

Nitrogen Addition Alleviates Cadmium Toxicity in Eleocarpus glabripetalus Seedlings

1
College of Forestry and Biotechnology, Zhejiang A&F University, Hangzhou 311300, China
2
Zhejiang Public Welfare Forest and State Forest Farm Management Station, Hangzhou 310012, China
3
East China Inventory and Planning Institute of National Forestry and Grassland Administration, Hangzhou 310019, China
*
Authors to whom correspondence should be addressed.
Forests 2023, 14(6), 1264; https://doi.org/10.3390/f14061264
Submission received: 15 May 2023
/
Revised: 13 June 2023
/
Accepted: 14 June 2023
/
Published: 19 June 2023
(This article belongs to the Special Issue Effects of Abiotic Stress on Tree Physiology and Ecology)
Abstract
:Cadmium (Cd) accumulation in soil is a serious form of heavy metal pollution affecting environmental safety and human health. In order to clarify the tolerance mechanisms to Cd-contaminated soils under N deposition, changes in plant growth, root architecture and physiological characteristics of Eleocarpus glabripetalus seedlings under combined nitrogen (N) and cadmium (Cd) treatments were determined in this study. The results indicated that Cd-induced negative effects inhibited the growth of E. glabripetalus seedlings through increased underground biomass allocation, and affected transpiration and respiratory processes, resulting in a decreased soluble sugars concentration in leaves and non-structural carbohydrates (NSC) in the roots. Root systems might play a major role in Cd absorption. Cd stress restricted the growth of fine roots (<0.5 mm), and affected the uptake of N and P. N addition alleviated the Cd-induced negative effect on plant growth through improving the root system, increasing starch and NSC contents in the roots and increasing total biomass. These findings have important implications for understanding the underlying tolerance mechanisms of Cd pollution under N deposition in arbor species.
1. Introduction
The rapid development of industrialization and urbanization has led to increasingly serious soil heavy metal contamination. The accumulation of heavy metals could affect plant growth and even cause ecological imbalance and environmental deterioration [1]. Cadmium (Cd) is a highly toxic heavy metal pollutant in agricultural soil [2] that could be accumulated in plant organs after being absorbed by plant roots, causing serious damage to plant growth [3,4]. Cd ions could affect the uptake, transportation and subsequent distribution of nutrient elements in plants [5]. When Cd2+ is excessively accumulated by plants, it can affect the physiology and biochemical activities of plants, such as photosynthesis, respiration, transpiration, and even the expression of related genes [6,7,8].
Global atmospheric nitrogen (N) deposition is steadily increasing with the increased burning of fossil fuels and the use of man-made fertilizers, and it has considerable effects on terrestrial ecosystems [9]. Moderate N deposition can contribute to an increase in plant biomass [10,11]. However, excessive N input can inhibit plant photosynthesis [12], and cause changes in soil physicochemical properties, resulting in decreased root growth and affecting plant root morphology [13]. As is widely known, nitrogen is essential for plant growth and development [14,15]. Nitrogen also plays a crucial role in plant stress resistance [15,16], and its supply may influence the plant’s ability to cope with abiotic stress, e.g., heavy metal pollution [17,18]. Adequate N application could enhance plant adaptability and resilience under various environmental conditions through promoting the synthesis of amino acids, enzymes and hormones [16,19,20].
Some research has revealed that N can effectively decrease the Cd toxicity for plants [21,22,23]. This alleviation effect of N addition on Cd toxicity differs among plant species and according to concentrations of Cd and N. It has been reported that appropriate N addition could promote root development and enhance Cd accumulation in plants [24]. High-concentration N supplementation could enhance plants’ growth, as well as the antioxidant enzymes’ activities, which may partially alleviate AOS accumulation induced by Cd stress [25,26]. However, the exact mechanism of how additional N alleviates cadmium-induced stress responses remains unknown. Additionally, many of the previous studies on Cd-stress in plants focused on vegetables [1,27,28,29] and crops [30,31], and only a few on woody plants, which are more susceptible to soil contamination because of their slow growth and the long generation time.
Eleocarpus glabripetalus is a dominant evergreen tree species in the subtropical forest of China. According to our previous studies, E. glabripetalus seedlings showed a strong plasticity to Cd stress and acid rain [32]. However, this could not fully explain the adaption of E. glabripetalus to soil Cd pollution. Therefore, in this study we aimed to understand the effects of Cd pollution on the morphology and physiology of E. glabripetalus under N deposition. We hypothesized that (1) N addition could influence eco-physiological traits under Cd stress, and (2) N addition could alleviate cadmium-induced stress responses.
2. Materials and Methods
2.1. Experiment Design
The experiment was carried out in a greenhouse of the Botanical Garden of Zhejiang A&F University (119°44′ E, 30°16′ N), East China. One-year-old healthy E. glabripetalus seedlings with a height of 60 cm and a ground diameter of about 0.9–1.0 cm were planted in plastic pots with 30 cm inner diameter and 40 cm depth. There was one seedling in each pot, with a PVC plate under the pot. E. glabripetalus seedlings were treated with Cd treatments (no Cd and 100 mg·kg−1 Cd), N additions (no N addition and 90 kg N·ha−1·yr−1 N addition), and a combined treatment of Cd and N. In each treatment, 12 seedlings were used as replicates to minimize sampling errors.
According to the reported average soil Cd concentration [33,34], Cd concentrations of 100 mg·kg−1 were selected. The Cd2+ solutions were prepared using CdCl2·5H2O (Chemical Co. Ltd., Shanghai, China) [32] and were sprayed evenly on the air-dried, reddish-brown forest soil and mixed well until the soil Cd concentration reached 100 mg kg−1. Then, the soil was potted, with 10 kg per pot. The same volume of soil with no Cd was also potted as the control. Then, E. glabripetalus seedlings were planted in pots in January 2019. After two months of growth, simulated nitrogen deposition treatment was carried out. According to the atmospheric N deposition of 59.1–70.5 kg N ha−1·yr−1 in eastern China [35], the average total N deposition of 80 kg N·hr−1·yr−1 [36], and the projected local N deposition value by 2050 [37], we applied 90 kg N·ha−1·yr−1 as N addition treatments in this study. Average weekly N deposition rate and N input amounts for each pot were calculated. A dissolved ammonium nitrate (NH4NO3) solution was sprayed evenly onto leaves and soil twice a week during the experimental period. The N treatments lasted 180 days, from April to October.
2.2. Measurements of Gas Exchange Parameters
Three individuals from each treatment were chosen randomly to measure leaf gas exchange from 8:00 a.m. to 11:30 a.m. after 180 days of treatments. The 3rd to 5th leaves with the same orientation were selected for gas exchange parameter measurements using LI-6400 system (Li-Cor Inc., Lincoln, NE, USA). The conditions were 25 °C leaf temperature, 50% relative air humidity, 400 ± 5 μmol·mol−1 CO2 concentration and 1000 μmol m−2·s−1 light intensity. The net photosynthetic rate (Pn), transpiration rate (Tr), stomatal conductance (gs) and intercellular CO2 concentration (Ci) of plants were recorded. Then, the intrinsic water use efficiency (WUE) was calculated with the following formula:
2.3. Measurements of Root Morphology and Biomass
At the end of treatments, four individuals of each treatment were harvested and separated into roots, leaves and stems after washing with deionized water. The root system was imaged using a scanner (LA2400 Scanner, Seiko Epson Corp., Nagano, Japan). A WinRHIZO System (Version 2012b, Regent Instruments Inc., Québec, QC, Canada) was used to analyze the total root length, average diameter, root diameter classes, root surface area, and root volume. Then, the samples were oven-dried to reach a constant dry weight. Total biomass (TB) was the sum of root biomass (RB), stem biomass (SB) and leaf biomass (LB). Biomass distributions were calculated with the following formula:
Then, dried leaves, stems and roots were ground separately and sieved through a 100-mesh sieve for determining the contents of N, phosphorus (P), Cd, starch and soluble sugar contents.
2.4. Measurements of N, P and Cd Contents, and Non-Structural Carbohydrate (NSC) Contents
The semi-micro-Kjeldahl acid-digestion method was used to determine the N content in leaves, stems and roots using an Alpkem Auto Analyzer (Kjektec System 2300 Distilling Unit, Tecator AB, Hoganas, Sweden). H2SO4-H2O2 decoction and molybdenum-antimony colorimetry was used to determined P content. ICP-MS was used to determine the Cd contents.
