
Effects of Forest Fires on Boreal Permafrost and Soil Microorganisms: A Review

1
Key Laboratory of Sustainable Forest Ecosystem Management of Ministry of Education, College of Forestry, Northeast Forestry University, Harbin 150040, China
2
State Key Laboratory of Frozen Soil Engineering, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
*
Author to whom correspondence should be addressed.
Forests 2024, 15(3), 501; https://doi.org/10.3390/f15030501
Submission received: 1 February 2024
/
Revised: 5 March 2024
/
Accepted: 5 March 2024
/
Published: 8 March 2024
(This article belongs to the Section Natural Hazards and Risk Management)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The frequency of forest fires has increased dramatically due to climate change. The occurrence of forest fires affects the carbon and nitrogen cycles and react to climate change to form a positive feedback mechanism. These effects further impact the distribution of microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) and the soil microbial community structure. In addition, permafrost degradation can significantly affect the microorganisms in the soil. Based on these findings, this review examines the effects of fire intensity and post-fire recovery time on permafrost, the soil microbial community, MBC, MBN, and their interrelationships. This review demonstrated that (1) fires alter the condition of surface vegetation, reduce the organic layer thickness, redistribute snow, accelerate permafrost degradation, and even lead to permanent changes, where the restoration of the pre-fire state would require several decades or even centuries; (2) soil microbial community structure, soil MBC, and MBN negatively correlate with fire intensity, and the effects become more pronounced with increasing fire intensity; and (3) the structural diversity and stability of the soil microbial community were improved with time, and the amount of MBC and MBN increases as the years after a fire go by; it would still take more than ten years to recover to the pre-fire level. However, the relationship between permafrost degradation and soil microbes after forest fires is still unclear due to a lack of quantitative research on the mechanisms underlying the changes in soil microorganisms resulting from fire-induced permafrost degradation. Therefore, expanding quantitative studies and analyses of the mechanisms of interactions between forest fires, permafrost, and soil microorganisms can provide a scientific basis for understanding ecosystem carbon pools and dual-carbon targets in Arctic–boreal permafrost regions.
1. Introduction
Forest fires destroy millions of hectares of forest annually and play a significant role in energy transport and nutrient cycling [1,2]. Fires affect forest ecosystems in two ways: high-intensity fire disturbances severely harm the equilibrium of forest ecosystems and cause the degradation of forest ecosystems; low-intensity fires help prevent major fires, preserve biodiversity, and promote natural regeneration [3,4,5]. Natural disasters such as fires are susceptible to meteorological factors [6]. The greenhouse effect has become increasingly severe in recent years due to the unusually dry climate and frequent extreme weather, leading to a longer duration of fires and a significant increase in their frequency and intensity [7].
Permafrost is a layer of soil that has remained frozen for more than two years [8]. Permafrost is widely distributed in the northern hemisphere, covering approximately 19 × 106 km2 or 15%~25% of the land surface [9], primarily in Russia, Canada, China, North America, and Mongolia (Figure 1) [10,11]. It is a crucial component of the cryosphere and significantly affects how the ecological environment in cold climates evolves over time. The area covered by boreal permafrost has drastically decreased in recent years, from 23.25 × 106 km2 in 1969~1973 to 21.64 × 106 km2 in 2014~2018, with the highest degradation rate occurring in Siberia, followed by Alaska, the Mongolian Plateau, northern Canada, and Greenland [12]. The cryosphere is the most sensitive to global warming, and alterations in the permafrost-induced natural environment directly affect the survival and development of humans [13]. Permafrost stores a large amount of organic carbon [14,15,16], which makes up approximately 60% of the terrestrial carbon on the planet and twice as much as in the atmosphere [17]. However, the Arctic has warmed significantly faster than the rest of the Earth in recent decades, giving rise to a phenomenon known as the Arctic amplification effect [18]. The average annual air temperature in the Arctic and boreal (subarctic) regions has increased by 0.31~0.44 °C/10 a between 1990 and 2021 [19], leading to rapid permafrost degradation [20] and a massive release of carbon in soil ecosystems, causing changes in soil functions [21].
Soil microorganisms are one of the most active components of soil [22], contributing significantly to the carbon and nitrogen cycles [23]. The structure and composition of soil microbial communities are crucial to the functioning of soil ecosystems because they serve as important indicators of soil quality [24] while directly affecting soil ecological functions and biochemical cycles [25]. In addition, they also contribute to the stability and sustainability of the soil ecosystem and can strengthen the buffer capacity to withstand the deterioration of the soil microecological environment [26]. Microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) are the total amounts of carbon and nitrogen, respectively, in living and dead organisms with volumes less than 5000 μm3 in the soil [27]. They are an indicator of changes in soil quality [28,29,30] and act as a link between the exchange of matter and energy in different spheres [31].
The dynamics of disturbances caused by fire are a response to climate change [32]. Climate warming, an increase in the number of days with high temperatures and droughts, a decrease in precipitation and relative humidity, an increase in wind speed, and a dry microclimate in forests are conducive to the occurrence and spread of fires, increasing the frequency and intensity of fires [18,33]. Forest fires disrupt the dynamic balance between permafrost and soil microorganisms in the boreal permafrost region [34,35], contributing to the ongoing effects of the permafrost environment. They increase the soil temperature and active layer thickness (ALT) [36] and cause rapid permafrost degradation [37,38], which alters the soil moisture content and respiration rates, affecting soil microbial activity [35]. Increased soil temperature rises the microbial respiration index [39], boosts microbial activity, and accelerates growth [40] causing MBC and MBN to accumulate in the soil [41]. MBC and MBN are prime factors that influence the composition of the soil microbial community [24]. Permafrost thawing caused by forest fires can alter the soil microbial community structure and its relative abundance [42]. These alterations can also affect the available nitrogen status and dissolved organic matter (DOM) content in forest soils. For example, research has shown that, whereas most bacteria have a significant positive correlation with DOM, nearly all fungi have a negative correlation with DOM [43].
The effects of fire disturbance on soil habitat and microorganisms have gained attention recently as a new area of study [44]. Current research on the effects of forest fires on soil microorganisms in permafrost has focused primarily on the upper soil horizons and their changing conditions. However, the mechanism underlying the influence of forest fires on the soil microbial community structure, MBC, and MBN in permafrost regions is unclear. To this end, this review attempts to study the effects of forest fires on soil microorganisms in permafrost, focusing on the following aspects: (1) changes in soil microbial community structure, MBC, and MBN content in boreal permafrost regions; (2) effects on permafrost characteristics; (3) effects of different fire intensities on soil microbial community structure, MBC, and MBN content in boreal permafrost regions; and (4) effects of different post-fire recovery times on soil microbial community structure, MBC, and MBN content in boreal permafrost regions. The study of soil MBC and MBN in permafrost is beneficial for improving the quantitative study and mechanistic analysis of wildfires, permafrost, and soil microorganisms in boreal permafrost regions and provides a theoretical basis for maintaining the carbon and nitrogen balance in permafrost areas and soil ecosystems. It can also provide a scientific basis as a global carbon pool and a two-carbon target.
2. Variations in Soil Microbial Community Structure, Carbon, and Nitrogen in Boreal Permafrost Regions
Permafrost in the northern hemisphere is rapidly deteriorating due to global warming [45]. Permafrost degradation alters ALT, soil temperature, moisture, ecosystem parameters (water balance, soil carbon content, carbon bioavailability, etc.), and soil microbial activities [46,47], which affect the soil microbial community structure, MBC, and MBN [48,49].
2.1. Changes in the Soil Microbial Community Structure
The primary cause of the alteration in the structure of the soil microbial community is the thawing of permafrost caused by climate change. Permafrost degradation changes several environmental parameters, including soil temperature, moisture, enzyme activity, pH, and species composition [45,50]. It also impacts how resources are distributed among soil microorganisms [51], ultimately affecting the structure and diversity of soil microbial communities [52,53,54]. With changes in habitat, the response pattern of bacterial alpha diversity (α-diversity) to permafrost degradation changes. The α–diversity of bacterial communities in forest ecosystems increased as the permafrost degraded in the Da Xing’anling Mountains of Northeast China, but it decreased in wetland ecosystems [55]. However, in the grassland and meadow ecosystems of the central and northern Tibetan Plateau, the α-diversity of bacteria and fungi decreased when the permafrost degraded [56]. Unlike bacteria, fungi are more resistant to permafrost degradation [57]. A study conducted in boreal permafrost regions of the Qinghai–Tibet Plateau revealed that, although permafrost degradation did not change the dominant community of soil microorganisms at depths of 0~20 cm, it significantly affected the bacterial community structure, with different soil layers exhibiting similar bacterial structures [58]. However, the structure of fungal communities in different soil layers did not change significantly between the undegraded and degraded permafrost [58]. Permafrost degradation on the Tibetan Plateau increased the abundance of Actinomycetes (from 6.2% to 17.2%) and decreased the abundance of Proteus (from 32.2% to 22.9%). The Parvarchaeota decreased from 5.7% to 0.3% during permafrost degradation, while the dominant fungi remained unchanged [59]. This suggests that the degradation of permafrost affects bacterial, fungal, and archaeal community structures. In the boreal permafrost regions of the Da Xing’anling Mountains, there were significant differences in the soil microbial community structure among the three forest types (Larix gmelinii, Pinus sylvestris, and Betula platyphylla). The types of phospholipid fatty acids in the soil were identical among the three forest types, but Betula platyphylla had the highest Shannon–Wiener diversity index (3.090), while Pinus sylvestris had the lowest (2.927). Additionally, the Larix gmelinii forest had a significantly greater Simpson dominance index (0.040) and McIntosh evenness index (6.297) than did the other two forests [60]. This indicates that the difference in forest type has a significant impact on the community composition and diversity of soil microorganisms. The structure and diversity of the soil microbial community varied with soil depth. For example, soil microbial diversity in the boreal permafrost regions of northwestern Canada peaked at 0–15 cm depth and decreased with increasing soil depth [61]. These results are consistent with those from the Siberian boreal permafrost regions [62]. Permafrost degradation has significantly increased the diversity of soil bacteria in the boreal permafrost regions of Alaska [63]. Overall, permafrost degradation causes changes in the soil microbial community structure and diversity. However, in permafrost areas, different ecosystems (forest, meadow, and wetland) vary somewhat; the soil microbial community structure and diversity index vary among different forest types. At present, the research on the change characteristics of the soil microbial community structure in permafrost has focused more on a soil depth of 0–30 cm, while research on the survival depth of soil microorganisms in permafrost region is less relevant. This requires the study of the structural characteristics of the microbial community in deep soil under permafrost regions.
