
Soil Phosphorus Availability Controls Deterministic and Stochastic Processes of Soil Microbial Community along an Elevational Gradient in Subtropical Forests
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location
2.2. Soil Samples and Data Collection
2.3. Soil Physicochemical Analyses
2.4. DNA Extraction and High-Throughput Amplicon Sequencing
2.5. Statistical Analyses
2.5.1. Soil Microbial Community Structure Analysis
2.5.2. Habitat Niche Breadth Analysis
2.5.3. Null Model Analysis
2.5.4. Correlation Analysis
3. Results
3.1. Soil Microbial Community Composition and Structure
3.2. Habitat Niche Breadth
3.3. Soil Microbial Community Assembly Processes and Their Influencing Factors
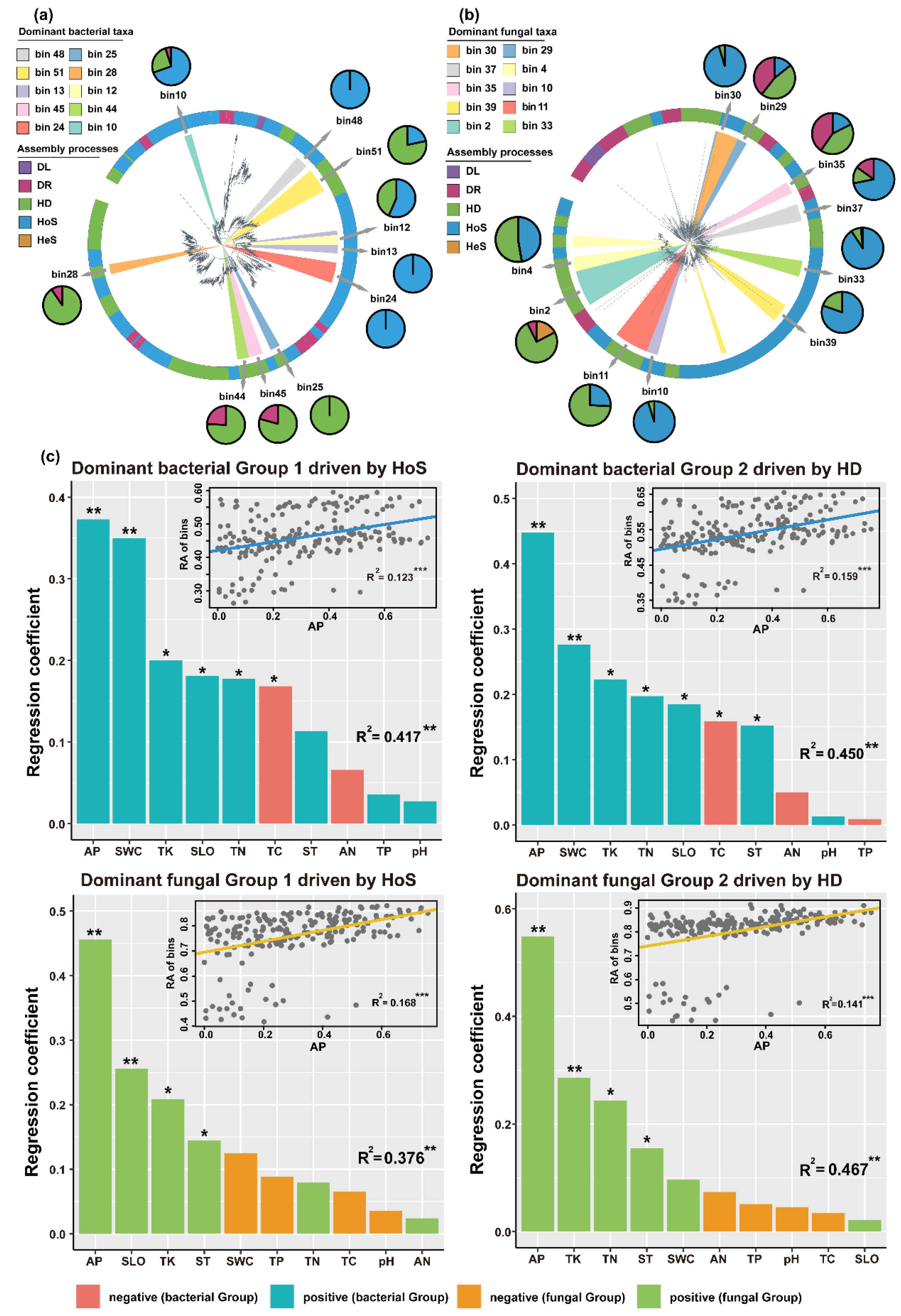
3.4. Soil Microbial Community Assembly Processes Based on Individual Lineages
3.5. Correlation between Environmental Factors and Dominant Microbial Taxa
4. Discussion
4.1. Structure of Soil Microbial Community along the Elevational Gradient
4.2. Mechanisms for the Soil Microbial Community Assembly
4.3. Assembly Processes of the Dominant Microbial Taxa
4.4. Limitations and Prospects of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, I.M.; Crawford, J.W. Interactions and self-organization in the soil-microbe complex. Science 2004, 304, 1634–1637. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth’s biogeochemical cycles. Science 2008, 320, 1034–1039. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.Z.; Ning, D.L. Stochastic community assembly: Does it matter in microbial ecology? Microbiol. Mol. Biol. Rev. 2017, 81, e00002–e00017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B.H. Beyond biogeographic patterns: Processes shaping the microbial landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef]
- Gao, C.; Guo, L.D. Progress on microbial species diversity, community assembly and functional traits. Biodivers. Sci. 2022, 30, 22429. (In Chinese) [Google Scholar] [CrossRef]
- Chalmandrier, L.; Pansu, J.; Zinger, L.; Boyer, F.; Coissac, E.; Génin, A.; Gielly, L.; Lavergne, S.; Legay, N.; Schilling, V.; et al. Environmental and biotic drivers of soil microbial β-diversity across spatial and phylogenetic scales. Ecography 2019, 42, 2144–2156. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.J.; Fredrickson, J.K.; Konopka, A.E. Estimating and mapping ecological processes influencing microbial community assembly. Front. Microbiol. 2015, 6, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martiny, J.B.H.; Eisen, J.A.; Penn, K.; Allison, S.D.; Horner-Devine, M.C. Drivers of bacterial β-diversity depend on spatial scale. Proc. Natl. Acad. Sci. USA 2011, 108, 7850–7854. [Google Scholar] [CrossRef] [PubMed]
- Stegen, J.C.; Lin, X.J.; Fredrickson, J.K.; Chen, X.Y.; Kennedy, D.W.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying community assembly processes and identifying features that impose them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef]
- Fargione, J.; Brown, C.S.; Tilman, D. Community assembly and invasion: An experimental test of neutral versus niche processes. Proc. Natl. Acad. Sci. USA 2003, 100, 8916–8920. [Google Scholar] [CrossRef] [PubMed]