The total NSCs contents were considered as the sum of soluble sugars and starch. Soluble sugars were extracted from dried leaves, stems, and roots in 80% (v/v) ethanol. Then, the extraction was centrifuged at 5000× g for 20 min. Then, 80% (v/v) ethanol was added to the pellet, and centrifugated for 5 min at 5000× g. The soluble sugars content in the supernatant was measured colorimetrically at 620 nm using the phenol-sulfuric method. Then, starch content was determined from the remaining pellet after the soluble sugars were extracted. After incubation in sodium acetate and amyloglucosidase solution, starch content was measured colorimetrically at 650 nm using the phenol-sulfuric method, as described by Newell et al. [38].
2.5. Data Analysis
Statistical analyses were performed using SPSS v22.0 software (SPSS, Inc., Chicago, IL, USA). The normality of the data were tested using Levene’s test before analysis, and, if necessary, natural log transformations were performed. One-way analysis of variance (ANOVA) was used to analyze the significant difference between the N treatments or Cd treatments. Two-way ANOVA was used to analyze the interactions of Cd and N treatments. Statistically significant difference was set at a 95% confidence level. All data were expressed as means ± standard error (SE, n ≥ 3). Correlation analyses between two variables were tested with Spearman’s rho correlation coefficients using Origin 2020 (Origin 2020, Origin Lab, Northampton, MA, USA). Graphs were prepared in Origin 2020.
3. Results
3.1. Gas Exchange
The Pn, gs and Tr of E.glabripetalus seedlings decreased significantly under Cd treatment and N treatment compared with the control group (p < 0.05) (Figure 1). However, the combined N + Cd treatment increased the Pn, gs and WUE of E. glabripetalus seedlings significantly compared with the Cd-treated group (p < 0.05). A significant interactive effect between Cd treatment and N treatment on Pn, gs and Tr was detected (p < 0.05).
3.2. Biomass
The total biomass and leaf and stem biomass in the Cd-treated group were significantly decreased compared with control plants (p < 0.05) (Figure 2). N addition significantly increased the leaf biomass, while the combined N and Cd treatment significantly increased the leaf biomass, root biomass and total biomass, compared to the plants under single cadmium treatment (p < 0.05). Compared with control plants, RSR was significantly increased under the Cd treatment and combined N + Cd treatment. The Cd and N treatment had a significant interactive effect on root biomass (p < 0.05).
3.3. Root Architecture
Cd treatment, N addition and their combination significantly altered the root phenotypic traits (Table 1). Cd treatment significantly increased the root average diameter, and the proportion of the length of fine roots with 0.5–1 mm- and 1–2 mm-diameter classes to total root length, but decreased the proportion of the length of <0.5 mm-diameter fine roots to the total root length. N treatment increased the total root length and total root surface area significantly compared with control groups (p < 0.05). Compared to the Cd- treated seedlings, the combined N + Cd treatment increased the total root length, total root surface area, total root volume and the proportion of fine roots (<0.5 mm in diameter) significantly (p < 0.05), but decreased the root mean diameter and length of roots with 1–2 mm diameter.
3.4. N, P and Cd Contents
N treatment and the combined N + Cd treatment significantly increased the leaf N content and N/P ratio (Figure 3). The phosphorus content in stems and leaves of E. glabripetalus seedlings grown under Cd treatment decreased compared with the control group, while the nitrogen–phosphorus ratio of stems and leaves was significantly increased. The highest Cd content was found in the roots of the Cd treatment group (Table 2). Compared to Cd treatment, Cd content in the above-ground parts was significantly increased under the combined N + Cd treatment, indicating combined N + Cd treatment had a significant impact on Cd accumulation in above-ground parts (p < 0.05).
3.5. Non-Structural Carbohydrate Content
Cd and N treatment decreased the soluble sugar contents in the roots and leaves of E. glabripetalus (p < 0.05) (Figure 4). However, soluble sugars in the stems remained relatively stable compared with control plants. Cd treatment decreased the stem starch content but increased the leaf starch content. Combined N and Cd treatments increased the root starch content but decreased the leaf starch content compared to Cd treatment. The NSCs contents in root and stem decreased significantly under Cd treatment (p < 0.05). N treatment increased NSCs contents in the root under Cd treatment. The soluble sugar/starch ratio in leaves was significantly decreased, by 30%, under Cd treatment compared with the control group. Cd and N treatment had a significant interactive effect on soluble sugars and the non-structural carbon of leaves, as well as the non-structural carbon of roots (p < 0.05).
3.6. Correlation between the Indicators in E. glabripetalus Seedlings
The leaf biomass and stem biomass were negatively correlated with leaf non-structural carbohydrate, leaf starch and root mean diameter, and positively correlated with N and P contents in plants. Root biomass was negatively correlated with root soluble sugar/starch ratio. Root length and root surface area were negatively correlated with leaf non-structural carbohydrate and root soluble sugar/starch ratio, and positively correlated with N contents in plants. Pn was positively correlated with non-structural carbohydrate in plants. Cd content in roots, stems and leaves was positively correlated with WUE, root biomass and root surface area, and negatively correlated with root soluble sugars and non-structural carbohydrates (Figure 5).
4. Discussion
4.1. The Impact of N and Cd Treatments on the Gas Exchange of E. glabripetalus Seedlings
It has been reported that Cd has adverse effects on the gas exchange system in plants [39,40,41]. Our results also found that E. glabripetalus seedlings responded to Cd with an inhibited gas exchange system, which makes them vulnerable to environmental stress. Previous studies have revealed that Cd could decrease gas exchange attributes [42,43]. Cd stress decreased the Pn, gs and Tr of E. glabripetalus in this study, indicating that Cd has deleterious effects on transpiration and respiratory processes and stomatal conductance, ultimately leading to a decline in photosynthesis [44,45]. Inhibited photosynthesis induced by Cd can be attributed mainly to stomatal and non-stomatal restriction [46]. The former refers to Cd causing stomatal closure, while the latter refers to the regulation of the plant’s intrinsic mechanisms. In addition, there was a significant linear correlation (p < 0.05) between Pn, gs and Tr in seedlings, suggesting that Cd treatments may limit Pn by affecting the stomatal factors of the plant. Since gs was not significantly correlated with Ci, the significant decrease in Pn may be due to a combination of stomatal and non-stomatal factors [46]. Ci was not significantly changed, indicating that leaf stomatal conductance was decreased in order to reduce leaf transpiration and prevent excessive tissue water deficit, keeping the rate of carbon dioxide, which indirectly led to the decrease in Pn. These results indicated that declined Pn, gs and Tr are closely related to plant growth [47,48].
N addition altered the negative effects induced by Cd stress, which was reflected by a significant increase in Pn, gs and WUE. This indicated that N addition could mitigate the damage of the gas exchange system induced by Cd. N fertilization could have a positive effect on Pn, gs and Tr, which are positively related to plant yield [48]. Nitrogen supplementation helps stomatal opening, slightly restores the level of photosynthesis, improves the water use efficiency, and reduces stomatal limitations induced by Cd stress. These results indicate that N addition could help plants to increase their sensitivity to other stresses [49,50,51].
4.2. The Impact of Cd and N Treatments on N, P and Cd Contents of E. glabripetalus Seedlings
In the present study, Cd accumulation in E. glabripetalus seedlings was mainly concentrated in the root system. Most studies have shown that plant root systems have a greater capacity to accumulate cadmium than the aboveground parts [52,53,54]. Normally, Cd absorbed by plants accumulates first in the root system and is subsequently distributed to individual tissues [53]. To avoid adverse effects of Cd on the normal physiological metabolism of the plant, Cd is usually stored in the underground parts, far from sensitive photosynthetic organs [52,55]. Most of the Cd accumulated in the roots can reduce the toxic effect of Cd on aboveground tissues and improve the Cd tolerance of seedlings to a certain extent, which is a self-protection mechanism [53,56]. These results showed that root systems have a major impact on heavy metal uptake. Root characteristics, such as root size, root surface area and the fineness of roots, may play the key role in Cd absorption.
