
The Alleviation of Nutrient Deficiency Symptoms in Changbai Larch (Larix olgensis) Seedlings by the Application of Exogenous Organic Acids


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Culture and Treatments
2.2. Measurements of Biomass
2.3. Nutrient Element Analysis
2.4. Physiological Parameter Measurements
2.5. Data Analysis
3. Results
3.1. Survival, Biomass, and Nutrient Element Contents of L. olgensis Seedlings
3.2. Lipid Peroxidation
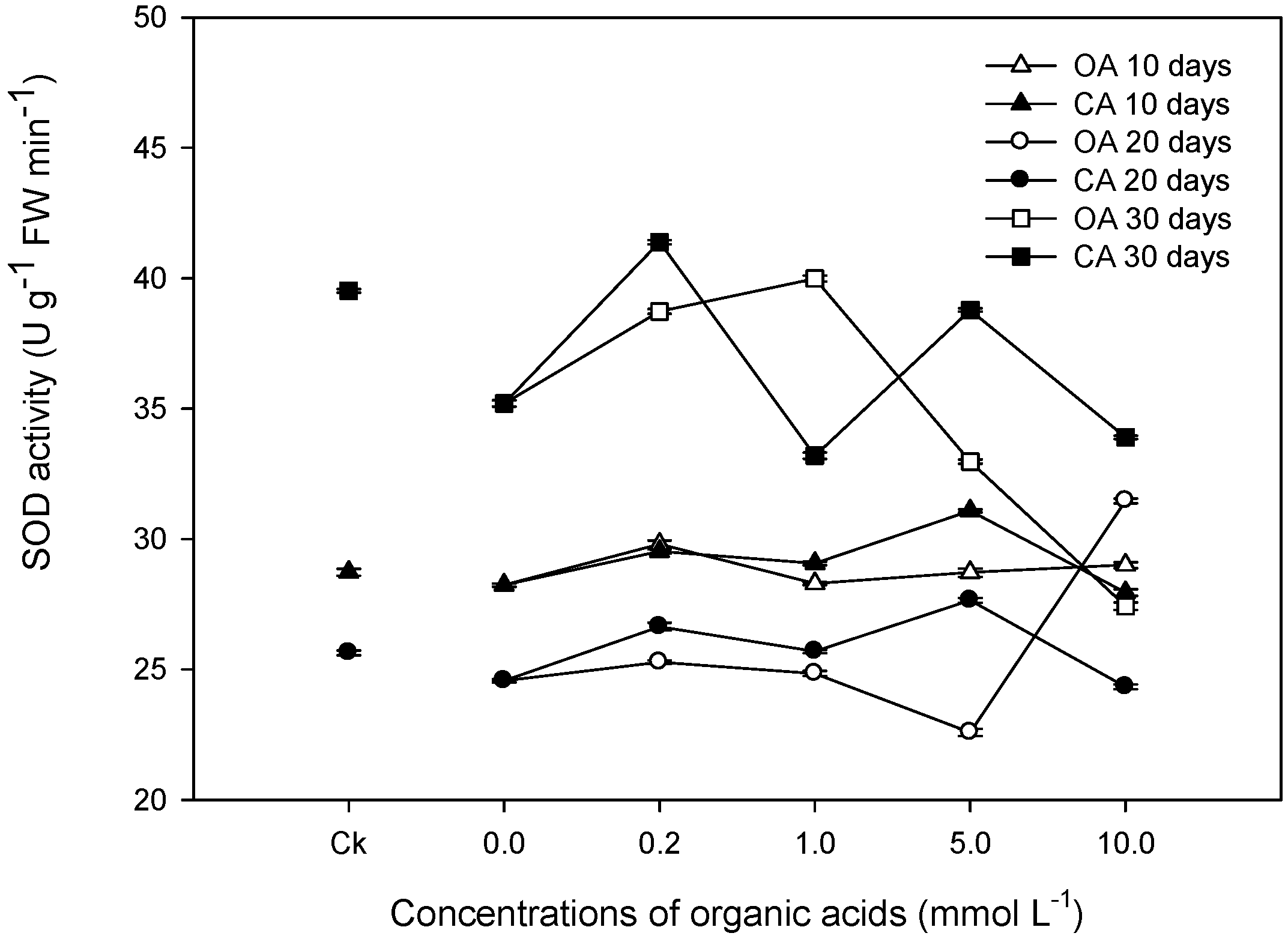
3.3. Activities of SOD
3.4. Proline Contents
3.5. Photosynthetic Pigment Contents and Chlorophyll Fluorescence
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, Z.M.; Song, Z.L.; Parr, J.F.; Wang, H.L. Occluded C in rice phytoliths: Implications to biogeochemical carbon sequestration. Plant Soil 2013, 370, 615–623. [Google Scholar] [CrossRef]
- Wu, J.S.; Xu, Q.F.; Jiang, P.K.; Cao, Z.H. Dynamics and distribution of nutrition elements in bamboos. J. Plant Nutr. 2009, 32, 489–501. [Google Scholar] [CrossRef]
- Xie, J.S.; Shao, C.H.; Tang, X.Y.; Shi, Q.H. Proteomics analysis of nutrient stress effects on rice leaf senescence at grain filling stage. Chin. J. Rice Sci. 2011, 25, 143–149. (In Chinese) [Google Scholar]
- Pizzeghello, D.; Zanella, A.; Carletti, P.; Nardi, S. Chemical and biological characterization of dissolved organic matter from silver fir and beech forest soils. Chemosphere 2006, 65, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Kpomblekou-A, K.; Tabatabai, M.A. Effect of low-molecular weight organic acids on phosphorus release and phytoavailabilty of phosphorus in phosphate rocks added to soils. Agric. Ecosyst. Environ. 2003, 100, 275–284. [Google Scholar] [CrossRef]