- Chave, J. Neutral theory and community ecology. Ecol. Lett. 2004, 7, 241–253. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Schmidt, S.K.; Fukami, T.; O’Neill, S.P.; Bilinski, T.M.; Stanish, L.F.; Knelman, J.E.; Darcy, J.L.; Lynch, R.C.; Wickey, P.; et al. Patterns and processes of microbial community assembly. Microbiol. Mol. Biol. Rev. 2013, 77, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Graham, E.B.; Crump, A.R.; Resch, C.T.; Fansler, S.; Arntzen, E.; Kennedy, D.W.; Fredrickson, J.K.; Stegen, J.C. Coupling spatiotemporal community assembly processes to changes in microbial metabolism. Front. Microbiol. 2016, 7, 1949. [Google Scholar] [CrossRef] [Green Version]
- Ning, D.L.; Yuan, M.T.; Wu, L.W.; Zhang, Y.; Guo, X.; Zhou, X.S.; Yang, Y.F.; Arkin, A.P.; Firestone, M.K.; Zhou, J.Z. A quantitative framework reveals ecological drivers of grassland microbial community assembly in response to warming. Nat. Commun. 2020, 11, 4717. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Yang, Y.F.; Feng, J.J.; Tian, R.M.; Guo, X.; Ning, D.L.; Hale, L.; Wang, M.M.; Cheng, J.M.; Wu, L.W.; et al. The spatial scale dependence of diazotrophic and bacterial community assembly in paddy soil. Glob. Ecol. Biogeogr. 2019, 28, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.D.; Li, S.P.; Yang, X.; Zhou, J.Z.; Shu, W.S.; Jiang, L. Mechanisms of soil bacterial and fungal community assembly differ among and within islands. Environ. Microbiol. 2020, 22, 1559–1571. [Google Scholar] [CrossRef]
- Zhang, B.; Xue, K.; Zhou, S.T.; Che, R.X.; Du, J.Q.; Tang, L.; Pang, Z.; Wang, F.; Wang, D.; Cui, X.Y.; et al. Phosphorus mediates soil prokaryote distribution pattern along a small-scale elevation gradient in Noijin Kangsang Peak, Tibetan Plateau. FEMS Microbiol. Ecol. 2019, 95, fiz076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.B.; Shen, Z.H.; Li, C.N.; Kou, Y.P.; Wang, Y.S.; Tu, B.; Zhang, S.H.; Li, X.Z. Stair-step pattern of soil bacterial diversity mainly driven by pH and vegetation types along the elevational gradients of Gongga mountain, China. Front. Microbiol. 2018, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- Shigyo, N.; Umeki, K.; Hirao, T. Plant functional diversity and soil properties control elevational diversity gradients of soil bacteria. FEMS Microbiol. Ecol. 2019, 95, fiz025. [Google Scholar] [CrossRef]
- Yeh, C.F.; Soininen, J.; Teittinen, A.; Wang, J.J. Elevational patterns and hierarchical determinants of biodiversity across microbial taxonomic scales. Mol. Ecol. 2019, 28, 86–99. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.C.; Gunina, A.; Luo, Y.; Wang, J.J.; He, J.Z.; Kuzyakov, Y.; Hemp, A.; Classen, A.T.; Ge, Y. Contrasting patterns and drivers of soil bacterial and fungal diversity across a mountain gradient. Environ. Microbiol. 