groups in the stem from combining with Pb in a normal manner, leading to excessive buildup of hydroxyl groups in the plant.
Absorption peaks in the 2920 cm−1 range were caused by O-H carboxylic acid and C-H methyl saturated in the cell wall and biofilm. The absorption peaks in the treatment group’s roots and leaves were higher than in the control group, indicating that P. fortunei, which experienced Pb stress, encouraged the formation of organic acids in roots and leaves, de-chelating heavy metals and increasing the amount of O-H carboxylic acid. The 800 mg·L−1 Pb treatment group’s P. fortunei stem absorption peak was higher than that of the 400 mg·L−1 treatment group, indicating that the high concentration of Pb stress caused the P. fortunei stem to create more organic acids to chelate Pb, which increased the level of O-H carboxylic acid.
The absorption peak near 3420 cm−1 came from the carboxyl group of amino acids and the amino group of proteins. Compared with the control group, the absorption peaks of roots and leaves in the treatment group were enhanced, indicating that under Pb stress, P. fortunei promoted the production of large amounts of amino acids and proteins by roots and leaves, which chelated Pb, increasing the amino and carboxyl groups. The absorption peak of the P. fortunei stem in the 800 mg L−1 Pb treatment group was enhanced compared with that in the 400 mg·L−1 treatment group, indicating that the highest concentration of Pb induced an in vivo increase in protein amino acids to protect cells, increasing the carboxyl groups and amino groups.
The C-O group of carbohydrates was responsible for the absorption peak near the observed 1040 cm−1. The absorption peaks of roots and leaves in the treatment group were higher than in the control group in this study, indicating that under Pb stress, P. fortunei encourages the accumulation of carbohydrates in roots and leaves that act as ligands, to which the heavy metals can bind, leading to an increase in C-O groups. The absorption peak of P. fortunei stems was higher in the 800 mg·L−1 Pb treatment group compared with the 400 mg·L−1 group, again indicating an accumulation of carbohydrates.
4. Discussion
In this study, the effect of Pb stress on the growth of P. fortunei showed a trend of initially promoting growth at low concentrations and inhibiting growth at high concentrations. P. fortunei was found to tolerate Pb concentrations up to 400 mg/L. However, at concentrations above this threshold, Pb had a clear toxic effect on P. fortunei by reducing the biomass, damaging the root system, and inhibiting growth. However, the accumulation and translocation coefficients decreased with the increasing Pb concentration, indicating that when the Pb concentration is too high, it limits the translocation, as also found in Lolium perenne L. by Feng et al. [29]. Pb was mainly concentrated in the roots, indicating that P. fortunei, similarly to tolerant woody plants, principally accumulate heavy metals in this organ. This is consistent with the research results obtained by Ma et al. [30], who concluded that the heavy metal concentration in the root system was higher than that in the aboveground part of the plant [30].
Pb was mainly concentrated in the cell wall and the soluble fraction, and in vesicles, whereas its concentration was lower in organelles, which is consistent with the findings of Basile [31] and Zhu [32], who also reported fixation on the cell wall and compartmentalization in the soluble fraction as important detoxification mechanisms for plant tolerance to heavy metals [33]. The Pb concentration in the cell wall was higher than that in the soluble fraction, indicating that the cell wall is the main barrier against the entry of heavy metals into the cell, inhibiting Pb translocation in plants [34]. As the concentration of Pb increased, the percentage of soluble fraction increased and the percentage of cell wall decreased, indicating that high concentrations of Pb caused damage to the cell wall, which led to an increase in the amount of Pb entering the soluble fraction. Therefore, it can be speculated that the protective role of the cell wall is crucial for plants at low Pb concentration stress, and compartmentalization of the soluble fraction becomes a key factor for plant protection at high Pb concentrations; similar results were found by Rongkui Su et al. for Koelreuteria paniculate [35].
Pb can exist in plants in many chemical forms, and there are significant differences in the transport capacity and activity of different forms of Pb. The chemical forms of Pb differ among different tissues of P. fortunei. In this experiment, Pb phosphate was the main chemical form in the roots and stems. In the leaves, Pb oxalate was the main chemical form. Similar results were reported by Cheng and Zhang [36], who found that the Pb forms in maize roots and shoots were mainly Pb phosphate and Pb oxalate, and values in the roots were slightly higher than those in the shoots. Indeed, the Pb in roots and shoots were mainly Pb phosphate and Pb oxalate; these values were slightly higher in the roots than in the shoots. Phosphate Pb was the predominant Pb chemical form in P. fortunei. We found that when the Pb concentration was low, Pb existed mainly in the weakly active form in the seedlings of P. fortunei; as the Pb concentration increased, the proportion of pectin- and protein-bound Pb, Pb inorganic, phosphate Pb, and Pb organic increased, while the proportion of oxalate Pb and residual Pb decreased. Indeed, under high Pb stress, the ability of plants to bind heavy metals is reduced, and Pb is present in a more active form, which can cause severe toxicity to plants [37].
