Conifer Recruitment in Trembling Aspen (Populus Tremuloides Michx.) Stands along an East-West Gradient in the Boreal Mixedwoods of Canada

Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area and Sample Plot Data


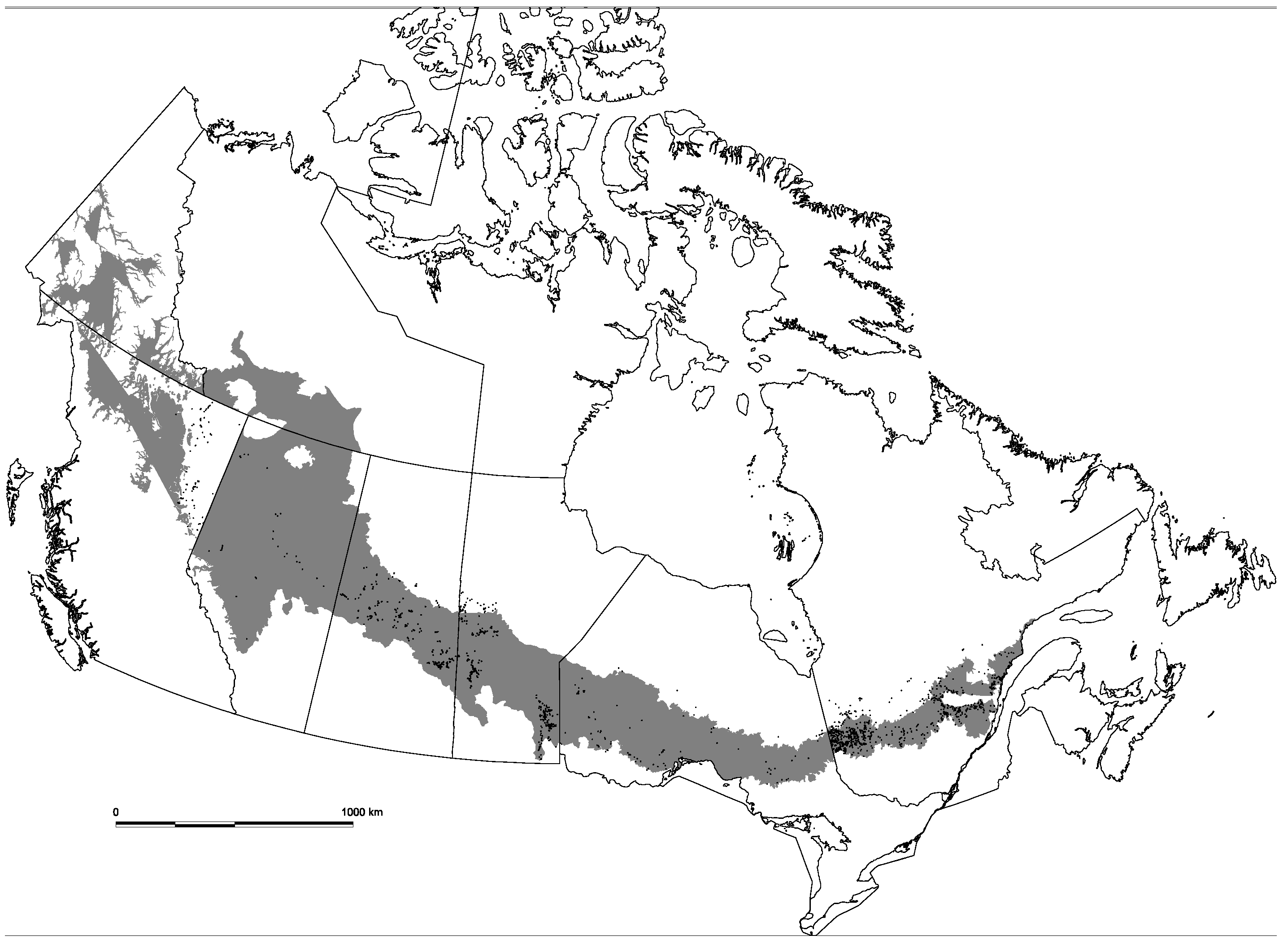{kind=link}
{kind=link}
{kind=link}
{kind=link}
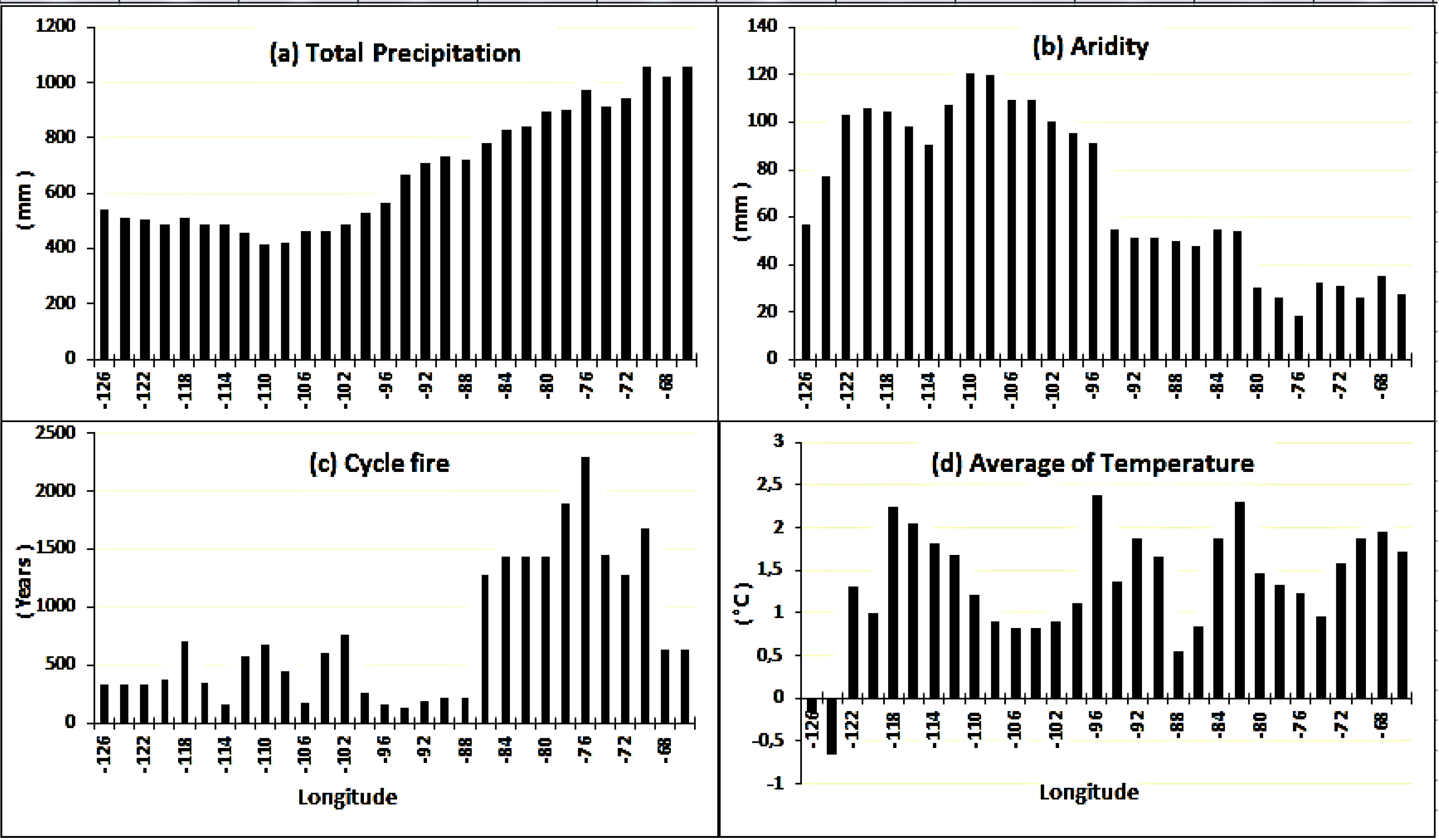
| Provinces a | No. of Stands b | Longitude (W) | Latitude (N) | T Mean (°C) c | Total Precipitation (mm) c | Relative Humidity (%) c | Drought Index (mm) c | Fire Cycle (years) c |
|---|---|---|---|---|---|---|---|---|
| BC | 100 | 120°04′–125°06′ | 55°11′–59°88′ | 0.8 (−1.3–2.9) | 504 (449–573) | 57 (53.6–60) | 97 (57.2–136.2) | 387.4 (326.8–397.9) |
| AB | 44 | 111°01′–119°43′ | 51°86′–58°58′ | 1.8 (−0.7–2.8) | 490 (415–560) | 54.4 (49.2–59.8) | 100.4 (69.7–126.7) | 332.7 (93–672.9) |
| SK | 470 | 101°75′–109°80′ | 52°33′–55°72′ | 0.9 (−0.3–1.4) | 461 (399–536) | 55.2 (50.8–60.4) | 107.7 (81–132.8) | 301.9 (50.9–1,225.6) |
| MB | 574 | 95°27′–101°92′ | 49°15′–55°36′ | 1.5 (−1.3–3.2) | 532 (445–657) | 57.3 (52.7–62.6) | 95.3 (72–129.9) | 325.7 (56.2–1,225.6) |
| ON | 155 | 79°85′–95°1′ | 47°5′–52°58′ | 1.6 (−0.4–2.7) | 736 (610–881) | 56.8 (52.5–61) | 51.1 (27.1–72.7) | 748.3 (83.9–1,440.2) |
