Abstract
This study focuses on the natural regeneration of Nothofagus glauca, a critical component of temperate forests in the Mediterranean region of the Southern Hemisphere in Chile. Various harvesting and canopy opening methods were evaluated to understand their impact on the regeneration of this species. A “mast seeding” behavior was observed in seed production, with alternating years of high and low production. Seed viability was affected by lepidopteran infestation and extreme weather events. The results indicate variable seed quantity and quality, influencing regeneration. First-year seedling mortality was significant, reaching up to 95%, and regeneration decreased annually. Additionally, changes in canopy opening and solar radiation transmission were analyzed, revealing significant post-harvest modifications. This study provides valuable insights for the sustainable management of Mediterranean forests, emphasizing the importance of considering seed production variability and regeneration dynamics in decision-making processes.
1. Introduction
Forest ecosystems play a fundamental role in biodiversity conservation and the provision of crucial ecosystem services for humanity. In particular, Southern Hemisphere Mediterranean forests harbor a rich diversity of tree species and constitute essential habitats for numerous forms of life. The Maulino Forest, part of the Mediterranean forest, stands out as a unique system transitioning between sclerophyllous shrubland and the southernmost temperate forests. It hosts a wide range of species adapted to varied environmental conditions, displaying a high degree of endemism.
The species Nothofagus glauca (Phil.) Krasser, known as hualo or maulino oak, is a key element in the Maulino Forest. Its natural distribution has significantly decreased due to human influence, leading to habitat degradation. This species plays a vital role in the structure and function of these forests, and consequently, its natural regeneration is essential for long-term persistence.
Forest regeneration is a dynamic process encompassing various stages, from flowering to seedling survival and growth, influenced by biotic and abiotic factors that vary in time and space [1,2]. Forest management should adopt an ecologically sustainable perspective, mimicking natural disturbance patterns and being compatible with plant regeneration strategies [3,4]. In particular, Nothofagus forests, as early successional species, colonize open areas after periodic disturbances of significant magnitude [5].
Silvicultural management in these forests involves successive shelterwood cuts to maintain even-aged stands [6,7,8]. However, selection cuts generating multi-aged stands are also possible [8,9]. This approach involves cutting larger trees at regular intervals, creating artificial clearings that provide optimal light conditions for regeneration, while surrounding trees offer protection against dehydration and freezing [3,4].
Despite the importance of N. glauca, scientific and practical information on sustainable regeneration and harvesting methods for this species remains insufficient. Some studies have reported the influence of canopy opening and soil moisture on seed germination [10]. However, the required degree of canopy opening for successful establishment is still unknown. Chilean forestry legislation has established guidelines for harvesting and regenerating these forests, but the effective implementation of management strategies promoting natural regeneration and biodiversity is a complex challenge. Addressing these aspects is crucial to ensure the conservation of this valuable ecosystem and its biotic components.
Nothofagus glauca is known to be shade-intolerant [11], but nursery studies have revealed that early protection against insolation is crucial for its survival [12]. Hualo plants grown in partially shaded nurseries have shown significantly higher survival rates than those exposed to full sunlight [13]. However, these results are preliminary and may be related to nursery container size.
It has been suggested that N. glauca’s regeneration strategy involves colonizing clearings within the forest after disturbances, such as tree falls, increasing light, and providing lateral protection [14]. Adult stands of this species are often relatively open, facilitating light penetration to the ground and regeneration. However, competition with other tolerant species is limited, allowing the self-replacement strategy to operate based on the generation of natural or disturbance-caused clearings.
Furthermore, the low dispersal capacity and viability of Nothofagus seeds pose additional challenges for regeneration [14]. These aspects emphasize the importance of studying the degree and type of protection necessary for the establishment of N. glauca forests, especially in areas prone to recurrent disturbances such as fires, which can have a significant impact on the biodiversity of these ecosystems.
In this context, this study aims to assess the effect of different regeneration cutting methods in N. glauca forests, evaluating the impact of varying canopy opening degrees over a 7-year period. We hypothesize that specific silvicultural interventions, aimed at altering canopy structure, will have discernible effects on seed production, seedling density, and overall forest regeneration dynamics over a 7-year observation period. Thus, the goal is to provide valuable and necessary information on the effects of these cuts on the natural regeneration of N. glauca. The results will contribute to the development of more effective and sustainable forest management strategies, as well as provide scientific and practical information supporting the conservation of this important ecosystem. Ultimately, it is expected that this work will drive informed decision making in temperate forest management and promote the conservation of biodiversity and ecological functions.
2. Materials and Methods
2.1. Study Area
The study was conducted on two properties located in the Andes Mountains (Fundo El Picazo and Cordillera property) (Figure 1).
