Douglas-Fir Tussock Moth- and Douglas-Fir Beetle-Caused Mortality in a Ponderosa Pine/Douglas-Fir Forest in the Colorado Front Range, USA

Abstract
:1. Introduction
2. Experimental Section
2.1. Study Area

2.2. Sampling
3. Results


{kind=link}
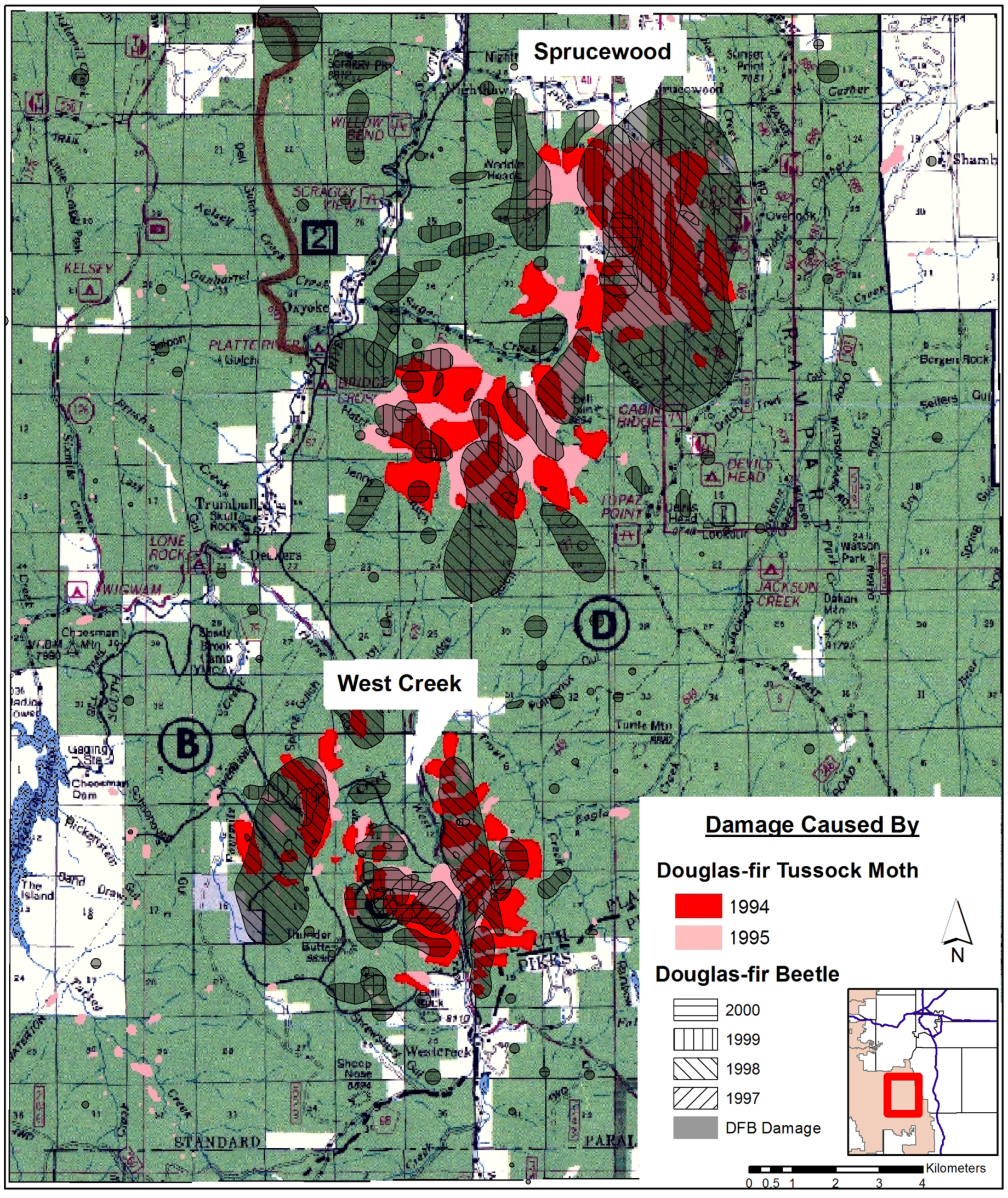{kind=link}
{kind=link}
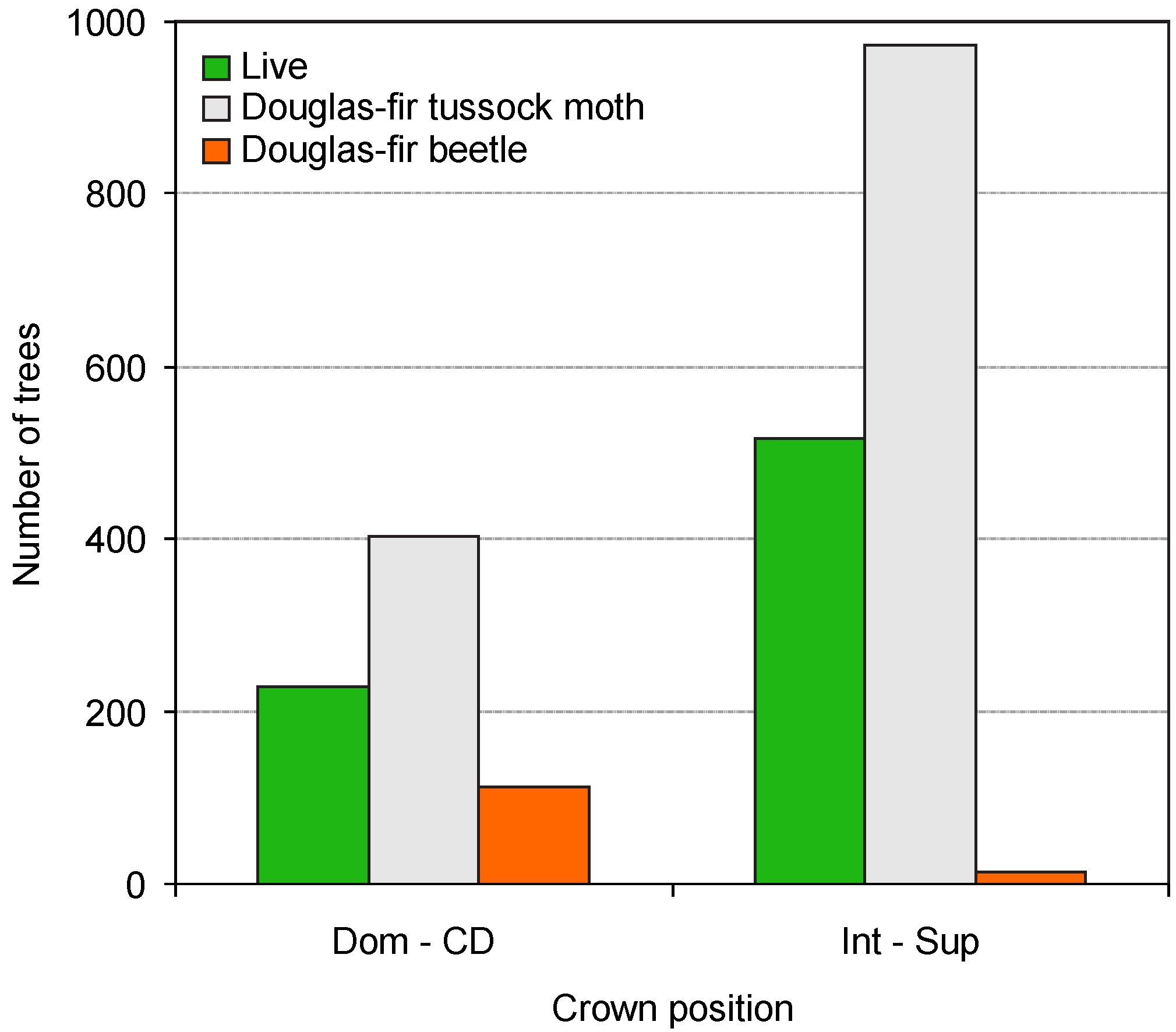{kind=link}
| Heavily Defoliated Stands (n = 6) | Lightly Defoliated Stands (n = 5) | |||||||
|---|---|---|---|---|---|---|---|---|
| Initial Condition | Post-Outbreak | DFTM-Killed | DFB-Killed | Initial Condition | Post-Outbreak | DFTM-Killed | DFB-Killed | |
| Stand Characteristics | ||||||||
| All Species | ||||||||
| Basal Area (m2/ha) | 25.1 (1.8) | 8.0 (1.2) * | -- | -- | 22.4 (0.8) | 18.6 (2.0) * | -- | -- |
| Density (tph) | 1115 (101) | 307 (31) * | -- | -- | 942 (102) | 845 (124) * | -- | -- |
| Douglas-fir | ||||||||
| Basal Area (m2/ha) | 19.4 (1.5) | 2.4 (0.4) * | 12.8 (2.1) * | 4.1 (1.7) | 15.0 (2.2) | 10.6 (3.6) * | 1.2 (0.6) * | 2.7 (1.4) |
| Density (tph) | 941 (104) | 67 (25) | 763 (113) * | 43.3 (17.5) | 657 (137) | 309 (186) | 68.0 (40) * | 30.0 (14.0) |
| % Basal Area | 74.8 (5.4) † | 17.5 (5.0) † | -- | -- | 63.9 (10.7) | 33.8 (16.2) | -- | -- |