| QC | 1293 | 66°98′–79°51′ | 47°54′–50°36′ | 1.4 (−0.7–3.7) | 931 (841–1256) | 55.8 (50.7–67.8) | 28 (10.4–64.5) | 1360.5 (625.7–3,706.6) |
| Total or ranges | 2636 | 66°98′–125°06′ | 47°5′–59°88′ | −1.3–3.7 | 399–1256 | 49.2–67.8 | 10.4–136.2 | 50.9–3706.6 |
2.2. Estimating Abundance and Occurrence of Regeneration
2.3. Predictive Variables
| Variable (Abbreviation) | Type | Description | Data Source |
|---|---|---|---|
| Climatic | |||
| Drought index (DI) | Continuous (Class) | Annual mean aridity (1981–2010) (mm) | Environment Canada |
| Degree-days (DD) | Continuous (Class) | Annual mean degree-day over 5 °C (1981–2010) (°C day) | Environment Canada |
| Fire regime | |||
| Fire cycle (FC) | Continuous (Class) | Year interval from cyclic return fire (years) | Boulanger et al., 2012 |
| Physical environment | |||
| Distance to the nearest great river, lake or bay (DistWater) ++ | Continuous (Class) | Distance to the nearest significant watercourse (km) | Natural resources of Canada, GeoBase |
| Hydrography (PctHydro) | Continuous (Class) | Percentage of area occupied by water in a radius of 20 km around the stand (%) | Natural resources of Canada, GeoBase |