- Kenji, M.; Yasuomi, T. Regulation of water, salinity, and cold stress responses by salicylic acid. Front. Plant Sci. 2014. [Google Scholar] [CrossRef]
- Mu, X.L.; Hu, X.L. Effects of salicylic acid on germination of cotton seeds and bud. Cotton China 2003, 30, 19–21. (In Chinese) [Google Scholar]
- Wu, X.P.; Liu, G.S.; Zhu, K.; Yang, C.; Jing, Y.F. Effect of applying organic acids on tobacco physiology and the leaf chemical components. Chin. Tob. Sci. 2003, 9, 23–27. (In Chinese) [Google Scholar]
- Shen, H.; Yan, X.L.; Zhao, M.; Zheng, S.L.; Wang, X.R. Exudation of organic acids in common bean as related to mobilization of aluminum- and iron-bound phosphates. Environ. Exp. Bot. 2001, 48, 1–9. [Google Scholar] [CrossRef]
- Liu, Y.; Mi, G.H.; Chen, F.J.; Zhang, J.H.; Zhang, F.S. Rhizosphere effect and root growth of two maize (Zea mays L.) genotypes with contrasting P efficiency at low P availability. Plant Sci. 2004, 167, 217–223. [Google Scholar] [CrossRef]
- Ryan, P.R.; Elhaize, E.D.; Randall, P.J. Malate efflux from rot apices: Evidence for a general mechanism of Al-tolerance in wheat. Aust. J. Plant Physiol. 1995, 22, 531–536. [Google Scholar] [CrossRef]
- Ashraf, M.; Iram, A. Drought stress induced changes in some organic substances in nodules and other plant parts of two potential legumes differing in salt tolerance. Flora 2005, 200, 535–546. [Google Scholar] [CrossRef]
- Song, J.F.; Cui, X.Y. Analysis of organic acids in selected forest litters of northeast China. J. For. Res. 2003, 14, 285–289. [Google Scholar]
- Cooperative Research Group on Chinese Soil Taxonomy. In Chinese Soil Taxonomy; Science Press: Beijing, China; New York, NY, USA, 2001.