2020, 22, 3287–3301. [Google Scholar] [CrossRef] [PubMed]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahbek, C.; Borregaard, M.K.; Colwell, R.K.; Dalsgaard, B.; Holt, B.G.; Morueta-Holme, N.; Nogues-Bravo, D.; Whittaker, R.J.; Fjeldså, J. Humboldt’s enigma: What causes global patterns of mountain biodiversity? Science 2019, 365, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.N.; Kolasa, J.; Cottenie, K. Contrasts between habitat generalists and specialists: An empirical extension to the basic metacommunity framework. Ecology 2009, 90, 2253–2262. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.L.; Xiang, W.H.; Ouyang, S.; Forrester, D.I.; Zhou, B.; Chen, L.X.; Ge, T.D.; Lei, P.F.; Chen, L.; Zeng, Y.L.; et al. Linkage between tree species richness and soil microbial diversity improves phosphorus bioavailability. Funct. Ecol. 2019, 33, 1549–1560. [Google Scholar] [CrossRef]
- Zhou, W.S.; Zhang, Y.X.; Zhang, S.; Yakimov, B.N.; Ma, K.M. Phylogenetic and Functional Traits Verify the Combined Effect of Deterministic and Stochastic Processes in the Community Assembly of Temperate Forests along an Elevational Gradient. Forests 2021, 12, 591. [Google Scholar] [CrossRef]
- Teste, F.P.; Lambers, H.; Enowashu, E.E.; Laliberté, E.; Marhan, S.; Kandeler, E. Soil microbial communities are driven by the declining availability of cations and phosphorus during ecosystem retrogression. Soil Biol. Biochem. 2021, 163, 108430. [Google Scholar] [CrossRef]
- Siles, J.A.; Margesin, R. Abundance and diversity of bacterial, archaeal, and fungal communities along an altitudinal gradient in Alpine forest soils: What are the driving factors? Microb. Ecol. 2016, 72, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.M.; Zhou, Z.; Chen, J.; Xu, H.; Ma, S.H.; Dippold, M.A.; Kuzyakov, Y. Long-term nitrogen and phosphorus fertilization reveals that phosphorus limitation shapes the microbial community composition and functions in tropical montane forest soil. Sci. Total Environ. 2023, 854, 158709. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.H.; Wang, Z.F.; Liu, Y.; Yang, T.Y.; Chen, W.M.; Wei, G.H.; Jiao, S. Soil phosphorus determines the distinct assembly strategies for abundant and rare bacterial communities during successional reforestation. Soil Ecol. Lett. 2021, 3, 342–355. [Google Scholar] [CrossRef]
- Xu, D.W.; Liu, J.F.; Marshall, P.; He, Z.S.; Zheng, S.Q. Leaf litter decomposition dynamics in unmanaged Phyllostachys pubescens stands at high elevations in the Daiyun Mountain National Nature Reserve. J. Mt. Sci. 2017, 14, 2246–2256. [Google Scholar] [CrossRef]
- He, Z.S.; Gu, X.G.; Jiang, L.; Xu, D.W.; Liu, J.F.; Li, W.Z.; Chen, W.W. Characteristics and its influencing factors of forest soil dominant bacterial community in different elevations on the southern slope of Daiyun Mountain, Fujian Province of eastern China. J. Beijing For. Univ. 2022, 44, 107–116. (In Chinese) [Google Scholar]
- Jiang, L.; He, Z.S.; Liu, J.F.; Xing, C.; Gu, X.G.; Wei, C.S.; Zhu, J.; Wang, X.L. Elevation gradient altered soil C, N, and P stoichiometry of Pinus taiwanensis forest on Daiyun mountain. Forests 2019, 10, 1089. [Google Scholar] [CrossRef] [Green Version]