| Physiographic regions (RegiPhysio) | Character | Three major regions of the physical geography of the Canadian landscape: Canadian Shield, Interior Plains and western Cordillera | Atlas of Canada |
| Anthropogenic | |||
| Distance to the nearest main road (DistRoad) | Continuous (Class) | Distance to the nearest carriage road (km) | Natural resources of Canada, GeoBase |
| Distance to agricultural area (DistAgri) | Continuous (Class) | Distance to the nearest farmland (km) | Natural resources of Canada, GeoBase |
| Road density (DensRoad) | Continuous (Class) | Total length of roads (km)/km2 area, in a radius of 15 and 25 km around the stand | Natural resources of Canada, GeoBase |
| Spatial | |||
| Longitude (Long) | Continuous | Geographic coordinates | Database forest inventory |
| Latitude (Lat) | Continuous | Geographic coordinates | Database forest inventory |
| Elevation (Elev) | Continuous | Geographic coordinates | Database forest inventory |
2.4. Data Analysis and Model Selection
3. Results
3.1. Abundance and Occurrence of Conifer Recruitment

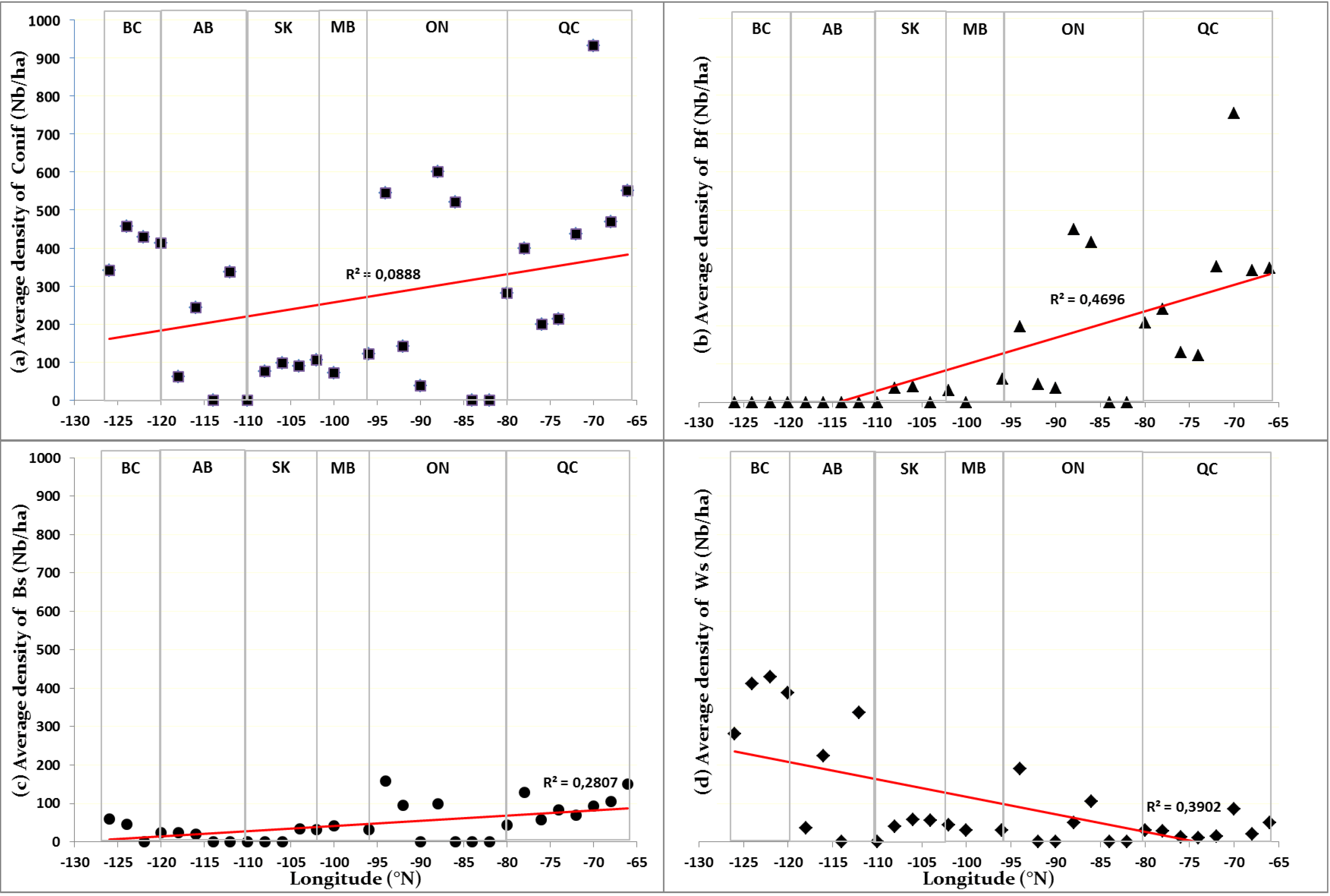
3.2. Variability in Environmental Conditions along the Study Gradient

 ), white spruce (
), white spruce (  ), black spruce (
), black spruce (  ) and balsam fir (
) and balsam fir (  ) respectively.
