Carbon Stocks and Climate Change: Management Implications in Northern Arizona Ponderosa Pine Forests

Abstract
:1. Introduction
- How might ponderosa pine stands respond to climate change at different elevations?
- Can management of ponderosa pine forests mitigate the negative effects of climate change on stand mortality and total carbon stocks?
2. Experimental Section
2.1. Data and Study Area


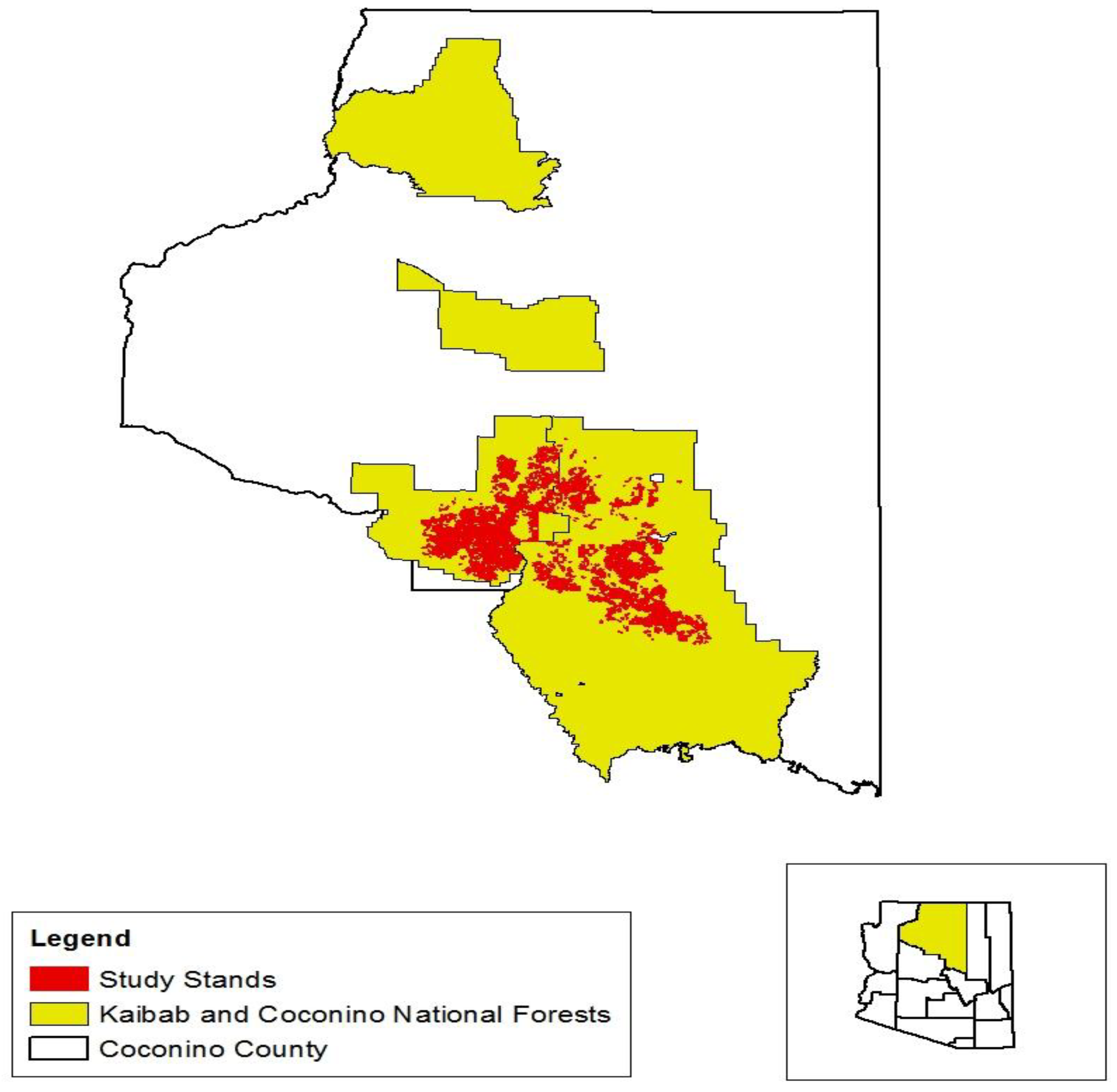{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation | TPH 1 | BA 2 | SDI 3 | QMD 4 | Can. Ht 5 | TI 6 | Live C 7 | Dead C 8 |
|---|---|---|---|---|---|---|---|---|
| Low 1950–2194 m | 510 | 36.7 | 366 | 14.0 | 3.5 | 57.5 | 91.0 | 12.6 |
| Mid 2194–2499 m | 338 | 28.0 | 259 | 19.1 | 4.7 | 135.8 | 68.8 | 10.1 |
| High 2499–2682 m | 213 | 42.5 | 363 | 21.3 | 4.4 | 58.6 | 105.8 | 15.0 |
| All Stands 1950–2682 m | 354 | 35.8 | 329 | 18.0 | 4.2 | 84.0 | 88.5 | 12.6 |
2.2. Representative Concentration Pathways
2.3. Climate Extension to the Forest Vegetation Simulator
2.4. Carbon Sub-Model of the Fire and Fuels Extension to the Forest Vegetation Simulator
2.5. Elevation
2.6. Management Scenarios
| Management Scenario | Initializing BA (m2/ha) | Residual Target BA (m2/ha) | Diameter Limit (Removal cm) | Treatment Frequency |
|---|---|---|---|---|
| Moderate | 37 | 28 | 41 cm | 20 years |
| Intensive | 28 | 18 | 61 cm | 10 years |