- ISSS/ISRIC/FAO. World Reference Base for Soil Resources; World Soil Resource Reports 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Wang, Z.Q.; Guo, D.L.; Wang, X.R.; Gu, J.C.; Mei, L. Fine root architecture, morphology and biomass, of different branch orders of two Chinese temperate tree species. Plant Soil 2006, 288, 155–171. [Google Scholar] [CrossRef]
- Pang, X.Q.; Cui, X.J.; Sun, Z.F.; Qi, Z.X. Application of Foliar fertilizer Potassium dihydrogen phosphate in culture of larch seedlings. Pract. For. Technol. 2010, 3, 22. (In Chinese) [Google Scholar]
- Xiao, Y.; Tang, Y.K.; Cao, Y.P.; Wang, J.G. Application of surfactants in fertilizer for foliage dressing and its progress. Phosphate Compd. Fertil. 2003, 18, 14–16. (In Chinese) [Google Scholar]
- Franco, A.C.; Herzog, B.; Hűbner, C.; De Mattos, E.A.; Scarano, F.R.; Ball, E.; Lűttge, U. Diurnal changes in chlorophyll a fluorescence, CO2-exchange and organic acid decarboxylation in the tropical CAM tree Clusia hilariana. Tree Physiol. 1999, 19, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. Principles and Techniques of Plant Physiological Biochemical Experiment; Higher Education Press: Beijing, China, 2000; pp. 134–263. (In Chinese) [Google Scholar]
- Song, J.F.; Ma, R.; Huang, W.B.; Yang, D.; Cui, X.Y. Exogenous organic acids protect Changbai Larch (Larix olgensis) seedlings against cadmium toxicity. Fresenius Environ. Bull. 2014, 23, 3459–3467. [Google Scholar]
- Chris, K.L.; Fox, T.R.; Carlson, C.A. Foliage and litter chemistry, decomposition, and nutrient release in Pinus taeda. Forests 2013, 4, 595–612. [Google Scholar]
- Panfili, F.; Schneider, A.; Vives, A.; Perrot, F.; Hubert, P.; Pellerin, S. Cadmium uptake by durum wheat in presence of citrate. Plant Soil 2009, 316, 299–309. [Google Scholar] [CrossRef]
- Krajnc, A.U.; Kristl, J.; Ivancic, A. Application of salicylic acid induces antioxidant defense responses in the phloem of Picea abies and inhibits colonization by Ips typographus. For. Ecol. Manag. 2011, 261, 416–426. [Google Scholar] [CrossRef]
- Sandalio, L.M.; Dalurzo, H.C.; Gomez, M.; Romero-Puertas, M.C.; DelRio, L.A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [PubMed]
- Belkhadi, A.; Hediji, H.; Abbes, Z.; Nouairi, I.; Barhoumi, Z.; Zarrouk, M.; Chaïbi, W.; Djebali, W. Effects of exogenous salicylic acid pre-treatment on cadmium toxicity and leaf lipid content in Linum usitatissimum L. Ecotoxicol. Environ. Saf. 2010, 73, 1004–1011. [Google Scholar] [CrossRef] [PubMed]
- Elastner, E.F. Oxygen activation and oxygen toxicity. Ann. Rev. Plant Physiol. 1982, 33, 73. [Google Scholar] [CrossRef]
- Sas, L.; Rengel, Z.; Tang, C. Excess cation uptake, and extrusion of protons and organic acid anions by Lupinus albus under phosphorus deficiency. Plant Sci. 2001, 160, 1191–1198. [Google Scholar] [CrossRef]
- Shakoor, M.B.; Ali, S.; Hameed, A.; Farid, M.; Hussain, S.; Yasmeen, T.; Najeeb, U.; Bharwana, S.A.; Abbasi, G.H. Citric acid improves lead (pb) phytoextraction in Brassica napus L. by mitigating pb-induced morphological and biochemical damages. Ecotoxicol. Environ. Saf. 2014, 109, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Fernández, R.; Bertrand, A.; Reis, R.; Mourato, M.P.; Martins, L.L.; González, A. Growth and physiological responses to cadmium stress of two populations of Dittrichia viscosa (L.) Greuter. J. Hazard. Mater. 2013, 244, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.Q.; Shi, G.X.; Jia, R.; Chen, L.; Tian, X.L.; Xu, J. Physiological and biochemical responses induced by lead stress in Spirodela polyrhiza. Plant Growth Regul. 2012, 67, 217–225. [Google Scholar] [CrossRef]