| % Density | 78.4(3.6) † | 18.3 (4.2) † | -- | -- | 65.0 (1.9) | 36.1 (15.5) | -- | -- |
| Mean dbh (cm) | 16.3 (1.6) | 14.0 (2.5) | 14.0 (1.0) | 34.9 (3.4) | 16.4 (1.4) | 12.6 (1.3) | 14.0 (2.0) | 35.6 (3.0) |
| Ponderosa pine | ||||||||
| Basal Area (m2/ha) | 5.6 (1.5) | 5.6 (1.4) | -- | -- | 7.1 (2.3) | 7.1 (2.3) | -- | -- |
| Density (tph) | 162 (31 ) | 160 (31) | -- | -- | 246 (94) | 246 (94) | -- | -- |
| % Basal Area | 24.8 (5.5) † | 63.0 (7.3) † | -- | -- | 32.9 (10.8) | 60.7 (15.0) | -- | -- |
| % Density | 20.4 (3.5) † | 60.6 (6.7) † | -- | -- | 31.1 (10.5) | 57.5 (15.0) | -- | -- |
| Mean dbh (cm) | 19.6 (1.0) | 18.9 (1.4) | -- | -- | 18.1 (1.8) | 18.1 (1.8) | -- | -- |
| Outbreak Effects Percent Douglas-fir mortality | ||||||||
| By basal area | 64.5 (8.4) * | 17.9 (7.9) | 14.0 (8.5) * | 14.4 (6.5) | ||||
| By density | 70.9 (15.4) * | 7.9 (4.6) | 15.4 (9.1) * | 6.2 (3.3) | ||||


4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- White, P.S.; Pickett, S.T.A. Natural disturbances and patch dynamics: An introduction. In The Ecology of Natural Disturbances and Patch Dynamics; White, P.S., Pickett, S.T.A., Eds.; Academic Press: San Diego, CA, USA, 1985; pp. 3–13. [Google Scholar]