) respectively.3.3. Selection of Predictive Models and Individual Effect of Variables
| Response | Candidate Models | Model ID | LL | AIC | ΔAIC | Wi |
|---|---|---|---|---|---|---|
| Conif | DI + DensRoad25Km + Lat | mod1 | −22,158 | 44,325 | 0.0 | 0.61 |
| DI + DensRoad25Km + DD | mod2 | −22,158 | 44,327 | 1.5 | 0.29 | |
| DI + DensRoad25Km | mod3 | −22,160 | 44,329 | 3.5 | 0.11 | |
| DI | mod4 | −22,169 | 44,344 | 19.2 | 0.00 | |
| RegiPhysio + DensRoad25Km + FC + DistAgri | mod5 | −22,175 | 44,364 | 39.1 | 0.00 | |
| FC + DensRoad25Km + Lat | mod6 | −22,186 | 44,381 | 56.1 | 0.00 | |
| DD + DensRoad25Km + FC + Elev | mod7 | −22,185 | 44,382 | 56.4 | 0.00 | |
| DD + DensRoad25Km + FC | mod8 | −22,192 | 44,393 | 68.0 | 0.00 | |
| Ws | DD + DensRoad25Km + Lat | mod1 | −17,922 | 35,855 | 0.0 | 0.88 |
| DD + DensRoad25Km + Long | mod2 | −17,925 | 35,860 | 4.5 | 0.09 | |
| DI + DistRoad + Lat | mod3 | −17,927 | 35,862 | 6.8 | 0.03 | |
| DD + DensRoad25Km | mod4 | −17,930 | 35,869 | 13.9 | 0.00 | |
| DD + Long + Lat | mod5 | −17,932 | 35,873 | 18.1 | 0.00 | |
| DI + DistRoad + Long | mod6 | −17,932 | 35,875 | 19.6 | 0.00 | |
| DD + Long | mod7 | −17,937 | 35,883 | 27.9 | 0.00 | |
| DI + Long | mod8 | −17,937 | 35,883 | 28.0 | 0.00 | |
| Bs | DI + DensRoad25Km + Lat | mod1 | −19,065 | 38,140 | 0.0 | 0.39 |
| DI + DensRoad25Km + Long | mod2 | −19,066 | 38,141 | 0.8 | 0.26 | |
| DI + FC + DensRoad25Km | mod3 | −19,066 | 38,141 | 1.1 | 0.23 | |
| DI + DensRoad25Km | mod4 | −19,067 | 38,142 | 2.3 | 0.12 | |
| DI | mod5 | −19,081 | 38,169 | 28.4 | 0.00 | |
| DD + FC + DensRoad25Km + Elev | mod6 | −19,081 | 38,174 | 33.6 | 0.00 | |
| DD + FC + DensRoad25Km | mod7 | −19,084 | 38,178 | 37.8 | 0.00 | |
| FC + DensRoad25Km | mod8 | −19,087 | 38,181 | 40.7 | 0.00 | |
| Bf | DI | mod1 | −21,874 | 43,754 | 0.0 | 0.99 |
| RegiPhysio + FC + DistWater | mod2 | −21,880 | 43,773 | 19.0 | 0.00 | |
| FC + DD + RegiPhysio | mod3 | −21,881 | 43,774 | 19.8 | 0.00 | |
| FC + RegiPhysio | mod4 | −21,884 | 43,778 | 24.1 | 0.00 | |
| FC + DistWater | mod5 | −21,898 | 43,805 | 51.0 | 0.00 | |
| FC + DD | mod6 | −21,899 | 43,807 | 52.7 | 0.00 | |
| FC | mod7 | −21,901 | 43,808 | 54.3 | 0.00 |
| Response | Parameters | Estimate | Lower 95% CI | Upper 95% CI | SE | t-Value | Pr (>|t|) |
|---|---|---|---|---|---|---|---|
| Conif | DI | −131.8 | −165.3 | −98.3 | 17.1 | −7.72 | 0.00 |
| DensRoad25Km | −123.4 | −187.4 | −59.4 | 32.6 | −3.78 | 0.00 | |
| Lat | 33.4 | 5.4 | 61.4 | 14.3 | 2.34 | 0.02 | |
| Ws | DD | −24.9 | −35.5 | −14.2 | 5.4 | −4.57 | 0.00 |
| DensRoad25Km | −31.5 | −44.4 | −18.7 | 6.6 | −4.80 | 0.00 | |
| Lat | 6.3 | 3.2 | 9.5 | 1.6 | 3.99 | 0.00 | |
| Bs | DI | −29.9 | −40.3 | −19.6 | 5.3 | −5.66 | 0.00 |
| DensRoad25Km | −49.6 | −69.4 | −29.8 | 10.1 | −4.91 | 0.00 | |
| Lat | 9.2 | 0.5 | 17.8 | 4.4 | 2.08 | 0.03 | |
| Bf | DI | −77.2 | −93.7 | −60.7 | 8.4 | −9.19 | 0.00 |
3.4. Variance Explained by Each Predictor Variable in the Best Model
| Model | Predictors | Variance (%) |
|---|---|---|
| Conif | DI | 45.8 |
| DensRoad25Km | 11.1 | |
| Lat | 4.3 | |
| Ws | DensRoad25Km | 29.4 |
| DD | 26.6 | |
| Lat | 20.3 | |
| Bs | DI | 39.8 |
| DensRoad25Km | 29.3 | |
| Lat | 3.1 | |
| Bf | DI | ‡ |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Appendix

| Response | Candidate Models | Model ID | LL | AIC | ΔAIC | Wi |
|---|---|---|---|---|---|---|
| Conif | DistAgri + DI + Lat + RegiPhysio + Long + Elev + DD + PctHydro | mod1 | −1,111 | 2,240 | 0.0 | 0.98 |
| DistAgri + DI + Lat + RegiPhysio + Long + Elev | mod2 | −1,117 | 2,248 | 8.3 | 0.02 | |
| DistAgri + DI + Lat + RegiPhysio + Long | mod3 | −1,123 | 2,257 | 17.2 | 0.00 | |
| DistAgri + DI + PctHydro + Lat + RegiPhysio + DD | mod4 | −1,132 | 2,278 | 38.1 | 0.00 | |
| DistAgri + DensRoad25Km + DI + PctHydro + Lat | mod5 | −1,136 | 2,283 | 43.2 | 0.00 | |
| DistAgri + DensRoad25Km + DI + DD | mod6 | −1,147 | 2,304 | 63.9 | 0.00 | |
| DistAgri + DensRoad25Km + DI | mod7 | −1,157 | 2,321 | 81.1 | 0.00 | |