- Zhong, Z.Y.; Lin, K.; Nora, F.Y.T. Lead stress in seedlings of Avicennia marina, a common mangrove species in South China, with and without cotyledons. Aquat. Bot. 2010, 92, 112–118. [Google Scholar]
- Fodor, J.; Gullner, G.; Adam, A.L.; Barna, B.; Komives, T.; Kiraly, Z. Local and systemic responses of antioxidants to tobacco mosaic virus infection and to salicylic acid in tobacco. Plant Physiol. 1997, 114, 1443–1451. [Google Scholar] [PubMed]
- Singh, I.; Shah, K. Evidences for suppression of cadmium induced oxidative stress in presence of sulphosalicylic acid in rice seedlings. Plant Growth Regul. 2015, 76, 99–110. [Google Scholar] [CrossRef]
- Guo, H.; Hong, C.; Chen, X.; Xu, Y.; Liu, Y.; Jiang, D.; Zheng, B. Different growth and physiological responses to cadmium of the three Miscanthus species. PLoS ONE 2016, 11, e0153475. [Google Scholar] [CrossRef] [PubMed]
- Scandalio, J.G. Molecular genetics of superoxide dismutases in plants. In Oxidative Stress and the Molecular Biology of Antioxidant Defenses; Scandalios, J.G., Ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1997; pp. 545–546. [Google Scholar]
- Xu, W.H.; Li, Y.R.; He, J.P.; Ma, Q.F.; Zhang, X.J.; Chen, G.Q.; Wang, H.X.; Zhang, H.B. Cd uptake in rice cultivars treated with organic acids and EDTA. J. Environ. Sci. 2010, 22, 441–447. [Google Scholar] [CrossRef]
- Cavalieri, A.J.; Huang, A.H.C. Evaluation of proline accumulation in the adaptation of diverse species of marsh halophytes to the saline environment. Am. J. Bot. 1979, 66, 307–312. [Google Scholar] [CrossRef]
- Boscaiu, M.; Lull, C.; Llinares, J.; Vicente, O.; Boira, H. Proline as a biochemical marker in relation to the ecology of two halophytic Juncus species. J. Plant Ecol. 2013, 6, 177–186. [Google Scholar] [CrossRef]
- Li, H.Y.; Liu, Y.G.; Zeng, G.M.; Zhou, L.; Wang, X.; Wang, Y.Q.; Wang, C.L.; Hu, X.J.; Xu, W.H. Enhanced efficiency of cadmium removal by Boehmeria nivea (L.) Gaud in the presence of exogenous citric and oxalic acids. J. Environ. Sci. 2014, 26, 2508–2516. [Google Scholar] [CrossRef] [PubMed]
- Fagbenro, J.A.; Agboola, A.A. Effect of different levels of humic acid on the growth and nutrient of teak seedlings. J. Plant Nutr. 1993, 16, 1465–1483. [Google Scholar] [CrossRef]
- Han, Y.L.; Huang, S.Z.; Yuan, H.Y.; Zhao, J.Z.; Gu, J.G. Organic acids on the growth, anatomical structure, biochemical parameters and heavy metal accumulation of Iris lactea var. chinensis seedling growing in Pb mine tailings. Ecotoxicology 2013, 22, 1033–1042. [Google Scholar] [PubMed]
- Yang, J.P.; Xu, H.M.; Wang, W.P.; Pan, J.B.; Xiao, N. The effects of SA and osmotic stress on physiological characteristics of maize seedlings. J. Beijing Agric. Coll. 2003, 18, 7–9. (In Chinese) [Google Scholar]
- Madrid, E.N.; Armitage, A.R.; Quigg, A. The response of photosystem II to soil salinity and nutrients in wetland plant species of the northwestern gulf of Mexico. J. Coast. Res. 2012, 28, 1197–1207. [Google Scholar] [CrossRef]
- Xue, J.M.; Clinton, P.W.; Davis, M.R.; Siddiqui, T.; Beets, P.N.; Leckie, A.C. Genotypic variation in foliar nutrient concentrations, δ13C, and chlorophyll fluorescence in relation to tree growth of radiata pine clones in a serpentine soil. J. Plant Nutr. Soil Sci. 2013, 176, 724–733. [Google Scholar] [CrossRef]