2.2. Changes in Soil Microbial Carbon and Nitrogen Characteristics
Soil MBC and MBN are the most active components of soil organic matter; these components are highly sensitive to environmental changes [64] and can reflect alterations in the soil microbial community and function [24]. Both climate change and fire-induced permafrost degradation reduce soil MBC and MBN contents [65,66,67]. Permafrost degradation affects ground ice, groundwater recharge sources and recharge, runoff pathways and discharge processes, and the exchange of groundwater and surface water [68], thus jeopardizing the ecological balance and stability of these regions [69]. Studies have shown that the levels of MBC and MBN are greater in stable permafrost on the Qinghai–Tibet Plateau than in unstable boreal permafrost regions [70]. At depths of 0–50 cm, the amounts of MBC and MBN were 0.014~0.620 g/kg and 0.644~12.770 mg/kg, respectively, in the stable boreal permafrost regions and 0.019~0.410 g/kg and 0.207~3.725 mg/kg, respectively, in the extremely unstable boreal permafrost regions. This indicates that the stability of permafrost has some effect on the MBC and MBN contents. At present, research on the microbial carbon and nitrogen contents of permafrost soil has focused more on short-term changes. With global climate change, permafrost has degraded, and environmental factors have changed, affecting soil microbial carbon and nitrogen. Therefore, it is necessary to strengthen long-term research on the carbon and nitrogen of permafrost soil.
3. Impact of Forest Fires on Permafrost
Fires alter soil temperature, moisture content, and ALT [71]. The effects are long-term [72] and require a lengthy recovery period to return to pre-fire levels [73]. In addition, permafrost degradation can also lead to various geological hazards and landscape changes [74,75].
3.1. Altered Characteristics Due to Forest Fire-Induced Permafrost Degradation
The first manifestation of permafrost degradation is a rising ground temperature and a deeper active layer. When the soil temperature rises, the permafrost thaws and becomes thinner [76,77]. Aboveground vegetation can lower the temperature of the soil surface by blocking direct sunlight, warming the soil, and reducing transpiration [78,79]. By delaying the thawing of permafrost and active layers and directly influencing the hydrothermal processes of permafrost, it safeguards the formation and conservation of permafrost [80,81]. It has been found that climate warming increases the intensity of fire disturbance, thereby exacerbating the severity of soil burns [82,83]. Fire disturbance affects the thickness of the soil organic layer. For example, in the Da Xing’anling Mountains in Northeast China, the soil organic layer decreased by 6.9 cm under low-intensity fire interference and 8.6 cm under high-intensity fire interference [84]. A reduction in the thickness of the organic layer can directly affect soil temperature by increasing the efficiency with which heat is transferred through the soil. In Alaska, permafrost studies revealed that a 12 cm decrease in the organic layer after fire disturbance increased the soil temperature by 2.6 °C in the top 20 cm of soil [85]. In a short period of time, the soil organic layer decreases, and the soil ALT can increase by 300%–600% [86]. As the organic layer recovers, so do the ALT and soil temperature [87]. Changes in available space and resources in permafrost may further affect the rate and magnitude of forest regeneration after fires due to increased fire disturbance intensity and organic layer depth [88]. Fires eliminate vegetation and organic layers, reducing their ability to shade and insulate the ground. As a result, more heat is transferred directly to the ground, thus increasing the soil temperature, thickening the active layer, and thawing the permafrost [89,90,91]. The understory vegetation coverage also decreased as the fire severity increased [92]. The permafrost ecosystem affected by fire may not return to its pre-fire state for at least ten years [73].
Variations in fire severity distinctly affect the temperature, moisture content, and ALT in permafrost soil. The high-severity fire disturbance areas in the boreal permafrost regions of the Da Xing’anling Mountains in Northeast China had higher soil temperatures than did the low-severity fire disturbance areas, but the soil moisture content was lower [93]. Eight years after a severe burn in the boreal permafrost regions of interior Alaska, the annual mean ground surface temperature increased by 2.7 °C, and the mean annual ground temperature (MAGT) increased by 2.1 °C at a depth of 1 m [94,95]. In the boreal permafrost regions of the Da Xing’anling Mountains in Northeast China, the MAGT at a depth of 1 m increased by 2.3 °C seven years after the severe burn and only by 0.1 °C after the light burn [96]. This is primarily because vegetation is burned more severely under high-intensity fire disturbance than under low-intensity fire disturbance. The thinner surface litter and organic matter layer and the partial or complete elimination of the insulation layer during high-intensity fire disturbances are additional contributing factors [71]. Thus, high-intensity fire disturbance areas have higher soil temperatures and soil moisture contents than low-intensity fire disturbance areas. The ALT trended upward in tandem with the increasing severity of the fire [97]. For example, three years after the Alaskan fire, the ALT was 60 cm in an unburned area, 59–64 cm in a lightly burned area, and increased from 99 cm to 179 cm in a moderately burned area [98]. Similarly, 13 years after the fire in the Da Xing’anling Mountains in Northeast China, the average ALT in severely burned areas was 102.63 cm, whereas it was only 61.93 cm in unburned areas [99]. In general, fire intensity and fire recovery time have different effects on permafrost, and the impact of forest fires on permafrost is long-term. Therefore, it is necessary to combine long-term and short-term research on permafrost to provide a theoretical basis for research on the stability of permafrost ecosystems.
3.2. Impact of Forest Fires on Permafrost Landscapes
Freeze–thaw disasters in permafrost areas are unique geological disasters caused by changes in the thermal–mechanical stability of rock and soil masses during the permafrost freezing and thawing process [100]. Forest fires can readily thaw and destroy permafrost structures. The interface between the active and frozen layers creates a sliding surface, and the thawed soil slowly glides downslope under its weight to form a mudflow [100]. Forest fires result in the increasing the amount of latent heat that can enter the ground, increasing soil temperatures, melting ground ice, triggering permafrost degradation, and affecting permafrost landscapes [20] via surface subsidence [75,101] and the formation of thermokarst ponds and lakes [74,102,103]. Forty percent of the permafrost in the northern hemisphere is affected by thermokarst subsidence, and new thermokarst landforms are constantly forming [104]. Due to permafrost degradation on the Tibetan Plateau, the total number of thermokarst lakes and ponds increased by 534, and the total area increase by 4.1 × 106 m2 from 1969 to 2010 [105]. Some studies in the boreal permafrost regions of Canada have shown that wildfires directly led to the development of thermokarst marshes of 2200 ± 1500 km2, accounting for 25% of all thermokarst marsh expansion during the same period. Even the impact of forest fires on permafrost peatlands can last for 30 years [36,94,106]. In the northern boreal permafrost regions of Alaska, due to forest fire disturbances and thermokarst landslides, thermokarst landforms, which are basically stable after five years, formed [107].
3.3. Effect of Fire Intensity on the Soil Microbial Community Structure
The structure of the soil microbial community is significantly correlated with soil pH, temperature, and moisture content [73,108,109]. Variations in soil temperature can potentially impact the structure and functions of soil microbial communities [109,110]. Given that soil microorganisms are heat-sensitive and that the ground surface temperature can increase from 50 °C to 300 °C after forest fires, it is inevitable that soil microorganisms will be killed or injured by forest fires, which will decrease their population [111] and make it more difficult for them to recover quickly [112]. Fire alters the aboveground vegetation and physicochemical properties of soil [113]. The slow infiltration of fire-generated ash also modifies the physicochemical properties of soil, which in turn influence soil microorganisms and alter their biomass and community structure [114].
Bacteria and fungi are the two main microbial groups in forest soil ecosystems [115,116]. Varying fire severities have different effects on the soil microbial community and diversity [117]. The molar mass concentration, relative abundance of phospholipid fatty acids, and total PLFA mass in the soil microbial community decrease with increasing fire intensity, while the relative abundance of actinomycetes and fungi gradually increase [118]. After high-severity fires in the boreal permafrost regions of Northeast China, the diversity of soil microbial communities declined and, 11 years later, the community structure resembled that of unburned areas [73]. High-severity fires in the boreal permafrost regions of Canada weakened the stability of the soil fungal and bacterial community structures, especially the diversity and richness of fungi, which declined as the fire intensity increased [119]. Due to the intensity of the fire, the structure and relative abundance and diversity indices of the soil fungal and bacterial microbial communities at different soil depths also changed accordingly [118]. For example, in the Da Xing’anling Mountains of Northeast China, the coefficient of the ratio of fungi to bacteria at a depth of 0~5 cm was lower in the severely burned area than in the lightly burned area, and the opposite was true at a depth of 5~10 cm [120]. In summary, the effects of forest fires on soil microorganisms in the boreal permafrost regions increased with increasing fire intensity. Forest fires decrease the structural stability of fungal and bacterial communities in soil. With increasing fire intensity, the diversity index of soil microbial communities decreased, but the relative abundance of actinomycetes and fungi increased, and it took more than ten years to recover to the pre-fire state. While forest fires affect soil ecosystems mainly by restricting microbial activity, where bacteria and fungi have high functional diversity and are usually distributed in phylogenetically distinct groups [121], we know little about the ecological roles of most taxa. Therefore, it is important to summarize the potential effects of fire on microbial communities.
3.4. Effect of Years Post-Fire on the Dynamics of the Soil Microbial Community Structure
Common microbial communities in permafrost soils include Proteobacteria, Firmicutes, Acidobacteria, Chloroflexi, Actinobacteria, and Bacteroidetes [109]. One year after the fire, the abundance of Firmicutes, Betaproteobacteria, Actinobacteria, and Proteobacteria increased in northern Canada, while the relative abundance of Betaproteobacteria decreased three years later [111]. Nine years after the fire in Alaska, the abundance of Firmicutes and Actinobacteria decreased [14]. Overall, the relative abundance of microorganisms in the soil after a fire first increases and then decreases with the increase in years post-fire [108,109].