3. Results and Discussion
3.1. Baseline Conditions
| Trees Per Hectare | Basal Area (m2/ha) | Quadratic Mean Diameter (cm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Elevation | Low | Mid | High | Low | Mid | High | Low | Mid | High |
| Year | |||||||||
| 2010 | 510 | 338 | 212 | 37 | 28 | 42 | 14 | 19 | 21 |
| 2020 | 478 | 333 | 214 | 40 | 31 | 45 | 15 | 20 | 22 |
| 2030 | 462 | 324 | 224 | 43 | 35 | 48 | 15 | 22 | 22 |
| 2040 | 425 | 287 | 233 | 45 | 37 | 50 | 16 | 24 | 23 |
| 2050 | 393 | 255 | 227 | 47 | 39 | 52 | 17 | 25 | 24 |
| 2060 | 374 | 229 | 227 | 48 | 41 | 54 | 18 | 27 | 25 |
| 2070 | 347 | 202 | 224 | 50 | 42 | 55 | 19 | 29 | 25 |
| 2080 | 333 | 177 | 207 | 51 | 43 | 56 | 19 | 30 | 26 |
| 2090 | 313 | 156 | 194 | 52 | 43 | 57 | 20 | 32 | 27 |
| 2100 | 296 | 139 | 177 | 53 | 43 | 58 | 21 | 34 | 29 |
| 2110 | 284 | 125 | 162 | 53 | 43 | 59 | 22 | 36 | 30 |
3.2. Stand Dynamics in a Changing Climate without Management
3.2.1. Effects of Climate Change in Lower Elevation Stands
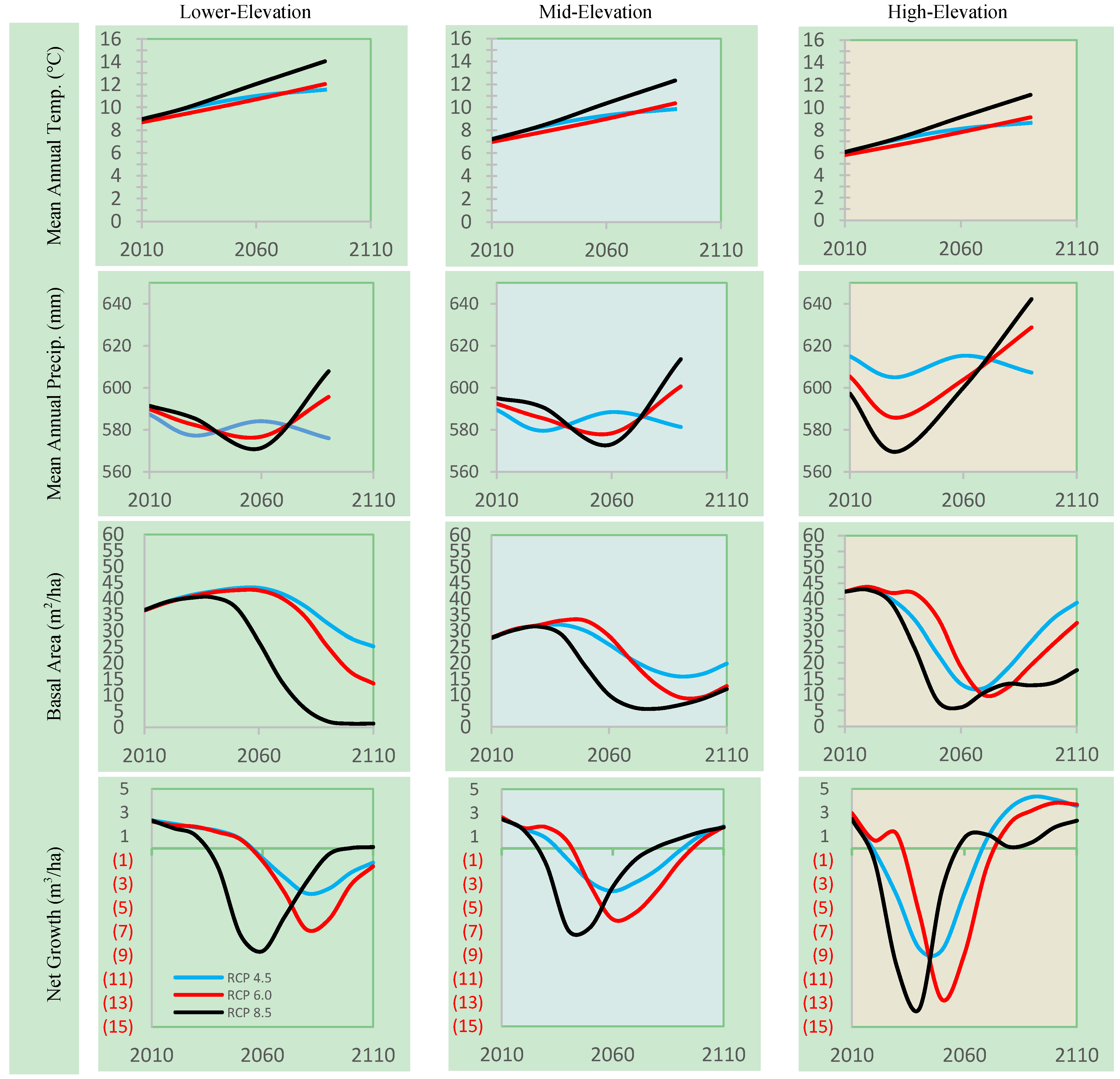
3.2.2. Effects of Climate Change on Mid-Elevation Stands
3.2.3. Effects of Climate Change on High-Elevation Stands
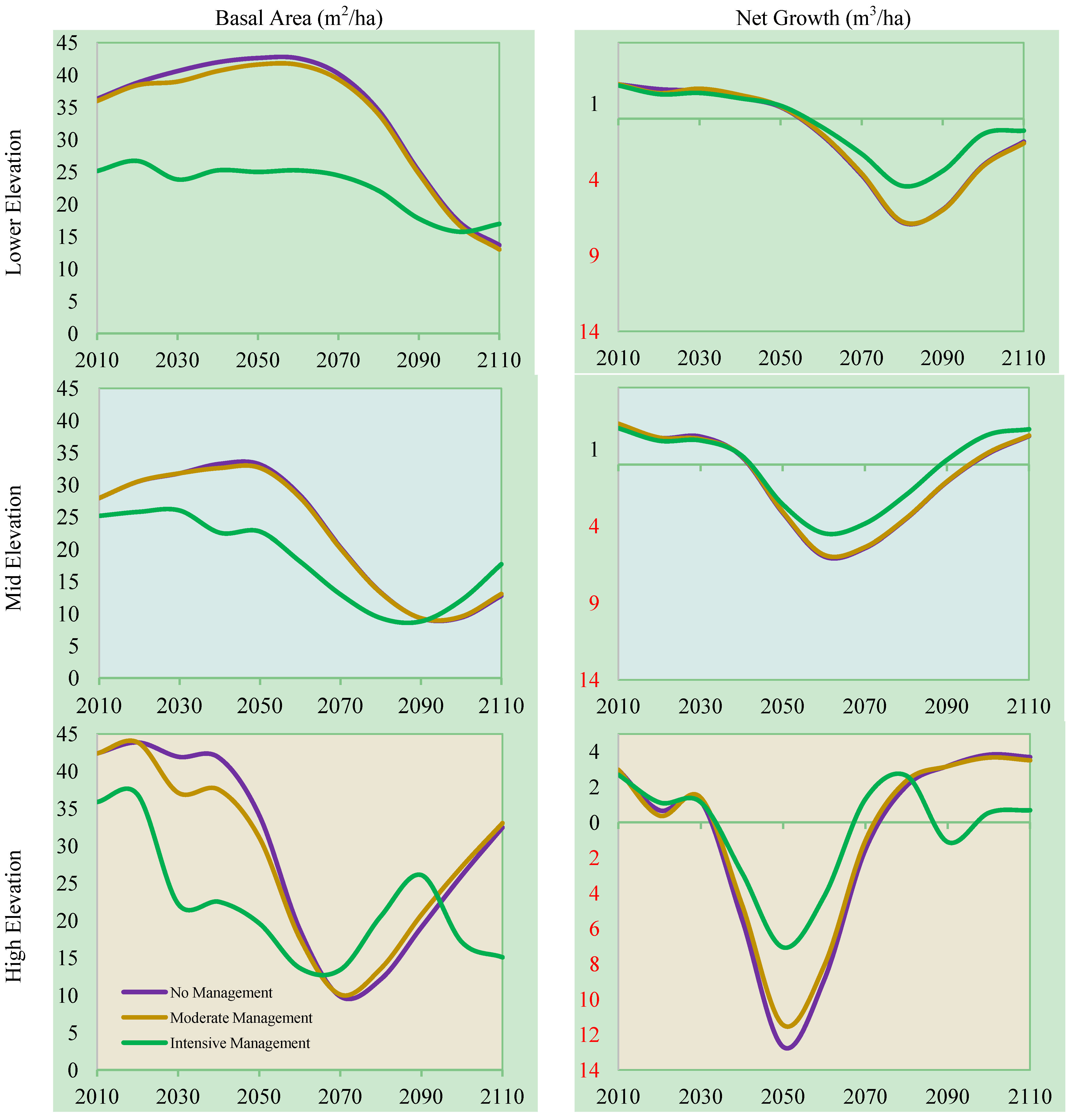
3.3. Management, Elevation, and Climate Change
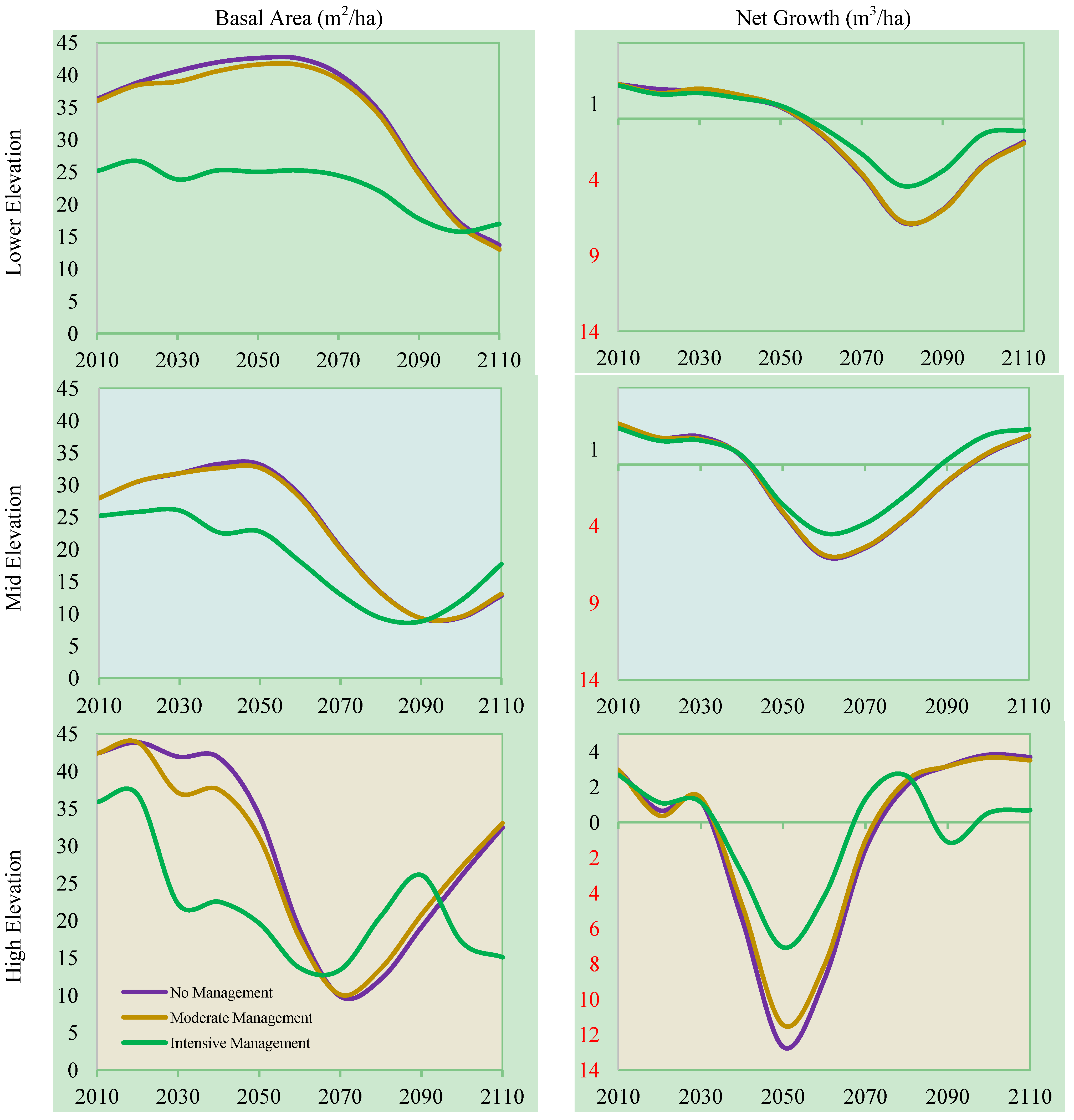
3.4. Carbon Storage, Management, and Climate Change