In order to avoid unfavorable environmental stresses, plants can adopt some adaptation strategies to adapt and protect themselves from stress [57]. For example, plants could regulate nutrient allocation strategies to meet the nutrient requirements of different organs under Cd stress [58]. The essential elements absorbed by plant root systems could be transported to stems and leaves, in which equilibrium can be disturbed by the presence of heavy metals, thus affecting the elements’ uptake and distribution in plant. This can be explained by the competition for the same transporters, a disturbance in water uptake as a result of Cd stress or disturbed key enzymes processes [59]. All these processes have effects on plant growth, such as on photosynthesis, non-structural carbon distribution and biomass allocation. According to the correlation analysis, the Cd content in plants was negatively correlated with N and P content in plants. Thus, it is hypothesized that the allocation of N and P in various plant parts may be affected by the accumulation of Cd through influencing the changes and equilibrium regulation in biomass and nonstructural carbon.
4.3. The Impact of N and Cd Treatments on the Non-Structural Carbohydrates of E. glabripetalus Seedlings
As is well known, NSCs represent the photosynthesis products and have important functions in plant development, carbon–nitrogen metabolism and resistance [60]. As the energy supply substances in plant growth metabolism, NSCs comprise mainly soluble sugars involved in nonstructural carbohydrate transport in plants or temporary storage, and starch stored for a long time in different plant tissues for future use, maintaining function when carbon demand is higher than supply under abiotic stress [61,62,63]. In this study, no matter what kind of treatment, starch was the main fraction of NSCs (>69%) in the roots and was the main storage system of carbohydrates in the roots. However, soluble sugars are the main fraction of leaf NSCs (>63%) of E. glabripetalus seedlings. Most of the NSCs were stored in the leaves and the roots, suggesting their importance in carbohydrate production and NSCs’ storage in E. glabripetalus. Carbon storage in the root plays a critical role in supporting plant growth under abiotic stress. Cd treatment decreased root NSCs contents, which may be because the adaption strategies of E. glabripetalus respond to Cd stress by decreasing carbohydrate temporary storage in the roots to restrict the new fine root growth, which inhibits Cd uptake. Soluble sugars contribute to the osmotic adjustment and can indicate an ability to adapt to environmental changes [64]. The decreased soluble sugars concentration induced by Cd in roots and leaves reflected a decrease in carbohydrates produced by photosynthesis, shown by the decreased Pn, and also as a result of carbohydrate conversion. These results indicated that the NSCs’ allocation pattern changes in roots and leaves reflected the dynamic balance between assimilated savings and growth investment allocation, and plant adaptation strategies to environmental changes [65,66].
N treatment increased the root starch and NSC contents under Cd stress, with a decrease in the soluble sugars to starch ratio, indicating the increased starch content to maintain carbon demand under abiotic stress [67]. N alleviated the adverse effects on root induced by Cd, since a large portion of starch and NSC were supplied to support the production of new tissues (such as new fine roots). Furthermore, some of starch and NSC were also used for plant respiration. The decrease in leaf starch and NSC under the combined N and Cd treatment indicated that increased stomatal conductance and photosynthetic rate resulted in less carbon synthesis than consumption [68], showing that N addition alleviated leaf growth restriction by Cd. These results indicated that carbohydrates invested in growth or storage may be affected by environmental changes.
4.4. The Impact of Cd and N Treatments on the Growth of E. glabripetalus Seedlings
It has been reported that Cd could have negative effects on plant growth [27,69], often as the indicator for the toxicity assessment of heavy metals in plants [70,71]. Similar to the findings of previous studies [32], this study also found that E. glabripetalus seedlings responded to Cd with decreased above-ground biomass and total biomass. Interestingly, Cd significantly increased the root biomass allocation, showing that more resources were allocated to underground parts to resist Cd stress, which reflected the optimal allocation partitioning in plant biomass [72]. More biomass allocation to the root system may be associated with a reduction in resource uptake and photosynthesis. In general, heavy metals may affect the ability of resources (e.g., carbon, nutrients, or water) to be transported upward from the roots, thereby affecting plant growth [64,73]. The decreased leaf and stem biomass might result from the large root Cd accumulation, which restricted the nutrient transportation to stems and leaves. Single N addition has no effect on total biomass accumulations. However, N addition improved the plant growth under Cd stress by increasing the total biomass. Meanwhile, nitrogen has a positive influence on root growth and development, as N treatment significantly increased root biomass and carbohydrate storage in the root, with increased starch and NSC contents. These results revealed that N addition effectively alleviated the inhibition of Cd on plant growth [74,75,76].
Root morphology is an important indicator for environmental stress tolerance assessment [77,78]. Roots have phenotypic plasticity under stress conditions, which allows plants to obtain more resources from the surrounding soil environment [79]. In this study, the roots of E. glabripetalus seedlings became significantly thicker, and the fine roots (<0.5 mm) were significantly inhibited under Cd treatment, as reported in other studies, where high Cd could inhibit root hair development and fine root production [80,81]. Lateral and fine roots could increase the volume of soil reached by the root. However, Cd reduced the proportion of the length of <0.5 mm fine roots to the total root length, making the root system shorter and thicker and reducing root vigor, which in turn affected the nutrient uptake [80]. As is well known, root systems are extremely important in responding to heavy metals because root systems can regulate plants’ responses to stress through coordinated nutrients absorption and distribution in the whole plant [82]. There was a significant correlation between the root morphology and biomass of E. glabripetalus seedlings, and the root system of E. glabripetalus seedlings was inhibited by Cd treatment, which affected the nutrient uptake and thus affected plant growth.
N treatment increased the root growth of E. glabripetalus under Cd treatment, which was beneficial for increasing water and nutrient absorption. In fact, many studies have found that moderate N addition could positively alleviate Cd’s adverse effects on plants [50,76,83]. The reason might be that adequate N supply could alter the bio-available Cd concentration in the soil [84], which is favorable for plants to absorb more Cd from the soil. More importantly, N could regulate cell wall isolation, chelation capacity and oxidative resistance to regulate Cd accumulation in plants [50]. Whether for N addition or Cd stress, roots are pivotal for elemental iron absorption. Improved root systems indicate that N addition alleviated Cd-induced toxicity.
Plants under adverse environmental conditions may show effective ecological strategies [85,86]. The changes after N addition indicated that N improved plant stress resistance by increasing photosynthesis and improving root growth, and affected carbon conversion. These indicated plants needed response time to adverse environments for plant growth, and often adopted the physiology-to-phenotype strategy. However, other influencing factors, e.g., interactions of plant–microbes in the soil, may also participate in Cd stress responses in E. glabripetalus, and should be considered in the future studies.
5. Conclusions
Our results suggested that Cd stress inhibited the growth of E. glabripetalus. Cd treatment affected transpiration and respiratory processes and stomatal conductance, resulting in a decline in photosynthesis, which led to decreased soluble sugar concentrations in leaves and NSC in the roots. Cd affected biomass allocation by decreasing the above-ground biomass and increasing underground biomass allocation. The growth of fine roots (<0.5 mm) was restricted by Cd. Most Cd was retained in roots, which affected the distribution of N and P in the plant. N addition improved the root system by increasing root growth, and increasing starch and NSC contents in the roots under Cd stress. With increased total biomass, these results indicated that N addition alleviated Cd-induced growth inhibition. These findings have important implications for understanding the underlying tolerance mechanisms of Cd pollution under N deposition in arbor species.
Author Contributions
Conceptualization, L.Y.; data analysis, H.G.; investigation, M.S.; writing—original draft preparation, M.L.; writing—review and editing, Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Zhejiang Provincial Natural Science Foundation of China, grant number LY19C160005.