The physiological and biochemical responses of plants to Pb reflect the activation of detoxification processes [38]. As a pigment closely related to photosynthesis, the change of chlorophyll content is, therefore, one of the important indicators of physiological changes in leaves. Indeed, if the concentration of external heavy metals reaches the critical value of plant growth, this will be affected. In this experiment, the total chlorophyll, chlorophyll a, chlorophyll b, and carotenoid contents of leaves showed a trend of increasing and then decreasing with the increase of Pb concentration, reaching the maximum value under 400 mg·L−1 Pb, which was similar to the findings of Zhu and Cheng [39], who considered that a low concentration of heavy metals can promote an increase in chlorophyll content to a certain extent. With the increase in heavy metal concentration, the chloroplast enzyme activity was inhibited and the chlorophyll content was reduced [40]. At the same time, when the concentration of Pb was higher than 400 mg·L−1, the value of chlorophyll a/b decreased rapidly. This phenomenon suggests that within this concentration range, Pb stress accelerated leaf aging and inhibited plant growth.
The toxicity of heavy metals to plants is mainly demonstrated through the plants’ production of excessive reactive oxygen free radicals, resulting in protein inactivation, membrane lipid peroxidation, and, finally, MDA formation [41]. The antioxidant system is an important pathway for plants to eliminate excess active free radicals. In this experiment, the SOD, CAT, and POD activities first increased and then decreased with the increase in Pb concentration. This is because the antioxidant defense system can eliminate the active free radicals induced by low concentrations of Pb (<400 mg/L) in time and maintain the normal metabolism of plant free radicals [42]. The activities of antioxidant enzymes will be inhibited and decrease sharply when the concentration in Pb is too high. At the same time, MDA increases with the increase of the Pb concentration, which indicates that Pb stress causes membrane lipid peroxidation, inducing an imbalance of intracellular osmotic pressure and inhibiting growth [19].
STEM results showed that the main toxic effect of Pb on roots is the destruction of the cell wall structure and the presence of black particles attached to the cell wall, speculated to be Pb-containing substances or Pb-induced metabolites. It has been proven that the cell wall can restrict the absorption of Pb into roots and act as an important barrier to protect root cells, as confirmed in [43]. The main toxic effects of Pb on the stem were concentrations of inclusion in the cell membrane and separation of the plasma wall. The main toxic effects of Pb on leaves were severe shrinkage or the deformation of cells and of thylakoid rupture. The results of the FTIR analysis showed that Pb could induce the excessive production of free hydroxyl groups. In addition, Pb stimulated the production of organic acids, amino acids, and carbohydrates to resist Pb stress [44]. Different organs of P. fortunei exhibit different responses to Pb. Roots and leaves can induce excessive hydroxyl radicals, amino acids, organic acids, and other substances when exposed to Pb stress. When exposed to low concentration of Pb, organic acids and amino acids in the stem of P. fortunei plants can effectively bind Pb and reduce the damaging effect of Pb on P. fortunei cells. However, under high concentrations of Pb, organic acids and amino acids in the stems of P. fortunei are unable to bind Pb, resulting in the excessive accumulation of this heavy metal in the stems, thus inhibiting the growth.
5. Conclusions
P. fortunei L. exhibits a degree of tolerance to Pb and tends to benefit from low levels of Pb stress; however, P. fortunei shows signs of toxicity and growth inhibition at concentrations exceeding 400 mg/L. When the Pb concentration was lower than 400 mg·L−1, Pb was fixed in the root cell wall in a less active and migratory form, reducing its transport to aboveground and alleviating stress suffered by the plant. At the same time, the low concentration of Pb promoted an increase in chlorophyll content to some extent. When the Pb treatment was greater than 400 mg·L−1, the content of Pb in the root cell wall fractions decreased. A high concentration of Pb inhibited the chloroplast enzyme activity and reduced the chlorophyll content, thus accelerating leaf aging and inhibiting plant growth. The proportions of Pb present in the tissues of P. fortunei as migratory and toxic inorganic salts and organic acid salts increased, indicating that a high concentration of Pb stress reduced the ability of the cell wall to fix Pb, which led to the entry of Pb into the interior of the cell and damage to the cell structure, thus increasing its highly active and toxic form and enabling entry to the aboveground part of the plant, causing tissue damage.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/1999-4907/14/6/1245/s1, Table S1: Percentage of different forms of Pb in P. fortunei.
Author Contributions
Conceptualization, Y.C. and R.S.; Methodology, L.D.; Formal analysis, H.Y.; Investigation, H.Y.; Data curation, J.X., L.H., Z.L. (Zhiyi Liu) and Z.L. (Zhiming Liu); Writing—original draft, H.Y.; Writing—review & editing, L.D.; Funding acquisition, L.D. and Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Youth Science Fund Project of the Hunan Provincial Department of Science and Technology [2021JJ41080] and the Scientific Research Foundation of the Hunan Provincial Education Department [20B595].
Data Availability Statement
No applicable.
Acknowledgments
We thank the College of Environmental Science and Engineering, Central South University of Forestry and Technology, for providing the experimental platform, as well as the anonymous reviewers for their constructive comments and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
Correction Statement
This article has been republished with a minor correction to the existing affiliation information. This change does not affect the scientific content of the article.
