
Grand Fir Nutrient Management in the Inland Northwestern USA


1
DRP Forestry, Hayden, ID 83835, USA
2
Forest, Rangeland and Fire Sciences, University of Idaho; Moscow, ID 83844, USA
*
Author to whom correspondence should be addressed.
Forests 2016, 7(11), 261; https://doi.org/10.3390/f7110261
Submission received: 31 August 2016
/
Revised: 4 October 2016
/
Accepted: 31 October 2016
/
Published: 4 November 2016
(This article belongs to the Special Issue Nutrient Cycling and Plant Nutrition in Forest Ecosystems)
Abstract
:Grand fir (Abies grandis (Douglas ex D. Don) Lindley) is widely distributed in the moist forests of the Inland Northwest. It has high potential productivity, its growth being nearly equal to western white pine, the most productive species in the region. There are large standing volumes of grand fir in the region. Nutritionally, the species has higher foliage cation concentrations than associated conifers, especially potassium (K) and calcium (Ca). In contrast, it has lower nitrogen (N) foliage concentrations, which creates favorable nutrient balance on N-limited sites. Despite concentration differences, grand fir stores proportionally more nutrients per tree than associated species because of greater crown biomass. Although few fertilization trials have examined grand fir specifically, its response is inferred from its occurrence in many monitored mixed conifer stands. Fertilization trials including grand fir either as a major or minor component show that it has a strong diameter and height growth response ranging from 15% to 50% depending in part on site moisture availability and soil geology. Grand fir tends to have a longer response duration than other inland conifers. When executed concurrently with thinning, fertilization often increases the total response. Late rotation application of N provides solid investment returns in carefully selected stands. Although there are still challenges with the post-fertilization effects on tree mortality, grand fir will continue to be an important species with good economic values and beneficial responses to fertilization and nutrient management.
1. Introduction
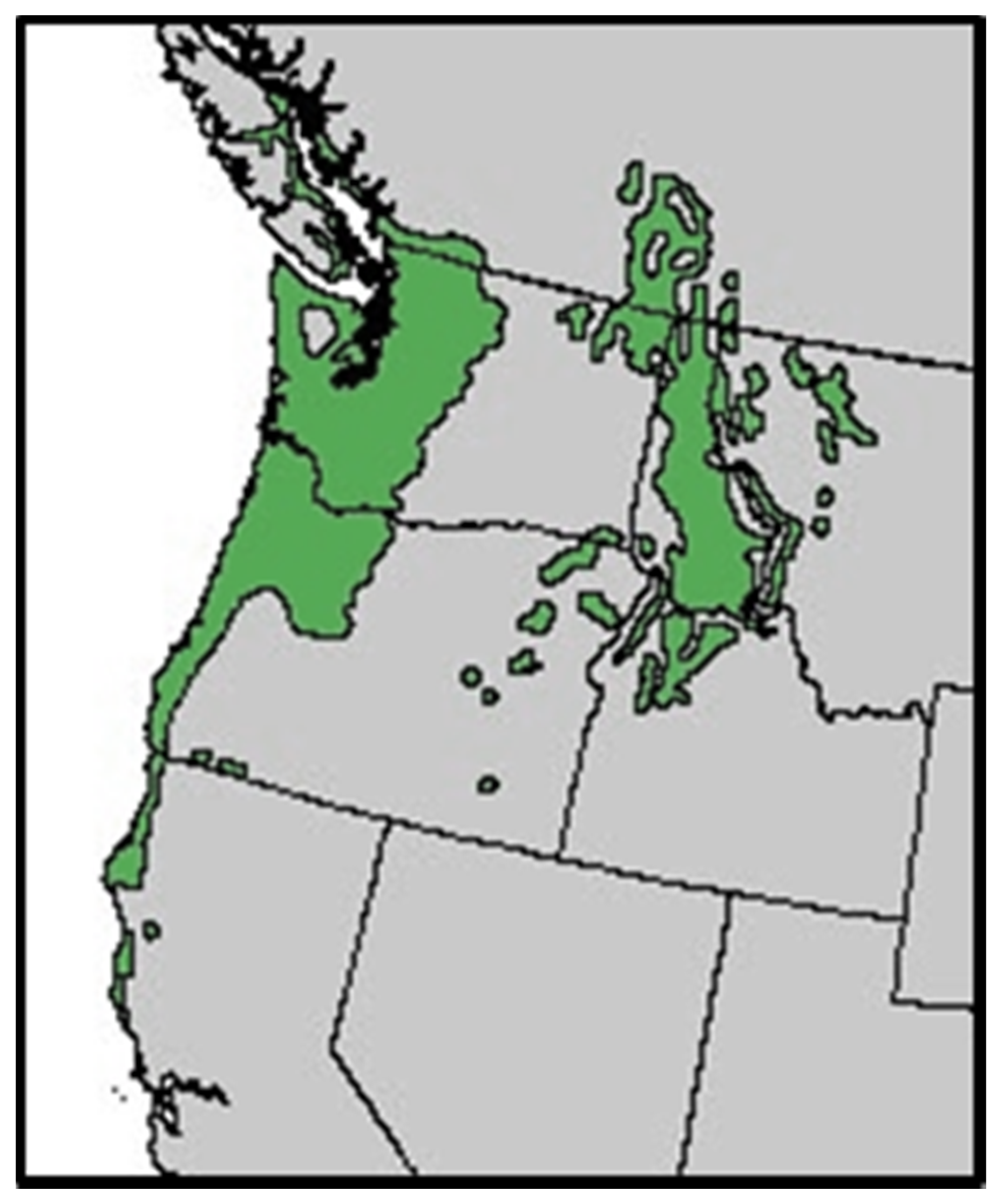
Grand fir is widely distributed in the moist forests of the Pacific Northwest (Figure 1). Its range of 39–51 degrees latitude and 114–125 degrees longitude includes southern Canada in the Okanogan and Kootenay Lakes region south through northeastern Washington, northern Idaho, western Montana, and northeastern Oregon [1]. In the Inland Northwest, it occurs on moister sites from 450 to 1800 m elevation on a wide variety of soils derived