Data Availability Statement
The data that support the findings of this study are available on request from the first author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.W.; Zia-Ur-Rehman, M.; Zahir, Z.A.; Rinklebe, J.; Tack, F.M.G.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Kubier, A.; Wilkin, R.T.; Pichler, T. Cadmium in soils and groundwater: A review. Appl. Geochem. 2019, 108, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sakouhi, L.; Kharbech, O.; Massoud, M.B.; Munemasa, S.; Murata, Y.; Chaoui, A. Oxalic acid mitigates cadmium toxicity in Cicer arietinum L. germinating seeds by maintaining the cellular redox homeostasis. J. Plant Growth Regul. 2021, 41, 697–709. [Google Scholar] [CrossRef]
- Yin, A.; Huang, B.; Xie, J.; Huang, Y.; Shen, C.; Xin, J. Boron decreases cadmium influx into root cells of Capsicum annuum by altering cell wall components and plasmalemma permeability. Environ. Sci. Pollut. Res. 2021, 28, 52587–52597. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, U.; Ayub, A.; Hussain, S.; Waraich, E.A.; El-Esawi, M.A.; Ishfaq, M.; Ahmad, M.; Ali, N.; Maqsood, M.F. Cadmium toxicity in plants: Recent progress on morpho-physiological effects and remediation strategies. J. Soil Sci. Plant Nutr. 2022, 22, 212–269. [Google Scholar] [CrossRef]
- Das, P.; Samantaray, S.; Rout, G.R. Studies on cadmium toxicity in plants: A review. Environ. Pollut. 1997, 98, 29–36. [Google Scholar] [CrossRef]
- Gill, S.S.; Khan, N.A.; Tuteja, N. Cadmium at high dose perturbs growth, photosynthesis and nitrogen metabolism while at low dose it up regulates sulfur assimilation and antioxidant machinery in garden cress (Lepidium sativum L.). Plant Sci. 2012, 182, 112–120. [Google Scholar] [CrossRef]
- Yamaguchi, C.; Khamsalath, S.; Takimoto, Y.; Suyama, A.; Mori, Y.; Ohkama-Ohtsu, N.; Maruyama-Nakashita, A. SLIM1 transcription factor promotes sulfate uptake and distribution to shoot, along with phytochelatin accumulation, under cadmium stress in Arabidopsis thaliana. Plants 2020, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.Y.; Wang, T.X.; Shi, Z.; Chiariello, N.R.; Docherty, K.; Field, C.B.; Gutknecht, J.; Gao, Q.; Gu, Y.F.; Guo, X.; et al. Long-term nitrogen deposition enhances microbial capacities in soil carbon stabilization but reduces network complexity. Microbiome 2022, 10, 112. [Google Scholar] [CrossRef]
- Li, W.; Jin, C.; Guan, D.; Wang, Q.; Wang, A.; Yuan, F.; Wu, J. The effects of simulated nitrogen deposition on plant root traits: A meta-analysis. Soil Biol. Biochem. 2015, 82, 112–118. [Google Scholar] [CrossRef]
- Li, W.B.; Zhang, H.X.; Huang, G.Z.; Liu, R.X.; Wu, H.J.; Zhao, C.Y.; McDowell, N.G. Effects of nitrogen enrichment on tree carbon allocation: A global synthesis. Glob. Ecol. Biogeogr. 2020, 29, 573–589. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, W.W.; Li, N.; Liu, Y.Y.; Zheng, X.B.; Hao, G.Y. Photosynthesis and growth responses of Fraxinus mandshurica Rupr. seedlings to a gradient of simulated nitrogen deposition. Ann. For. Sci. 2018, 75, 1. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.X.; Li, B.Y.; Penuelas, J.; Sardans, J.; Yu, H.; Chen, X.P.; Deng, X.Y.; Cheng, D.L.; Zhong, Q.L. Response of functional traits in Machilus pauhoi to nitrogen addition is influenced by differences of provenances. For. Ecol. Manag. 2022, 513, 120207. [Google Scholar] [CrossRef]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.Y.; Yang, F.; He, X.Y.; Du, X.Y.; Mu, P.; Ma, W.J. Advances in the functional study of glutamine synthetase in plant abiotic stress tolerance response. Crop J. 2022, 10, 917–923. [Google Scholar] [CrossRef]
- Meng, S.; Zhang, C.; Su, L.; Li, Y.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Maglovski, M.; Gregorová, Z.; Rybanský, Ľ.; Mészáros, P.; Moravčíková, J.; Hauptvogel, P.; Adamec, L.; Matušíková, I. Nutrition supply affects the activity of pathogenesis-related β-1,3-glucanases and chitinases in wheat. Plant Growth Regul. 2017, 81, 443–453. [Google Scholar] [CrossRef]
- Kasera, N.; Kolar, P.; Hall, S.G. Nitrogen-doped biochars as adsorbents for mitigation of heavy metals and organics from water: A review. Biochar 2022, 4, 17. [Google Scholar] [CrossRef]
- Wångstrand, H.; Eriksson, J.; Öborn, I. Cadmium concentration in winter wheat as affected by nitrogen fertilization. Eur. J. Agron. 2007, 26, 209–214. [Google Scholar] [CrossRef]
- Li, Z.; Wang, X.; Liu, Y.; Zhou, Y.; Qian, Z.; Yu, Z.; Wu, N.; Bian, Z. Water uptake and hormone modulation responses to nitrogen supply in Populus simonii under PEG-induced drought stress. Forests 2022, 13, 907. [Google Scholar] [CrossRef]
- Zhang, F.; Li, J.; Huang, J.; Lin, L.; Wan, X.; Zhao, J.; Dong, J.; Sun, L.; Chen, Q. Transcriptome profiling reveals the important role of exogenous nitrogen in alleviating cadmium toxicity in poplar plants. J. Plant Growth Regul. 2017, 36, 942–956. [Google Scholar] [CrossRef]
- Yu, F.; Yi, L.; Mao, X.; Song, Q.; Korpelainen, H.; Liu, M. Nitrogen addition alleviated sexual differences in responses to cadmium toxicity by regulating the antioxidant system and root characteristics, and inhibiting Cd translocation in mulberry seedlings. Ecotoxicol. Environ. Saf. 2022, 232, 113288. [Google Scholar] [CrossRef]
- Wang, S.; Wei, M.; Wu, B.; Cheng, H.; Wang, C. Combined nitrogen deposition and Cd stress antagonistically affect the allelopathy of invasive alien species Canada goldenrod on the cultivated crop lettuce. Sci. Hortic. 2020, 261, 108955. [Google Scholar] [CrossRef]
- Lin, Z.; Dou, C.; Li, Y.; Wang, H.; Niazi, N.K.; Zhang, S.; Liu, D.; Zhao, K.; Fu, W.; Li, Y.; et al. Nitrogen fertilizer enhances zinc and cadmium uptake by hyperaccumulator Sedum alfredii Hance. J. Soil. Sediment. 2020, 20, 320–329. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.; Chen, J.; Zhang, S.; Xu, J.; Han, X.; Feng, Y.; Chen, Y.; Zhang, X.; Dong, G. Xylem development, cadmium bioconcentration, and antioxidant defense in Populus × euramericana stems under combined conditions of nitrogen and cadmium. Environ. Exp. Bot. 2019, 164, 1–9. [Google Scholar] [CrossRef]
- Yang, C.; Qiu, W.; Chen, Z.; Chen, W.; Li, Y.; Zhu, J.; Rahman, S.U.; Han, Z.; Jiang, Y.; Yang, G. Phosphorus influence Cd phytoextraction in Populus stems via modulating xylem development, cell wall Cd storage and antioxidant defense. Chemosphere 2020, 242, 125154. [Google Scholar] [CrossRef] [PubMed]
- Altas, S.; Uzal, O. Mitigation of negative impacts of cadmium stress on physiological parameters of curly lettuce (Lactuca sativa var. Crispa) by proline treatments. J. Elementol. 2022, 27, 351–365. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, J.; Sun, Y.; Wang, H.; Zhan, J.; Huang, Y.; Zou, J.; Wang, L.; Su, N.; Cui, J. Mechanisms of calcium sulfate in alleviating cadmium toxicity and accumulation in pak choi seedlings. Sci. Total Environ. 2022, 805, 150115. [Google Scholar] [CrossRef]