- Seidl, R.; Fernandes, P.M.; Fonseca, T.F.; Gillet, F.; Jönssong, A.M.; Merganiˇcová, K.; Netherer, S.; Arpaci, A.; Bontemps, J.; Bugmann, H.; et al. Modelling natural disturbances in forest ecosystems: A review. Ecol. Model. 2011, 222, 903–924. [Google Scholar] [CrossRef]
- Knight, D.H. Parasites, lightning and the vegetation mosaic in wilderness landscapes. In Landscape Heterogeneity and Disturbance; Turner, M.G., Ed.; Springler-Verlag: New York, NY, USA, 1987; pp. 59–83. [Google Scholar]
- Peet, R.K. Forest vegetation of the Colorado Front Range. Vegetatio 1981, 45, 3–77. [Google Scholar] [CrossRef]
- Furniss, R.L.; Carolin, V.M. Western Forest Insects; Miscellaneous Publication No. 1339; U.S. Department of Agriculture Forest Service: Portland, OR, USA, 1977; p. 654.
- Tunnock, S. The Douglas-fir Tussock Moth in the Northern Region: A Cartographic History of Outbreaks from 1928 to 1973; Report No. 73–27; U.S. Department of Agriculture, Forest Service, Division of State and Private Forestry, Northern Region: Missoula, MT, USA, 1973.
- Wickman, B.E. Tree Mortality and Top-Kill Related to Defoliation by the Douglas-fir Tussock Moth in the Blue Mountains Outbreak; USDA Forest Service Research Paper PNW-233; Pacific Northwest Forest and Range Experiment Station, U.S. Department of Agriculture: Portland, OR, USA, 1978.