References
- Usman, K.; Abu-Dieyeh, M.H.; Zouari, N.; Al-Ghouti, M.A. Lead (Pb) bioaccumulation and antioxidative responses in Tetraena qataranse. Sci. Rep. 2020, 10, 17070. [Google Scholar] [CrossRef] [PubMed]
- Moshchenko, D.; Kolesnikov, S.I.; Kuzina, A.A.; Kazeev, K.S.; Minkina, T.M.; Mezhenkov, A.A.; Litvinov, Y.A.; Shende, S.S.; Mandzhieva, S.S.; Sushkova, S.N. Comparative Assessment of the Resistance to Lead (Pb) Pollution of Forest, Forest-Steppe, Steppe, and Mountain-Meadow Soils of the Central Ciscaucasia and the Caucasus Regions. Forests 2022, 13, 1528. [Google Scholar] [CrossRef]
- Bouchard, M.F.; Bellinger, D.C.; Weuve, J.; Matthews-Bellinger, J.; Gilman, S.E.; Wright, R.O.; Schwartz, J.; Weisskopf, M.G. Blood lead levels and major depressive disorder, panic disorder, and generalized anxiety disorder in US young adults. Arch. Gen. Psychiatry 2009, 66, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Lanphear, B.P.; Hornung, R.; Khoury, J.; Yolton, K.; Baghurst, P.; Bellinger, D.C.; Canfield, R.L.; Dietrich, K.N.; Bornschein, R.; Greene, T.; et al. Low-level environmental lead exposure and children’s intellectual function: An international pooled analysis. Environ. Health Perspect. 2005, 113, 894–899. [Google Scholar] [CrossRef]
- Pourrut, B.; Jean, S.; Silvestre, J.; Pinelli, E. Lead-induced DNA damage in Vicia faba root cells: Potential involvement of oxidative stress. Mutat. Res. -Genet. Toxicol. Environ. Mutagen. 2011, 726, 123–128. [Google Scholar] [CrossRef]
- Tauqeer, H.M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Iftikhar, U.; Ahmad, R.; Farid, M.; Abbasi, G.H. Phytoremediation of heavy metals by Alternanthera bettzickiana: Growth and physiological response. Ecotoxicol. Environ. Saf. 2016, 126, 138–146. [Google Scholar] [CrossRef]
- Shahid, M.; Ferrand, E.; Schreck, E.; Dumat, C. Behavior and Impact of Zirconium in the Soil-Plant System: Plant Uptake and Phytotoxicity. Rev. Environ. Contam. Toxicol. 2013, 221, 107–127. [Google Scholar]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Menon, M.; Hermle, S.; Abbaspour, K.C.; Günthardt-Goerg, M.S.; Oswald, S.E.; Schulin, R. Water regime of metal-contaminated soil under juvenile forest vegetation. Plant Soil 2005, 271, 227–241. [Google Scholar] [CrossRef]
- Wu, Y.; Qi, L.; Wang, B.Y.; Medley, P.; Drake, J.; Vernon, J.; Ibeanusi, V.; Chen, G. Assess long-term As, Pb and Cr contamination and uptake by Eriocaulon decangulare in the Apalachicola National Forest. Sci. Total Environ. 2022, 838, 156040. [Google Scholar] [CrossRef]
- Wang, D.; Guo, W.; Zhang, G.; Zhou, L.; Wang, M.; Lu, Y.; Cai, D.; Wu, Z. Remediation of Cr (VI)-contaminated acid soil using a nanocomposite. ACS Sustain. Chem. Eng. 2017, 5, 2246–2254. [Google Scholar] [CrossRef]
- Rigoletto, M.; Calza, P.; Gaggero, E.; Malandrino, M.; Fabbri, D. Bioremediation methods for the recovery of lead-contaminated soils: A review. Appl. Sci. 2020, 10, 3528. [Google Scholar] [CrossRef]
- Su, R.; Ou, Q.; Wang, H.; Luo, Y.; Dai, X.; Wang, Y.; Chen, Y.; Shi, L. Comparison of Phytoremediation Potential of Nerium indicum with Inorganic Modifier Calcium Carbonate and Organic Modifier Mushroom Residue to Lead–Zinc Tailings. Int. J. Environ. Res. Public Health 2022, 19, 10353. [Google Scholar] [CrossRef] [PubMed]
- Mohtadi, A.; Ghaderian, S.M.; Schat, H. A comparison of lead accumulation and tolerance among heavy metal hyperaccumulating and non-hyperaccumulating metallophytes. Plant Soil 2012, 352, 267–276. [Google Scholar] [CrossRef]
- Jaffré, T.; Pillon, Y.; Thomine, S.; Merlot, S. The metal hyperaccumulators from New Caledonia can broaden our understanding of nickel accumulation in plants. Front. Plant Sci. 2013, 4, 279. [Google Scholar] [CrossRef] [PubMed]
- Capuana, M. Heavy metals and woody plants-biotechnologies for phytoremediation. Iforest-Biogeosciences For. 2011, 4, 7. [Google Scholar] [CrossRef]
- Gallagher, F.J.; Pechmann, I.; Bogden, J.D.; Grabosky, J.; Weis, P. Soil metal concentrations and vegetative assemblage structure in an urban brownfield. Environ. Pollut. 2008, 153, 351–361. [Google Scholar] [CrossRef]