- Hu, M.; Dou, Q.; Cui, X.; Lou, Y.; Zhuge, Y. Polyaspartic acid mediates the absorption and translocation of mineral elements in tomato seedlings under combined copper and cadmium stress. J. Integr. Agric. 2019, 18, 1130–1137. [Google Scholar] [CrossRef]
- Chen, H.; Li, Y.; Ma, X.; Guo, L.; He, Y.; Ren, Z.; Kuang, Z.; Zhang, X.; Zhang, Z. Analysis of potential strategies for cadmium stress tolerance revealed by transcriptome analysis of upland cotton. Sci. Rep. 2019, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Li, X.; Xu, Z.; Lin, G. Application of fluorescence analysis technology in the study of crop response to cadmium stress. Spectrosc. Spect. Anal. 2020, 40, 3118–3122. [Google Scholar]
- Liu, M.; Korpelainen, H.; Dong, L.; Yi, L. Physiological responses of Elaeocarpus glabripetalus seedlings exposed to simulated acid rain and cadmium. Ecotoxicol. Environ. Saf. 2019, 175, 118–127. [Google Scholar] [CrossRef]
- Jin, C.; Zhou, Q.; Zhou, Q.; Fan, J. Effects of chlorimuron-ethyl and cadimum on biomass growth and cadimum accumulation of wheat in the phaiozem area, northeast China. Bulle. Environ. Contam. Toxicol. 2010, 84, 395–400. [Google Scholar] [CrossRef]
- Zhou, Q.X.; Kong, F.X.; Zhu, L. Ecotoxicology; Science Press: Beijing, China, 2004. (In Chinese) [Google Scholar]
- Ti, C.; Gao, B.; Luo, Y.; Wang, S.; Chang, S.X.; Yan, X. Dry deposition of N has a major impact on surface water quality in the Taihu Lake region in southeast China. Atmos. Environ. 2018, 190, 1–9. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef]
- Galloway, J.; Dentener, F.; Boyer, E.; Howarth, R.; Seitzinger, S.; Asner, G.; Cleveland, C.; Green, P.; Holland, E.; Karl, D.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Newell, E.A.; Mulkey, S.S.; Wright, J.S. Seasonal patterns of carbohydrate storage in four tropical tree species. Oecologia 2002, 131, 333–342. [Google Scholar] [CrossRef]
- Pereira, T.S.; Pereira, T.S.; Figueredo de Carvalho Souza, C.L.; Alvino Lima, E.J.; Batista, B.L.; da Silva Lobato, A.K. Silicon deposition in roots minimizes the cadmium accumulation and oxidative stress in leaves of cowpea plants. Physiol. Mol. Biol. Plants 2018, 24, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Guan, J.; Liang, Q.; Zhang, X.; Hu, H.; Zhang, J. Effects of cadmium stress on growth and physiological characteristics of sassafras seedlings. Sci. Rep. 2021, 11, 9913. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Elkelish, A.; Soliman, M.; Elansary, H.O.; Zaid, A.; Wani, S.H. Serratia marcescens BM1 enhances cadmium stress tolerance and phytoremediation potential of soybean through modulation of osmolytes, leaf gas exchange, antioxidant machinery, and stress-responsive genes expression. Antioxidants 2020, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yang, W.; Yang, T.; Chen, Y.; Ni, W. Effects of cadmium stress on leaf chlorophyll fluorescence and photosynthesis of Elsholtzia argyi-A cadmium accumulating plant. Int. J. Phytopharm. 2015, 17, 85–92. [Google Scholar]
- Khan, M.I.R.; Nazir, F.; Asgher, M.; Per, T.S.; Khan, N.A. Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. J. Plant Physiol. 2015, 173, 9–18. [Google Scholar] [CrossRef]
- Hasan, S.A.; Hayat, S.; Ahmad, A. Brassinosteroids protect photosynthetic machinery against the cadmium induced oxidative stress in two tomato cultivars. Chemosphere 2011, 84, 1446–1451. [Google Scholar] [CrossRef]
- Ci, D.; Jiang, D.; Wollenweber, B.; Dai, T.; Jing, Q.; Cao, W. Cadmium stress in wheat seedlings: Growth, cadmium accumulation and photosynthesis. Acta Physiol. Plant. 2010, 32, 365–373. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Choudhary, S.P.; Chen, S.; Xia, X.; Shi, K.; Zhou, Y.; Yu, J. Role of brassinosteroids in alleviation of phenanthrene–cadmium co-contamination-induced photosynthetic inhibition and oxidative stress in tomato. J. Exp. Bot. 2013, 64, 199–213. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Effah, Z.; Li, L.; Xie, J.; Liu, C.; Xu, A.; Karikari, B.; Anwar, S.; Zeng, M. Regulation of nitrogen metabolism, photosynthetic activity, and yield attributes of spring wheat by nitrogen fertilizer in the semi-arid Loess Plateau region. J. Plant Growth Regul. 2022, 42, 1120–1133. [Google Scholar] [CrossRef]
- Leiter, T.d.S.; Monteiro, F.A. Partial replacement of nitrate by ammonium increases photosynthesis and reduces oxidative stress in Tanzania guinea grass exposed to cadmium. Ecotoxicol. Environ. Saf. 2019, 174, 592–600. [Google Scholar] [CrossRef]
- Yang, Y.; Xiong, J.; Tao, L.; Cao, Z.; Tang, W.; Zhang, J.; Yu, X.; Fu, G.; Zhang, X.; Lu, Y. Regulatory mechanisms of nitrogen (N) on cadmium (Cd) uptake and accumulation in plants: A review. Sci. Total Environ. 2020, 708, 135186. [Google Scholar] [CrossRef]
- Song, J.; Finnegan, P.M.; Liu, W.; Li, X.; Yong, J.W.H.; Xu, J.; Zhang, Q.; Wen, Y.; Qin, K.; Guo, J.; et al. Mechanisms underlying enhanced Cd translocation and tolerance in roots of Populus euramericana in response to nitrogen fertilization. Plant Sci. 2019, 287, 110206. [Google Scholar] [CrossRef]
- Chi, K.; Zou, R.; Wang, L.; Huo, W.; Fan, H. Cellular distribution of cadmium in two amaranth (Amaranthus mangostanus L.) cultivars differing in cadmium accumulation. Environ. Sci. Pollut. Res. 2019, 26, 22147–22158. [Google Scholar] [CrossRef]
- Shackira, A.M.; Puthur, J.T. Cd2+ influences metabolism and elemental distribution in roots of Acanthus ilicifolius L. Int. J. Phytoremediat. 2019, 21, 866–877. [Google Scholar] [CrossRef]
- Zhang, D.; Dong, F.; Zhang, Y.; Huang, Y.; Zhang, C. Mechanisms of low cadmium accumulation in storage root of sweet potato (Ipomoea batatas L.). J. Plant Physiol. 2020, 254, 153262. [Google Scholar] [CrossRef]
- Yu, S.; Deng, H.; Zhang, B.; Liu, Z.; Lin, J.; Sheng, L.; Pan, J.; Huang, L.; Qi, J. Physiological response of Vetiveria zizanioides to cadmium stress revealed by Fourier transform infrared spectroscopy. Spectrosc. Lett. 2022, 55, 157–165. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, C.; Li, S.; Li, B.; Li, Q.; Chen, G.; Chen, W.; Wang, F. Cadmium adsorption, chelation and compartmentalization limit root-to-shoot translocation of cadmium in rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2017, 24, 11319–11330. [Google Scholar] [CrossRef]
- Tan, S.; Liu, Z.; Zeng, Q.; Zhu, M.; Wang, A.; Chen, B. Nutrient allocation might affect the cadmium accumulation of Bermuda grass (Cynodon dactylon). Chemosphere 2020, 252, 126512. [Google Scholar] [CrossRef]
- Yang, J.; Sun, H.; Qin, J.; Wang, X.; Chen, W. Impacts of Cd on temporal dynamics of nutrient distribution pattern of Bletilla striata, a traditional Chinese medicine plant. Agriculture 2021, 11, 594. [Google Scholar] [CrossRef]