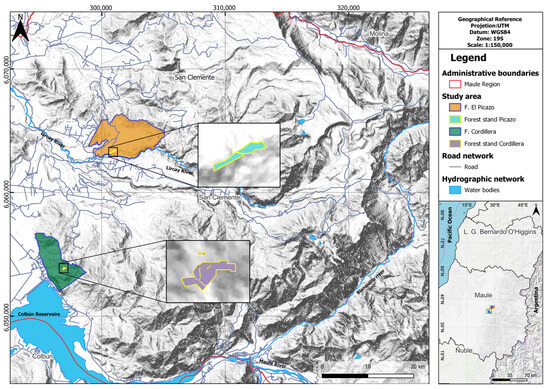
Figure 1.
Study area.
The stand at Fundo El Picazo consists of a mixed stand, composed of the species N. glauca and Nothofagus obliqua (Mirb.) Oerst. as the main species, associated with Lithraea caustica (Molina) Hook. et Arn, Quillaja saponaria Molina, Persea lingue Miers ex Bertero Nees, Cryptocarya alba (Molina) Looser, Aristotelia chilensis (Molina) Stuntz, and Azara integrifolia Ruiz & Pav. Structurally, the stand is a regularly bi-stratified stand, with N. glauca and N. obliqua occupying the upper canopy (fustal development stage), and suppressed and etiolated individuals beneath the canopy (understory and subcanopy). Secondary species are mostly present in the understory, mainly sclerophyllous species, a common situation in ecotonal areas where both forest types coexist. There is also the presence of sectors with invasive exotics, such as the regeneration of Acacia melanoxylon R.Br. and some individuals of Pinus radiata D. Don. The forest’s origin is a medium stand, an unmanaged stand with a recent age estimated at 50–60 years, showing a widespread condition of multiple stems per stump. Sanitarily, the stand exhibits a regular state for formations of these characteristics, with an abundance of etiolated individuals in the lower canopy due to the species’ intolerance. The upper canopy of Nothofagus glauca was systematically assessed using photographic material captured during various stages of the study. The evaluation focused on several key indicators of canopy health, including foliage color, density, and overall crown structure. In the images, the foliage exhibited vibrant shades of green, suggesting active photosynthesis and good chlorophyll content. The density of leaves within the upper canopy appeared consistent and robust, indicating a healthy and well-developed foliage layer. The soil, belonging to the Matacabritos series, is a sedimentary soil with a loamy-clay texture, very dark brown on the surface, clayey in depth, resting on a substrate composed of volcanic tuff with a sandy texture that forms a sandstone, with gently undulating topography, good drainage, moderate permeability, and moderate surface runoff, with effective depth ranging from 40 to 80 cm [15].
The stand at the Cordillera property also corresponds to a mixed stand, composed of the species N. glauca as the main species, associated with L. caustica, Q. saponaria, P. lingue, Laureliopsis philippiana (Looser) R. Schodde, C. alba, A. chilensis, and A. integrifolia. Structurally, the stand is a regularly bi-stratified stand, with N. glauca occupying the upper canopy (fustal development stage) and the presence of understory regeneration and suppressed and etiolated individuals beneath the canopy (understory and subcanopy). Secondary species are mostly present in the understory, mainly sclerophyllous species (C. alba being the accompanying species with the highest presence), and some hydrophilic species like P. lingue, L. philippiana, Citronella mucronata (Ruiz et Pav.) D. Don, and scarce Aextoxicon punctatum Ruiz et Pav., a common situation in ecotonal areas where both forest types coexist. The forest’s origin is a medium stand, an unmanaged stand with a recent age estimated at 50–60 years. Sanitarily, the stand exhibits a regular state for formations of these characteristics, with an abundance of etiolated individuals in the lower canopy due to the species’ intolerance. The soils, belonging to the Sierra Bellavista Association series, are moderately shallow (25 to 50 cm), without stoniness, with a moderately coarse texture, and topography featuring slopes between 30 and 50% [15].
Regarding the climate, the temperature varies between a January maximum of 29.1 °C (maximum of 29.9 °C and minimum of 27.4 °C) and a July minimum of 3.5 °C (maximum of 4.2 °C and minimum of 2.9 °C). There is an average of 211 consecutive frost-free days, with an annual average of 20 frost occurrences. The period of temperatures conducive to vegetative activity lasts for 9 months. Annually, it records an average of 1535 degree-days and 949 accumulated chill hours until 31 July. The annual average precipitation is 1137 mm, with a 5-month dry period and a water deficit of 846 mm/year. The wet period lasts for 5 months, during which there is a water surplus of 528 mm [16].