| DistAgri + RegiPhysio + FC + DensRoad25Km | mod8 | −1,198 | 2,406 | 165.8 | 0.00 | |
| Ws | DensRoad25Km + DI + Lat + Long + Elev + DD | mod1 | −506 | 1,026 | 0.0 | 0.88 |
| DensRoad25Km + DI + Lat + Long + Elev | mod2 | −509 | 1,030 | 3.9 | 0.12 | |
| PctHydro + DensRoad25Km + DI + Lat + Long | mod3 | −519 | 1,049 | 23.1 | 0.00 | |
| PctHydro + DensRoad25Km + DI + Lat + DistAgri | mod4 | −519 | 1,051 | 24.5 | 0.00 | |
| PctHydro + DensRoad25Km + DI + Lat | mod5 | −523 | 1,055 | 28.7 | 0.00 | |
| DD + DistAgri + PctHydro + DensRoad25Km + Long + DI | mod6 | −525 | 1,065 | 38.5 | 0.00 | |
| DD + DistAgri + PctHydro + DensRoad25Km | mod7 | −534 | 1,079 | 52.4 | 0.00 | |
| DI + DD + FC | mod8 | −541 | 1,090 | 64.0 | 0.00 | |
| Bs | DistAgri + DensRoad25Km + DI + Lat + Long + RegiPhysio | mod1 | −585 | 1,184 | 0.0 | 0.39 |
| DistAgri + DI + Lat + Long + RegiPhysio + DistRoad | mod2 | −585 | 1,184 | 0.2 | 0.35 | |
| DistAgri + DensRoad25Km + DI + Lat + Long | mod3 | −587 | 1,185 | 0.9 | 0.25 | |
| DI + DD + FC + DistAgri + DensRoad25Km | mod4 | −591 | 1,195 | 10.5 | 0.00 | |
| DensRoad25Km + DI + Lat + Long | mod5 | −593 | 1,196 | 11.9 | 0.00 | |
| DistAgri + DensRoad25Km + DI + Lat + Long + RegiPhysio | mod6 | −593 | 1,197 | 12.7 | 0.00 | |
| DI + DD + FC + DistAgri | mod7 | −595 | 1,200 | 15.8 | 0.00 | |
| DI + DD + FC | mod8 | −602 | 1,212 | 28.3 | 0.00 | |
| Bf | RegiPhysio + PctHydro + DistAgri + DI + Long + FC | mod1 | −767 | 1,548 | 0.0 | 0.87 |
| RegiPhysio + PctHydro + DistAgri + DI + Long | mod2 | −770 | 1,552 | 3.8 | 0.13 | |
| RegiPhysio + DistAgri + DI + Long + FC | mod3 | −775 | 1,563 | 14.7 | 0.00 | |
| DistAgri + DI + Long + FC | mod4 | −781 | 1,572 | 23.8 | 0.00 | |
| RegiPhysio + PctHydro + DensRoad15Km + DistAgri + DI | mod5 | −783 | 1,578 | 30 | 0.00 | |
| RegiPhysio + PctHydro + DensRoad15Km + FC + DistAgri | mod6 | −799 | 1,611 | 62.6 | 0.00 | |
| RegiPhysio + PctHydro + DensRoad15Km + FC | mod7 | −803 | 1,615 | 67.2 | 0.00 |
Conflicts of Interest
References
- Qian, H.; Klinka, K.; Økland, R.H.; Krestov, P.; Kayahara, G.J. Understorey vegetation in boreal Picea Mariana and Populus Tremuloides stands in British Columbia. J. Veg. Sci. 2003, 14, 173–184. [Google Scholar]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley and Sons, Inc.: New York, NY, USA, 1996. [Google Scholar]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Gauthier, S.; Grandpré, L.; Bergeron, Y. Differences in forest composition in two boreal forest ecoregions of Quebec. J. Veg. Sci. 2000, 11, 781–790. [Google Scholar]
- Bergeron, Y.; Gauthier, S.; Flannigan, M.; Kafka, V. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Quebec. Ecology 2004, 85, 1916–1932. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y. Fire, logging, and overstory affect understory abundance, diversity, and composition in boreal forest. Ecol. Monogr. 2008, 78, 123–140. [Google Scholar] [CrossRef]
- Chen, H.Y.; Popadiouk, R.V. Dynamics of North American boreal mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Bergeron, Y.; Fenton, N. Boreal forests of eastern Canada revisited: Old growth, nonfire disturbances, forest succession, and biodiversity. Botany 2012, 90, 509–523. [Google Scholar] [CrossRef]
- Peters, V.S.; Macdonald, S.E.; Dale, M.R.T. Patterns of initial versus delayed regeneration of white spruce in boreal mixedwood succession. Can. J. For. Res. 2006, 36, 1597–1609. [Google Scholar] [CrossRef]
- Stocks, B.J.; Mason, J.A.; Todd, J.B.; Bosch, E.M.; Wotton, B.M.; Amiro, B.D.; Flannigan, M.D.; Hirsch, K.G.; Logan, K.A.; Martell, D.L.; et al. Large forest fires in Canada, 1959–1997. J. Geophys. Res. 2002, 107. [Google Scholar] [CrossRef]
- Bergeron, Y.; Richard, P.J.H.; Carcaillet, C.; Gauthier, S.; Flannigan, M.; Prairie, Y.T. Variability in fire frequency and forest composition in Canada’s southeastern boreal forest: A challenge for sustainable forest management. Conserv. Ecol. 1998, 2, pp. 1–11. Available online: ://www.consecol.org/vol2/iss2/art6/ (accessed on 17 December 2013).