- Kong, S.X.; Li, H.K.; Chao, J.; Cui, X.S.; Guo, Y.H. Effects of Pb Stress on Photosynthetic Pigment Biosynthesis and Growth of Rabdosia rubescens. Zhong Yao Cai 2015, 38, 215–220. [Google Scholar]
- Giannakoula, A.; Therios, I.; Chatzissavvidis, C. Effect of lead and copper on photosynthetic apparatus in citrus (Citrus aurantium L.) plants. The role of antioxidants in oxidative damage as a response to heavy metal stress. Plants 2021, 10, 155. [Google Scholar] [CrossRef]
- Peng, X.; Yang, B.; Deng, D.; Dong, J.; Chen, Z. Lead tolerance and accumulation in three cultivars of Eucalyptus urophyllaXE. grandis: Implication for phytoremediation. Environ. Earth Sci. 2012, 67, 1515–1520. [Google Scholar] [CrossRef]
- Kanwal, U.; Ibrahim, M.; Abbas, F.; Yamin, M.; Jabeen, F.; Shahzadi, A.; Farooque, A.A.; Imtiaz, M.; Ditta, A.; Ali, S. Phytoextraction of Lead Using a Hedge Plant [Alternanthera bettzickiana (Regel) G. Nicholson]: Physiological and Biochemical Alterations through Bioresource Management. Sustainability 2021, 13, 5074. [Google Scholar] [CrossRef]
- Silbergeld, E.K. Implications of new data on lead toxicity for managing and preventing exposure. Environ. Health Perspect. 1990, 89, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.N.; Chen, Y.H.; Du, L.; Zhang, M.Y.; Han, L.Z. Accumulation and subcellular distribution of heavy metal in Paulownia fortunei cultivated in lead-zinc slag amended with peat. Int. J. Phytoremediation 2019, 21, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Chen, G.; Xiong, L.; Liu, Q.; Yang, W. Accurate digitization of the chlorophyll distribution of individual rice leaves using hyperspectral imaging and an integrated image analysis pipeline. Front. Plant Sci. 2017, 8, 1238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Chen, Y.; Du, L.; Wu, Y.; Liu, Z.; Han, L. The potential of Paulownia fortunei seedlings for the phytoremediation of manganese slag amended with spent mushroom compost. Ecotoxicol. Environ. Saf. 2020, 196, 110538. [Google Scholar] [CrossRef]
- Deng, H.; Xu, D.; Li, M.; Li, J. Comparison of different digestion methods in analyzing heavy metals content in soils. J. Guangxi Norm. Univ. Nat. Sci. Ed. 2010, 28, 80–83. [Google Scholar]
- Cai, B.; Chen, Y.; Du, L.; Liu, Z.; He, L. Spent mushroom compost and calcium carbonate modification enhances phytoremediation potential of Macleaya cordata to lead-zinc mine tailings. J. Environ. Manag. 2021, 294, 113029. [Google Scholar] [CrossRef]
- Han, L.; Chen, Y.; Chen, M.; Wu, Y.; Su, R.; Du, L.; Liu, Z. Mushroom residue modification enhances phytoremediation potential of Paulownia fortunei to lead-zinc slag. Chemosphere 2020, 253, 126774. [Google Scholar] [CrossRef]
- Feng, P.; Sun, L.; Shen, X.; Li, R.; Jiang, C.; Zheng, H.; Li, Z.; Li, Z.; Guo, W.; Han, X. Response and accumulation ability of perennial ryegrass to plumbum and cadmium stress. Fresenius Environ. Bull. 2017, 26, 598–606. [Google Scholar]
- Ma, L.; Sun, H.; Chen, L.; Zhao, J. Relation between heavy metal fraction in soils and plants enrichment in pilot scale experiment on land application of sewage sludge. J. Food Agric. Environ. 2011, 9, 967–973. [Google Scholar]
- Basile, A.; Sorbo, S.; Conte, B.; Cardi, M.; Esposito, S. Ultrastructural changes and Heat Shock Proteins 70 induced by atmospheric pollution are similar to the effects observed under in vitro heavy metals stress in Conocephalum conicum (Marchantiales-Bryophyta). Environ. Pollut. 2013, 182, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.-X.; Xiao, H.; Guo, Q.J.; Zhang, Z.Y.; Yang, X.; Kong, J. Subcellular Distribution and Chemical Forms of Heavy Metals in Three Types of Compositae Plants from Lead-Zinc Tailings Area. Huan Jing Ke Xue Huanjing Kexue 2017, 38, 3054–3060. [Google Scholar] [PubMed]
- Dai, F.W.; Luo, G.; Li, Z.; Wei, X.; Wang, Z.; Lin, S.; Tang, C. Physiological and transcriptomic analyses of mulberry (Morus atropurpurea) response to cadmium stress. Ecotoxicol. Environ. Saf. 2020, 205, 10. [Google Scholar] [CrossRef] [PubMed]