2.2. Establishment of Trials
In August 2017, the forest at both sites was harvested according to regeneration cuts considered to assess the natural regeneration of N. glauca. The evaluated harvest methods included clear-cutting, conventional shelterwood cut, shelterwood cut with light gaps, shelterwood cut with strips, and seed tree or parent tree cut. For each type of cut, the intervened area per site was 3000 m2, totaling an area of 30,000 m2 (15,000 m2 per site). In clear-cutting, all trees were harvested. The conventional shelterwood cut involved the gradual extraction of the entire stand mass in a series of partial cuts extending over part of the rotation, leaving a residual basal area of ~65–70%, aiming to maintain a homogeneous canopy cover. This residual basal area is the same as that left in the light gap cut, which is a variant of the shelterwood method where the lateral canopy effect regulates environmental conditions within the intervened area. In the strip shelterwood cut, forest strips alternate with cut strips, where the entire canopy is removed. These strips are made along contour lines, and their widths are multiples of the average canopy height. In the seed tree cut, the area is completely harvested, except for certain trees called seed trees or parent trees. Seed trees represent less than 10% of the original volume.
2.3. Seeding and Natural Regeneration
After a period of vegetative growth and with an annual frequency (from 2017 to 2023), the frequency and development of N. glauca seeding and regeneration were evaluated. For this, in each of the 3000 m2 plots described in the previous section, 9 traps of 1 m2 each were placed for seed collection. Additionally, after seeding, 9 square subplots of 1 m2 (100 cm × 100 cm) were used for each 3000 m2 plot, where the number of seedlings regenerated from seeds and vegetatively, as well as their height, was measured. A seedling is defined as an individual that has developed its first true leaves, with a practical height consideration of 5 cm. The effects of harvest methods on natural regeneration through high stand could be observed in the Cordillera property, as it was the only site where seeding occurred (in 2017). Both traps and transects were randomly located.
2.4. Statistical Analysis
Three independent variables (year, origin, and type of cut) were considered in the analysis and treated as categorical variables. The dependent variable was the quantity of seeds produced per sampling unit, expressed as the number of seeds per hectare. To determine the influence of the independent variables (year, origin, and type of cut) on N. glauca seeding, a three-way analysis of variance (ANOVA) was conducted using R Studio software Version 4.3.2. A three-way ANOVA model was applied to assess the interaction of these three variables in seed production. Multiple comparison tests (Tukey) were also conducted to identify specific differences between the levels of the independent variables. A significance level of α = 0.05 was used for all statistical tests. All necessary assumptions for conducting the statistical analysis were rigorously addressed. Normality of the dependent variable was confirmed. Additionally, other key assumptions, such as homoscedasticity and independence of observations, were carefully examined and met. These measures were taken to uphold the robustness and validity of the statistical analyses conducted in this study.
2.5. Canopy Coverage and Solar Radiation Transmission
Hemispherical photography analysis determined patterns characterizing the tree canopy and solar radiation transmission within the forest dominated by mature N. glauca individuals, before and after the application of regeneration-promoting treatments (i.e., light gaps, strips, conventional regeneration cut, seed tree, and clear-cutting). Canopy patterns correspond to canopy openness and leaf area index. Solar radiation transmission components, including direct, diffuse, and global radiation, were estimated [17].
During January 2017, hemispherical photographs were taken inside the N. glauca forest before undergoing the aforementioned cuts (Figure 2). A total of 13 hemispherical photographs were taken in the Cordillera property, and 11 hemispherical photographs were taken in the Picazo property. These photographs served as control (untreated) for statistical analysis.

Figure 2.
Hemispherical photographs taken in Nothofagus glauca forests before silvicultural interventions such as light gap creation, strip cutting, conventional regeneration cut, seed tree, and clear-cutting.
After the cuts, during the months of the vegetative growth period (between January and March from 2018 to 2023), hemispherical photographs were taken annually for six years for each treatment. In the Cordillera property, three photographs were taken per treatment in each year on plots where regeneration was monitored. In the Picazo property, one photograph was taken in the middle of the cut treatment in 2018, and in the years 2019, 2020, 2021, 2022, and 2023, hemispherical photographs were taken over the regeneration plots. Unfortunately, photographs could not be retaken in the Picazo property over the clear-cut treatment plots due to vandalism.
Hemispherical photographs were taken using a Canon EOS 60D® digital camera (Canon Inc., Tokyo, Japan) with a Sigma 4.5 mm f/2.8 EX DC® fisheye lens (Sigma GmbH, Rödermark, Germany). The camera was mounted at a height of 1.3 m above ground level, on a tripod leveled with the horizon, and with the top facing north. Photographs were taken with automatic aperture and shutter speed settings [18].
