
Effect of Chitosan on Disease Control and Growth of Scots Pine (Pinus sylvestris L.) in a Forest Nursery

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Filed Experiment, Application of Chitosan and Fungicide
2.3. Seedling Inventory and Growth Measurement
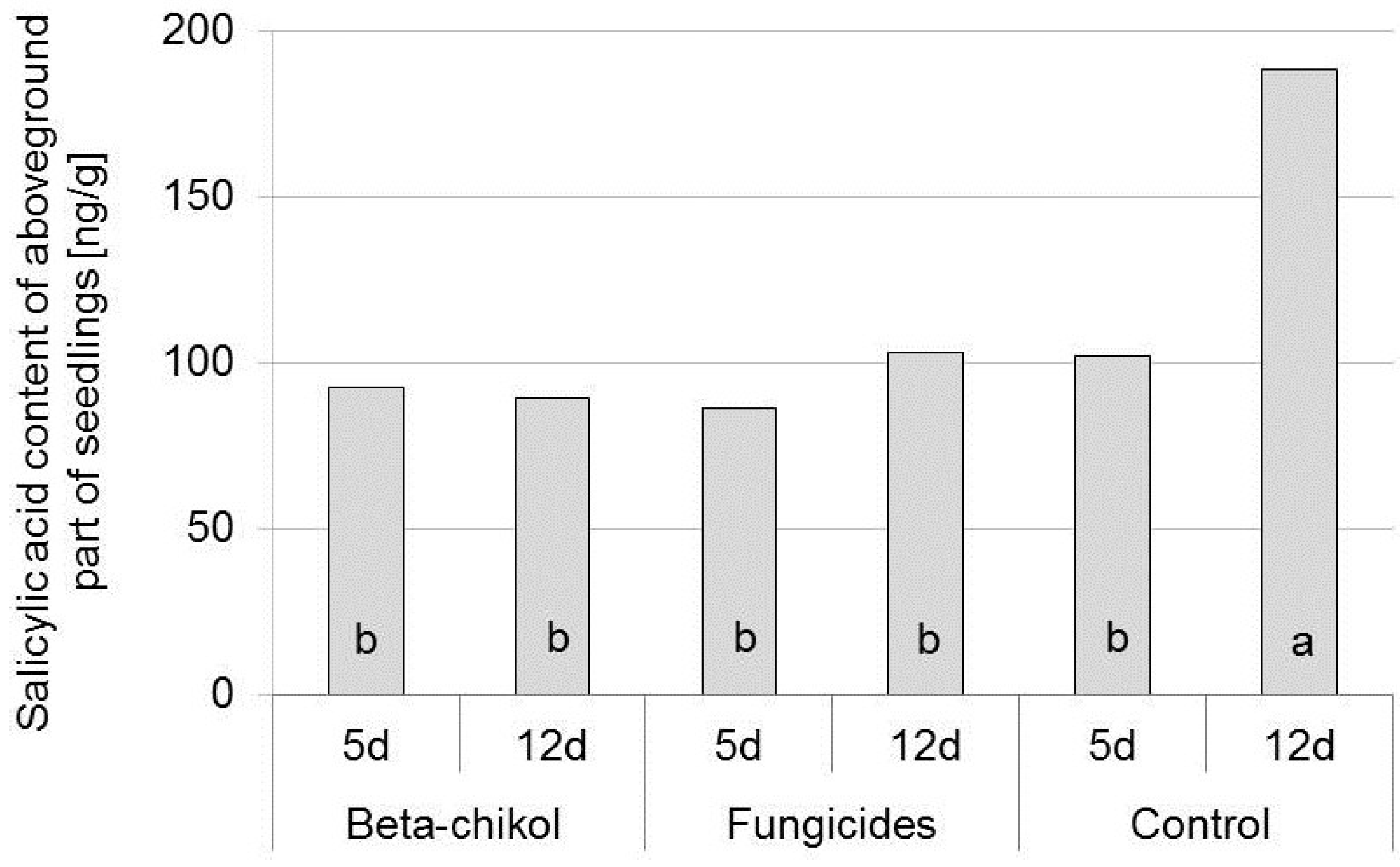
2.4. Salicylic Acid Content
2.5. Statistical Analyses
3. Results


{kind=link}
{kind=link}
| Treatment | Shoot Length (cm) | Root Collar Diameter (mm) | Aboveground Dry Mass (g) | Root Dry Mass (g) | No. of Lateral Shoots | Root Length (cm) | No. of Root Tips |
|---|---|---|---|---|---|---|---|
| Beta‑chikol | 9.3 a (1.9) | 2.2 a (0.4) | 0.737 a (0.287) | 0.318 a (0.138) | 3.9 a (1.7) | 332.7 a (107.5) | 795 a (222) |
| Fungicides | 8.1 b (1.8) | 1.8 b (0.4) | 0.515 b (0.261) | 0.183 b (0.091) | 3.2 b (2.5) | 178.4 b (85.1) | 424 b (182) |
| Control | 7.6 b (1.5) | 1.6 b (0.4) | 0.455 b (0.208) | 0.169 b (0.078) | 2.3 b (2.3) | 192.0 b (89.7) | 486 b (175) |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thakur, V.K.; Kesster, M.R. Self-healing polymer nanocomposite materials: A review. Polymer 2015, 69, 369–383. [Google Scholar] [CrossRef]
- Thakur, V.K.; Thakur, M.K. Recent advances in green hydrogels from lignin: A review. Int. J. Biol. Macromol. 2015, 72, 834–847. [Google Scholar] [CrossRef] [PubMed]
- Pappu, A.; Patil, V.; Jain, S.; Mahindrakar, A.; Haque, R.; Thakur, V.K. Advances in industrial prospective of cellulosic macromolecules enriched banana biofibre resources: A review. Int. J. Biol. Macromol. 2015, 79, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.K.; Thakur, M.K.; Gupta, R.K. Review: Raw natural fiber-based polymer composites. Int. J. Polym. Anal. Charact. 2014, 19, 256–271. [Google Scholar] [CrossRef]
- Thakur, V.K.; Thakur, M.K. Processing and characterization of natural cellulose fibers/thermoset polymer composites. Carbohydr. Polym. 2014, 109, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.K.; Thakur, M.K. Recent advances in graft copolymerization and applications of chitosan: A review. ACS Sustain. Chem. Eng. 2014, 2, 2637–2652. [Google Scholar] [CrossRef]
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B 2011, 366, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Raafat, D.; Sahl, H.-G. Chitosan and its antimicrobial potential—A critical literature survey. Microb. Biotechnol. 2009, 2, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, G.; Niranjan Raj, S.; Prashanth, G.N.; Deepak, S.; Amruthesh, K.N.; Shetty, H.S. Nitric oxide is involved in chitosan-induced systemic resistance in pearl millet against downy mildew disease. Pest Manag. Sci. 2009, 65, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Zuppini, A.; Baldan, B.; Millioni, R.; Favaron, F.; Navazio, L.; Mariani, P. Chitosan induces Ca2+-mediated programmed cell death in soybean cells. New Phytol. 