Soil pH is one of the primary determinants of soil microbial community structure [55,117]. The pH increases at the onset of forest fires [14] and is consistently higher than the pre-fire value in the short term, returning to or even being lower than the pre-fire value nine years later [122,123]. In boreal forests, the pH and fungal diversity of the soil after a fire initially increase and then progressively decrease [124]. A higher soil pH is more conducive to bacterial development [106], and usually increases Actinobacteria abundance and decreases Acidobacteria abundance [125]. The increase in soil pH after a forest fire reduces the diversity of soil microbial communities [75]. In addition, it would take a minimum of 11 years for the composition and diversity of the soil microbial community to revert to their pre-fire levels [73]. Studies have shown that fires are more lethal to soil fungi than to bacteria [73,97]. Thus, we can infer that soil microorganisms progressively recover in the years following the fire. In addition, there were significant differences in the microbial communities in the soil at various time points post-fire. The relative abundance of microorganisms in permafrost soil increased in the short term in response to fire interference (e.g., Firmicutes, β Proteobacteria, and Actinobacteria) and decreased with recovery time. It took approximately 11 years to return to the pre-fire disturbance community structure and diversity [73].
3.5. Effects of Forest Fires on the Soil Microbial Community Structure in the Boreal Permafrost Regions
The structure of the soil microbial community is significantly correlated with soil pH, temperature, and moisture content [76,111]. Variations in soil temperature can potentially impact the structure and functions of soil microbial communities [112,113]. Some studies on the Canadian permafrost found that the relative abundance of Acidimicrobiia and Holophagae, Thermoleophilia, Bacilli, and Blastocatellia increased under the influence of fire interference compared with those in unburned samples, but the relative abundance of Solibacteres, Phycisphaerae, and Opitutae decreased [111,112]. Fire alters the aboveground vegetation and physicochemical properties of soil [116]. The slow infiltration of fire-generated ash also modifies the physicochemical properties of soil, which in turn influences soil microorganisms and alters their biomass and community structure [117].
4. Effects of Forest Fire on MBC and MBN in the Boreal Permafrost Regions
Forest fires can cause soil temperature to increase [39], ALT to increase, and permafrost to degrade rapidly [40,41]. These factors can damage the permafrost environment, affect soil microbial activities, and disrupt the dynamic balance between permafrost and soil microorganisms [38], resulting in changes in the composition and quantity of the soil microbial community structure and affecting the soil MBC and MBN content [44]. MBC and MBN are the primary components affecting the soil microbial community structure [30].
4.1. Effect of Fire Severity on Soil MBC and MBN
Fire severity can alter factors such as the permafrost environment and soil organic matter (SOM) content [126,127], the SOM content decreases with increasing fire intensity and exacerbates the degradation of permafrost [75], affecting the survival and development of soil microorganisms [75]. The levels of MBC and MBN in the soil decrease with increasing fire severity [128,129]. For example, in Alaska, the contents of MBC and MBN were highest in unburned areas, followed by lightly burned areas, and lowest in severely burned areas [130]. MBC and MBN levels progressively decrease with increasing depth [131,132], indicating a close relationship between depth and biomass content. Forest fires can increase soil temperature and cause permafrost degradation, thus increasing the activity space of soil microorganisms [128,131]. This helps boost the soil microbial population, alter the microbial community structure, and affect the MBC and MBN contents [24]. SOM is an energy source for soil microorganisms [133] that can improve the physical environment of soil, increase porosity, and improve permeability to provide an optimal environment for the activities of soil microorganisms [134]. On the Hebei Plain in China, MBC and MBN decreased after different fire intensities and with increasing soil depth. At depths of 0–10 cm, the MBC content did not change significantly after light or moderate burn injury but decreased significantly after severe burn injury. At a depth of 10–30 cm, the MBC and MBN contents were the highest at the unburned site, followed by the lightly burned and severely burned site, and were the lowest at the moderately burned site [135].
4.2. Effect of Years Post-Fire on MBC and MBN
The effects of forest fires on MBC and MBN gradually weakened and recovered for a long time after the fire. In the permafrost region of North America, compared with those in unburned plots, MBC and MBN levels decreased (3.8%–12.7%) one year after mild fire disturbance [136]. This is because permafrost is resilient and can recover to a certain extent after a fire disturbance, but it is a long process, and the recovery time is different due to the fire intensity, climatic conditions, and geographical conditions [137]. However, it is becoming more difficult for permafrost to return to its pre-fire state because of ongoing climate change and forest fire disturbance. The severity of the fire will slow the response ability of soil microorganisms. Permafrost degradation after a fire will change the living environment of soil microorganisms. This will impact soil microbial survival and development, which in turn will alter the levels of MBC and MBN [138]. A study that simulated MBC and MBN in boreal forests revealed that 17.2 years after a severe burn, 3.8 years after a moderate burn, and 0.05 years after a light burn, the impacts on MBC and MBN changed from negative to positive [128]. In the above study, the MBC and MBN decreased under the influence of mild fire disturbance compared with those in unburned plots. The greater the fire interference intensity was, the greater the threshold for MBC and MBN content which changed from a decreasing trend to an increasing trend.
5. Discussion
In this review, we showed that fire intensity has an impact on the physical and chemical properties of permafrost soil. Compared with the unburned site in the permafrost region, the soil temperature increases more after high-intensity fires [94,95,96] than after low intensity fires. With increasing fire intensity, the thickness of the active layer increases [98], and the soil moisture content decreases [93]. Forest fires cause permafrost degradation, destroy permafrost structures, and cause the formation of geological landscapes, such as surface settlements, thaw depressions, and thermokarst lakes [74,102,103]. Through research and investigations of the physical and chemical properties of permafrost, we can understand the specific impact of forest fire disturbance on permafrost and provide a scientific basis for forest and wetland protection and ecological environment restoration in cold regions. With increasing fire intensity, the diversity index of the soil microbial community structure decreases, and the molar mass concentration and relative abundance of PLFAs in the soil microbial communities decreases, but the relative abundance of actinomycetes and fungi gradually increases [121]. However, some studies have shown that the microbial community structure of Canadian permafrost soil, such as the relative abundance of Firmicutes, Proteobacteria, and Actinomyces [123], increased one year after fire disturbance, but the relative abundance of Proteobacteria and the numbers of Firmicutes and Actinomycetes decreased 3 years after the fire [14]. This indicates that the soil microbial community structure is not only affected by the fire intensity, but also that the recovery time is crucial for soil microbial community structure. This may be because forest fires reduce the stability of soil microorganisms [139], which use genetic variation to adapt to external interference and maintain the relative stability of community composition and structure to improve the resistance and resilience of soil microbial systems [140,141]. The carbon and nitrogen contents in the permafrost area decrease [128,129] with increasing fire intensity, and the carbon and nitrogen contents decrease with increasing soil depth [132]. When we study the microbial carbon and nitrogen content after forest fires, soil depth should be considered comprehensively.
6. Prospects
Even though there have been some studies on the relationships among wildfires, permafrost, and soil microorganisms, many research findings are still very tentative and even contradict each other. Moreover, there are few studies on the effects of forest fires on MBC, MBN, and soil microorganisms in boreal permafrost regions. Therefore, future research should focus on the following areas:
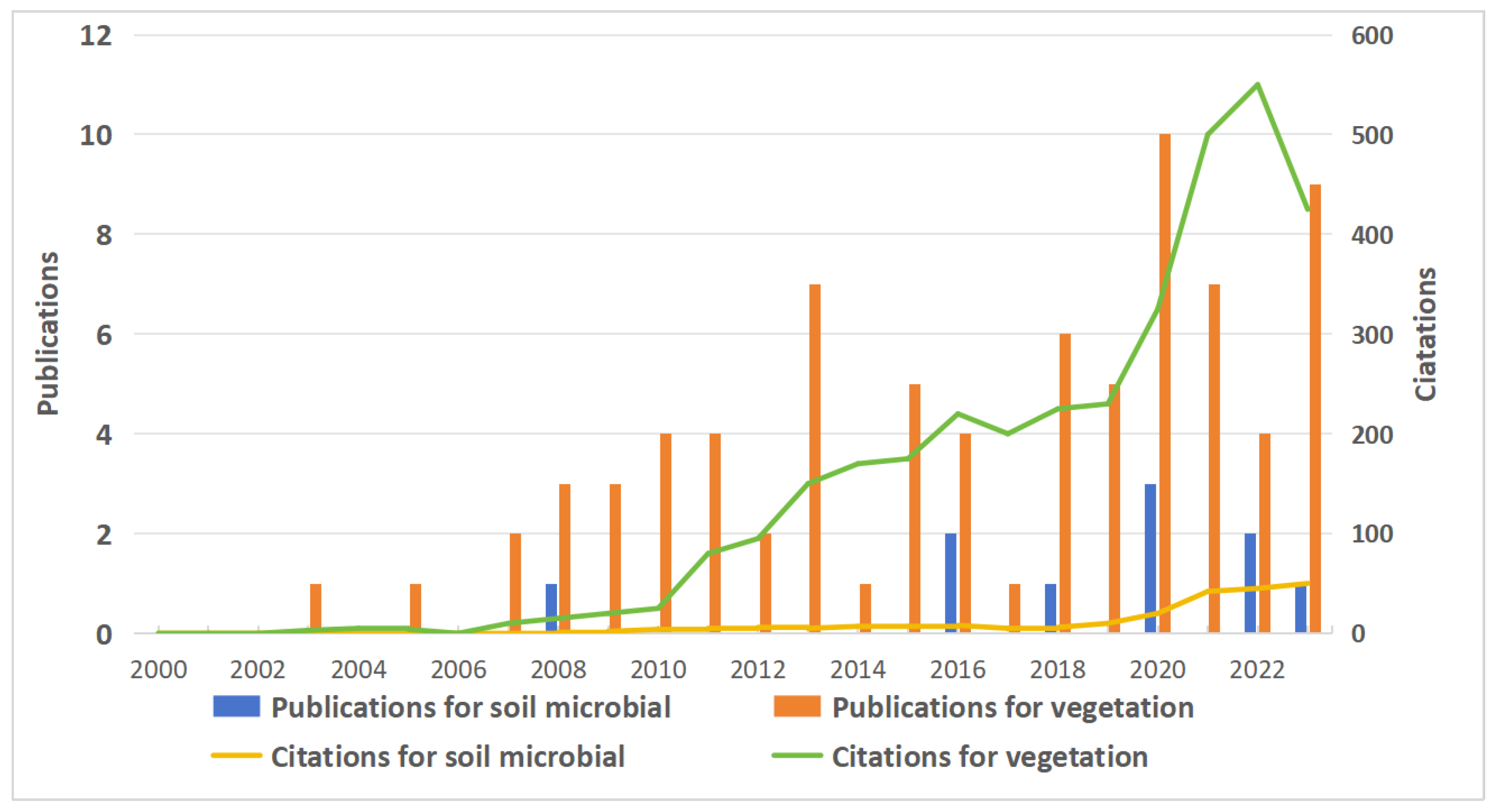
The combined short- and long-term monitoring of the soil microbial community structure, MBC, and MBN content after fires in boreal permafrost regions should be improved. Soil microbial community structure is a key determinant of soil quality [23] that influences soil ecological function and biochemical cycling and is critical for soil ecosystems [25]. MBC and MBN are important conversion links of soil carbon and nitrogen pools and serve as a storehouse for soil nutrients [27]. At present, research on the impact of forest fires and induced permafrost degradation on soil microbial changes is limited and short-term (1–20 years) (Figure 2). However, the effects of forest fires on soil microbial properties are long-term [73,112], with some studies confirming that even 25 years after a fire, the soil microbial community structure and MBC and MBN contents are still significantly different from those in unburned areas [14]. But the long-term effects (>30 years) research is not yet available. And more research was focused on seasonally frozen ground region (Figure 2). Thus, to precisely assess the varying characteristics and recovery trends of soil microorganisms following fires in boreal permafrost regions, it is imperative to develop combined long- and short-term monitoring programs.