3.5. Broader Implications of Results
3.6. Study Limitations
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Breshears, D.D. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. 2005, 102, 15144–15148. [Google Scholar] [CrossRef]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. 2008, 105, 11823–11826. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Ferguson, D.E.; Crookston, N.L. Aspen, climate, and sudden decline in western USA. For. Ecol. Manag. 2009, 258, 2353–2364. [Google Scholar] [CrossRef]
- Williams, A.P.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D.; et al. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang. 2012, 3, 292–298. [Google Scholar] [CrossRef]
- Brusca, R.C.; Wiens, J.F.; Meyer, W.M.; Eble, J.; Franklin, K.; Overpeck, J.T.; Moore, W. Dramatic response to climate change in the Southwest: Robert Whittaker’s 1963 Arizona Mountain plant transect revisited. Ecol. Evol. 2013, 3, 3307–3319. [Google Scholar]
- Swetnam, T.W.; Betancourt, J.L. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. J. Clim. 1997, 11, 3128–3147. [Google Scholar] [CrossRef]
- Christensen, J.H.; Hewitson, B.; Busuioc, A.; Chen, A.; Gao, X.; Held, I.; Jones, R.; Kolli, R.K.; Kwon, W.T.; Laprise, R.; et al. Regional Climate Projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 847–940. [Google Scholar]