The digital images were subsequently processed and analyzed following the protocol for hemispherical photograph evaluation described in Promis et al. [17]. All images were converted to pixels with white or black colors using an automatic threshold adjustment through the SideLook 1.1.01 program (http://www.appleco.ch (accessed on 18 October 2023), Nobis [19]), which is based on an automatic edge detection method [20]. Subsequently, all images were analyzed using the HemiView version 2.1 program (Delta-T Devices, Cambridge, UK). Lens distortion correction was adjusted using specific coefficients for the lens, which were entered into the HemiView program [21]. To describe the intensity of diffuse solar radiation, the Universal Overcast Sky model (UOC) was selected. This model assumes that all regions of the sky are equally bright. Since there are no current measurements of direct and diffuse solar radiation in the forest locations, a relative proportion of 0.5 was assumed for each of them [17].
3. Results
3.1. Seed Production
In the year 2017, there was seed production in the Cordillera property, while it was absent in the El Picazo property. In both 2018 and 2019, seed production was absent in both properties. In the year 2020, seed production occurred again in the Cordillera property. In both 2021 and 2022, there was no seed production in either property, followed by a return to abundant seed production in the year 2023. These data confirm the mast seeding behavior of the species, with years of prolific seed production alternating with years of poor or no seed production, without establishing a clear pattern.
Table 1 displays some data on seed production in both properties for the years 2017, 2020, and 2023.

Table 1.
Seed production (seeds/ha) ± error in 2017, 2020, and 2023 for Cordillera and Picazo locations.
In Table 2, the ANOVA results are shown for seed production data in both properties for the years 2017, 2020, and 2023.
Table 2.
ANOVA results for seed production data in both properties for the years 2017, 2020, and 2023. *** p < 0.001; ** p < 0.01.
The results indicate that the type of harvest, the year, and the origin significantly affect the seed production of Nothofagus glauca. Furthermore, there are significant interactions among these variables, suggesting that their combinations also influence seed production. Conducting the Tukey test indicated that there is a significant difference in the number of seeds per hectare based on the origin, with significantly fewer seeds per hectare in the Picazo Property.
3.2. Natural Regeneration
In this regard, considering the amount of fallen seeds, the obtained results are presented in Table 3.
Table 3.
Average density of regeneration from seed germination on Cordillera property from 2018 to 2023.
It can be observed how first-year mortality varies depending on the harvesting method, ranging from 77% in shelterwood cuts to 95% in light gaps. Additionally, it is noted that by the second year of establishment, seedlings continue to die, with seed regeneration decreasing each year, reaching only 3.2% in the case of light gaps.
Table 4 presents the results of the ANOVA conducted on the data of regeneration density from seed germination on Cordillera property from 2018 to 2023.
Table 4.
ANOVA results for regeneration density from seed germination data on Cordillera property from 2018 to 2023. *** p < 0.001.
The results indicate that both the harvest type and the year, along with their interaction, have a significant impact on the variable of seedlings per hectare. The p-value is very close to zero in all cases, suggesting that these differences are not random and are statistically significant.
3.3. Coverage and Radiation Transmission
The Nothofagus glauca forest in the Cordillera property before the intervention had a high canopy coverage, with an average value of 23.8% canopy openings and a leaf area index of 1.4 m2 m−2 (Table 5). By 2023, the sixth summer period after the interventions, almost all harvesting activities maintain a significant modification (p < 0.05) of the canopy coverage. Clear-cutting, conventional cutting, and seed tree cutting appear to be the treatments with the lowest tree canopy coverage, between 1.4 and 1.0 times the value recorded before cutting (Table 5). The forest’s leaf area index also significantly decreased after the application of the harvests (p < 0.05), with clear-cutting reducing the leaf area index by up to 76.2% (Table 5 and Figure 3).
Table 5.
Average values of canopy opening estimation (%), leaf area index (LAI, m2 m−2), and direct (SRD, %), diffuse (SRDif, %), and global (SRG, %) solar radiation transmission during the vegetative growth period (October–March) in Nothofagus glauca forests before and after five seasons of silvicultural interventions to promote regeneration in light gaps, strips, conventional cutting, seed tree, and clear-cutting in the Cordillera property, Maule region. Hemispherical photographs were taken in January 2023. Asterisks indicate significant differences between the untreated forest situation and the silvicultural treatment (Mann–Whitney U test, * indicates significant differences with p < 0.05, ** p < 0.01).
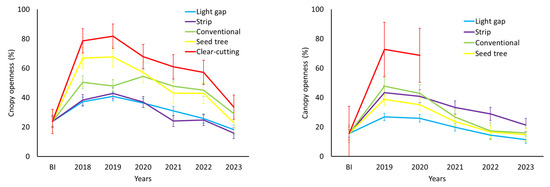
Figure 3.