- He, Z.S.; Wang, Z.W.; Zhu, J.; Chen, J.J.; Gu, X.G.; Jiang, L.; Chen, B.; Wu, Z.Y.; Liu, J.F.; Chen, W.W. Forest soil microbial community structure characteristics and its influencing factors at different elevations on the southern slope of Daiyun mountain. Environ. Sci. 2022, 43, 2802–2811. (In Chinese) [Google Scholar]
- Wang, W.T.; Sun, Z.H.; Mishra, S.; Xia, S.W.; Lin, L.X.; Yang, X.D. Body size determines multitrophic soil microbiota community assembly associated with soil and plant attributes in a tropical seasonal rainforest. Mol. Ecol. 2022. early view. [Google Scholar] [CrossRef] [PubMed]
- Li, M.J.; He, Z.S.; Jiang, L.; Gu, X.G.; Jin, M.G.; Chen, B.; Liu, J.F. Distribution pattern and driving factors of species diversity and phylogenetic diversity along altitudinal gradient on the south slope of Daiyun Mountain. Acta Ecol. Sin. 2021, 41, 1148–1157. (In Chinese) [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Dai, T.J.; Zhang, Y.; Tang, Y.S.; Bai, Y.H.; Tao, Y.L.; Huang, B.; Wen, D.H. Identifying the key taxonomic categories that characterize microbial community diversity using full-scale classification: A case study of microbial communities in the sediments of Hangzhou Bay. FEMS Microbiol. Ecol. 2016, 92, fiw 150. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.T.; Xiao, X.; Nuccio, E.E.; Yuan, M.T.; Zhang, N.; Xue, K.; Cohan, F.M.; Zhou, J.Z.; Sun, B. Differentiation strategies of soil rare and abundant microbial taxa in response to changing climatic regimes. Environ. Microbiol. 2020, 22, 1327–1340. [Google Scholar] [CrossRef]
- Subramanian, B.; Gao, S.H.; Lercher, M.J.; Hu, S.N.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.X.; Lu, H.P.; Sastri, A.; Yeh, Y.C.; Gong, G.C.; Chou, W.C.; Hsieh, C.H. Contrasting the relative importance of species sorting and dispersal limitation in shaping marine bacterial versus protist communities. ISME J. 2018, 12, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.L. Spaa 0.2.2—Species Association Analysis, R Package Version 0.2.2; The R Project for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Yang, L.Y.; Ning, D.L.; Yang, Y.F.; He, N.P.; Li, X.Z.; Cornell, C.R.; Bates, C.T.; Filimonenko, E.; Kuzyakov, Y.; Zhou, J.Z.; et al. Precipitation balances deterministic and stochastic processes of bacterial community assembly in grassland soils. Soil Biol. Biochem. 2022, 168, 108635. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Lü, X.T.; Yao, J.; Wang, Z.W.; Deng, Y.; Cheng, W.X.; Zhou, J.Z.; Han, X.G. Habitat-specific patterns and drivers of bacterial β-diversity in China’s drylands. ISME J. 2017, 11, 1345–1358. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package, R package version 2.4-3; The R Project for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Lichstein, J.W. Multiple regression on distance matrices: A multivariate spatial analysis tool. Plant Ecol. 2007, 188, 117–131. [Google Scholar] [CrossRef]
- Chen, L.; Xiang, W.H.; Wu, H.L.; Ouyang, S.; Lei, P.F.; Hu, Y.J.; Ge, T.D.; Ye, J.; Kuzyakov, Y. Contrasting patterns and drivers of soil fungal communities in subtropical deciduous and evergreen broadleaved forests. Appl. Microbiol. Biotechnol. 2019, 103, 5421–5433. [Google Scholar] [CrossRef]