- Grissino-Mayer, H.D.; Swetnam, T.W. Century-scale climate forcing of fire regimes in the American Southwest. Holocene 2000, 10, 213–229. [Google Scholar]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western US forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negron, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: direct and indirect effects. Biosci. 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fulé, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; Veblen, T.T. Widespread increase in tree mortality rates in the Western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef]
- Breshears, D.D.; Allen, C.D. The importance of rapid, disturbance-induced losses in carbon management and sequestration. Glob. Ecol. Biogeog. 2002, 11, 1–5. [Google Scholar] [CrossRef]
- Schubert, G.H. Silviculture of Southwestern Ponderosa pine: The Status of our Knowledge; General Technical Report RMRS-GTR-123; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 1974; p. 7. [Google Scholar]
- Merriam, C.H. Results of a Biological Survey on the San Francisco Mountain Region and Desert of the Little Colorado, Arizona; North. Am.; Fauna No. 3; U.S. Department of Agriculture: Washington, DC, USA, 1890; pp. 7–13. [Google Scholar]
- Ogle, K.; Whitham, T.G.; Cobb, N.S. Tree-ring variation in pinyon pine predicts likelihood of death following severe drought. Ecology 2000, 81, 3237–3243. [Google Scholar] [CrossRef]
- Nevo, E. Evolution of genome-phenome diversity under environmental stress. Proc. Natl. Acad. Sci. 2001, 98, 6233–6240. [Google Scholar] [CrossRef]
- Rehfeldt, G.E. A spline model of climate for the Western United States; Genetic. Techicle. Report. RMRS-GTR-165; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2006; pp. 1–20. [Google Scholar]
- Gray, S.T.; Betancourt, J.L.; Jackson, S.T.; Eddy, R.G. Role of multidecadal climate variability in a range extension of pinyon pine. Ecology 2006, 87, 1124–1130. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; Yepez, E.A. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Gitlin, A.R.; Sthultz, C.M.; Bowker, M.A.; Stumpf, S.; Paxton, K.L.; Kennedy, K.; Muňoz, A.; Bailey, J.K.; Whitham, T.G. Mortality gradients within and among dominant plant populations as barometers of ecosystem change during extreme drought. Cons. Biol. 2006, 5, 1477–1486. [Google Scholar]
- Bowker, M.A.; Muňoz, A.; Martinez, T.; Lau, M.K. Rare drought-induced mortality of juniper is enhanced by edaphic stressors and influenced by stand density. J. Arid Environ. 2012, 76, 9–16. [Google Scholar] [CrossRef]
- NASA Earth Observatory. Earth’s Energy Balance. Available online: http://earthobservatory.nasa.gov/Features/EnergyBalance/page4.php (accessed on 16 December 2013).