Temporal change in canopy openness (%) measured through hemispherical photographs in silvicultural interventions conducted in Nothofagus glauca forests in Cordillera property (left) and Picazo property (right). (BI: Before intervention).
In the Picazo property, the canopy opening in the untreated area averages 15.6%, with a leaf area index of 1.97 m2 m−2. After six seasons of vegetative growth, strip cutting shows a statistically significant difference (p < 0.005) in canopy opening and leaf area index compared to the condition before cutting (Table 6). The other interventions show no statistical differences compared to the pre-cutting condition, indicating canopy closure over time (Table 6 and Figure 4).
Table 6.
Average values of canopy openness (%), leaf area index (LAI, m2 m−2), and direct solar radiation transmission (DSR, %), diffuse solar radiation transmission (DSR-Diff, %), and global solar radiation transmission (DSR-Global, %) during the vegetative growth period (October–March) in Nothofagus glauca forests before and after four seasons of silvicultural interventions to promote regeneration in light gaps, strips, conventional, seed tree, and clear-cutting areas in the Picazo property, Maule region. Hemispherical photographs were taken in March 2023, during the fourth vegetative growth period. No data are shown for clear-cutting due to the inability to locate the plots. Asterisks indicate significant differences between the untreated forest situation and the silvicultural treatment (Mann–Whitney U test, * denotes significant differences with p < 0.05).
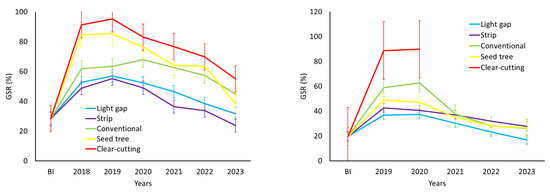
Figure 4.
Temporal change in global solar radiation transmission (%) estimated through hemispherical photographs in silvicultural interventions conducted in Nothofagus glauca forests in Cordillera property (left) and Picazo property (right). (BI: before intervention).
Changes in Solar Radiation Transmission into the Forest
In the Nothofagus glauca forest of the Cordillera property, direct, diffuse, and global solar radiation correspond to 29.1%, 27.8%, and 28.3% of what would be over the tree canopy. The solar radiation transmitted into the treatments of forest cutting remains statistically higher than that recorded inside the forest (p < 0.05). Clear-cutting and conventional cutting interventions increase solar radiation transmission by 2.2 and 1.7 times, while light gaps and strips show slightly lower increases (between 0.8 and 1.0 times) (Table 5). The trend over time in the reduction in tree canopy openings and the transmission of global solar radiation by type of silvicultural intervention can be observed in Figure 3 and Figure 4.
Solar radiation within the Nothofagus glauca forest of the Picazo property averages 18.9%, 20.3%, and 19.7% of the components of direct, diffuse, and global solar radiation received over the tree canopy, respectively (Table 6). Solar radiation transmissions, both direct and global, are statistically similar between the harvests and the pre-intervention situation (p > 0.05). Therefore, in these interventions, spontaneous revegetation inside the forest, with the establishment of the regeneration of tree species and an increase in understory coverage, tends to reduce solar radiation transmission to the forest floor (Table 6). The trend over time in the reduction in tree canopy openings and the transmission of global solar radiation by type of silvicultural intervention can be observed in Figure 3 and Figure 4.
4. Discussion
The natural regeneration of Nothofagus glauca is of great importance for the conservation and sustainable management of temperate forests in the Mediterranean region of Chile. This study focused on evaluating the effect of different regeneration harvest methods and canopy openings on the natural regeneration of N. glauca. Through the assessment of seeding, natural regeneration, and solar radiation transmission, valuable information is provided for sustainable forest management in this region.
It can be observed that the quantity of seeds varies, but the most notable aspect is that almost all collected seeds were perforated by a lepidopteran, rendering them non-viable for the natural regeneration of the species through high forests. Many aborted fruits were also found, possibly as a result of high temperatures due to apparent climate change. The variability in seeding observed in the Cordillera and El Picazo properties over the years is consistent with the mast seeding of Nothofagus glauca, a common pattern in many Nothofagus species [22]. This mast seeding is reflected in the alternation of years with high and low seed production, which can have a significant impact on natural regeneration and tree cohort composition [23]. The lack of seeding in certain years may be related to various factors, including seed viability, insect predation, and climatic conditions, especially high temperatures associated with climate change manifested in extreme events such as heatwaves [24].