- Taylor, A.R.; Chen, H.Y. Multiple successional pathways of boreal forest stands in central Canada. Ecography 2011, 34, 208–219. [Google Scholar] [CrossRef]
- Nilsson, M.-C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Bergeron, Y.; Dubuc, M. Succession in the southern part of the Canadian boreal forest. Vegetatio 1988, 79, 51–63. [Google Scholar] [CrossRef]
- Johnstone, J.; Chapin, F.S., III. Fire interval effects on successional trajectory in boreal forests of northwest Canada. Ecosystems 2006, 9, 268–277. [Google Scholar] [CrossRef]
- Cumming, S.G.; Schmiegelow, F.K.A.; Burton, P.J. Gap dynamics in boreal aspen stands: Is the forest older than we think? Ecol. Appl. 2000, 10, 744–759. [Google Scholar]
- Caners, R.; Kenkel, N. Forest stand structure and dynamics at Riding Mountain National Park, Manitoba, Canada. Community Ecol. 2003, 4, 185–204. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Chapin, F.S., III; Foote, J.; Kemmett, S.; Price, K.; Viereck, L. Decadal observations of tree regeneration following fire in boreal forests. Can. J. For. Res. 2004, 34, 267–273. [Google Scholar] [CrossRef]
- Galipeau, C.; Kneeshaw, D.D.; Bergeron, Y. White spruce and balsam fir colonization of a site in the southeastern boreal forest as observed 68 years after fire. Can. J. For. Res. 1997, 27, 139–147. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.-P. Paysages Régionaux du Quebec Méridional. In MRN (Ministère des Ressources Naturelles); Gouvernement du Québec, Les publications du Québec: Québec, Canada, 1998; p. 213. [Google Scholar]
- Flannigan, M.D.; Bergeron, Y.; Engelmark, O.; Wotton, B.M. Future wildfire in circumboreal forests in relation to global warming. J. Veg. Sci. 1998, 9, 469–476. [Google Scholar] [CrossRef]
- Bergeron, Y.; Cyr, D.; Girardin, M.P.; Carcaillet, C. Will climate change drive 21st century burn rates in Canadian boreal forest outside of its natural variability: Collating global climate model experiments with sedimentary charcoal data. Int. J. Wildland Fire 2010, 19, 1127–1139. [Google Scholar] [CrossRef]
- Wotton, B.M.; Nock, C.A.; Flannigan, M.D. Forest fire occurrence and climate change in Canada. Int. J. Wildland Fire 2010, 19, 253–271. [Google Scholar] [CrossRef]
- Girardin, M.P.; Ali, A.A.; Carcaillet, C.; Gauthier, S.; Hely, C.; LeGoff, H.; Terrier, A.; Bergeron, Y. Fire in managed forests of eastern Canada: Risks and options. For. Ecol. Manage. 2013, 294, 238–249. [Google Scholar] [CrossRef]
- Bergeron, Y.; Gauthier, S.; Kafka, V.; Lefort, P.; Lesieur, D. Natural fire frequency for the eastern Canadian boreal forest: Consequences for sustainable forestry. Can. J. For. Res. 2001, 31, 384–391. [Google Scholar] [CrossRef]
- Bergeron, Y.; Chen, H.Y.H.; Kenkel, N.C.; Leduc, A.L.; Macdonald, S.E. Boreal mixedwood stand dynamics: Ecological processes underlying multiple pathways. For. Chron. 2014, 90, 202–213. [Google Scholar] [CrossRef]
- Reyes, G.P.; Kneeshaw, D.; de Grandpré, L. The relative importance of natural disturbances and local site factors on woody vegetation regeneration diversity across a large, contiguous forest region. Open J. For. 2013, 3, 88–98. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Stadt, K.J.; Navratil, S. Age structure and growth of understory white spruce under aspen. Can. J. For. Res. 1996, 26, 1002–1007. [Google Scholar] [CrossRef]
- Greene, D.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.-J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- MacIsaac, D.A.; Comeau, P.G.; Macdonald, S.E. Dynamics of regeneration gaps following harvest of aspen stands. Can. J. For. Res. 2006, 36, 1818–1833. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Armstrong, G.W.; Stadt, K.J.; Marenholtz, E.H. Forest regeneration standards: Are they limiting management options for Alberta’s boreal mixedwoods? For. Chron. 2008, 84, 76–82. [Google Scholar] [CrossRef]
- Arbour, M.L.; Bergeron, Y. Effect of increased Populus cover on Abies regeneration in the Picea-feathermoss boreal forest. J. Veg. Sci. 2011, 22, 1132–1142. [Google Scholar] [CrossRef]
- Hart, S.A.; Chen, H.Y. Understory vegetation dynamics of North American boreal forests. Crit. Rev. Plant Sci. 2006, 25, 381–397. [Google Scholar] [CrossRef]
- Boulanger, Y.; Gauthier, S.; Burton, P.J.; Vaillancourt, M.-A. An alternative fire regime zonation for Canada. Int. J. Wildland Fire 2012, 21, 1052–1064. [Google Scholar] [CrossRef]
- McIntire, E.J.B.; Duchesneau, R.; Kimmins, J.P. Seed and bud legacies interact with varying fire regimes to drive long-term dynamics of boreal forest communities. Can. J. For. Res. 2005, 35, 2765–2773. [Google Scholar] [CrossRef]