- Parrotta, L.; Guerriero, G.; Sergeant, K.; Cai, G.; Hausman, J.F. Target or barrier? The cell wall of early- and later-diverging plants vs cadmium toxicity: Differences in the response mechanisms. Front. Plant Sci. 2015, 6, 16. [Google Scholar] [CrossRef]
- Su, R.; Xie, T.; Yao, H.; Chen, Y.; Wang, H.; Dai, X.; Wang, Y.; Shi, L.; Luo, Y. Lead Responses and Tolerance Mechanisms of Koelreuteria paniculata: A Newly Potential Plant for Sustainable Phytoremediation of Pb-Contaminated Soil. Int. J. Environ. Res. Public Health 2022, 19, 14968. [Google Scholar] [CrossRef]
- Cheng, H.-K.; Zhang, B.; Jing, X.-X.; Yang, S.-Q.; Zhao, P.; Sun, X.-X.; Zhou, Z.-Y. Response of Maize to Lead Stress and Relevant Chemical Forms of Lead. Huan Jing Ke Xue Huanjing Kexue 2015, 36, 1468–1473. [Google Scholar]
- Staszak, A.M.; Małecka, A.; Ciereszko, I.; Ratajczak, E. Differences in stress defence mechanisms in germinating seeds of Pinus sylvestris exposed to various lead chemical forms. PLoS ONE 2020, 15, 15. [Google Scholar] [CrossRef]
- He, L.; Su, R.; Chen, Y.; Zeng, P.; Du, L.; Cai, B.; Zhang, A.; Zhu, H. Integration of manganese accumulation, subcellular distribution, chemical forms, and physiological responses to understand manganese tolerance in Macleaya cordata. Environ. Sci. Pollut. Res. 2022, 29, 39017–39026. [Google Scholar] [CrossRef]
- Zhu, X.; Cheng, H.; Ji, L.; Ru, G.; Zhao, Z.; Cai, Y.; Wen, D. Physiological and enrichment characteristics of Paulownia fortunei seedlings under zinc, cadmium and their combined stress. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2021, 37, 2463–2473. [Google Scholar]
- Shakya, K.; Chettri, M.K.; Sawidis, T. Impact of heavy metals (copper, zinc, and lead) on the chlorophyll content of some mosses. Arch. Environ. Contam. Toxicol. 2008, 54, 412–421. [Google Scholar] [CrossRef]
- Sharma, I.; Sharma, A.; Pati, P.; Bhardwaj, R. Brassinosteroids Reciprocates Heavy Metals Induced Oxidative Stress in Radish by Regulating the Expression of Key Antioxidant Enzyme Genes. Braz. Arch. Biol. Technol. 2018, 61, 9. [Google Scholar] [CrossRef]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzałka, K.; Prasad, M. Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, C.; Huang, M.; Luo, J.; Hou, X.; Wu, P.; Ma, X. Lead tolerance mechanism in Conyza canadensis: Subcellular distribution, ultrastructure, antioxidative defense system, and phytochelatins. J. Plant Res. 2016, 129, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Vlot, A.C.; Sales, J.H.; Lenk, M.; Bauer, K.; Brambilla, A.; Sommer, A.; Chen, Y.; Wenig, M.; Nayem, S. Systemic propagation of immunity in plants. New Phytol. 2021, 229, 1234–1250. [Google Scholar] [CrossRef]
Figure 1.
Proportion of chemical forms of Pb in P. fortunei. Note: FE represents Pb inorganic; FW represents Pb organic; FNaCl represents pectin- and protein-bound Pb; FHAc represents Pb phosphate; FHCl represents Pb oxalate; FR represents Pb residual state.
Figure 1.
Proportion of chemical forms of Pb in P. fortunei. Note: FE represents Pb inorganic; FW represents Pb organic; FNaCl represents pectin- and protein-bound Pb; FHAc represents Pb phosphate; FHCl represents Pb oxalate; FR represents Pb residual state.

Figure 2.
Effects of Pb on antioxidant defense system in P. fortunei leaves. Note: (a) indicates superoxide dismutase (SOD) activity, (b) indicates catalase (CAT) activity, (c) indicates peroxidase (POD) activity, (d) indicates malondialdehyde (MDA) in P. fortunei leaves treated with different Pb concentrations. Different lowercase letters indicate significant differences between treatments (p < 0.05).
Figure 2.
Effects of Pb on antioxidant defense system in P. fortunei leaves. Note: (a) indicates superoxide dismutase (SOD) activity, (b) indicates catalase (CAT) activity, (c) indicates peroxidase (POD) activity, (d) indicates malondialdehyde (MDA) in P. fortunei leaves treated with different Pb concentrations. Different lowercase letters indicate significant differences between treatments (p < 0.05).