Considering that in recent years there has been no seeding (except in 2023) and that in seed tree, conventional shelterwood cut, and strip shelterwood cut treatments, the number of plants per hectare increased in 2021, this would indicate that there is also understorey regeneration, probably from the root. However, in the 2022–2023 season, the number of plants decreased again. The regeneration was less than the fallen seeds, which was expected, and may be due to various factors, such as seed viability, predation, and microclimatic conditions, among others.
The results of natural regeneration show that in the Cordillera property, seedling mortality is high during the first year after seed fall, especially in conventional shelterwood cut and light gap treatments. These results are consistent with the idea that protection against direct solar radiation is crucial for the survival of N. glauca seedlings [12]. The high mortality of seedlings in light gap treatments may be related to excessive exposure to sunlight, which can desiccate and damage seedlings [10].
The significant reduction in canopy cover and leaf area index after harvests, especially in the Cordillera property, confirms that management treatments succeeded in opening the canopy and increasing solar radiation transmission. This is crucial to provide optimal light conditions for N. glauca regeneration [25]. However, clear-cutting had the most drastic effect on reducing canopy cover and leaf area index, which could pose a challenge to forest biodiversity and ecosystem services associated with tree cover [26].
The long-term trend in solar radiation transmission shows a gradual decrease in canopy opening and solar radiation transmission to the forest floor in most treatments. This indicates a possible recovery of tree canopy cover and competition for light between regeneration and adult trees. Resource competition is a critical factor that can influence the success of natural regeneration.
The adaptive capacity of Nothofagus glauca is inherently linked to its genetic diversity, a crucial factor influencing the species’ ability to respond to environmental changes. While our study has focused on short-term regeneration dynamics and the impact of silvicultural interventions, it is pertinent to consider the genetic aspects shaping the observed patterns. Genetic diversity plays a pivotal role in the adaptation and resilience of tree populations [27,28]. Variability in genetic makeup can influence traits such as seed viability, resistance to pests, and tolerance to climatic variations [29]. In the case of Nothofagus glauca, understanding the genetic factors affecting regeneration is essential for predicting the species’ long-term sustainability. Future research endeavors could delve deeper into the genetic composition of Nothofagus glauca populations in different regions, exploring how distinct genetic traits may contribute to variations in seed production, seedling survival, and overall forest dynamics [30]. Long-term studies incorporating genetic analyses would provide valuable insights into the adaptive potential of Nothofagus glauca under changing environmental conditions.
In examining the regeneration patterns of Nothofagus glauca, it is crucial to consider potential confounding factors that might have influenced the observed dynamics. While our study primarily focused on silvicultural interventions and short-term regeneration outcomes, acknowledging and exploring additional variables contributes to a more nuanced interpretation of the results. One significant factor to consider is the influence of microclimatic variations on seed germination and seedling establishment. Microclimatic conditions, such as temperature and humidity, can play a pivotal role in determining the success of natural regeneration [31,32]. Additionally, soil characteristics, including nutrient composition and structure, may affect seedling growth and survival [33]. Furthermore, the presence of specific mycorrhizal associations has been documented to impact the establishment of tree seedlings [34]. Mycorrhizal symbiosis can influence nutrient uptake and overall plant health, thereby affecting regeneration success. In addition to these ecological factors, anthropogenic influences, such as invasive species or land-use history, could act as confounding variables. Human-induced alterations to the landscape may have repercussions on seed dispersal mechanisms and the competitive dynamics among different species within the ecosystem [35].
5. Conclusions
The conclusions drawn from this study shed light on the complex dynamics of natural regeneration of Nothofagus glauca in the Mediterranean region of Chile. The evidence of mast seeding in seed production highlights the importance of considering annual variability when designing forest management strategies. The presence of seeds perforated by lepidopterans and aborted fruits suggests potential negative impacts of climate change, as evidenced during extreme temperature events such as heatwaves.
Silvicultural interventions, particularly clear-cutting, have demonstrated a substantial impact on forest structure, significantly reducing tree cover and leaf area index. This result poses significant challenges for forest biodiversity and underscores the need to balance canopy opening with the conservation of key ecosystem elements.
Regarding natural regeneration, there is high seedling mortality, especially in conventional shelterwood cut and light gap treatments. The importance of protection against direct solar radiation for seedling survival highlights the need for specific considerations in the planning of silvicultural interventions.
In the long term, a gradual decrease in canopy opening and solar radiation transmission is foreseen, indicating a potential recovery of tree canopy cover and an intensification of competition for light. This phenomenon underscores the importance of competition as a critical factor that can influence the success of natural regeneration.
In terms of sustainable management, the conclusions point to the need for continuous monitoring of natural regeneration and forest structure. This adaptive approach will allow for adjusting management practices in response to changing ecosystem dynamics, ensuring the long-term preservation of Nothofagus glauca and its ecological functions in the Mediterranean region of Chile. These conclusions not only contribute to fundamental scientific knowledge but also provide valuable guidance for forestry professionals in making informed and sustainable decisions.