- Wang, X.L.; Liu, J.F.; He, Z.S.; Xing, C.; Zhu, J.; Gu, X.G.; Lan, Y.Q.; Wu, Z.Y.; Liao, P.C.; Zhu, D.H. Forest gaps mediate the structure and function of the soil microbial community in a Castanopsis kawakamii forest. Ecol. Indic. 2021, 122, 107288. [Google Scholar] [CrossRef]
- Lv, X.F.; Yu, J.B.; Fu, Y.Q.; Ma, B.; Qu, F.Z.; Ning, K.; Wu, H.F. A meta-analysis of the bacterial and archaeal diversity observed in wetland soils. Sci. World J. 2014, 2014, 437684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Angulo, J.; de la Cruz, M.; Chacón-Labella, J.; Illuminati, A.; Matesanz, S.; Pescador, D.S.; Pías, B.; Sánchez, A.M.; Escudero, A. The role of root community attributes in predicting soil fungal and bacterial community patterns. New Phytol. 2020, 228, 1070–1082. [Google Scholar] [CrossRef] [PubMed]
- Dee, L.E.; Cowles, J.; Isbell, F.; Pau, S.; Gaines, S.D.; Reich, P.B. When do ecosystem services depend on rare species? Trends Ecol. Evol. 2019, 34, 746–758. [Google Scholar] [CrossRef]
- Liu, L.M.; Yang, J.; Yu, Z.; Wilkinson, D.M. The biogeography of abundant and rare bacterioplankton in the lakes and reservoirs of China. ISME J. 2015, 9, 2068–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.N.; Hu, H.F.; Ma, W.H.; Deng, Y.; Wang, Q.G.; Luo, A.; Meng, J.H.; Feng, X.J.; Wang, Z.H. Relative importance of deterministic and stochastic processes on soil microbial community assembly in temperate grasslands. Microorganisms 2021, 9, 1929. [Google Scholar] [CrossRef]
- Jiao, S.; Lu, Y.H. Abundant fungi adapt to broader environmental gradients than rare fungi in agricultural fields. Glob. Change Biol. 2020, 26, 4506–4520. [Google Scholar] [CrossRef]
- Xu, M.Y.; Zhang, Q.; Xia, C.Y.; Zhong, Y.M.; Sun, G.P.; Guo, J.; Yuan, T.; Zhou, J.Z.; He, Z.L. Elevated nitrate enriches microbial functional genes for potential bioremediation of complexly contaminated sediments. ISME J. 2014, 8, 1932–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Liang, Y.T.; Zhou, S.; Zhuang, S.Y.; Sun, B. Fungal community reveals less dispersal limitation and potentially more connected network than that of bacteria in bamboo forest soils. Mol. Ecol. 2018, 27, 550–563. [Google Scholar] [CrossRef]
- Catano, C.P.; Dickson, T.L.; Myers, J.A. Dispersal and neutral sampling mediate contingent effects of disturbance on plant beta-diversity: A meta-analysis. Ecol. Lett. 2017, 20, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xue, K.; Zhou, S.T.; Wang, K.; Liu, W.J.; Xu, C.; Cui, L.Z.; Li, L.F.; Ran, Q.W.; Wang, Z.S.; et al. Environmental selection overturns the decay relationship of soil prokaryotic community over geographic distance across grassland biotas. eLife 2022, 11, e70164. [Google Scholar] [CrossRef]
- Jiao, S.; Chu, H.Y.; Zhang, B.G.; Wei, X.R.; Chen, W.M.; Wei, G.H. Linking soil fungi to bacterial community assembly in arid ecosystems. iMeta 2022, 1, e2. [Google Scholar] [CrossRef]