Extending the scope of studies on permafrost degradation and soil microbial properties following fire disturbance is crucial. Currently, more research has focused on the variation in soil microorganisms after different fire severities and various times post-fire. However, quantitative and process-revealing studies on the effects of fire severity, post-fire duration, and fire frequency on soil microorganisms in boreal permafrost regions are lacking; in particular, the effect of fire frequency on soil microorganisms is still unclear. Studies have shown that the frequency of fire may affect the physicochemical properties of the soil, such as the soil temperature, soil moisture content, soil pH, and SOM [139,142]. A single forest fire may promote the formation of soil nutrients in the short-term, but frequent, long-term forest fires may reduce or even eliminate soil nutrients [143]. These findings are consistent with the intermediate disturbance hypothesis [144]. An increase in fire frequency negatively affects the extent of permafrost recovery and the ability of forests to store carbon and nitrogen [145]. However, most research on fire frequency in boreal permafrost regions has focused on aboveground vegetation, with only a few studies on soil microorganisms (Figure 3). Moreover, there is a dearth of research on the mechanisms controlling the effects of MBC and MBN in boreal permafrost regions, with a greater emphasis on non-permafrost areas. Therefore, quantitative research and the mechanistic analysis of forest fires and their rapid permafrost degradation, as well as the soil microbial community structure, MBC, and MBN are urgently needed. This study can provide a scientific basis and data support for soil ecosystem research and carbon reserves in Arctic–boreal permafrost regions.
7. Conclusions
The frequency of forest fires is increasing in tandem with the severity of global warming. Forest fires induce rapid permafrost degradation and affect soil microbial activity, which modifies the soil microbial community structure, MBC, and MBN content. In this paper, we reviewed previous studies on forest fires, permafrost, and soil microorganisms and drew the following conclusions.
Forest fires burn vegetation in the permafrost region, reducing the thickness of the organic layer and surface albedo, causing snow cover redistribution, allowing more latent heat to reach the ground, accelerating permafrost degradation, and even causing irreversible changes. Restoring the state prior to the fire would require decades or possibly hundreds of years.
The diversity of the soil microbial community structure decreased with increasing fire intensity and was greatest after a light burn; however, the proportion coefficient of fungi and bacteria in the soil microbial community structure decreased after moderate and severe burns. As the severity of the fire increased, so did the molar mass concentration of phospholipid fatty acids in the soil microbial community and the corresponding richness, dominance, and evenness indices.
The contents of MBC and MBN in the permafrost region decreased as the fire severity increased, and recovered as the amount of time after the fire increased. Time and fire severity interact to influence the recovery of MBC and MBN, with MBC and MBN recovering more quickly following a light burn than following a severe burn.
As the recovery time after the fire increase and the contents of MBC and MBN increase, the diversity and stability of the soil microbial community structure gradually improves. However, it would take more than ten years to return to the pre-fire level of community structure.
Author Contributions
Conceptualization, J.L. (Jing Liu), T.X., Y.H. and Y.S.; methodology, J.L. (Jing Liu), J.L. (Jingtao Li) and K.C.; writing—original draft preparation, J.L. (Jing Liu) and X.L.; writing—review and editing, J.L. (Jing Liu) and X.L.; supervision, X.L.; project administration, X.L.; funding acquisition, X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been supported by the National Natural Science Foundation of China (Grant Nos. 32241032 and 42001052), the Heilongjiang Excellent Youth Fund (Grant No. YQ2022D002), the State Key Laboratory of Frozen Soils Engineering Open Fund (Grant No. SKLFSE202008), and the Fundamental Research Fund for the Central Universities (Grant Nos. 2572023CT01 and 2572021BA03).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hu, H.; Luo, S.; Luo, B. Effects of forest fire disturbance on soil organic carbon in forest ecosystems: A review. Acta Ecol. Sin. 2020, 40, 1839–1850. (In Chinese) [Google Scholar]
- Knorre, A.A.; Siegwolf, R.T.; Kirdyanov, A.V.; Saurer, M.; Churakova, O.V.; Prokushkin, A.S. Fire as a major factor in dynamics of tree-growth and stable δ13C and δ18O variations in larch in the permafrost zone. Forests 2022, 13, 725. [Google Scholar] [CrossRef]
- Wu, Z.; Li, M.; Wang, B.; Tian, Y.; Quan, Y.; Liu, J. Analysis of factors related to forest fires in different forest ecosystems in China. Forests 2022, 13, 1021. [Google Scholar] [CrossRef]
- Liang, S.; Hurteau, M.D. Novel climate–fire–vegetation interactions and their influence on forest ecosystems in the western USA. Funct. Ecol. 2023, 37, 2126–2142. [Google Scholar] [CrossRef]
- Zylstra, P.; Wardell-Johnson, G.; Falster, D.; Howe, M.; McQuoid, N.; Neville, S. Mechanisms by which growth and succession limit the impact of fire in a south-western Australian forested ecosystem. Funct. Ecol. 2023, 37, 1350–1365. [Google Scholar] [CrossRef]
- Zhao, P.; Liu, L. Prediction of forest fire danger rating based on meteorological and space-time factors. J. For. Res. 2018, 3, 102–110. [Google Scholar]
- Hu, Y.; Zhao, F.; Chen, F.; Chen, F.; Shu, L. Impacts of global warming and large-scale climate fluctuation on forest fires and forest carbon emissions. Terr. Ecosyst. Conserv. 2021, 1, 75–81. (In Chinese) [Google Scholar]
- Subcommittee, P. Glossary of permafrost and related ground-ice terms. Assoc. Comm. Geotech. Res. NRC 1988, 156, 63–64. [Google Scholar]
- Obu, J. How much of the earth’s surface is underlain by permafrost? J. Geophys. Res. 2021, 126, e2021JF006123. [Google Scholar] [CrossRef]
- Zhang, T.; Heginbottom, J.A.; Barry, R.G.; Brown, J. Further statistics on the distribution of permafrost and ground ice in the Northern Hemisphere. Polar. Geogr. 2000, 24, 126–131. [Google Scholar] [CrossRef]
- Ran, Y.; Li, X.; Cheng, G.; Che, J.; Aalto, J.; Karjalainen, O.; Hjort, J.; Luoto, M.; Jin, H.; Obu, J.; et al. New high-resolution estimates of the permafrost thermal state and hydrothermal conditions over the Northern Hemisphere. Earth Syst. Sci. Data Discuss. 2021, 14, 865–884. [Google Scholar] [CrossRef]
- Li, G.; Zhang, M.; Pei, W.; Melnikov, A.; Khristoforov, I.; Li, R.; Yu, F. Changes in permafrost extent and active layer thickness in the Northern Hemisphere from 1969 to 2018. Sci. Total Environ. 2021, 804, 150182. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Liu, M.; Chen, H.; Jiang, X.; Zang, S. Effects of snow cover change on soil microbial community structure in permafrost region of Great Hing’an Mountains. Acta Ecol. Sin. 2020, 40, 789–799. (In Chinese) [Google Scholar]
- Taş, N.; Prestat, E.; McFarland, J.W.; Wickland, K.P.; Knight, R.; Berhe, A.A.; Jorgenson, T.; Waldrop, M.P.; Jansson, J.K. Impact of fire on active layer and permafrost microbial communities and metagenomes in an upland Alaskan boreal forest. ISME J. 2014, 8, 1904–1919. [Google Scholar] [CrossRef] [PubMed]
- McGuire, A.D.; Koven, C.; Lawrence, D.M.; Clein, J.S.; Xia, J.; Beer, C.; Burke, E.; Chen, G.; Chen, X.; Delire, C.; et al. Variability in the sensitivity among model simulations of permafrost and carbon dynamics in the permafrost region between 1960 and 2009. Glob. Biogeochem. Cycles 2016, 30, 1015–1037. [Google Scholar] [CrossRef]
- Xu, Q.; Xiao, C.; Feng, Y.; Du, Z.; Wang, L.; Wei, Z. Advances in microbe mediated key processes of the carbon cycle in thermokarst lakes. Adv. Earth Sci. 2023, 38, 470–482. (In Chinese) [Google Scholar]
- Ni, J.; Wu, T.; Zhao, L. Carbon cycle in circum-Arctic permafrost regions: Progress and prospects. J. Glaciol. Geocryol. 2019, 41, 845–857. (In Chinese) [Google Scholar]