2003, 161, 557–568. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Balasubramanian, R. Chitosan induce resistance components in Arachis hypogaea against leaf rust caused by Puccinia arachidis Speg. Crop Prot. 1998, 17, 307–313. [Google Scholar] [CrossRef]
- Reddy, M.V.B.; Arul, J.; Angers, P.; Couture, L. Chitosan treatment of wheat seeds induces resistance to Fusarium graminearun and improves seed quality. J. Agric. Food Chem. 1999, 47, 1208–1216. [Google Scholar] [CrossRef]
- Fajardo, J.E.; Waniska, R.D.; Cuero, R.G.; Pettit, R.E. Phenolic compounds in peanut seeds: Enhanced elicitation by chitosan and effects on growth and aflatoxin B1 production by Aspergillus flavus. Food Biotechnol. 1995, 9, 59–78. [Google Scholar] [CrossRef]
- Pospieszny, H.; Chirkov, S.; Atabekov, J. Induction of antiviral resistance in plants by chitosan. Plant Sci. 1991, 79, 63–68. [Google Scholar] [CrossRef]
- Li, B.; Liu, B.; Shan, C.; Ibrahim, M.; Lou, Y.; Wang, Y.; Xie, G.; Li, H.; Sun, G. Antibacterial activity of two chitosan solutions and their effect on rice bacterial leaf blight and leaf streak. Pest Manag. Sci. 2013, 69, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, P.; Benhamou, N.; Bussières, G.; Dessureault, M. Differential effects of chitosan on root rot fungal pathogens in forest nurseries. Can. J. Bot. 2000, 77, 1460–1468. [Google Scholar] [CrossRef]
- Hernández-Lauzardo, A.N.; Vega-Pérez, J.; Velázquez-Del Valle, M.G.; Sánchez, N.S.; Peña, A.; Guerra-Sánchez, G. Changes in the functionality of plasma membrane of Rhizopus stolonifer by addition of chitosan. J. Phytopathol. 2011, 159, 563–568. [Google Scholar] [CrossRef]
- Cho, M.H.; No, H.K.; Prinyawiwatkul, W. Chitosan treatments affect growth and selected quality of sunflower sprouts. J. Food Sci. 2008, 73, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Manjunatha, G.; Roopa, K.S.; Prashanth, G.N.; Shetty, H.S. Chitosan enhances disease resistance in pearl millet against downy mildew caused by Sclerospora graminicola and defence-related enzyme activation. Pest Manag. Sci. 2008, 64, 1250–1257. [Google Scholar] [CrossRef] [PubMed]
- Sharp, R.G. A review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 2013, 3, 757–793. [Google Scholar] [CrossRef]
- Raskin, I.; Skubatz, H.; Tang, W.; Meeuse, B.J.D. Salicylic acid levels in thermogenic and non-thermogenic plants. Ann. Bot. 1990, 66, 369–373. [Google Scholar]