Author Contributions
Conceptualization, R.S.-M. and A.M.C.-A.; methodology, R.S.-M., Á.P. and A.M.C.-A.; software, R.S.-M., Á.P. and A.M.C.-A.; validation, R.S.-M., Á.P. and A.M.C.-A.; formal analysis, R.S.-M., Á.P. and A.M.C.-A.; investigation, R.S.-M., Á.P., M.P.-A. and A.M.C.-A.; resources, R.S.-M.; data curation, R.S.-M., Á.P., M.P.-A. and A.M.C.-A.; writing—original draft preparation, R.S.-M., Á.P. and A.M.C.-A.; writing—review and editing, R.S.-M., Á.P., M.P.-A. and A.M.C.-A.; visualization, R.S.-M., Á.P. and A.M.C.-A.; supervision, R.S.-M.; project administration, R.S.-M.; funding acquisition, R.S.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fondo de Investigación del Bosque Nativo (project number: 008/2014).
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Pulido, F.J.; Díaz, M. Regeneration of a Mediterranean oak: A whole-cycle approach. Écoscience 2005, 12, 92–102. [Google Scholar] [CrossRef]
- Martínez Pastur, G.J.; Lencinas, M.; Peri, P.; Cellini, J. Flowering and seeding patterns in unmanaged and managed Nothofagus pumilio forests with a silvicultural variable retention system. Forstarchiv 2008, 79, 60–65. [Google Scholar]
- Aplet, G.H.; Johnson, N.C.; Olson, J.T.; Sample, V.A. Defining Sustainable Forestry; Island Press: Washington, DC, USA, 1993; p. 328. [Google Scholar]
- Smith, D.; Larson, B.; Kelty, M.; Ashton, P. The Practice of Silviculture: Applied Forest Ecology; John Wiley & Sons: New York, NY, USA, 1997. [Google Scholar]
- Veblen, T.T. Nothofagus regeneration in treefall gaps in northern Patagonia. Can. J. For. Res. 1989, 19, 265–371. [Google Scholar] [CrossRef]
- Grosse, H.; Quiroz, I. Silvicultura de los bosques de segundo crecimiento de roble, raulí y coigüe en la región centro-sur de Chile. In Silvicultura de los Bosques Nativos de Chile; Donoso, C., Lara, A., Eds.; Editorial Universitaria: Santiago, Chile, 1998; pp. 95–128. [Google Scholar]
- Lara, A.; Donoso, C.; Donoso, P.; Nuñez, P.; Cavieres, A. Normas de manejo para raleo de renovales del tipo forestal roble-raulí-coigüe. In Silvicultura de los Bosques Nativos de Chile; Donoso, C., Lara, A., Eds.; Editorial Universitaria: Santiago, Chile, 1998; pp. 129–144. [Google Scholar]
- Martínez-Velásquez, A. Silvicultura práctica en renovales puros y mixtos, y bosques remanentes originales del tipo forestal roble-raulí-coigüe. In Silvicultura de los Bosques Nativos de Chile; Donoso, C., Lara, A., Eds.; Editorial Universitaria: Santiago, Chile, 1998; pp. 145–175. [Google Scholar]
- Chauchard, L.; Forgues, A.A.; Fernández, M. Plan de Manejo Cerro Tren; Asentamiento Universitario San Martín de los Andes, Universidad Nacional del Comahue: San Martín de los Andes, Argentina, 1994; p. 21. [Google Scholar]
- Burgos, A.; Grez, A.A.; Bustamante, R.O. Seed production, pre-dispersal seed predation and germination of Nothofagus glauca (Nothofagaceae) in a temperate fragmented forest in Chile. For. Ecol. Manag. 2008, 255, 1226–1233. [Google Scholar] [CrossRef]
- Donoso, C. Ecología Forestal, el Bosque y su Ambiente; Editorial Universitaria: Santiago, Chile; University Austral de Chile: Santiago, Chile, 1981; p. 369. [Google Scholar]
- Santelices, R.; Herrera, L.; Osores, J. Cultivo en vivero del hualo (Nothofagus glauca (Phil.) Krasser) bajo diferentes gradientes de luminosidad y espaciamiento. Cienc. For. 1995, 10, 3–13. [Google Scholar]
- Quiroz, I.; Pincheira, M.; Hernández, A. Restauración ecológica en la zona central de Chile: Técnicas Silvícolas para la Supervivencia y Crecimiento de Ruil, Hualo, y Peumo bajo Condiciones de Estrés Hídrico. Chile For. 2009, 363, 50–53. [Google Scholar]
- Donoso, C. Bosques Templados de Chile y Argentina. Variación, Estructura y Dinámica; Editorial Universitaria: Santiago, Chile, 1993; p. 484. [Google Scholar]