- Rantanen, M.; Karpechko, A.Y.; Lipponen, A.; Nordling, K.; Hyvärinen, O.; Ruosteenoja, K.; Vihma, T.; Laaksonen, A. The Arctic has warmed nearly four times faster than the globe since 1979. Commun. Earth Environ. 2022, 3, 168–178. [Google Scholar] [CrossRef]
- Li, W.; Yan, D.; Weng, B.; Zhu, L. Research progress on hydrological effects of permafrost degradation in the Northern Hemisphere. Geoderma 2023, 438, 116629. [Google Scholar] [CrossRef]
- Biskaborn, B.K.; Smith, S.L.; Noetzli, J.; Matthes, H.; Vieira, G.; Streletskiy, D.A.; Schoeneich, P.; Romanovsky, V.E.; Lewkowicz, A.G.; Abramov, A.; et al. Permafrost is warming at a global scale. Nat. Commun. 2019, 10, 264–275. [Google Scholar] [CrossRef]
- Wang, G.; Chen, L.; Zhang, D.; Chang, R. Divergent trajectory of soil autotrophic and heterotrophic respiration upon permafrost thaw. Environ. Sci. Technol. 2022, 56, 10483–10493. [Google Scholar] [CrossRef]
- Zhao, M. Analysis situation in the study of elevational patterns of soil microbial diversity. J. Green Sci. Technol 2020, 23–25. (In Chinese) [Google Scholar]
- Qu, Z.; Liu, B.; Ma, Y.; Sun, H. Differences in bacterial community structure and potential functions among eucalyptus plantations with different ages and species of trees. Appl. Soil. Ecol. 2020, 149, 103515. [Google Scholar] [CrossRef]
- Tang, Y.; Wei, Z.; Yan, T.; Yang, L.; Ci, E. Biological indicator of soil quality: A review. Soils 2017, 39, 157–163. (In Chinese) [Google Scholar]
- Yang, Y.-L.; Ma, X.-S.; Xie, H.-T.; Bao, X.-L.; Liang, C.; Zhu, X.-F.; He, H.-B.; Zhang, X.-D. Effects of conservation tillage on soil microbial community and the function of soil carbon cycling. Chin. J. Appl. Ecol. 2021, 32, 2675–2684. (In Chinese) [Google Scholar]
- Fang, Y.; Wang, W.; Yao, X.; Peng, X. Soil microbial community composition and environmental controls in northern temperate steppe of China. Acta Sci. Nat. PKU 2017, 53, 142–150. (In Chinese) [Google Scholar]
- Li, R.-B. Review of Research Advances of soil microbial biomass carbon. Forest Environ Sci. 2008, 24, 65–69. (In Chinese) [Google Scholar]
- Mazzarino, M.J.; Szott, L.; Jimenez, M. Dynamics of soil total C and N, microbial biomass, and water-soluble C in tropical agroecosystems. Soil Biol. Biochem. 1993, 25, 205–214. [Google Scholar] [CrossRef]
- Mabuhay, J.A.; Nakagoshi, N.; Isagi, Y. Influence of erosion on soil microbial biomass, abundance and community diversity. Land Degrad. Dev. 2004, 15, 183–195. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, Y. Research progress of forest soil microbial metrology. J. Guizhou Norm. Univ. (Nat. Sci.) 2020, 38, 118–122. (In Chinese) [Google Scholar]
- Shi, B.D.; Han, D.D.; Li, Z.G.; Quan, X.K.; Yu, H.Z.; Di, X.Y.; Yang, G. Characteristics of soil microbial biomass carbon and nitrogen in burned area in Larix gmelinii forest. J. Northeast Forest Univ. 2022, 50, 78–82+98. (In Chinese) [Google Scholar]
- Hu, H.Q.; Wei, S.J.; Wei, S.; Sun, L. Effect of fire disturbance on forest ecosystem carbon cycle under the background of climate warming. J. Catastrophol. 2012, 27, 37–41. (In Chinese) [Google Scholar]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlström, A.; Canadell, J.G.; Cong, N.; et al. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nat. Commun. 2014, 5, 5018–5025. [Google Scholar] [CrossRef]
- Song, X.; Wang, G.; Ran, F.; Chang, R.; Song, C.; Xiao, Y. Effects of topography and fire on soil CO2 and CH4 flux in boreal forest underlain by permafrost in northeast China. Ecol. Eng. 2017, 106, 35–43. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, J.F.; Liu, J.P.; Du, H. A review of main studies on permafrost degradation in northeast China. Sci. Technol. Eng. 2022, 22, 13151–13161. (In Chinese) [Google Scholar]
- Gibson, C.M.; Chasmer, L.E.; Thompson, D.K.; Quinton, W.L.; Flannigan, M.D.; Olefeldt, D. Wildfire as a major driver of recent permafrost thaw in boreal peatlands. Nat. Commun. 2018, 9, 3041. [Google Scholar] [CrossRef]
- Smith, S.L.; Romanovsky, V.E.; Lewkowicz, A.G.; Burn, C.R.; Allard, M.; Clow, G.D.; Yoshikawa, K.; Throop, J. Thermal state of permafrost in North America: A contribution to the international polar year. Permafrost Periglac. 2010, 21, 117–135. [Google Scholar] [CrossRef]
- O’neill, H.B.; Burn, C.R.; Allard, M.; Arenson, L.U.; Bunn, M.I.; Connon, R.F.; Kokelj, S.V.; LeBlanc, A.-M.; Morse, P.D.; Smith, S.L. Permafrost thaw and northern development. Nat. Clim. Chang. 2020, 10, 722–723. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Scott, J.J.; Saltonstall, K.; Broders, K.; Montero-Sanchez, M.; Püspök, J.; Bååth, E.; Meir, P. Microbial diversity declines in warmed tropical soil and respiration rise exceed predictions as communities adapt. Nat. Microbiol. 2022, 7, 1650–1660. [Google Scholar] [CrossRef]
- Zheng, Y.; Shi, C.; Wu, F.; Liu, H.; Zhao, Z.; Hu, C. Research progress on influencing factors of soil microbial activity. Chin. J. Soil Sci. 2009, 40, 1209–1214. (In Chinese) [Google Scholar]
- Wang, D.; Zang, S.; Wang, L.; Ma, D.; Li, M. Effects of permafrost degradation on soil carbon and nitrogen cycling in permafrost wetlands. Front. Earth Sci. 2022, 10, 911314. [Google Scholar] [CrossRef]
- Monteux, S.; Weedon, J.T.; Blume-Werry, G.; Gavazov, K.; Jassey, V.E.; Johansson, M.; Keuper, F.; Olid, C.; Dorrepaal, E. Long-term in situ permafrost thaw effects on bacterial communities and potential aerobic respiration. ISME J. 2018, 12, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Heal, K.; Wang, S.; Cao, S.; Zhou, C. Chemodiversity of soil dissolved organic matter and its association with soil microbial communities along a chronosequence of Chinese fir monoculture plantations. Front. Microbiol. 2021, 12, 729344. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; Chen, K.; Li, H.; Wei, J.; Zhao, P.; Zhou, M. Research progress on the effects of forest fire disturbance on soil carbon release in high-latitude permafrost regions. Ecol. Environ. Sci. 2022, 31, 1278–1284. (In Chinese) [Google Scholar]
- Tian, J.; He, N.; Hale, L.; Niu, S.; Yu, G.; Liu, Y.; Blagodatskaya, E.; Kuzyakov, Y.; Gao, Q.; Zhou, J. Soil organic matter availability and climate drive latitudinal patterns in bacterial diversity from tropical to cold temperate forests. Funct. Ecol. 2018, 32, 61–70. [Google Scholar] [CrossRef]
- Lin, L.; Zhu, B.; Chen, C.; Zhang, Z.; Wang, Q.; He, J. Precipitation overrides warming in mediating soil nitrogen pools in an alpine grassland ecosystem on the Tibetan Plateau. Sci. Rep. 2016, 6, 31438. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, S.; Pu, Y.; Xu, X.; Li, Y. Distribution characteristics and impact factors of soil microbial biomass carbon, nitrogen and phosphorus in western Sichuan plain. Chin. J. Eco-Agric. 2019, 27, 1607–1616. (In Chinese) [Google Scholar]
- Euskirchen, E.S.; Edgar, C.W.; Turetsky, M.R.; Waldrop, M.P.; Harden, J.W. Differential response of carbon fluxes to climate in three peatland ecosystems that vary in the presence and stability of permafrost. J. Geophys. Res. Biogeosci. 2014, 119, 1576–1595. [Google Scholar] [CrossRef]
- Schädel, C.; Schuur, E.A.G.; Bracho, R.; Elberling, B.; Knoblauch, C.; Lee, H.; Luo, Y.; Shaver, G.R.; Turetsky, M.R. Circumpolar assessment of permafrost C quality and its vulnerability over time using long-term incubation data. Glob. Chang. Biol. 2014, 20, 641–652. [Google Scholar] [CrossRef]
- Ganie, M.A.; Mukhtar, M.; Dar, M.A.; Ramzan, S. Soil microbiological activity and carbon dynamics in the current climate change scenarios: A Review. Pedosphere 2016, 26, 577–591. [Google Scholar]
- Liang, Y.; Jiang, Y.; Wang, F.; Wen, C.; Deng, Y.; Xue, K.; Qin, Y.; Yang, Y.; Wu, L.; Zhou, J.; et al. Long-term soil transplant simulating climate change with latitude significantly alters microbial temporal turnover. ISME J. 2015, 9, 2561–2572. [Google Scholar] [CrossRef]
- Mitzscherling, J.; Horn, F.; Winterfeld, M.; Mahler, L.; Kallmeyer, J.; Overduin, P.P.; Schirrmeister, L.; Winkel, M.; Grigoriev, M.N.; Wagner, D.; et al. Microbial community composition and abundance after millennia of submarine permafrost warming. Biogeosciences 2019, 16, 3941–3958. [Google Scholar] [CrossRef]