- Bahram, M.; Põlme, S.; Kõljalg, U.; Zarre, S.; Tedersoo, L. Regional and local patterns of ectomycorrhizal fungal diversity and community structure along an altitudinal gradient in the Hyrcanian forests of northern Iran. New Phytol. 2012, 193, 465–473. [Google Scholar] [CrossRef]
- Peay, K.G.; Garbelotto, M.; Bruns, T.D. Evidence of dispersal limitation in soil microorganisms: Isolation reduces species richness on mycorrhizal tree islands. Ecology 2010, 91, 3631–3640. [Google Scholar] [CrossRef]
- Nemergut, D.R.; Costello, E.K.; Hamady, M.; Lozupone, C.; Jiang, L.; Schmidt, S.K.; Fierer, N.; Townsend, A.R.; Cleveland, C.C.; Stanish, L.; et al. Global patterns in the biogeography of bacterial taxa. Environ. Microbiol. 2011, 13, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Tian, J.Q.; Bai, C.M.; Xiang, M.C.; Sun, J.Z.; Liu, X.Z. The biogeography of fungal communities in wetland sediments along the Changjiang River and other sites in China. ISME J. 2013, 7, 1299–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, G.H.; Luo, X.Z.; Hou, E.Q.; Zheng, M.H.; Zhang, L.L.; He, X.J.; Shen, W.J.; Wen, D.Z. Mycorrhizal fungi and phosphatase involvement in rhizosphere phosphorus transformations improves plant nutrition during subtropical forest succession. Soil Biol. Biochem. 2021, 153, 108099. [Google Scholar] [CrossRef]
- He, X.L.; Zhou, J.; Wu, Y.H.; Bing, H.J.; Sun, H.Y.; Wang, J.P. Leaching disturbed the altitudinal distribution of soil organic phosphorus in subalpine coniferous forests on Mt. Gongga, SW China. Geoderma 2018, 326, 144–155. [Google Scholar] [CrossRef]
- Zhou, J.Z.; Deng, Y.; Shen, L.N.; Wen, C.Q.; Yan, Q.Y.; Ning, D.L.; Qin, Y.J.; Xue, K.; Wu, L.Y.; He, Z.L.; et al. Temperature mediates continental-scale diversity of microbes in forest soils. Nat. Commun. 2016, 7, 12083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, S.; Lu, Y.H. Soil pH and temperature regulate assembly processes of abundant and rare bacterial communities in agricultural ecosystems. Environ. Microbiol. 2020, 22, 1052–1065. [Google Scholar] [CrossRef]
- Ni, Y.Y.; Yang, T.; Ma, Y.Y.; Zhang, K.P.; Soltis, P.S.; Soltis, D.E.; Gilbert, J.A.; Zhao, Y.P.; Fu, C.X.; Chu, H.Y. Soil pH determines bacterial distribution and assembly processes in natural mountain forests of eastern China. Glob. Ecol. Biogeogr. 2021, 30, 2164–2177. [Google Scholar] [CrossRef]
- Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Küsel, K.; Rillig, M.C.; Rivett, D.W.; Salles, J.F.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef]
- Liu, S.S.; Wang, F.; Xue, K.; Sun, B.; Zhang, Y.G.; He, Z.L.; Van Nostrand, J.D.; Zhou, J.Z.; Yang, Y.F. The interactive effects of soil transplant into colder regions and cropping on soil microbiology and biogeochemistry. Environ. Microbiol. 2015, 17, 566–576. [Google Scholar] [CrossRef]