- Martin-DeMoor, J.; Lieffers, V.J.; Macdonald, S.E. Natural regeneration of white spruce in aspen-dominated boreal mixedwoods following harvesting. Can. J. For. Res. 2010, 40, 585–594. [Google Scholar] [CrossRef]
- Payette, S. Fire as a controlling process in the North American boreal fores. In A Systems Analysis of the Global Boreal Forest; Shugart, H.H., Leemans, R., Bonan, G.B., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 144–169. [Google Scholar]
- Bergeron, Y.; Flannigan, M.D. Predicting the effects of climate change on fire frequency in the southeastern Canadian boreal forest. Water Air Soil Pollut. 1995, 82, 437–444. [Google Scholar] [CrossRef]
- Bergeron, Y.; Leduc, A.; Harvey, B.D.; Gauthier, S. Natural fire regime: A guide for sustainable management of the Canadian boreal forest. Silva Fenn. 2002, 36, 81–95. [Google Scholar] [CrossRef]
- Chhin, S.; Hogg, E.; Lieffers, V.J.; Huang, S. Potential effects of climate change on the growth of lodgepole pine across diameter size classes and ecological regions. For. Ecol. Manag. 2008, 256, 1692–1703. [Google Scholar] [CrossRef]
- Johnson, E.A.; Miyanishi, K.; Weir, J.M.H. Wildfires in the western Canadian boreal forest: Landscape patterns and ecosystem management. J. Veg. Sci. 1998, 9, 603–610. [Google Scholar] [CrossRef]
- Baldwin, K.A.; MacKenzie, W.H.; Pfalz, J.; Meades, W.J.; Meidinger, D.V.; Robitaille, A.; Saucier, J.-P.; Uhlig, P. Level 4 Map, Version 1, Canadian Component of the Circumboreal Vegetation Map (CBVM); Canadian regional team of the Circumboreal Vegetation Map Project. Natural Resources Canada, Canadian Forest Service, Great Lakes Forestry Centre: Sault Ste. Marie, ON, Canada, 2012. [Google Scholar]
- Régnière, J.; Saint-Amant, R. BioSIM 9—Manuel de L’utilisateur. Ressources Naturelles Canada, Service Canadien des Forêts, Centre de Foresterie des Laurentide. 2008. Available online: ftp://www.ftp.cfl.scf.rncan.gc.ca/regniere/Data/Weather/ (accessed on 4 February 2012).
- Environment Canada. National Climate Data and Information Archive. 2013. Available online: http://climate.weatheroffice.gc.ca/ (accessed on 23 September 2013).
- Villeneuve, G.O. Glossaire de Métérologie et de Climatologie, 2nd ed.; Les Presses de l’université Laval: Québec, QC, Canada, 1980; p. 651. [Google Scholar]
- Kaennel, M.; Schweingruber, F.H. Multilingual Glossary of Dendrochronology; Birmensdorf, Swiss Federal Institute for Forest, Snow and Landscape Research: Berne, Stuttgart, Vienna, 1995; p. 467. [Google Scholar]
- Laquerre, S.; Leduc, A.; Harvey, B.D. Augmentation du couvert en peuplier faux-tremble dans les pessières noires du nord-ouest du Québec après coupe totale. Ecoscience 2009, 16, 483–491. [Google Scholar] [CrossRef]
- R-Development-Core-Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. ISBN 3-900051-07-0. Available online: http://www.R-project.org/ (accessed on 15 November 2013).
- Mazerolle, M.J. Improving data analysis in herpetology: Using Akaike’s Information Criterion (AIC) to assess the strength of biological hypotheses. Amphib.Reptil. 2006, 27, 169–180. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer-Verlag: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Lesieur, D.; Gauthier, S.; Bergeron, Y. Fire frequency and vegetation dynamics for the south-central boreal forest of Quebec, Canada. Can. J. For. Res. 2002, 32, 1996–2009. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Lieffers, V.J. Growth of Populus tremuloides in association with Calamagrostis canadensis. Can. J. For. Res. 1998, 28, 396–401. [Google Scholar] [CrossRef]
- Zasada, J.C.; Sharik, T.L.; Nygren, M. The reproductive process in boreal forest trees. In A Systems Analysis of the Global Boreal Forest; Shugart, H.H., Leemans, R., Bonan, G.B., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 85–125. [Google Scholar]
- Messaoud, Y.; Bergeron, Y.; Asselin, H. Reproductive potential of balsam fir (Abies balsamea), white spruce (Picea glauca), and black spruce (P. mariana) at the ecotone between mixedwood and coniferous forests in the boreal zone of western Quebec. Am. J. Bot. 2007, 94, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Frank, R.M. Balsam fir. Silvics of North America; Agriculture Handbook 654. Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; p. 26. Available online: http://na.fs.fed.us/spfo/pubs/silvics_manual/Volume_1/abies/balsamea.htm (accessed on 28 March 2014).