- Liu, F.; Kou, D.; Chen, Y.; Xue, K.; Ernakovich, J.G.; Chen, L.; Yang, G.; Yang, Y. Altered microbial structure and function after thermokarst formation. Global Change Biol. 2021, 27, 823–835. [Google Scholar] [CrossRef]
- Huang, Y.; Ye, T.; Du, Y.; Cong, R.; Yang, L.; He, S.; Wang, X. Research progress on frozen soil and its microorganisms. J. Temp. For. Res. 2021, 4, 13–18. (In Chinese) [Google Scholar]
- Dong, X.; Man, H.; Liu, C.; Wu, X.; Zhu, J.; Zheng, Z.; Ma, D.; Li, M.; Zang, S. Changes in soil bacterial community along a gradient of permafrost degradation in Northeast China. Catena 2023, 222, 106870. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, F.; Kang, L.; Zhang, D.; Kou, D.; Mao, C.; Qin, S.; Zhang, Q.; Yang, Y. Large-scale evidence for microbial response and associated carbon release after permafrost thaw. Glob. Chang. Biol. 2021, 24, 3218–3229. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ma, D.; Jiang, X.; Liu, M.; Zang, S. Effects of seasonal freeze-thaw on soil microbial community structures and extracellular enzyme activities in Zhalong wetland. Acta Sci. Circumst. 2020, 40, 1443–1451. (In Chinese) [Google Scholar]
- Zhang, N.; Chen, K.; Wang, H.; Yang, Y. Changes of microbial community characteristics in freezing-thaw degradation of alpine wetland. Ecol. Sci. 2022, 41, 20–28. (In Chinese) [Google Scholar]
- Wu, M.-H.; Chen, S.-Y.; Chen, J.-W.; Xue, K.; Chen, S.-L.; Wang, X.-M.; Chen, T.; Kang, S.-C.; Rui, J.-P.; Thies, J.E.; et al. Reduced microbial stability in the active layer is associated with carbon loss under alpine permafrost degradation. Proc. Nat. Acad. Sci. USA 2021, 118, e2025321118. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Ma, D.; Zang, S.; Wu, X. Characteristics of soil microbial community structure under different forest types of permafrost regions in the Greater Khingan Mountains. J. Glaciol. Geocryol. 2018, 40, 1028–1036. (In Chinese) [Google Scholar]
- Frank-Fahle, B.A.; Yergeau, É.; Greer, C.W.; Lantuit, H.; Wagner, D. Microbial functional potential and community composition in permafrost-affected soils of the NW Canadian Arctic. PLoS ONE 2014, 9, e84761. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Kobabe, S.; Liebner, S. Bacterial community structure and carbon turnover in permafrost-affected soils of the Lena Delta, northeastern Siberia. Can. J. Microbiol. 2009, 55, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Kong, W.; Liang, C.; Zhou, T.; Jia, H.; Dong, X. Permafrost thawing exhibits a greater influence on bacterial richness and community structure than permafrost age in Arctic permafrost soils. Cryosphere 2020, 14, 3907–3916. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Y.; Shi, R.; Han, S.; Li, H.; Xu, H. Temporal variations of soil microbial biomass and enzyme activities during the secondary succession of primary broadleaved-pinuskoraiensis forests in Changbai Mountains of Northeast China. Chin. J. Appl. Ecol. 2013, 24, 366–372. (In Chinese) [Google Scholar]
- Chen, S.; Zhao, Q.; Liu, W.; Zhang, Z.; Li, S.; Li, H.; Nie, Z.; Zhou, L.; Kang, S. Effects of freeze-thaw cycles on soil N2O concentration and flux in the permafrost regions of the Qinghai-Tibetan Plateau. Sci. Cold Arid. Reg. 2018, 10, 69–79. [Google Scholar]
- Wang, J.; Han, Y.; Song, C.; Xi, F. Effects of freezing-thawing cycles on soil organic carbon mineralization in the peatland ecosystems from continuous permafrost zone, Great Hinggan Mountains. Clim. Chang. Res. 2018, 14, 59–66. (In Chinese) [Google Scholar]
- Chen, L.; Gao, Y. Global climate change effects on soil microbial biomass stoichiometry in alpine ecosystems. Land 2022, 11, 1661. [Google Scholar] [CrossRef]
- Cheng, G.; Zhao, L.; Li, R.; Wu, X.; Sheng, Y.; Hu, G.; Zou, D.; Jin, H.; Li, X.; Wu, Q. Characteristics, changes and impacts of permafrost on the Tibetan Plateau. Chin. Sci. Bull. 2019, 64, 2783–2795. (In Chinese) [Google Scholar]
- Oliva, M.; Pereira, P.; Antoniades, D. The environmental consequences of permafrost degradation in a changing climate. Sci. Total Environ. 2017, 616, 435–437. [Google Scholar] [CrossRef]
- Wu, M.; Qu, D.; Li, T.; Liu, F.; Gao, Y.; Chen, Y.; Chen, T. Effects of permafrost degradation on soil microbial biomass carbon and nitrogen in the Shule River Headwaters, the Qilian Mountains. Sci. Geogr. Sin. 2021, 41, 177–186. (In Chinese) [Google Scholar]
- Lucash, M.S.; Marshall, A.M.; Weiss, S.A.; McNabb, J.W.; Nicolsky, D.J.; Flerchinger, G.N.; Link, T.E.; Vogel, J.G.; Scheller, R.M.; Abramoff, R.Z.; et al. Burning trees in frozen soil: Simulating fire, vegetation, soil, and hydrology in the boreal forests of Alaska. Ecol. Model. 2023, 481, 110367. [Google Scholar] [CrossRef]
- Han, A.; Qing, S.; Bao, Y.; Na, L.; Bao, Y.; Liu, X.; Zhang, J.; Wang, C. Short-term effects of fire severity on vegetation based on Sentinel-2 satellite ddata. Sustainability 2021, 13, 432. [Google Scholar] [CrossRef]
- Xiang, X.; Shi, Y.; Yang, J.; Kong, J.; Lin, X.; Zhang, H.; Zeng, J.; Chu, H. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci. Rep. 2014, 4, 3829. [Google Scholar] [CrossRef]
- Strauss, J.; Schirrmeister, L.; Grosse, G.; Fortier, D.; Hugelius, G.; Knoblauch, C.; Romanovsky, V.; Schädel, C.; von Deimling, T.S.; Schuur, E.A.; et al. Deep Yedoma permafrost: A synthesis of depositional characteristics and carbon vulnerability. Earth-Sci. Rev. 2017, 172, 75–86. [Google Scholar] [CrossRef]
- Antonova, S.; Sudhaus, H.; Strozzi, T.; Zwieback, S.; Kääb, A.; Heim, B.; Langer, M.; Bornemann, N.; Boike, J. Thaw subsidence of a yedoma landscape in northern Siberia, measured in situ and estimated from TerraSAR-X interferometry. Remote Sens. 2018, 10, 494–521. [Google Scholar] [CrossRef]
- Jin, H.; Wang, S.; Lu, L.; Yu, S. Features of permafrost degradation in Hinggan Mountains, Northeastern China. Sci. Geogr. Sin. 2009, 29, 223–228. (In Chinese) [Google Scholar]
- Li, Y.; Qing, Y.; Lyu, M.; Chen, S.; Yang, Z.; Lin, C.; Yang, Y. Effects of artificial warming on different soil organic carbon and nitrogen pools in a subtropical plantation. Soil Biol. Biochem. 2018, 124, 161–167. [Google Scholar] [CrossRef]
- Guo, L.; Chang, J.; Xu, H.; Sun, W. Modelling plant canopy effects on water-heat exchange in the freezing-thawing processes of active layer on the Qinghai-Tibet Plateau. J. Mt. Sci. 2021, 18, 1564–1579. [Google Scholar] [CrossRef]
- Yang, H.; Hong, X.; Yuan, Z.; He, X. Modelling the influence of vegetation on the hydrothermal processes of frozen soil in the Qinghai–Tibet Plateau. Water 2023, 15, 1692–1710. [Google Scholar] [CrossRef]
- Kelley, A.M.; Epstein, H.E.; Walker, D.A. Role of vegetation and climate in permafrost active layer depth in arctic tundra of northern Alaska and Canada. J. Glaciol. Geocryol. 2004, 26, 269–274. [Google Scholar]
- Chang, X.L.; Jin, H.J.; Wang, Y.P.; Zhang, Y.L.; Zhou, G.Y.; Che, F.Q.; Zhao, Y.M. Influences of vegetation on permafrost: A review. Acta Ecol. Sin. 2012, 32, 7981–7990. (In Chinese) [Google Scholar] [CrossRef]
- Schaphoff, S.; Reyer, C.P.O.; Schepaschenko, D.; Gerten, D.; Shvidenko, A. Tamm Review: Observed and projected climate change impacts on Russia’s forests and its carbon balance. For. Ecol. Manag. 2016, 361, 432–444. [Google Scholar] [CrossRef]
- Turetsky, M.R.; Kane, E.S.; Harden, J.W.; Ottmar, R.D.; Manies, K.L.; Hoy, E.; Kasischke, E.S. Recent acceleration of biomass burning and carbon losses in Alaskan forests and peatlands. Nat. Geosci. 2011, 4, 27–31. [Google Scholar] [CrossRef]
- Kong, J.; Yang, J. Effects of fire on soil properties and nutrient availability in a Dahurian larch forest in Great Xing’an Mountains of Northeast China. Chin. J. Ecol. 2013, 32, 2837–2843. [Google Scholar]
- Jiang, Y.; Rocha, A.V.; O’Donnell, J.A.; Drysdale, J.A.; Rastetter, E.B.; Shaver, G.R.; Zhuang, Q. Contrasting soil thermal responses to fire in Alaskan tundra and boreal forest. J. Geophys. Res. Earth Surf. 2015, 120, 363–378. [Google Scholar] [CrossRef]
- Smith, S.L.; Riseborough, D.W.; Bonnaventure, P.P. Eighteen year record of forest fire effects on ground thermal regimes and permafrost in the Central Mackenzie Valley, NWT, Canada. Permafr. Periglac. Process 2015, 26, 289–303. [Google Scholar] [CrossRef]