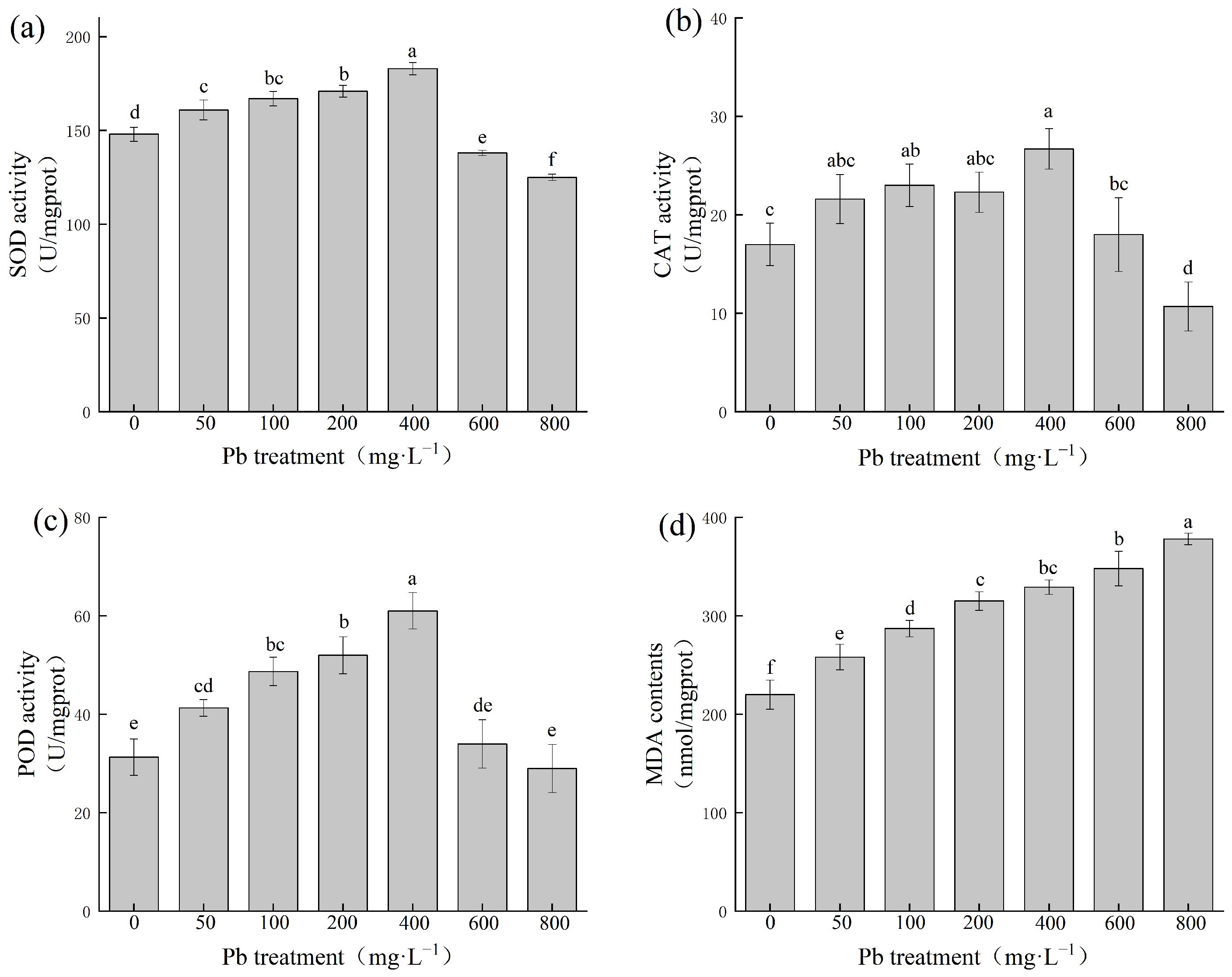
Figure 3.
Effect of Pb on the microstructure of P. fortunei. Note: CW = cell wall, SG = starch granule, V = vacuole, and Chl = chloroplast. Red arrows point out heavy metal particles. (A,D,H) are the root, stem, and leaves, respectively, of P. fortunei in the control group; (B,E,I) are the root, stem, and leaves, respectively, of P. fortunei under 400 mg L−1 of Pb; (C,F,G) are the root, stem, and leaves, respectively, of P. fortunei under 800 mg L−1 of Pb.
Figure 3.
Effect of Pb on the microstructure of P. fortunei. Note: CW = cell wall, SG = starch granule, V = vacuole, and Chl = chloroplast. Red arrows point out heavy metal particles. (A,D,H) are the root, stem, and leaves, respectively, of P. fortunei in the control group; (B,E,I) are the root, stem, and leaves, respectively, of P. fortunei under 400 mg L−1 of Pb; (C,F,G) are the root, stem, and leaves, respectively, of P. fortunei under 800 mg L−1 of Pb.

Figure 4.
Effect of Pb on functional group of P. fortunei. Note: (a–c) correspond to the root, stem, and leaves of P. fortunei, respectively. (1–4) indicate the absorption peaks near 3420 cm−1, 2920 cm−1, 1630 cm−1, and 1040 cm−1, respectively.