- Wickman, B.E. Mortality and Growth Reduction of White Fir Following Defoliation by the Douglas-fir Tussock Moth; USDA Forest Service Research Paper PSW-7; Pacific Southwest Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Berkeley, CA, USA, 1963.
- Wickman, B.E. Increased growth of white fir after a Douglas-fir tussock moth outbreak. J. For. 1980, 78, 31–33. [Google Scholar]
- Alfaro, R.I.; Shepherd, R.F. Tree-ring growth of interior Douglas-fir after one year’s defoliation by Douglas-fir tussock moth. For. Sci. 1991, 37, 959–964. [Google Scholar]
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph. In Great Basin Naturalist Memoirs, Number 6; Brigham Young University: Provo, UT, USA, 1982. [Google Scholar]
- Wright, L.C.; Berryman, A.A.; Wickman, B.E. Abundance of the fir engraver, Scolytus ventralis, and the Douglas-fir beetle, Dendroctonus pseudotsugae, following tree defoliation by the Douglas-fir tussock moth, Orgyia pseudotsugata. Can. Entomol. 1984, 116, 293–305. [Google Scholar] [CrossRef]
- Furniss, M.M. Susceptibility of fire-injured Douglas-fir to bark beetle attack in Southern Idaho. J. For. 1965, 63, 8–14. [Google Scholar]
- Rudinsky, J.A. Host selection and invasion by the Douglas-fir beetle, Dendroctonus pseudotsugae Hopkins, in coastal Douglas-fir forests. Can. Entomol. 1966, 98, 98–111. [Google Scholar] [CrossRef]
- Wright, K.H.; Lauterbach, P.G. Ten Year Study of Mortality in a Douglas-fir Saw-Timber Stand in Coos and Douglas Counties, Oregon; USDA Forest Service Research Paper PNW-27; U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1958.
- Furniss, M.M.; Livingston, R.L.; McGregor, M.D. Development of a stand susceptibility classification for Douglas-fir beetle. In Hazard-Rating Systems in Forest Insect Pest Management: Symposium Proceedings; Hedden, R.L., Barras, S.J., Coster, J.E., Eds.; General Technical Report WO-27; USDA Forest Service Washington Office: Washington, DC, USA, 1981; pp. 115–128. [Google Scholar]
- Lessard, E.D.; Schmid, J.M. Emergence, attack densities, and host relationships for the Douglas-fir beetle (Dendroctonus pseudotsugae Hopkins) in northern Colorado. Gt. Basin Nat. 1990, 50, 333–338. [Google Scholar]
- Johnson, N.E.; Belluschi, P.G. Host-finding behavior of the Douglas-fir beetle. J. For. 1969, 67, 290–295. [Google Scholar]
- Furniss, M.M.; McGregor, M.D.; Foiles, M.W.; Partridge, A.D. Chronology and Characteristics of a Douglas-fir Beetle Outbreak in Northern Idaho; USDA Forest Service General Technical Report INT-59; Intermountain Forest and Range Experiment Station, USDA Forest Service: Boise, ID, USA, 1979.
- Negron, J.F.; Schaupp, W.C., Jr.; Gigson, K.E.; Anhold, J.; Hansen, D.; Their, R.; Mocettini, P. Estimating extent of mortality associated with the Douglas-fir beetle in the Central and Southern Rockies. West. J. Appl. For. 1999, 14, 121–127. [Google Scholar]
- Powers, S.P.; Sollins, P.; Harmon, M.E.; Jones, J.A. Plant-pest interactions in time and space: A Douglas-fir bark beetle outbreak as a case study. Landsc. Ecol. 1999, 14, 105–120. [Google Scholar] [CrossRef]
- Negron, J.F. Probability of infestation and extent of mortality associated with the Douglas-fir beetle in the Colorado Front Range. For. Ecol. Manag. 1998, 107, 71–85. [Google Scholar] [CrossRef]
- Negron, J.F.; Schaupp, W.C., Jr.; Pederson, L. Flight Periodicity of the Douglas-fir beetle, Dendroctonus pseudotsugae Hopkins (Coleoptera: Curculionidae: Scolytinae) in Colorado, USA. Coleopt. Bull. 2011, 65, 182–184. [Google Scholar] [CrossRef]
- Schaupp, W.C.; Costello, S.L.; Ciesla, W.M. The History of Douglas-fir Tussock Moth in Colorado and Wyoming; Technical Report R2–67; Rocky Mountain Region, Renewable Resources, USDA Forest Service: Lakewood, CO, USA, 2008.