- Malamy, J.; Carr, J.P.; Klessig, D.F.; Raskin, I. Salicylic acid: A likely endogenous signal in the resistance response of tobacco to viral infection. Science 1990, 250, 1002–1004. [Google Scholar] [CrossRef] [PubMed]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Janda, T.; Szalai, G.; Tari, I.; Páldi, E. Hydroponic treatment with salicylic acid decreases the effects of chilling injury in maize (Zea mays L.) plants. Planta 1999, 208, 175–180. [Google Scholar] [CrossRef]
- Kaydan, D.; Yagmur, M.; Okut, N. Effects of salicylic acid on the growth and some physiological characters in salt stressed wheat (Triticum aestivum L.). Tarim Bilim. Derg. 2007, 13, 114–119. [Google Scholar]
- Żurek, A.; Grzyś, E. Wpływ kwasu salicylowego na parametry fizjologiczne siewek ogórka traktowanych herbicydem Reglone. Post. Ochr. Roślin 2007, 47, 325–329, (In Polish with English summary). [Google Scholar]
- Matysiak, K.; Adamczewski, K. Regulatory wzrostu i rozwoju roślin—Kierunki badań w Polsce i na świecie. Post. Ochr. Roślin 2009, 49, 1810–1816, (In Polish with English summary). [Google Scholar]
- Handro, W.; Mello, C.M.; Manzano, M.A.; Floh, E.I.S. Enhancement of stem elongation and flower bud regeneration by salicylic acid. Rev. Bras. Fisiol. Veg. 1997, 9, 139–142. [Google Scholar]
- Hadwiger, L.A. Multiple effects of chitosan on plant system: Solid science or hype. Plant Sci. 2013, 208, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Forest Management Plan 2012–2021, Spała Forest District: Spała, Poland, Unpublished work. 2012. (In Polish)
- Głowacka, B. Środki Ochrony Roślin Oraz Produkty do Rozkładu Pni Drzew Leśnych Zalecane do Stosowania w Leśnictwie w Roku 2013; IBL: Sękocin Stary, Poland, 2012; pp. 1–72. [Google Scholar]
- Ziani, K.; Ursúa, B.; Maté, J.I. Application of bioactive coatings based on chitosan for artichoke seed protection. Crop Prot. 2010, 29, 853–859. [Google Scholar] [CrossRef]
- Guan, Y.J.; Hu, J.; Wang, X.J.; Shao, C.X. Seed priming with chitosan improves miaze germination and seedling growth in relation to physiological changes under low temperature stress. J. Zhejiang Univ.—Sci. B 2009, 10, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Duda, B.; Oszako, T.; Piwnicki, J. Possibilities of chitosan use in forestry. Bull. Pol. Acad. Sci. Biol. Sci. 2003, 51, 213–220. [Google Scholar]
- Patkowska, E.; Pięta, D.; Pastucha, A. The Effect of Biochikol 020 PC on Microorganism Communities in the Rhizosphere of Fabaceae Plants; Polish Chitin Society: Łódź, Poland, 2006; pp. 171–178. [Google Scholar]