- Mourato, M.; Pinto, F.; Moreira, I.; Sales, J.; Leitão, I.; Martins, L.L. Chapter 13—The effect of Cd stress in mineral nutrient uptake in plants. In Cadmium Toxicity and Tolerance in Plants; Hasanuzzaman, M., Prasad, M.N.V., Fujita, M., Eds.; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Zhang, C.; Yi, X.; Gao, X.; Wang, M.; Shao, C.; Lv, Z.; Chen, J.; Liu, Z.; Shen, C. Physiological and biochemical responses of tea seedlings (Camellia sinensis) to simulated acid rain conditions. Ecotoxicol. Environ. Saf. 2020, 192, 110315. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Ruecker, A.; Schmidt, A.; Gleixner, G.; Gershenzon, J.; Trumbore, S.; Hartmann, H. Production of constitutive and induced secondary metabolites is coordinated with growth and storage in Norway spruce saplings. Tree Physiol. 2020, 40, 928–942. [Google Scholar] [CrossRef] [Green Version]
- Ravi, S.; Bader, M.K.F.; Young, T.; Duxbury, M.; Clearwater, M.; Macinnis-Ng, C.; Leuzinger, S. Are the well-fed less thirsty? Effects of drought and salinity on New Zealand mangroves. J. Plant Ecol. 2022, 15, 85–99. [Google Scholar] [CrossRef]
- Ravi, S.; Young, T.; Macinnis-Ng, C.; Nyugen, T.V.; Duxbury, M.; Alfaro, A.C.; Leuzinger, S. Untargeted metabolomics in halophytes: The role of different metabolites in New Zealand mangroves under multi-factorial abiotic stress conditions. Environ. Exp. Bot. 2020, 173, 103993. [Google Scholar] [CrossRef]
- Xu, L.; Wu, X.; Zhou, Z. Effects of physiological integration and fertilization on heavy metal remediation in soil by a clonal grass. Pol. J. Environ. Stud. 2016, 25, 395–404. [Google Scholar] [CrossRef]
- Li, M.H.; Cherubini, P.; Dobbertin, M.; Arend, M.; Xiao, W.F.; Rigling, A. Responses of leaf nitrogen and mobile carbohydrates in different Quercus species/provenances to moderate climate changes. Plant Biol. 2013, 15, 177–184. [Google Scholar] [CrossRef]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Liu, F. Carbon allocation of Chinese pine seedlings along a nitrogen addition gradient. For. Ecol. Manag. 2014, 334, 114–121. [Google Scholar] [CrossRef]
- Cui, Z.; Yang, Z.; Xu, D.; Li, X. Drought could promote the heartwood formation in Dalbergia odorifera by enhancing the transformation of starch to soluble sugars. Scand. J. For. Res. 2022, 37, 23–32. [Google Scholar] [CrossRef]
- Shang, X.; Xue, W.; Jiang, Y.; Zou, J. Effects of calcium on the alleviation of cadmium toxicity in Salix matsudana and its effects on other minerals. Pol. J. Environ. Stud. 2020, 29, 2001–2010. [Google Scholar] [CrossRef]
- Cheng, Y.; Zou, Z.; Ye, Z.; Cheng, Y.; Song, Y.; Li, R.; Li, J. Effect of iron oxide nanoparticles on cadmium stress in melon. Fresenius Environ. Bull. 2020, 29, 8988–8997. [Google Scholar]
- Zhu, T.; Li, L.; Duan, Q.; Liu, X.; Chen, M. Progress in our understanding of plant responses to the stress of heavy metal cadmium. Plant Signal. Behav. 2021, 16, 1836884. [Google Scholar] [CrossRef]
- Mccarthy, M.C.; Enquist, B.J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Funct. Ecol. 2007, 21, 713–720. [Google Scholar] [CrossRef]
- Bari, M.A.; Akther, M.S.; Reza, M.A.; Kabir, A.H. Cadmium tolerance is associated with the root-driven coordination of cadmium sequestration, iron regulation, and ROS scavenging in rice. Plant Physiol. Biochem. 2019, 136, 22–33. [Google Scholar] [CrossRef]
- Zhang, F.; Wan, X.; Zhong, Y. Nitrogen as an important detoxification factor to cadmium stress in poplar plants. J. Plant Interact. 2014, 9, 249–258. [Google Scholar] [CrossRef]
- Huang, J.; Wu, X.; Tian, F.; Chen, Q.; Luo, P.; Zhang, F.; Wan, X.; Zhong, Y.; Liu, Q.; Lin, T. Changes in proteome and protein phosphorylation reveal the protective roles of exogenous nitrogen in alleviating cadmium toxicity in poplar plants. Int. J. Mol. Sci. 2020, 21, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Bi, J.; Liu, X.; Kang, J.; Korpelainen, H.; Niinemets, U.; Li, C. Microstructural and physiological responses to cadmium stress under different nitrogen levels in Populus cathayana females and males. Tree Physiol. 2020, 40, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Edaphic stress interactions: Important yet poorly understood drivers of plant production in future climates. Field Crops Res. 2022, 283, 108547. [Google Scholar] [CrossRef]
- Hannan, A.; Hassan, L.; Hoque, M.N.; Tahjib-Ul-Arif, M.; Robin, A.H.K. Increasing new root length reflects survival mechanism of rice (Oryza sativa L.) genotypes under PEG-induced osmotic stress. Plant Breed. Biotechnol. 2020, 8, 46–57. [Google Scholar] [CrossRef]
- Malamy, J.E. Intrinsic and environmental response pathways that regulate root system architecture. Plant Cell Environ. 2010, 28, 67–77. [Google Scholar] [CrossRef]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Kuriakose, S.V.; Prasad, M.N.V. Cadmium stress affects seed germination and seedling growth in Sorghum bicolor (L.) Moench by changing the activities of hydrolyzing enzymes. Plant Growth Regul. 2008, 54, 143–156. [Google Scholar] [CrossRef]
- Yu, L.; Wang, X.; Li, X.; Wang, Y.; Kang, H.; Chen, G.; Fan, X.; Sha, L.; Zhou, Y.; Zeng, J. Protective effect of different forms of nitrogen application on cadmium-induced toxicity in wheat seedlings. Environ. Sci. Pollut. Res. 2019, 26, 13085–13094. [Google Scholar] [CrossRef]
- Wang, S.; van Dijk, J.; de Boer, H.J.; Wassen, M.J. Source and sink activity of Holcus lanatus in response to absolute and relative supply of nitrogen and phosphorus. Funct. Plant Biol. 2021, 48, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ouyang, Y.; Xu, J.; Liu, K. Cadmium remobilization from shoot to grain is related to pH of vascular bundle in rice. Ecotoxicol. Environ. Saf. 2018, 147, 913–918. [Google Scholar] [CrossRef]
- Shaar-Moshe, L.; Hayouka, R.; Roessner, U.; Peleg, Z. Phenotypic and metabolic plasticity shapes life-history strategies under combinations of abiotic stresses. Plant Direct 2019, 3, e00113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertens, D.; Boege, K.; Kessler, A.; Koricheva, J.; Thaler, J.S.; Whiteman, N.K.; Poelman, E.H. Predictability of biotic stress structures plant defence evolution. Trends Ecol. Evol. 2021, 36, 444–456. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of Cd and N addition on leaf net photosynthesis rate (Pn) (A), stomatal conductance (gs) (B), intercellular CO2 concentration (Ci) (C), transpiration (Tr) (D) and water use efficiency (WUE) (E) in E. glabripetalus seedlings. Different letters indicate significant differences between groups (mean ± SE, n = 3) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.
Figure 1.
Effects of Cd and N addition on leaf net photosynthesis rate (Pn) (A), stomatal conductance (gs) (B), intercellular CO2 concentration (Ci) (C), transpiration (Tr) (D) and water use efficiency (WUE) (E) in E. glabripetalus seedlings. Different letters indicate significant differences between groups (mean ± SE, n = 3) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.