- Negron, J.F.; Schaupp, W.C.; Johnson, E. Development and validation of a fixed-precision sequential sampling plan for estimating brood adult density of Dendroctonus pseudotsugae (Coleoptera: Scolytidae). Can. Entomol. 2000, 132, 119–133. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Riging, A.; Breshears, D.D.; Hogg, E.H.; et al. A global review of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Johnston, B.C. Plant Associations of Region 2, Potential Plant Communities of Wyoming, South Dakota, Nebraska, and Kansas; R2-ECOL-87-2; Rocky Mountain Region, Range, Wildlife, Fisheries, and Ecology, USDA Forest Service: Lakewood, CO, USA, 1987.
- Lynch, A.M. Comparison of fixed-area plot designs for estimating stand characteristics and western spruce budworm damage in southwestern USA forests. Can. J. For. Res. 2003, 33, 1245–1255. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT® 9.2 User’s Guide, SAS Institute Inc: Cary, NC, USA, 2008.
- Mielke, P.W., Jr.; Berry, K.J. Permutation Methods: A Distance Function Approach; Springer-Verlag: New York, NY, USA, 2001; p. 427. [Google Scholar]
- Kaufmann, M.R.; Regan, C.M.; Brown, P.M. Heterogeneity in ponderosa pine/Douglas-fir forests: Age and size structure in unlogged and logged landscapes in central Colorado. Can. J. For. Res. 2000, 30, 698–711. [Google Scholar] [CrossRef]
- Stoszek, K.J.; Mika, P.G.; Moore, J.A.; Osborne, H.L. Relationships of Douglas-fir tussock moth defoliation to site and stand characteristics in northern Idaho. For. Sci. 1981, 27, 431–442. [Google Scholar]
- Coleman, T.W.; Jones, M.I.; Courtial, B.; Graves, A.D.; Woods, M.; Roques, A.; Seybold, S.J. Impact of the first recorded outbreak of the Douglas-fir tussock moth, Orgyia pseudotsugata, in southern California and the extent of its distribution in the Pacific Southwest region. For. Ecol. Manag. 2014, 329, 295–305. [Google Scholar] [CrossRef]
- Hadley, K.S.; Veblen, T.T. Stand response to western spruce budworm and Douglas-fir bark beetle outbreaks, Colorado Front Range. Can. J. For. Res. 1993, 23, 479–491. [Google Scholar] [CrossRef]
- Hadley, K.S. The role of disturbance, topography, and forest structure in the development of a montane forest landscape. B. Torrey Bot. Club 1994, 121, 47–61. [Google Scholar] [CrossRef]
- Shepherd, R.F.; Gray, T.G.; Chorney, R.J.; Daterman, G.E. Pest management of Douglas-fir tussock moth, Orgyia pseudotsugata (Lepidoptera: Lymantriidae): Monitoring endemic populations with pheromone traps to detect incipient outbreaks. Can. Entomol. 1985, 117, 839–848. [Google Scholar] [CrossRef]
- Mason, R.R. Dynamic behavior of Douglas-fir tussock moth populations in the Pacific Northwest. For. Sci. 1996, 42, 182–191. [Google Scholar]
- Mitchell, R.G. Dispersal of early instars of the Douglas-fir tussock moth. Ann. Entomol. Soc. Am. 1979, 72, 291–297. [Google Scholar]
- Negron, J.F.; Anhold, J.A.; Munson, A.S. Within-stand spatial distribution of tree mortality caused by the Douglas-Fir Beetle (Coleoptera: Scolytidae). Environ. Entomol. 2001, 30, 215–224. [Google Scholar] [CrossRef]
- Withrow, J.R.; Lundquist, J.E.; Negron, J.F. Spatial dispersal of Douglas-fir beetle populations in Colorado and Wyoming. ISRN For. 2013, 2013. [Google Scholar] [CrossRef]
- Schmid, J.M.; Mata, S.A. Natural Variability of Specific Forest Insect Populations and Their Associated Effects in Colorado; Technical Report RM-GTR-275; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 1996.