- Shenoy, A.; Johnstone, J.F.; Kasischke, S.; Kielland, K. Persistent effects of fire severity on early successional forests in interior Alaska. For. Ecol. Manag. 2011, 261, 381–390. [Google Scholar] [CrossRef]
- Alexander, H.D.; Natali, S.M.; Loranty, M.M.; Ludwig, S.M.; Spektor, V.V.; Davydov, S.; Zimov, N.; Trujillo, I.; Mack, M.C. Impacts of increased soil burn severity on larch forest regeneration on permafrost soils of far northeastern Siberia. For. Ecol. Manag. 2018, 417, 144–153. [Google Scholar] [CrossRef]
- Li, X.; Jin, H.; He, R.; Huang, Y.; Luo, D.; Jin, X.; Lü, L. Impacts of hazardous fires on permafrost environment: A review. J. Glaciol. Geocryol. 2017, 39, 317–327. (In Chinese) [Google Scholar]
- Holloway, J.E.; Lewkowicz, A.G.; Douglas, T.A.; Li, X.; Turetsky, M.R.; Baltzer, J.L.; Jin, H. Impact of wildfire on permafrost landscapes: A review of recent advances and future prospects. Permafr. Periglac. Process. 2020, 31, 371–382. [Google Scholar] [CrossRef]
- Li, T.; Jeřábek, J.; Winkler, J.; Vaverková, M.D.; Zumr, D. Effects of prescribed fire on topsoil properties: A small-scale straw burning experiment. J. Hydrol. Hydromech. 2022, 70, 450–461. [Google Scholar] [CrossRef]
- Zhang, Y.; Qin, B.; Sun, M.; Zhou, Z. Impact of forest fire on understory vegetation and soil in typical forest types of Daxing’an Mountains, northeastern China. J. Beijing For. Univ. 2012, 34, 7–13. (In Chinese) [Google Scholar]
- Bai, Q.; Zhou, M.; Qi, Q.; Meng, R.; Cui, X.; Wen, D. The changes of the frozen soil temperature and humidity of the forest fire forest in the Great Higgnan mountains. J. Arid. Land Res. Environ. 2013, 27, 69–75. (In Chinese) [Google Scholar]
- Nossov, D.R.; Jorgenson, M.T.; Kielland, K.; Kanevskiy, M.Z. Edaphic and microclimatic controls over permafrost response to fire in interior Alaska. Environ. Res. Lett. 2013, 8, 035013. [Google Scholar] [CrossRef]
- Li, X.Y.; Jin, H.J.; Wang, H.W.; Marchenko, S.S.; Shan, W.; Luo, D.L.; He, R.X.; Spektor, V.; Huang, Y.D.; Li, X.Y.; et al. Influences of forest fires on the permafrost environment: A review. Adv. Clim. Chang. Res. 2021, 12, 48–65. [Google Scholar] [CrossRef]
- Li, X.; Jin, H.; He, R.; Huang, Y.; Wang, H.; Luo, D.; Jin, X.; Lü, L.; Wang, L.; Li, W.; et al. Effects of forest fires on the permafrost environment in the northern Da Xing’anling (Hinggan) mountains, Northeast China. Permafr. Periglac. Process. 2019, 30, 163–177. [Google Scholar] [CrossRef]
- Jiang, X.; Cai, H.; Yang, X. Response of active layer thickening to wildfire in the pan-Arctic region: Permafrost type and vegetation type influences. Sci. Total Environ. 2023, 902, 166132. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Jorgenson, M.T.; Douglas, T.A.; Romanovsky, V.E.; Kielland, K.; Hiemstra, C.; Euskirchen, E.S.; Ruess, R.W. Interactive effects of wildfire and climate on permafrost degradation in Alaskan lowland forests. J. Geophys. Res. Biogeosci. 2015, 120, 1619–1637. [Google Scholar] [CrossRef]
- Hou, C.; Xie, F.; Li, X.; Xiao, D. Changes of permafrost active layer in greater Khingan mountains under fire disturbance. J. Univ. Jinan Sci. Technol. 2010, 24, 277–281. (In Chinese) [Google Scholar]
- Zhang, Z.; Wu, Q.; Zhou, Z. Risk assessment of freeze thawing disaster in permafrost zone. J. Nat. Disasters 2012, 21, 142–149. [Google Scholar]
- Anders, K.; Marx, S.; Boike, J.; Herfort, B.; Wilcox, E.J.; Langer, M.; Marsh, P.; Höfle, B. Multitemporal terrestrial laser scanning point clouds for thaw subsidence observation at Arctic permafrost monitoring sites. Earth Surf. Process. Landf. 2020, 45, 1589–1600. [Google Scholar] [CrossRef]
- Bouchard, F.; MacDonald, L.A.; Turner, K.W.; Thienpont, J.R.; Medeiros, A.S.; Biskaborn, B.K.; Korosi, J.; Hall, R.I.; Pienitz, R.; Wolfe, B.B. Paleolimnology of thermokarst lakes: A window into permafrost landscape evolution. Arct. Sci. 2016, 3, 91–117. [Google Scholar] [CrossRef]
- Turetsky, M.R.; Abbott, B.W.; Jones, M.C.; Anthony, K.W.; Olefeldt, D.; Schuur, E.A.G.; Grosse, G.; Kuhry, P.; Hugelius, G.; Koven, C.; et al. Carbon release through abrupt permafrost thaw. Nat. Geosci. 2020, 13, 138–143. [Google Scholar] [CrossRef]
- Mu, C. Thermokarst terrains change landscape and earth surface processes. Chin. J. Nat. 2020, 42, 386–392. (In Chinese) [Google Scholar]
- Luo, J.; Niu, F.; Lin, Z.; Liu, M.; Yin, G. Thermokarst lake changes between 1969 and 2010 in the Beilu River Basin, Qinghai–Tibet Plateau, China. Sci. Bull. 2015, 60, 556–564. [Google Scholar] [CrossRef]
- Smith, N.R.; Kishchuk, B.E.; Mohn, W.W. Effects of wildfire and harvest disturbances on forest soil Bacterial communities. Appl. Environ. Microbiol. 2008, 74, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.M.; Grosse, G.; Arp, C.D.; Miller, E.; Liu, L.; Hayes, D.J.; Larsen, C.F. Recent Arctic tundra fire initiates widespread thermokarst development. Sci. Rep. 2015, 5, 15865. [Google Scholar] [CrossRef] [PubMed]
- Ferrenberg, S.; O’neill, S.P.; Knelman, J.E.; Todd, B.; Duggan, S.; Bradley, D.; Robinson, T.; Schmidt, S.K.; Townsend, A.R.; Williams, M.W.; et al. Changes in assembly processes in soil bacterial communities following a wildfire disturbance. ISME J. 2013, 7, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Santalahti, M.E.; Pumpanen, J.; Köster, K.; Berninger, F.; Raffaello, T.; Asiegbu, F.O.; Heinonsalo, J. Bacterial community structure and function shift across a northern boreal forest fire chronosequence. Sci. Rep. 2016, 6, 32411. [Google Scholar] [CrossRef] [PubMed]
- Smithwick, E.A.H.; Naithani, K.J.; Balser, T.C.; Romme, W.H.; Turner, M.G. Post-fire spatial patterns of soil nitrogen mineralization and microbial abundance. PLoS ONE 2012, 7, e50597. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Sun, H.; Sietiö, O.M.; Pumpanen, J.; Heinonsalo, J.; Köster, K.; Berninger, F. Wildfire effects on soil bacterial community and its potential functions in a permafrost region of Canada. Appl. Soil Ecol. 2020, 156, 103713. [Google Scholar] [CrossRef]
- Pressler, Y.; Moore, J.C.; Cotrufo, M.F. Belowground community responses to fire: Meta-analysis reveals contrasting responses of soil microorganisms and mesofauna. Oikos 2019, 128, 309–327. [Google Scholar] [CrossRef]
- Jian, M.; Berli, M.; Ghezzehei, T.A. Soil structural degradation during low-severity burns. Geophys. Res. Lett. 2018, 45, 5553–5561. [Google Scholar] [CrossRef]
- She, R.; Yang, X.; Xiao, W. Research status on forest soil microbes under fire disturbance. J. Sichuan For. Sci. Technol. 2021, 42, 94–101. (In Chinese) [Google Scholar]
- Chen, F.; Zheng, H.; Ou-Yang, Z.; Zhang, K.; Tu, N. Responses of microbial community structure to the leaf litter composition. Acta Pedol. Sin. 2011, 48, 603–611. (In Chinese) [Google Scholar]
- Tao, Y.; Di, X. Fire interference on forest soil microbial communities and the mechanism: A review. Sci. Silvae. Sin. 2013, 49, 146–157. (In Chinese) [Google Scholar]
- Köster, K.; Aaltonen, H.; Berninger, F.; Heinonsalo, J.; Köster, E.; Ribeiro-Kumara, C.; Sun, H.; Tedersoo, L.; Zhou, X.; Pumpanen, J. Impacts of wildfire on soil microbiome in Boreal environments. Curr. Opin. Environ. Sci. Health 2021, 22, 100258. [Google Scholar] [CrossRef]
- Wang, D. Analysis of the influence of forest fire interference on the structure of soil microbial community in Pinus tabulaeformis forests. J. Temp. For. Res. 2023, 6, 84–86. (In Chinese) [Google Scholar]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.-A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]
- Peng, Y.; Cao, Y.; Qu, L. Effects of fire intensity on the soil microbial community of larch gmelinii forests. Bull. Bot. Res. 2017, 37, 549–555. (In Chinese) [Google Scholar]
- Gao, H.; Wang, Q. Research progress on the effects of logging and burning on forest soil microorganisms. Intern. J. Ecol. 2021, 10, 472–480. (In Chinese) [Google Scholar] [CrossRef]
- Granged, A.J.; Zavala, L.M.; Jordán, A.; Bárcenas-Moreno, G. Post-fire evolution of soil properties and vegetation cover in a Mediterranean heathland after experimental burning: A 3-year study. Geoderma 2011, 164, 85–94. [Google Scholar] [CrossRef]