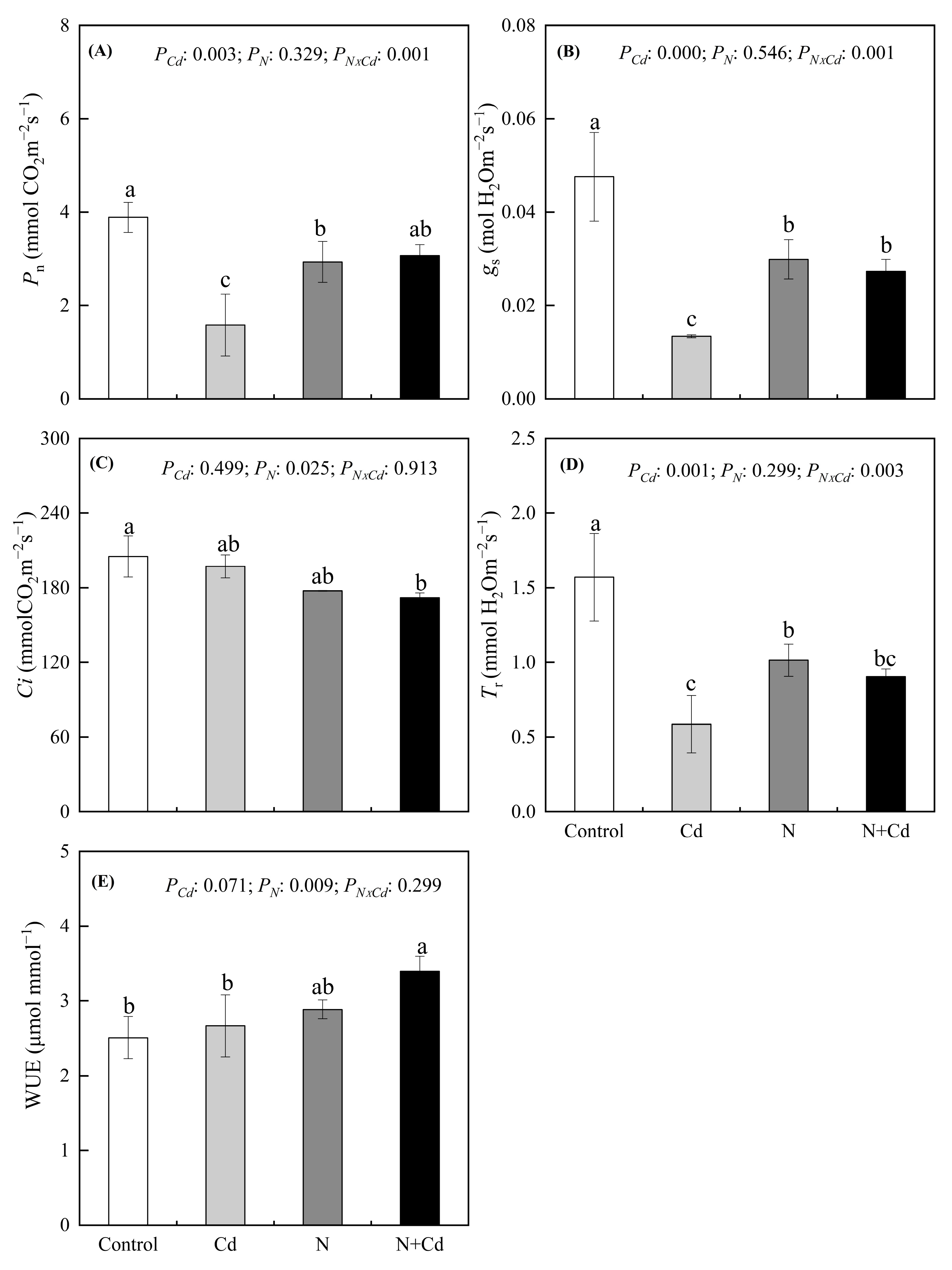
Figure 2.
Effects of Cd and N addition on leaf biomass (A), stem biomass (B), root biomass (C), total biomass (D) and root to shoot ratio (E) in E. glabripetalus. Different letters indicate significant differences between groups (mean ± SE, n = 4) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.
Figure 2.
Effects of Cd and N addition on leaf biomass (A), stem biomass (B), root biomass (C), total biomass (D) and root to shoot ratio (E) in E. glabripetalus. Different letters indicate significant differences between groups (mean ± SE, n = 4) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.

Figure 3.
Effects of Cd and N addition on N contents (A), P contents (B), and N/P ratio (C) in roots, stem and leaves of E. glabripetalus. Different letters indicate significant differences between groups (mean ± SE, n = 3) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.
Figure 3.
Effects of Cd and N addition on N contents (A), P contents (B), and N/P ratio (C) in roots, stem and leaves of E. glabripetalus. Different letters indicate significant differences between groups (mean ± SE, n = 3) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.
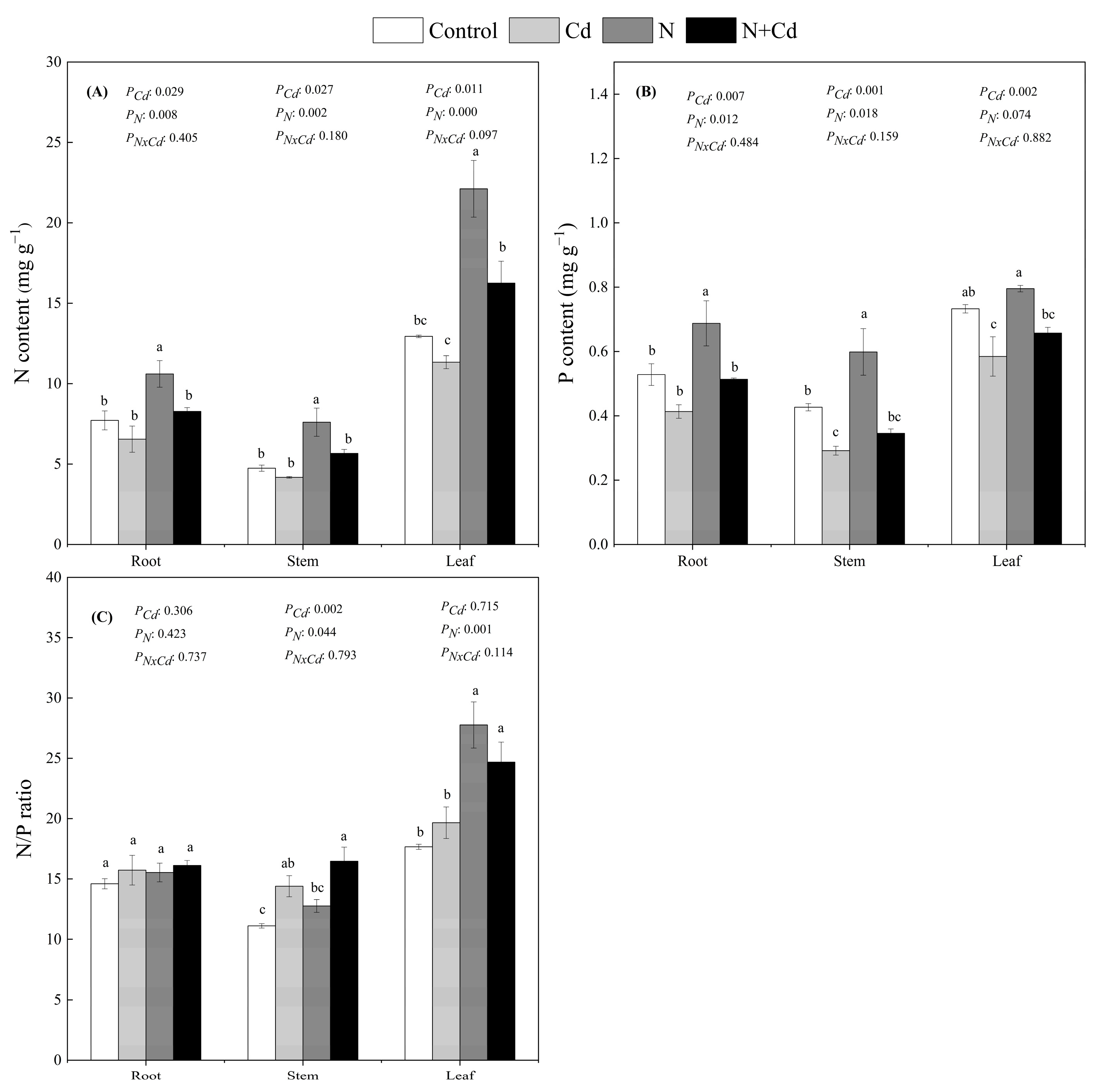
Figure 4.
Effects of Cd and N addition on soluble sugars (A), starch (B), non-structural carbohydrate (C) and sugar/starch ratio (D) in roots, stems and leaves of E. glabripetalus. Different letters indicate significant differences between groups (mean ± SE, n = 3) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.
Figure 4.
Effects of Cd and N addition on soluble sugars (A), starch (B), non-structural carbohydrate (C) and sugar/starch ratio (D) in roots, stems and leaves of E. glabripetalus. Different letters indicate significant differences between groups (mean ± SE, n = 3) (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd.