- Le Treut, H.; Somerville, R.; Cubasch, U.; Ding, Y.; Mauritzen, C.; Mokssit, A.; Peterson, T.; Prather, M. Historical Overview of Climate Change. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 93–127. [Google Scholar]
- Meehl, G.A.; Stocker, T.F.; Collins, W.D.; Friedlingstein, P.; Gaye, A.T.; Gregory, J.M.; Kitoh, A.; Knutti, R.; Murphy, J.M.; Noda, A.; et al. Climate Projections. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 747–846. [Google Scholar]
- Allen, C.D.; Breshears, D.D. Drought induced shift of a forest-woodland ecotones: Rapid landscape response to climate variation. Proc. Natl. Acad. Sci. 1998, 95, 14389–14842. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Hughes, L.; Poulsen, M. Predicting species distributions: Use of climatic parameters in Bioclim and its impact on predictions of species’ current and future distributions. Ecol. Model. 2005, 186, 250–269. [Google Scholar]
- Rehfeldt, G.E.; Crookston, N.L.; Warwell, M.V.; Evans, J.S. Empirical analyses of plant-climate relationships for the Western United-States. Int. J. Plant Sci. 2006, 167, 1123–1150. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C. Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology. 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Crimmins, S.M.; Dobrowski, S.Z.; Greenberg, J.A.; Abatzoglou, J.T.; Mynsberge, A.R. Changes in climatic water balance drive downhill shifts in plant species’ optimum elevations. Science. 2011, 331, 324–326. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Fulé, P.Z.; Huffman, D.W.; Crouse, J.; Laliberté, E. Climatic constraints on trait-based forest assembly. J. Ecol. 2011, 99, 1489–1499. [Google Scholar] [CrossRef]
- Covington, W.W.; Moore, M.M. Post-settlement changes in natural fire regimes and forest structure: Ecological restoration of old-growth ponderosa pine forests. J. Sust. For. 1994, 2, 153–181. [Google Scholar] [CrossRef]
- Covington, W.W.; Fulé, P.Z.; Moore, M.M.; Hart, S.C.; Kolb, T.E.; Mast, J.N.; Sackett, S.S.; Wagner, M.R. Restoring ecosystem health in ponderosa pine forests of the southwest. J. For. 1997, 95, 23–29. [Google Scholar]
- Reynolds, R.T.; Sánchez-Meador, A.J.; Youzt, J.A.; Nicolet, T.; Matonis, M.S.; Jackson, P.L.; DeLorenzo, D.G.; Graves, A.D. Restoring composition and structure in Southwestern frequent-fire forests: A science-based framework for improving ecosystem resiliency; Genetic. Technical. Report. RMRS-GTR-310; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; p. 76. [Google Scholar]
- Finkral, A.J.; Evans, A.M. The effects of a thinning treatment on carbon stocks in a northern Arizona ponderosa pine forest. For. Ecol. Manag. 2008, 255, 2743–2750. [Google Scholar] [CrossRef]
- Wiedinmyer, C.; Hurteau, M.D. Prescribed fire as a means of reducing forest carbon emissions in the Western United States. Environ. Sci. Technol. 2010, 44, 1926–1932. [Google Scholar] [CrossRef]
- Hurteau, M.D.; Koch, G.W.; Hungate, B.A. Carbon protection and fire risk reduction: Toward a full accounting of forest carbon offsets. Fron. Ecol. Envir. 2008, 6, 493–498. [Google Scholar] [CrossRef]
- Vegh, T.; Huang, C.H.; Finkral, A.J. Carbon credit possibilities and economic implications of fuel reduction treatements. West. J. Appl. For. 2013, 28, 57–65. [Google Scholar] [CrossRef]
- Sorensen, C.; Finkral, A.J.; Kolb, T.E.; Huang, C.H. Short- and long-term effects of thinning and prescribed fire on carbon stocks in ponderosa pine stands in Northern Arizona. For. Ecol. Manag. 2011, 261, 460–472. [Google Scholar] [CrossRef]