- Swetnam, T.W.; Lynch, A.M. A tree-ring reconstruction of western spruce budworm history in the Southern Rocky Mountains. For. Sci. 1989, 35, 962–986. [Google Scholar]
- Fellin, D.G.; Shearer, R.C.; Carlson, C.E. Western spruce budworm in the northern Rocky Mountains: Biology, ecology and impacts. West. Wildlands 1983, 9, 2–7. [Google Scholar]
- Allen, C.D. Historical fire regime patterns in the southwestern United States since AD 1700. In Fire Effects in Southwestern Forests, General Technical Report RM-GTR-286; In Proceedings of the 2nd La Mesa Fire Symposium; Los Alamos, NM, USA, 29–31 March 1994, Allen, C.D. USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 1996; pp. 11–32. [Google Scholar]
- Fulé, P.Z.; Covington, W.W.; Moore, M.M. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecol. Appl. 1997, 7, 895–908. [Google Scholar] [CrossRef]
- Mast, J.N.; Fulé, P.Z.; Moore, M.M.; Covington, W.W.; Waltz, A.E.M. Restoration of presettlement age structure of an Arizona ponderosa pine forest. Ecol. Appl. 1999, 9, 228–239. [Google Scholar] [CrossRef]
- Brown, P.M.; Kaufmann, M.R.; Shepperd, W.D. Long-term, landscape patterns of past fire events in a montane ponderosa pine forest of central Colorado. Landsc. Ecol. 1999, 4, 513–532. [Google Scholar] [CrossRef]
- Sherriff, R.L.; Veblen, T.T. A spatially-explicit reconstruction of historical fire occurrence in the ponderosa pine zone of the Colorado Front Range. Ecosystems 2007, 10, 311–323. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Donnegan, J. Climatic and human influences on fire regimes in ponderosa pine forests in the Colorado Front Range. Ecol. Appl. 2000, 10, 1178–1195. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Lynch, A.M. Multicentury, regional-scale patterns of western spruce budworm outbreaks. Ecol. Monogr. 1993, 63, 399–424. [Google Scholar] [CrossRef]
- Touchan, R.; Allen, C.D.; Swetnam, T.W. Fire history and climatic patterns in ponderosa pine and mixed-conifer forests of the Jemez Mountains, Northern New Mexico; USDA Forest Service GTR-RM-286. In Proceedings of the second La Mesa Fire symposium, USDA Forest Service GTR-RM-286. Los Alamos, NM, USA, 29–31 March 1996; pp. 33–46.
- Swetnam, T.W.; Allen, C.D.; Betancourt, J.L. Applied historical ecology: Using the past to manage for the future. Ecol. Appl. 1999, 9, 1189–1206. [Google Scholar] [CrossRef]
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T.; et al. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Lynch, A.M. What tree-ring reconstruction tells us about conifer defoliator outbreaks. In Insect Outbreaks Revisited; Barbosa, P., Letourneau, D.K., Agrawal, A.A., Eds.; Wiley-Blackwell: Chichester, UK, 2012; pp. 126–154. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Negrón, J.F.; Lynch, A.M.; Schaupp, W.C., Jr.; Mercado, J.E. Douglas-Fir Tussock Moth- and Douglas-Fir Beetle-Caused Mortality in a Ponderosa Pine/Douglas-Fir Forest in the Colorado Front Range, USA. Forests 2014, 5, 3131-3146. https://doi.org/10.3390/f5123131
Negrón JF, Lynch AM, Schaupp WC Jr., Mercado JE. Douglas-Fir Tussock Moth- and Douglas-Fir Beetle-Caused Mortality in a Ponderosa Pine/Douglas-Fir Forest in the Colorado Front Range, USA. Forests. 2014; 5(12):3131-3146. https://doi.org/10.3390/f5123131
Chicago/Turabian StyleNegrón, José F., Ann M. Lynch, Willis C. Schaupp, Jr., and Javier E. Mercado. 2014. "Douglas-Fir Tussock Moth- and Douglas-Fir Beetle-Caused Mortality in a Ponderosa Pine/Douglas-Fir Forest in the Colorado Front Range, USA" Forests 5, no. 12: 3131-3146. https://doi.org/10.3390/f5123131
APA StyleNegrón, J. F., Lynch, A. M., Schaupp, W. C., Jr., & Mercado, J. E. (2014). Douglas-Fir Tussock Moth- and Douglas-Fir Beetle-Caused Mortality in a Ponderosa Pine/Douglas-Fir Forest in the Colorado Front Range, USA. Forests, 5(12), 3131-3146. https://doi.org/10.3390/f5123131