- Yi, M.L.; Fang, Y.; Hu, G.P.; Liu, S.F.; Ni, J.R.; Liu, T. Distinct community assembly processes underlie significant spatiotemporal dynamics of abundant and rare bacterioplankton in the Yangtze River. Front. Environ. Sci. Eng. 2021, 16, 79. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Pylro, V.S.; Baldrian, P.; van Elsas, J.D.; Salles, J.F. Ecological succession reveals potential signatures of marine–terrestrial transition in salt marsh fungal communities. ISME J. 2016, 10, 1984–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, X.H.; Chen, X.L.; Huang, Z.Q.; Chen, H.Y.H. Global soil microbial biomass decreases with aridity and land-use intensification. Glob. Ecol. Biogeogr. 2021, 30, 1056–1069. [Google Scholar] [CrossRef]
- Baldock, J.A.; Skjemstad, J.O. Role of the soil matrix and minerals in protecting natural organic materials against biological attack. Org. Geochem. 2000, 31, 697–710. [Google Scholar] [CrossRef]
- Carletti, P.; Vendramin, E.; Pizzeghello, D.; Concheri, G.; Zanella, A.; Nardi, S.; Squartini, A. Soil humic compounds and microbial communities in six spruce forests as function of parent material, slope aspect and stand age. Plant Soil 2009, 315, 47–65. [Google Scholar] [CrossRef]
- Fanin, N.; Kardol, P.; Farrell, M.; Kempel, A.; Ciobanu, M.; Nilsson, M.C.; Gundale, M.J.; Wardle, D.A. Effects of plant functional group removal on structure and function of soil communities across contrasting ecosystems. Ecol. Lett. 2019, 22, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.B.; Su, F.L.; Pang, Z.Q.; Mao, Q.G.; Zhong, B.Q.; Xiong, Y.M.; Mo, J.M.; Lu, X.K. The removal of understory vegetation can rapidly alter the soil microbial community structure without altering the community assembly in a primary tropical forest. Geoderma 2023, 429, 116180. [Google Scholar] [CrossRef]
- Looby, C.I.; Treseder, K.K. Shifts in soil fungi and extracellular enzyme activity with simulated climate change in a tropical montane cloud forest. Soil Biol. Biochem. 2018, 117, 87–96. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Homage to Santa Rosalia or why are there so many kinds of animals? Am. Nat. 1959, 93, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Delgado-Baquerizo, M.; Ding, J.; Gillings, M.R.; Zhu, Y.G. Trophic level drives the host microbiome of soil invertebrates at a continental scale. Microbiome 2021, 9, 189. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, R.; Zhu, J.; Jiang, L.; Liu, L.; Gao, C.; Chen, B.; Xu, D.; Liu, J.; He, Z. Soil Phosphorus Availability Controls Deterministic and Stochastic Processes of Soil Microbial Community along an Elevational Gradient in Subtropical Forests. Forests 2023, 14, 1475. https://doi.org/10.3390/f14071475
Chen R, Zhu J, Jiang L, Liu L, Gao C, Chen B, Xu D, Liu J, He Z. Soil Phosphorus Availability Controls Deterministic and Stochastic Processes of Soil Microbial Community along an Elevational Gradient in Subtropical Forests. Forests. 2023; 14(7):1475. https://doi.org/10.3390/f14071475
Chicago/Turabian StyleChen, Rongzhi, Jing Zhu, Lan Jiang, Lan Liu, Cheng Gao, Bo Chen, Daowei Xu, Jinfu Liu, and Zhongsheng He. 2023. "Soil Phosphorus Availability Controls Deterministic and Stochastic Processes of Soil Microbial Community along an Elevational Gradient in Subtropical Forests" Forests 14, no. 7: 1475. https://doi.org/10.3390/f14071475