- Huang, C.H.; Finkral, A.; Sorensen, C.; Kolb, T. Toward full economic valuation of forest fuels-reduction treatments. J. Environ. Manag. 2013, 130, 221–231. [Google Scholar] [CrossRef]
- Mason, C.L.; Lippke, B.R.; Zobrist, K.W.; Blaxton, T.D., Jr.; Ceder, K.R.; Comnick, J.M.; McCarter, J.B.; Rogers, H.K. Investments in fuel removals to avoid forest fires result in substantial benefits. J. For. 2006, 104, 27–31. [Google Scholar]
- Snider, G.; Daugherty, P.; Wood, D. Irrationality of continued fire suppression: an avoided cost analysis of fire hazard reduction treatments versus no treatment. J. For. 2006, 104, 431–437. [Google Scholar]
- Fulé, P.Z. Does it make sense to restore wildland fire in changing climate? Restor. Ecol. 2008, 16, 526–531. [Google Scholar] [CrossRef]
- Crookston, N.L.; Rehfeldt, G.E.; Dixon, G.E.; Weiskittel, A.R. Addressing climate change in the forest vegetation simulator to assess impacts on landscape forest dynamics. For. Ecol. Manag. 2010, 260, 1198–1211. [Google Scholar] [CrossRef]
- United States Department of Agriculture Collaborative Forest Landscape Restoration Program. Available online: http://www.fs.fed.us/restoration/CFLRP/ (accessed on 16 December 2013).
- National Climatic Data Center. Annual Climatological Summary: 1979–2009, Flagstaff Pulliam Airport, AZ. Dataset available upon request to corresponding author.
- Torching Index (TI) is the 20 foot wind speed necessary, in miles-per-hour, for a surface fire to ignite a crown fire [46]. Torching index is calculated in the Fire and Fuels Extension to the Forest Vegetation Simulator (FFE-FVS).
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.; et al. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Crookston, N.L. Climate-FVS Version 2: Content, User’s Guide, Applications, and Behavior. (Review Draft). 2013. Available online: http://www.fs.fed.us/fmsc/ftp/fvs/docs/gtr/ClimateFVS_UsersGuide.pdf. (accessed on 11 December 2013).
- Dixon, G.E. Essential FVS: A User’s Guide to the Forest Vegetation Simulator; Internal Report, comp. (Revised: November 19, 2013); Department of Agriculture, Forest Service, Forest Management Service Center: Fort Collins, CO, USA, 2002; p. 226. [Google Scholar]
- Daniels, A.E.; Morrison, J.F.; Joyce, L.A.; Crookston, N.L.; Chen, S.C.; McNulty, S.G. Climate projections FAQ; Genetic Techicle Report RMRS-GTR-277WWW; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2012; p. 32. [Google Scholar]
- Breiman, L. Random forests. Machine Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and regression by random forest. R News 2002, 2, 18–22. [Google Scholar]
- Rebain, S.A. The Fire and Fuels Extension to the Forest Vegetation Simulator: Updated Model Documentation; Internal Report; Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2013; p. 408. [Google Scholar]
- Veblen, T.T.; Kitzberger, T.; Donnegan, J. Climatic and human influences on fire regimes in ponderosa pine forests in the Colorado Front Range. Ecol. Appl. 2000, 10, 1178–1195. [Google Scholar] [CrossRef]
- Shorter fire-return intervals in lower-elevation ponderosa pine stands are characteristic of the historic fire regimes, i.e., pre-European settlement, before widespread fire-suppression policies were implemented in the Western United States.
- McCusker, N. Four-Forest Restoration Initiative Coconino and Kaibab National Forests Silviculture Specialist Report; US Forest Service: Flagstaff, AZ, USA, 2013. Available online: https://fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5413017.pdf (accessed on 16 December 2013).