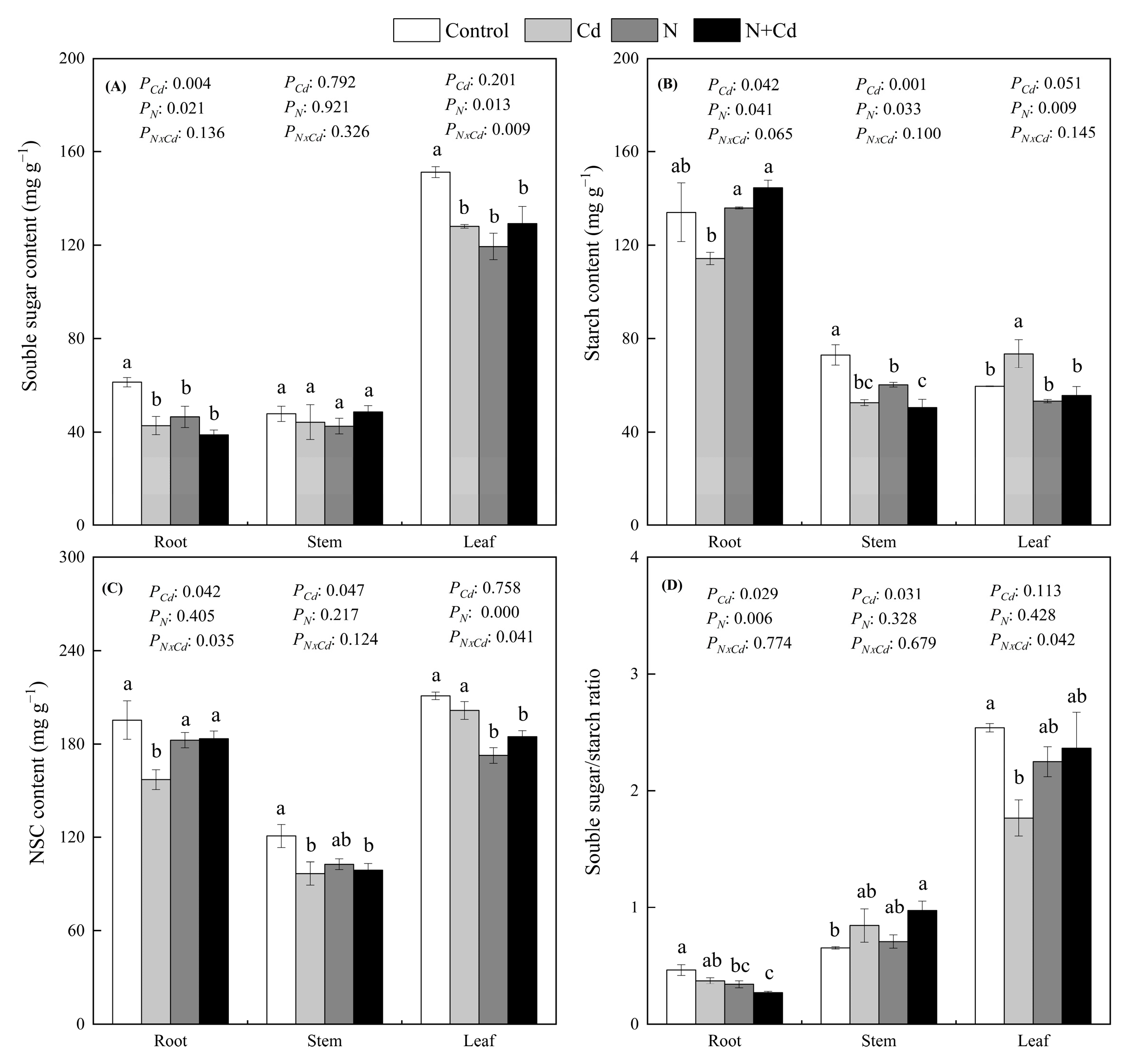
Figure 5.
Correlation between the indicators in E. glabripetalus seedlings. Data are Spearman correlation coefficients. Positive correlations are shown in red and negative correlations are shown in blue, and the darker the color, the stronger the correlation. * indicates a significant correlation at the 0.05 level.
Figure 5.
Correlation between the indicators in E. glabripetalus seedlings. Data are Spearman correlation coefficients. Positive correlations are shown in red and negative correlations are shown in blue, and the darker the color, the stronger the correlation. * indicates a significant correlation at the 0.05 level.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of Cd and N addition on root morphology of E. glabripetalus.
| N (kg N ha−1 yr−1) | Cd (mg kg−1) | Total Root Length (cm) | Total Root Surface Area (cm2) | Average Root Diameter (mm) | Total Root Volume (cm3) | The Proportion of the Length of Fine Roots with Different Diameter Classes to Total Root Length (%) | ||
|---|---|---|---|---|---|---|---|---|
| 0–0.5 mm | 0.5–1 mm | 1–2 mm | ||||||
| 0 | 0 | 6236.20 ± 626.70 b | 1080.60 ± 34.40 b | 0.52 ± 0.02 b | 16.79 ± 0.54 b | 66.88 ± 1.00 ab | 24.97 ± 0.04 bc | 8.15 ± 0.97 b |
| 0 | 100 | 6445.24 ± 1815.82 b | 1342.86 ± 389.49 b | 0.64 ± 0.01 a | 23.99 ± 3.72 b | 56.53 ± 0.16 c | 31.40 ± 0.26 a | 12.07 ± 0.09 a |
| 90 | 0 | 15,991.44 ± 1747.08 a | 2318.75 ± 250.20 a | 0.50 ± 0.02 b | 23.18 ± 4.68 b | 72.09 ± 3.01 a | 21.88 ± 2.16 c | 6.02 ± 0.91 b |
| 90 | 100 | 14,579.63 ± 3224.05 a | 2602.361 ± 549.36 a | 0.55 ± 0.01 b | 39.36 ± 5.53 a | 64.37 ± 1.32 b | 27.86 ± 0.59 ab | 7.77 ± 0.73 b |
| PCd | NS | NS | ** | * | * | * | * | |
| PN | ** | ** | * | * | * | NS | * | |
| PN × Cd | NS | NS | NS | NS | NS | NS | NS | |
Mean values ± SE (n = 4) are shown. Different letters indicate significant differences between groups (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd. NS, no significance; * p < 0.05 and ** p ≤ 0.01.
Table 2.
Effects of Cd and N addition on Cd contents in the root, stem and leaves of E. glabripetalus seedlings.
Table 2.
Effects of Cd and N addition on Cd contents in the root, stem and leaves of E. glabripetalus seedlings.
| N (kg N·ha−1 yr−1) | Cd (mg·kg−1 Dry Soil) | Root Cd (mg·kg−1) | Stem Cd (mg·kg−1) | Leaf Cd (mg·kg−1) |
|---|---|---|---|---|
| 0 | 0 | 1.95 ± 0.35 c | 1.78 ± 0.19 c | 0.18 ± 0.05 c |
| 0 | 100 | 1701.01 ± 53.06 b | 12.2 ± 0.64 b | 2.08 ± 1.13 b |
| 90 | 0 | 2.85 ± 0.02 c | 2.78 ± 0.71 c | 0.48 ± 0.11 bc |
| 90 | 100 | 2451.07 ± 392.77 a | 15.62 ± 0.93 a | 5.43 ± 0.14 a |
| PCd | ** | ** | ** | |
| PN | NS | * | * | |
| PN × Cd | NS | NS | * |
Mean values ± SE (n = 3) are shown. Different letters indicate significant differences between groups (p < 0.05). PCd, Cd effect; PN, N effect; PN × Cd, the interactive effect of N and Cd. NS, no significance; * p < 0.05 and ** p ≤ 0.01.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, M.; Shi, M.; Gao, H.; Zheng, Y.; Yi, L. Nitrogen Addition Alleviates Cadmium Toxicity in Eleocarpus glabripetalus Seedlings. Forests 2023, 14, 1264. https://doi.org/10.3390/f14061264
AMA Style
Liu M, Shi M, Gao H, Zheng Y, Yi L. Nitrogen Addition Alleviates Cadmium Toxicity in Eleocarpus glabripetalus Seedlings. Forests. 2023; 14(6):1264. https://doi.org/10.3390/f14061264
Chicago/Turabian StyleLiu, Meihua, Mengjiao Shi, Haili Gao, Yu Zheng, and Lita Yi. 2023. "Nitrogen Addition Alleviates Cadmium Toxicity in Eleocarpus glabripetalus Seedlings" Forests 14, no. 6: 1264. https://doi.org/10.3390/f14061264
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.