- Chatzistathis, T.A.; Papadakis, I.E.; Therios, I.N.; Giannakoula, A.; Dimassi, K. Is chlorophyll fluorescence technique a useful tool to assess manganese deficiency and toxicity stress in olive plants? J. Plant Nutr. 2011, 34, 98–114. [Google Scholar] [CrossRef]
- Guo, H.; Yao, J.T.; Sun, Z.M.; Duan, D.L. Effects of salinity and nutrients on the growth and chlorophyll fluorescence of Caulerpa lentillifera. Chin. J. Oceanol. Limnol. 2015, 33, 410–418. [Google Scholar] [CrossRef]
- Osório, J.; Osório, M.L.; Correia, P.J.; Varennes, A.D.; Pestana, M. Chlorophyll fluorescence imaging as a tool to understand the impact of iron deficiency and resupply on photosynthetic performance of strawberry plants. Sci. Hortic. 2014, 165, 148–155. [Google Scholar] [CrossRef]
- Boyce, R.L. Chlorophyll fluorescence response of red spruce and balsam fir to a watershed calcium fertilization experiment in New Hampshire. Can. J. For. Res. 2007, 37, 1518–1522. [Google Scholar] [CrossRef]
- Strand, M. Effect of mineral nutrient content on oxygen exchange and chlorophyll a fluorescence in needles of Norway spruce. Tree Physiol. 1997, 17, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Sun, K.; Su, X.; Pan, Y.X.; Zhang, Y.F.; Wang, X.P. Physiological responses and tolerance mechanisms to Pb in two xerophils: Salsola passerina Bunge and Chenopodium album L. J. Hazard. Mater. 2012, 205–206, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Ye, J.Y. Effects of oxaloacetate and malate on photosynthesis in leaves and in intact chloroplasts from spinach. Acta Phytophystol. Sin. 2001, 27, 478–482. (In Chinese) [Google Scholar]
- Palomo, L.; Claassen, N.; Jones, D.L. Differential mobilization of P in the maize rhizosphere by citric acid and potassium citrate. Soil Biol. Biochem. 2006, 38, 683–692. [Google Scholar] [CrossRef]
- Burckhard, S.R.; Schwab, A.P.; Banks, M.K. The effects of organic acids on the leaching of heavy metals from mine tailings. J. Hazard. Mater. 1995, 41, 135–145. [Google Scholar] [CrossRef]
- Cui, F.X.; Song, J.F.; Yang, D. Effects of exogenous organic acids on phosphorus availability of dark brown forest soils and phosphorus absorption and accumulation of Larix olgensis seedlings with nutrient deficiency. J. Soil Water Conserv. 2012, 26, 116–121. (In Chinese) [Google Scholar]
- Song, J.F.; Cui, X.Y.; Wang, Z.Q. Effects of low molecular-weight organic acids on P, Fe and K availability of dark brown forest soils and absorption of forest seedlings. J. Soil Water Conserv. 2011, 25, 123–127. (In Chinese) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil horizon | pH (H2O) | CEC a (cmol·kg−1) b | Organic Matter (g·kg−1) | Total N (g·kg−1) | Total P (g·kg−1) | Available P (mg·kg−1) c | Soil Texture | Clay (<2 µm) (g·kg−1) |
|---|---|---|---|---|---|---|---|---|
| A1 | 5.36 | 40.5 | 108.0 | 6.41 | 2.10 | 46.1 | loam d | 134 |
| B | 5.10 | 21.0 | 12.2 | 1.10 | 1.32 | 10.3 | loam d | 191 |
| Treatment | Ck | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 |
|---|---|---|---|---|---|---|---|---|---|---|
| Soil | A1 a | A1:B b | A1:B | A1:B | A1:B | A1:B | A1:B | A1:B | A1:B | A1:B |
| Organic acid type | - c | - | OA | OA | OA | OA | CA | CA | CA | CA |
| Organic acid concentration (mmol·L−1) | 0 | 0 | 0.2 | 1.0 | 5.0 | 10.0 | 0.2 | 1.0 | 5.0 | 10.0 |