- U.S. Forest Service Four Forest Restoration Initiative. Draft Environmental Impact Statement for the Four-Forest Restoration Initiative; U.S. Department of Agriculture, Forest Service, Southwest Region: Fort Collins, CO, USA, February 2013; p. 716. Available online: http://www.4fri.org/pdfs/documents/NEPA/draft-eis-feb2013.pdf (accessed on 12 December 2013).
- Four Forest Restoration Initiative Stakeholders Group. Old-Growth Protection and Large Tree Retention Strategy. Comp; Four Forest Restoration Initiative: Flagstaff, AZ, USA, 2011. (revised August 2012). Available online: http://www.4fri.org/pdfs/documents/collaboration/old_growth_protection-revised080812.pdf (accessed on 30 November 2013).
- Fulé, P.Z.; Covington, W.W.; Moore, M.M. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecol. Appl. 1997, 7, 895–908. [Google Scholar] [CrossRef]
- Fulé, P.Z.; Crouse, J.E.; Roccaforte, J.P.; Kalies, E.L. Do thinning and/or burning treatments in western USA ponderosa or Jeffrey pine-dominated forests help restore natural fire behavior? For. Ecol. Manag. 2012, 269, 68–81. [Google Scholar] [CrossRef]
- DeGomez, T. Guidelines for thinning ponderosa pine for improved forest health and fire prevention. Cooperative Extension: The University of Arizona College of Agriculture and Life Sciences; Publication AZ: Tucson, AZ, USA, 1397. Available online: http://cals.arizona.edu/pubs/natresources/az1397.pdf (accessed on 24 October 2013).
- Bailey, J.D.; Covington, W.W. Evaluating ponderosa pine regeneration rates following ecological restoration treatments in northern Arizona, USA. For. Ecol. Manag. 2002, 155, 271–278. [Google Scholar] [CrossRef]
- The FVS has calibrated its tree-growth equations using data primarily collected during the 20th Century [50]. The climate in North America during this period of time was relatively constant, up until the 1990s when increases in MAT became more apparent [7]. Therefore, running simulations into the 21st Century without the Climate Extension to the FVS outputs data derived from equations that assume tree-growth will not be affected by climate change.
- Crown competition factor is a measure of area occupied by each tree’s canopy [50]. A tree’s crown competition factor is the percentage of an acre the tree’s crown would occupy if that tree were to be grown in open conditions. A crown competition factor of 100 means that all tree-crowns would just touch. This metric is a measure of stand density and competition between trees.
- Hurteau, M.; North, M. Fuel treatment effects on tree-based forest carbon storage and emissions under modeled wildfire scenarios. Front. Ecol. Environ. 2009, 7, 409–414. [Google Scholar] [CrossRef]
- Soulé, P.T.; Knapp, P.A. Radial growth rate increases in naturally occurring ponderosa pine trees: A late-20th century CO2 fertilization effect? New Phytol. 2006, 171, 379–390. [Google Scholar] [CrossRef]
- McDowell, N.; Allen, C.D.; Marshall, L. Growth, carbon-isotope discrimination, and drought-associated mortality across a Pinus ponderosa elevational transect. Glob. Chang. Biol. 2010, 16, 399–415. [Google Scholar] [CrossRef]
- Hurteau, M.D.; Stoddard, M.T.; Fulé, P.Z. The carbon costs of mitigating high-severity wildfire in southwestern ponderosa pine. Glob. Chang. Biol. 2011, 17, 1516–1521. [Google Scholar] [CrossRef]
- Keith, H.; Mackey, B.G.; Lindenmayer, D.B. Re-evaluation of forest biomass carbon stocks and lessons from the world’s most carbon-dense forests. Proc. Natl. Acad. Sci. 2009, 11635–11640. [Google Scholar]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. Chapter 1: Silviculture and its place in forestry. In The Practice of Silviculture: Applied Ecology, 9th ed.; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bagdon, B.; Huang, C.-H. Carbon Stocks and Climate Change: Management Implications in Northern Arizona Ponderosa Pine Forests. Forests 2014, 5, 620-642. https://doi.org/10.3390/f5040620
Bagdon B, Huang C-H. Carbon Stocks and Climate Change: Management Implications in Northern Arizona Ponderosa Pine Forests. Forests. 2014; 5(4):620-642. https://doi.org/10.3390/f5040620
Chicago/Turabian StyleBagdon, Benjamin, and Ching-Hsun Huang. 2014. "Carbon Stocks and Climate Change: Management Implications in Northern Arizona Ponderosa Pine Forests" Forests 5, no. 4: 620-642. https://doi.org/10.3390/f5040620