- Harman, G.E. Multifunctional fungal plant symbionts: New tools to enhance plant growth and productivity. New Phytol. 2011, 189, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Viswanathan, R.; Raguchander, T.; Prakasam, V.; Samiyappan, R. Induction of systemic resistance by plant growth promoting rhizobacteria in crop plants against pests and diseases. Crop Prot. 2001, 20, 1–11. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Jansson, H.-B.; Salinas, J.; Lopez-Llorca, L.V. Effect of chitosan on hyphal growth and spore germination of plant pathogenic and biocontrol fungi. J. Appl. Microbiol. 2008, 104, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Bornet, A.; Teissedre, P.L. Chitosan, chitin-glucan and chitin effects on minerals (iron, lead, cadmium) and organic (ochratoxin A) contaminants in wines. Eur. Food Res. Technol. 2008, 226, 681–689. [Google Scholar] [CrossRef]
- Aleksandrowicz-Trzcińska, M.; Hamera-Dzierżanowska, A.; Żybura, H.; Drozdowski, S. Wpływ mykoryzacji i chitozanu na wzrost sosny zwyczajnej (Pinus sylvestris L.) w szkółce i na uprawie. Sylwan 2013, 157, 899–908, (In Polish with English summary). [Google Scholar]
- Ohta, K.; Morishita, S.; Suda, K.; Kobayashi, N.; Hosoki, T. Effects of chitosan soil mixture treatment in the seedling stage on the growth and flowering of several ornamental plants. J. Jap. Soc. Hortic. Sci. 2004, 73, 66–68. [Google Scholar] [CrossRef]
- Ohta, K.; Taniguchi, A.; Konishi, N.; Hosoki, T. Chitosan treatment affects plant growth and flower quality in Eustoma grandiflorum. HortScience 1999, 34, 233–234. [Google Scholar]
- Ramírez, M.Á.; Rodriguez, A.T.; Alfonso, L.; Peniche, C. Chitin and its derivatives as biopolymers with potential agricultural applications. Biotecnol. Apl. 2010, 27, 270–276. [Google Scholar]
- Ravi Kumar, M.N.V. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleksandrowicz-Trzcińska, M.; Bogusiewicz, A.; Szkop, M.; Drozdowski, S. Effect of Chitosan on Disease Control and Growth of Scots Pine (Pinus sylvestris L.) in a Forest Nursery. Forests 2015, 6, 3165-3176. https://doi.org/10.3390/f6093165
Aleksandrowicz-Trzcińska M, Bogusiewicz A, Szkop M, Drozdowski S. Effect of Chitosan on Disease Control and Growth of Scots Pine (Pinus sylvestris L.) in a Forest Nursery. Forests. 2015; 6(9):3165-3176. https://doi.org/10.3390/f6093165
Chicago/Turabian StyleAleksandrowicz-Trzcińska, Marta, Anna Bogusiewicz, Michał Szkop, and Stanisław Drozdowski. 2015. "Effect of Chitosan on Disease Control and Growth of Scots Pine (Pinus sylvestris L.) in a Forest Nursery" Forests 6, no. 9: 3165-3176. https://doi.org/10.3390/f6093165
APA StyleAleksandrowicz-Trzcińska, M., Bogusiewicz, A., Szkop, M., & Drozdowski, S. (2015). Effect of Chitosan on Disease Control and Growth of Scots Pine (Pinus sylvestris L.) in a Forest Nursery. Forests, 6(9), 3165-3176. https://doi.org/10.3390/f6093165