| Effect | n | Day | Acid | Conc | Day × Acid | Acid × Conc | Day × Conc |
|---|---|---|---|---|---|---|---|
| Survival | 10 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Bioleaf | 10 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Biostem | 10 | <0.01 | 0.067 | <0.01 | <0.01 | <0.01 | <0.01 |
| Bioroot | 10 | <0.01 | <0.01 | <0.01 | 0.035 | <0.01 | <0.01 |
| Mgroot | 3 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Kroot | 3 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Feroot | 3 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Mgleaf | 3 | <0.01 | <0.01 | <0.01 | 0.155 | 0.165 | <0.01 |
| Kleaf | 3 | <0.01 | 0.239 | <0.01 | <0.01 | <0.01 | <0.01 |
| Feleaf | 3 | <0.01 | 0.137 | <0.01 | <0.01 | <0.01 | <0.01 |
| MDA | 3 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| SOD | 3 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Proline | 3 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Car | 3 | <0.01 | 0.110 | <0.01 | <0.01 | <0.01 | <0.01 |
| Chl | 3 | <0.01 | 0.359 | <0.01 | <0.01 | <0.01 | <0.01 |
| Fv/Fm | 10 | 0.351 | <0.01 | <0.01 | 0.069 | <0.01 | <0.01 |
| Treatment (mmol·L−1) | 10 Days | 20 Days | 30 Days | |||
|---|---|---|---|---|---|---|
| Total Chl a | Car b | Total Chl | Car | Total Chl | Car | |
| Ck | 0.834 ± 0.033 b | 0.145 ± 0.006 abc | 1.017 ± 0.163 ab | 0.168 ± 0.024 bc | 1.192 ± 0.144 f | 0.158 ± 0.023 cd |
| A1:B + acid 0 | 0.798 ± 0.008 a | 0.139 ± 0.008 ab | 0.96 ± 0.188 a | 0.157 ± 0.03 ab | 1.085 ± 0.271 d | 0.146 ± 0.04 abc |
| A1:B + OA 0.2 | 0.904 ± 0.047 cd | 0.14 ± 0.015 ab | 0.962 ± 0.185 a | 0.152 ± 0.034 a | 1.038 ± 0.009 c | 0.139 ± 0.003 a |
| A1:B + OA 1.0 | 0.877 ± 0.228 c | 0.152 ± 0.034 bcd | 1.124 ± 0.131 c | 0.166 ± 0.027 b | 1.21 ± 0.165 f | 0.161 ± 0.03 de |
| A1:B + OA 5.0 | 0.803 ± 0.05 a | 0.133 ± 0.01 a | 1.112 ± 0.139 c | 0.166 ± 0.019 b | 1.166 ± 0.311 e | 0.172 ± 0.047 e |
| A1:B + OA 10.0 | 0.835 ± 0.037 b | 0.148 ± 0.003 bcd | 0.962 ± 0.046 a | 0.147 ± 0.015 a | 1.361 ± 0.335 g | 0.206 ± 0.046 f |
| A1:B + CA 0.2 | 0.922 ± 0.12 d | 0.158 ± 0.019 cd | 1.045 ± 0.201 b | 0.179 ± 0.032 cd | 0.924 ± 0.041 a | 0.138 ± 0.002 a |
| A1:B + CA 1.0 | 0.917 ± 0.074 d | 0.161 ± 0.008 d | 1.212 ± 0.148 d | 0.191 ± 0.023 d | 1.09 ± 0.133 d | 0.155 ± 0.023 bcd |
| A1:B + CA 5.0 | 0.88 ± 0.071 c | 0.157 ± 0.011 cd | 1.348 ± 0.18 e | 0.184 ± 0.028 d | 0.934 ± 0.232 ab | 0.135 ± 0.038 a |
| A1:B + CA 10.0 | 0.894 ± 0.062 cd | 0.16 ± 0.009 d | 1.109 ± 0.088 c | 0.155 ± 0.017 ab | 0.96 ± 0.136 b | 0.143 ± 0.022 ab |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Markewitz, D.; Liu, Y.; Liu, X.; Cui, X. The Alleviation of Nutrient Deficiency Symptoms in Changbai Larch (Larix olgensis) Seedlings by the Application of Exogenous Organic Acids. Forests 2016, 7, 213. https://doi.org/10.3390/f7100213
Song J, Markewitz D, Liu Y, Liu X, Cui X. The Alleviation of Nutrient Deficiency Symptoms in Changbai Larch (Larix olgensis) Seedlings by the Application of Exogenous Organic Acids. Forests. 2016; 7(10):213. https://doi.org/10.3390/f7100213
Chicago/Turabian StyleSong, Jinfeng, Daniel Markewitz, Yong Liu, Xingping Liu, and Xiaoyang Cui. 2016. "The Alleviation of Nutrient Deficiency Symptoms in Changbai Larch (Larix olgensis) Seedlings by the Application of Exogenous Organic Acids" Forests 7, no. 10: 213. https://doi.org/10.3390/f7100213