- Greene, D.F.; Noel, J.; Bergeron, Y.; Rousseau, M.; Gauthier, S. Recruitment of Picea mariana, Pinus banksiana, and Populus tremuloides across a burn severity gradient following wildfire in the southern boreal forest of Quebec. Can. J. For. Res. 2004, 34, 1845–1857. [Google Scholar] [CrossRef]
- Rowe, J.S. Forest regions of Canada; Fisheries and Environment Canada, Canadian Forest Service: Ottawa, ON, Canada, 1972. Available online: http://cfs.nrcan.gc.ca/pubwarehouse/pdfs/24040.pdf (accessed on 11 February 2014).
- Williams, J.W.; Shuman, B.N.; Webb, T., III; Bartlein, P.J.; Leduc, P.L. Late-Quaternary vegetation dynamics in North America: Scaling from taxa to biomes. Ecol. Monogr. 2004, 74, 309–334. [Google Scholar] [CrossRef]
- National Oceanic Atmospheric Administration. WDC for Paleoclimatology. 2013. Available online: http://www.ncdc.noaa.gov/paleo/pollen/viewer/webviewer.html (accessed on 20 June 2014). [Google Scholar]
- Gärtner, S.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and management of natural regeneration of white spruce in the boreal forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]
- Greene, D.F.; Messier, C.; Asselin, H.; Fortin, M.-J. The effect of light availability and basal area on cone production in Abies balsamea and Picea glauca. Can. J. Bot. 2002, 80, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, J.F.; Chapin, F.S., III. Effects of soil burn severity on post-fire tree recruitment in boreal forest. Ecosystems 2006, 9, 14–31. [Google Scholar] [CrossRef]
- Girardin, M.P.; Tardif, J.C.; Flannigan, M.D.; Wotton, B.M.; Bergeron, Y. Trends and periodicities in the Canadian Drought Code and their relationships with atmospheric circulation for the southern Canadian boreal forest. Can. J. For. Res. 2004, 34, 103–119. [Google Scholar] [CrossRef]
- Amiro, B.D.; Logan, K.; Wotton, B.; Flannigan, M.; Todd, J.; Stocks, B.; Martell, D. Fire weather index system components for large fires in the Canadian boreal forest. Int. J. Wildland Fire 2005, 13, 391–400. [Google Scholar] [CrossRef]
- Girardin, M.P.; Mudelsee, M. Past and future changes in Canadian boreal wildfire activity. Ecol. Appl. 2008, 18, 391–406. [Google Scholar] [CrossRef]
- Girardin, M.P.; Wotton, B.M. Summer moisture and wildfire risks across Canada. J. Appl. Meteo. Climat. 2009, 48, 517–533. [Google Scholar] [CrossRef]
- Boucher, Y.; Arseneault, D.; Sirois, L. Logging-induced change (1930–2002) of a preindustrial landscape at the northern range limit of northern hardwoods, eastern Canada. Can. J. For. Res. 2006, 36, 505–517. [Google Scholar] [CrossRef]
- Archambault, L.; Morissette, J.; Bernier-Cardou, M. Forest succession over a 20-year period following clearcutting in balsam fir-yellow birch ecosystems of eastern Quebec, Canada. For. Ecol. Manag. 1998, 102, 61–74. [Google Scholar] [CrossRef]
- Terrail, R. Influence de la Colonisation sur les Transformations du Paysage Forestier Depuis L’époque Préindustrielle Dans l’Est du Québec (Canada). Ph.D. Thesis, Université du Québec à Rimouski, Rimouski, Juillet, Canada, 2013. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nlungu-Kweta, P.; Leduc, A.; Bergeron, Y. Conifer Recruitment in Trembling Aspen (Populus Tremuloides Michx.) Stands along an East-West Gradient in the Boreal Mixedwoods of Canada. Forests 2014, 5, 2905-2928. https://doi.org/10.3390/f5112905
Nlungu-Kweta P, Leduc A, Bergeron Y. Conifer Recruitment in Trembling Aspen (Populus Tremuloides Michx.) Stands along an East-West Gradient in the Boreal Mixedwoods of Canada. Forests. 2014; 5(11):2905-2928. https://doi.org/10.3390/f5112905
Chicago/Turabian StyleNlungu-Kweta, Pierre, Alain Leduc, and Yves Bergeron. 2014. "Conifer Recruitment in Trembling Aspen (Populus Tremuloides Michx.) Stands along an East-West Gradient in the Boreal Mixedwoods of Canada" Forests 5, no. 11: 2905-2928. https://doi.org/10.3390/f5112905
APA StyleNlungu-Kweta, P., Leduc, A., & Bergeron, Y. (2014). Conifer Recruitment in Trembling Aspen (Populus Tremuloides Michx.) Stands along an East-West Gradient in the Boreal Mixedwoods of Canada. Forests, 5(11), 2905-2928. https://doi.org/10.3390/f5112905