- Centro de Información de Recursos Naturales (CIREN). Estudio Agrologico VII Region. Erosión de Suelos. Descripciones de Suelos, Materiales y Símbolos; Publicación CIREN: Santiago, Chile, 1997; Volume 2023. [Google Scholar]
- Santibáñez, F. Atlas Agroclimático de Chile. Estado Actual y Tendencias del Clima. Tomo III: Regiones de Valparaíso, Metropolitana, O’Higgins y Maule; Disponible: Santiago, Chile, 2017. [Google Scholar]
- Promis, A.; Gärtner, S.; Butler Manning, D.; Durán Rangel, C.; Reif, A.; Cruz, G.; Hernández, L. Comparison of four different programs for the analysis of hemispherical photographs using parameters of canopy structure and solar radiation transmittance. Wald. Landschaftsforschung Naturschutz 2011, 11, 19–33. [Google Scholar]
- Inoue, A.; Yamamoto, K.; Mizoue, N.; Kawahara, Y. Effects of image quality, size and camera type on forest light environment estimates using digital hemispherical photography. Agr. Forest Meteorol. 2004, 126, 89–97. [Google Scholar] [CrossRef]
- Nobis, M. SideLook 1.1. Imaging Software for the Analysis of Vegetation Structure with True-Colour Photographs. Available online: http://www.appleco.ch (accessed on 18 October 2023).
- Nobis, M.; Hunziker, U. Automatic thresholding for hemispherical canopy-photographs based on edge detection. Agric. For. Meteorol. 2005, 128, 243–250. [Google Scholar] [CrossRef]
- Webb, N. User Manual for the Self Levelling Mount Type SLM8; Delta-T Devices Ltd.: Cambridge, UK, 2009; p. 20. [Google Scholar]
- Santelices Moya, R.; Vergara, R.; Cabrera Ariza, A.; Espinoza Meza, S.; Silva Flores, P. Variación intra-específica en Nothofagus glauca una especie endémica de los bosques mediterráneos de Chile %J Bosque (Valdivia). Bosque 2020, 41, 221–231. [Google Scholar] [CrossRef]
- Monks, A.; Kelly, D. Testing the resource-matching hypothesis in the mast seeding tree Nothofagus truncata (Fagaceae). Austral. Ecol. 2006, 31, 366–375. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Ashton, M.; Kelty, M. The Practice of Silviculture: Applied Forest Ecology, 10th ed.; Wiley: Hoboken, NJ, USA, 2018. [Google Scholar]
- Lindenmayer, D.B.; Laurance, W.F.; Franklin, J.F. Global Decline in Large Old Trees. Science 2012, 338, 1305–1306. [Google Scholar] [CrossRef] [PubMed]
- Leimu-Brown, R.; Fischer, M. A Meta-Analysis of Local Adaptation in Plants. PLoS ONE 2008, 3, e4010. [Google Scholar] [CrossRef]
- Allendorf, F.; Luikart, G. Conservation and Genetics of Populations; Blackwell Publishing: Hoboken, NJ, USA, 2006. [Google Scholar]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Petit, R.; Hampe, A. Some Evolutionary Consequences of Being a Tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef]
- Clark, J.S.; Bell, D.M.; Kwit, M.C.; Zhu, K. Competition-interaction landscapes for the joint response of forests to climate change. Glob. Change Biol. 2014, 20, 1979–1991. [Google Scholar] [CrossRef] [PubMed]
- LePage, P.T.; Canham, C.D.; Coates, K.D.; Bartemucci, P. Seed abundance versus substrate limitation of seedling recruitment in northern temperate forests of British Columbia. Can. J. For. Res. 2000, 30, 415–427. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L. The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. J. Ecol. 2009, 97, 1202–1214. [Google Scholar] [CrossRef]
- Teste, F.P.; Simard, S.W.; Durall, D.M.; Guy, R.D.; Jones, M.D.; Schoonmaker, A.L. Access to mycorrhizal networks and roots of trees: Importance for seedling survival and resource transfer. Ecology 2009, 90, 2808–2822. [Google Scholar] [CrossRef] [PubMed]
- Vellend, M.; Baeten, L.; Myers-Smith, I.H.; Elmendorf, S.C.; Beauséjour, R.; Brown, C.D.; De Frenne, P.; Verheyen, K.; Wipf, S. Global meta-analysis reveals no net change in local-scale plant biodiversity over time. Proc. Natl. Acad. Sci. USA 2013, 110, 19456–19459. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).