- Alcañiz, M.; Outeiro, L.; Francos, M.; Farguell, J.; Úbeda, X. Long-term dynamics of soil chemical properties after a prescribed fire in a Mediterranean forest (Montgrí Massif, Catalonia, Spain). Sci. Total Environ. 2016, 572, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Santalahti, M.; Pumpanen, J.; Köster, K.; Berninger, F.; Raffaello, T.; Jumpponen, A.; Asiegbu, F.O.; Heinonsalo, J. Fungal community shifts in structure and function across a boreal forest fire chronosequence. Appl. Environ. Microbiol. 2015, 81, 7869–7880. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.; Whiteley, A.S. The bacterial biogeography of British soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef] [PubMed]
- Wieting, C.; Ebel, B.A.; Singha, K. Quantifying the effects of wildfire on changes in soil properties by surface burning of soils from the Boulder Creek Critical Zone Observatory. J. Hydrol. Reg. Stud. 2017, 13, 43–57. [Google Scholar] [CrossRef]
- Aaltonen, H.; Köster, K.; Köster, E.; Berninger, F.; Zhou, X.; Karhu, K.; Biasi, C.; Bruckman, V.; Palviainen, M.; Pumpanen, J. Forest fires in Canadian permafrost region: The combined effects of fire and permafrost dynamics on soil organic matter quality. Biogeochemistry 2019, 143, 257–274. [Google Scholar] [CrossRef]
- Hu, M.; Wang, J.; Lu, L.; Gui, H.; Wan, S. Global recovery patterns of soil microbes after fire. Soil Biol. Biochem. 2023, 183, 109057. [Google Scholar] [CrossRef]
- Das, S.; Deb, S.; Sahoo, S.S.; Sahoo, U.K. Soil microbial biomass carbon stock and its relation with climatic and other environmental factors in forest ecosystems: A review. Acta Ecol. Sin. 2023, 43, 933–945. [Google Scholar] [CrossRef]
- Holden, S.R.; Rogers, B.M.; Treseder, K.K.; Randerson, J.T. Fire severity influences the response of soil microbes to a boreal forest fire. Environ. Res. Lett. 2016, 11, 035004. [Google Scholar] [CrossRef]
- Cui, Y.; Shen, Y.; Zhang, Z. Influence of environmental factors on distribution and development of Xing’an-Baikal permafrost. J. Jilin Univ. Earth Sci. Ed. 2021, 51, 1427–1440. (In Chinese) [Google Scholar]
- Wang, F.; Tian, L.; Song, A.; Sang, Y.; Zhang, J.; Gao, J. Seasonal dynamics of microbial biomass carbon and nitrogen in soil of Robinia pseudoacacia forests and near-naturally restored vegetation in Northern China. Sci. Silvae Sin. 2015, 51, 16–24. (In Chinese) [Google Scholar]
- Jia, W.; Zhou, H.; Xie, W.; Guan, C.; Gao, C.; Shi, Y. Effects of long-term inorganic fertilizer combined with organic manure on microbial biomass C, N and enzyme activity in cinnamon soil. Plant Nutr. Fert. Sci. 2008, 14, 700–705. (In Chinese) [Google Scholar]
- Zhang, Y.; Pang, X.; Bao, W.; You, C.; Tang, H.; Hu, T. A review of soil organic matter and its research methods. World Sci. Tech. RD 2005, 27, 72–78. (In Chinese) [Google Scholar]
- Li, W.; Liu, X.; Niu Li, B.; Liu, G.; Chu, Y. Impact of fire on soil microbial biomass of Pinus tabuliformis forest in Pingquan County, Hebei of northern China. J. Beijing For. Univ. 2017, 39, 70–77. (In Chinese) [Google Scholar]
- Dove, N.C.; Taş, N.; Hart, S.C. Ecological and genomic responses of soil microbiomes to high-severity wildfire: Linking com-munity assembly to functional potential. ISME J. 2022, 16, 1853–1863. [Google Scholar] [CrossRef]
- Yang, Y.; Cheng, D.; Fu, H. Engineering geological characterstics and evaluations of permafrost in Daxing′ an Mountains. J. Eng. Geol. 2008, 16, 657–662. (In Chinese) [Google Scholar]
- Min, K.; Slessarev, E.; Kan, M.; McFarlane, K.; Oerter, E.; Pett-Ridge, J.; Nuccio, E.; Berhe, A.A. Active microbial biomass decreases, but microbial growth potential remains similar across soil depth profiles under deeply-vs. shallow-rooted plants. Soil Biol. Biochem. 2021, 162, 108401. [Google Scholar] [CrossRef]
- Zhao, B.; Sun, L.; Hu, H.; Sun, Z. Post-fire soil microbial biomass and nutrient content of Larix gmelinii forest in autumn. J. Nat. Res. 2011, 26, 450–459. (In Chinese) [Google Scholar]
- Allison, G. The influence of species diversity and stress intensity on community resistance and resilience. Ecol. Monogr. 2004, 74, 117–134. [Google Scholar] [CrossRef]
- Van Elsas, J.D.; Chiurazzi, M.; Mallon, C.A.; Elhottovā, D.; Krištůfek, V.; Salles, J.F. Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Natl. Acad. Sci. USA 2012, 109, 1159–1164. [Google Scholar] [CrossRef]
- Shen, J.-P.; Chen, C.R.; Lewis, T. Long term repeated fire disturbance alters soil bacterial diversity but not the abundance in an Australian wet sclerophyll forest. Sci. Rep. 2016, 6, 19639. [Google Scholar] [CrossRef]
- Pellegrini, A.F.A.; Ahlström, A.; Hobbie, S.E.; Reich, P.B.; Nieradzik, L.P.; Staver, A.C.; Scharenbroch, B.C.; Jumpponen, A.; Anderegg, W.R.L.; Randerson, J.T.; et al. Fire frequency drives decadal changes in soil carbon and nitrogen and ecosystem productivity. Nature 2018, 553, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.H. Diversity in tropical rain forests and coral reefs: High diversity of trees and corals is maintained only in a nonequilibrium state. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Hoy, E.E.; Turetsky, M.R.; Kasischke, E.S. More frequent burning increases vulnerability of Alaskan boreal black spruce forests. Environ. Res. Lett. 2016, 11, 095001. [Google Scholar] [CrossRef]
Figure 1.
Map of boreal−Arctic permafrost zonation. (Permafrost distribution data and figure are derived from Ran et al., 2021 [11]).
Figure 1.
Map of boreal−Arctic permafrost zonation. (Permafrost distribution data and figure are derived from Ran et al., 2021 [11]).

Figure 2.
Comparison of the number of publications and citations on soil microbial communities in permafrost and seasonally frozen ground regions from 2000 to 2023 (Data are derived from the citation index of the Web of Science: https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/02773368-6061-41eb-9254-0eefe2a1763d-d136b106?page=1 and https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/80c93092-41fd-48cd-838d-749c90f4486e-d1392c9f?page=1, accessed on 30 January 2024).
Figure 2.
Comparison of the number of publications and citations on soil microbial communities in permafrost and seasonally frozen ground regions from 2000 to 2023 (Data are derived from the citation index of the Web of Science: https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/02773368-6061-41eb-9254-0eefe2a1763d-d136b106?page=1 and https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/80c93092-41fd-48cd-838d-749c90f4486e-d1392c9f?page=1, accessed on 30 January 2024).
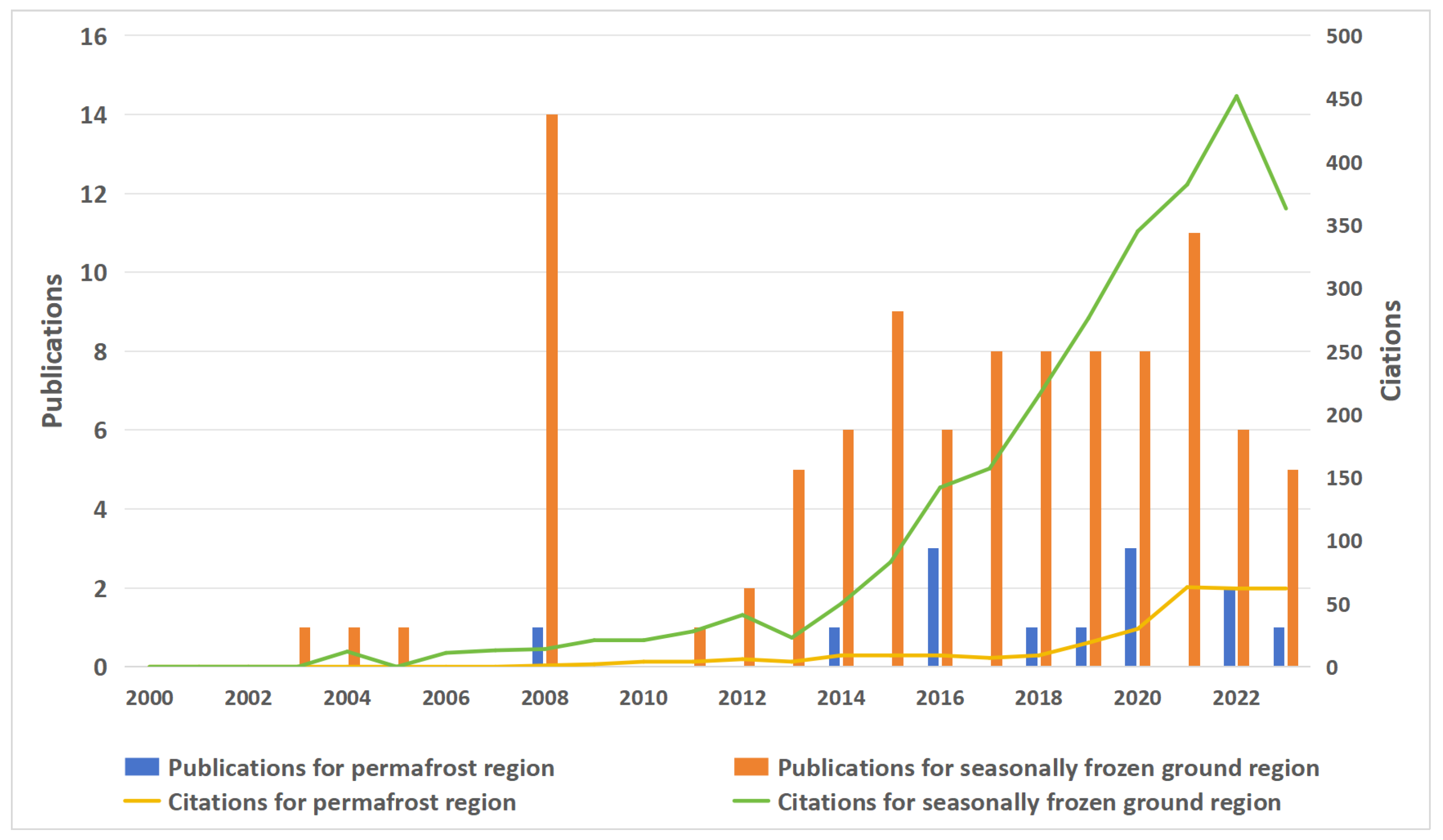
Figure 3.
Comparison of the number of publications and citations on the impact of forest fires on vegetation and soil microbes in the permafrost region from 2000 to 2023 (Data are derived from the citation index of the Web of Science: https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/99eedb70-36fa-49cc-a31d-e1cb17405f0d-d13477de?page=1 and https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/644aa2a4-202e-4da5-9b68-f41a15666c4f-d135bcfd?page=1, accessed on 30 January 2024).
Figure 3.
Comparison of the number of publications and citations on the impact of forest fires on vegetation and soil microbes in the permafrost region from 2000 to 2023 (Data are derived from the citation index of the Web of Science: https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/99eedb70-36fa-49cc-a31d-e1cb17405f0d-d13477de?page=1 and https://libvpn.nefu.edu.cn/piswebofscience.clarivate.cn/wos/alldb/citation-report/644aa2a4-202e-4da5-9b68-f41a15666c4f-d135bcfd?page=1, accessed on 30 January 2024).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, J.; Li, X.; Xu, T.; Han, Y.; Li, J.; Shen, Y.; Chen, K. Effects of Forest Fires on Boreal Permafrost and Soil Microorganisms: A Review. Forests 2024, 15, 501. https://doi.org/10.3390/f15030501
AMA Style
Liu J, Li X, Xu T, Han Y, Li J, Shen Y, Chen K. Effects of Forest Fires on Boreal Permafrost and Soil Microorganisms: A Review. Forests. 2024; 15(3):501. https://doi.org/10.3390/f15030501
Chicago/Turabian StyleLiu, Jing, Xiaoying Li, Tao Xu, Yilun Han, Jingtao Li, Yang Shen, and Kui Chen. 2024. "Effects of Forest Fires on Boreal Permafrost and Soil Microorganisms: A Review" Forests 15, no. 3: 501. https://doi.org/10.3390/f15030501
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.