Figure 4.
Effect of Pb on functional group of P. fortunei. Note: (a–c) correspond to the root, stem, and leaves of P. fortunei, respectively. (1–4) indicate the absorption peaks near 3420 cm−1, 2920 cm−1, 1630 cm−1, and 1040 cm−1, respectively.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of Pb stress on the biomass parameters of P. fortunei.
| Pb Concentration (mg·L−1) | Plant Height (cm) | Total Biomass (g) | Root Length (cm) | Root Surface Area (cm)2 | Average Root Diameter (mm) | Root Tip Number (a) |
|---|---|---|---|---|---|---|
| 0 | 30.1 ± 2.9 d | 7.3 ± 1.8 d | 59.3 ± 1.5 c | 17.7 ± 1.7 d | 0.6 ± 0.4 e | 433 ± 25 d |
| 50 | 34.0 ± 1.3 c | 12.2 ± 1.1 c | 60.0 ± 4.3 c | 22.1 ± 0.9 c | 1.3 ± 0.5 d | 569 ± 44 c |
| 100 | 39.2 ± 1.1 b | 15.9 ± 1.9 c | 66.1 ± 3.8 b | 24.8 ± 1.6 c | 2.5 ± 0.2 c | 606 ± 25 c |
| 200 | 39.4 ± 1.7 b | 22.4 ± 2.7 b | 73.8 ± 1.0 a | 30.6 ± 2.2 b | 3.8 ± 0.2 b | 737 ± 28 b |
| 400 | 45.2 ± 1.4 a | 32.6 ± 2.4 a | 78.5 ± 1.7 a | 36.4 ± 1.9 a | 4.4 ± 0.3 a | 818 ± 18 a |
| 600 | 25.1 ± 1.0 e | 5.5 ± 0.4 de | 34.2 ± 1.8 d | 8.1 ± 1.3 e | 2.4 ± 0.2 c | 351 ± 36 e |
| 800 | 16.0 ± 1.3 f | 3.3 ± 0.5 e | 18.7 ± 1.7 e | 4.8 ± 0.5 f | 1.4 ± 0.2 d | 269 ± 24 f |
Note: Data are mean ± standard deviation. Different letters in the same column indicate significant difference in P. fortunei growth index under different treatments (p < 0.05).
Table 2.
Heavy metal content of P. fortunei under Pb stress.
| Pb Treatment mg·L−1 | Pb Content (mg·kg−1) | BCF | TF | ||
|---|---|---|---|---|---|
| Roots | Stems | Leaves | |||
| 50 | 229.8 ± 15.18 f | 7.5 ± 0.85 d | 14.1 ± 1.08 f | 0.18 ± 0.01 a | 0.09 ± 0.01 e |
| 100 | 460.5 ± 9.55 e | 25.7 ± 3.98 cd | 29.8 ± 2.34 e | 0.19 ± 0.01 a | 0.12 ± 0.01 d |
| 200 | 821.0 ± 25.69 d | 55.2 ± 3.38 c | 63.7 ± 3.78 d | 0.15 ± 0.01 b | 0.15 ± 0.01 d |
| 400 | 1108 ± 41.56 c | 130.1 ± 15.33 b | 175.5 ± 10.58 c | 0.12 ± 0.01 c | 0.28 ± 0.02 c |
| 600 | 1349.4 ± 17.46 b | 177.0 ± 7.46 a | 386.1 ± 12.60 a | 0.09 ± 0.003 d | 0.42 ± 0.01 a |
| 800 | 1587.0 ± 23.41 a | 186.9 ± 7.58 a | 339.2 ± 38.00 b | 0.06 ± 0.006 e | 0.33 ± 0.02 b |
Note: Pb content is related to the total biomass of Pb in each organ of P. fortunei. Data are mean± standard deviation. Different letters in the same column indicate significant difference in heavy metal content of P. fortunei under different treatments (p < 0.05).
Table 3.
Subcellular distribution of Pb in P. fortunei.
| Pb Treatment mg·L−1 | Pb Content (mg·kg−1) Percentage (%) | |||||
|---|---|---|---|---|---|---|
| Root Stem Leaves | Root Stem Leaves | |||||
| F1 | 159.9 ± 3.4 a | 4.99 ± 0.2 a | 8.3 ± 0.2 a | 75.7 | 71.1 | 60.1 |
| 50 F2 | 22.1 ± 0.3 b | 1.0 ± 0.02 b | 2.4 ± 0.1 c | 10.5 | 9.5 | 17.4 |
| F3 | 26.5 ± 2.2 b | 1.4 ± 0.04 b | 3.1 ± 0.1 b | 13.8 | 19.4 | 22.5 |
| F1 | 300.7 ± 7.8 a | 13.9 ± 0.8 a | 13.4 ± 1.8 a | 71.8 | 63.5 | 49.6 |
| 100 F2 | 43.0 ± 2.5 c | 3.2 ± 0.2 c | 6.6 ± 0.1 c | 10.8 | 10.5 | 20.8 |
| F3 | 72.7 ± 2.4 b | 5.7 ± 0.3 b | 8.0 ± 0.3 b | 17.4 | 26.0 | 29.6 |
| F1 | 549.2 ± 5.9 a | 29.4 ± 1.7 a | 30.4 ± 1.4 a | 68.3 | 60.4 | 47.6 |
| 200 F2 | 96.3 ± 2.2 c | 5.8 ± 0.1 c | 14.2 ± 1.3 c | 12.0 | 11.9 | 21.1 |
| F3 | 157.8 ± 2.1 b | 13.4 ± 2.0 b | 20.0 ± 3.7 b | 19.7 | 27.7 | 31.3 |
| F1 | 689.4 ± 6.0 a | 68.0 ± 3.3 a | 74.9 ± 1.8 a | 65.2 | 56.9 | 45.5 |
| 400 F2 | 112.7 ± 5.7 c | 13.5 ± 3.6 c | 34.5 ± 1.8 c | 11.3 | 11.3 | 21.0 |
| F3 | 248.6 ± 2.9 b | 36.0 ± 2.9 b | 55.6 ± 3.2 b | 23.5 | 31.8 | 33.5 |
| F1 | 830.8 ± 5.2 a | 86.0 ± 5.4 a | 155.6 ± 4.7 a | 61.7 | 49.3 | 41.3 |
| 600 F2 | 192.7 ± 4.1 c | 32.5 ± 2.2 c | 95.9 ± 2.9 c | 14.3 | 18.6 | 25.4 |
| F3 | 325.9 ± 15.8 b | 59.4 ± 1.0 b | 126.1 ± 5.4 b | 24.0 | 32.1 | 33.3 |
| F1 | 837.0 ± 8.9 a | 84.4 ± 9.0 a | 123.6 ± 7.8 a | 54.8 | 46.0 | 38.0 |
| 800 F2 | 261.6 ± 3.4 c | 40.5 ± 2.6 c | 87.4 ± 2.8 b | 17.1 | 20.8 | 26.8 |
| F3 | 430.0 ± 3.4 b | 61.0 ± 0.8 b | 114.2 ± 3.3 a | 28.1 | 33.2 | 35.2 |
Note: Pb content is related to the total biomass of Pb in each organ. Different letters in the same column indicate significant difference in heavy metal content of P. fortunei under different treatments (p < 0.05). F1 represents the cell wall component; F2 represents organelle and nuclear components; F3 represents the soluble component.
Table 4.
Effect of Pb treatment on pigment content in P. fortunei leaves.
| Pb Concentration | Chlorophyll a (mg·g−1) | Chlorophyll b (mg·g−1) | Carotenoids (mg·g−1) | Total Chlorophyll (mg·g−1) | Chlorophyll a/b |
|---|---|---|---|---|---|
| 0 | 9.3 ± 1.63 c | 5.4 ± 0.28 c | 2.0 ± 0.27 b | 1.9 ± 0.35 c | 1.8 ± 0.74 c |
| 50 | 9.3 ± 1.03 c | 5.5 ± 0.40 bc | 2.0 ± 0.11 b | 2.0 ± 0.04 c | 1.7 ± 0.10 c |
| 100 | 11.8 ± 0.80 b | 6.0 ± 0.21 a | 2.1 ± 0.09 b | 2.3 ± 0.05 b | 2.0 ± 0.07 b |
| 200 | 12.7 ± 1.59 b | 6.1 ± 0.20 a | 2.3 ± 0.08 ab | 2.5 ± 0.06 b | 2.1 ± 0.21 b |
| 400 | 14.9 ± 0.73 a | 5.9 ± 0.19 ab | 2.5 ± 0.19 a | 2.8 ± 0.11 a | 2.5 ± 0.08 a |
| 600 | 7.4 ± 0.44 c | 5.3 ± 0.14 c | 1.7 ± 0.16 c | 1.6 ± 0.04 d | 1.4 ± 0.05 d |
| 800 | 5.0 ± 0.17 d | 4.4 ± 0.04 d | 1.2 ± 0.12 d | 1.2 ± 0.11 e | 1.1 ± 0.03 e |
Note: Different letters in the same column indicate significant difference in chlorophyll content under different treatments (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Du, L.; Yang, H.; Xie, J.; Han, L.; Liu, Z.; Liu, Z.; Chen, Y.; Su, R. The Potential of Paulownia fortunei L. for the Phytoremediation of Pb. Forests 2023, 14, 1245. https://doi.org/10.3390/f14061245
AMA Style
Du L, Yang H, Xie J, Han L, Liu Z, Liu Z, Chen Y, Su R. The Potential of Paulownia fortunei L. for the Phytoremediation of Pb. Forests. 2023; 14(6):1245. https://doi.org/10.3390/f14061245
Chicago/Turabian StyleDu, Lu, Hang Yang, Juan Xie, Liangze Han, Zhiyi Liu, Zhiming Liu, Yonghua Chen, and Rongkui Su. 2023. "The Potential of Paulownia fortunei L. for the Phytoremediation of Pb" Forests 14, no. 6: 1245. https://doi.org/10.3390/f14061245
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.