from basalts, granites, gneiss, and sandstone as long as moisture is adequate [1]. Grand fir does best throughout the region where a deep layer of volcanic ash occurs [2,3]. There are similar occurrences of grand fir throughout western Montana habitat types and northeastern Washington and Oregon plant associations [4,5]. Grand fir is shade tolerant and often takes a subordinate position in the stand in mixture with other inland species.
Grand fir has high potential productivity. Once established, it grows rapidly nearly equal to Douglas-fir (Pseudotsuga mensiesii (Mirb.) Franco) on the Pacific Coast and western white pine (Pinus monticola Dougl. ex D. Don) in Idaho [1]. As it approaches 50 years old, height is comparable to other associated species in the Inland Northwest including western larch (Larix occidentalis Nutt.). At 100-years-old, grand fir may approach or surpass other species in total height and volume yield. Table 1 is a general comparison of grand fir and its associates in the Inland Region using data from various historical authors [7,8,9]. The last two columns of Table 1 include data for coastal Douglas-fir for an approximate comparison with inland Douglas-fir [10].
There are large standing volumes of grand fir in the Inland Northwest. It ranks first in total growing stock volume at 26 percent [11]. Douglas-fir ranks second at 23 percent. In Idaho, grand fir also makes up the largest percentage of species harvested, ranking first at 36 percent with Douglas-fir second at 26 percent [12].
Grand fir log and lumber values are lower than Douglas-fir, which is the framing lumber standard in the Inland Region. Grand fir has compensating wood quality advantages over Douglas-fir, such as higher log scale, lighter weight, less shrinkage during drying, and excellent fiber characteristics. Alternative forest products that add value are Christmas trees, floral boughs, and oils obtained from the foliage.
2. Nutrient Characteristics
Grand fir has some unique nutritional characteristics compared to other inland conifers. When grand fir was compared to other conifers in over 160 stands, it had higher foliar base cation levels and also higher variation [13]. Grand fir N foliar concentrations are slightly less than associated conifers (Figure 2a). However, foliar K is higher (Figure 2b) and Ca levels are even higher (Figure 2c). Grand fir also has higher foliar concentrations of sulfur (S) and boron (B) than the other species (not shown). Foliar nutrient data from several studies confirm that N concentrations were similar for all species but other cations were higher in grand fir [14].
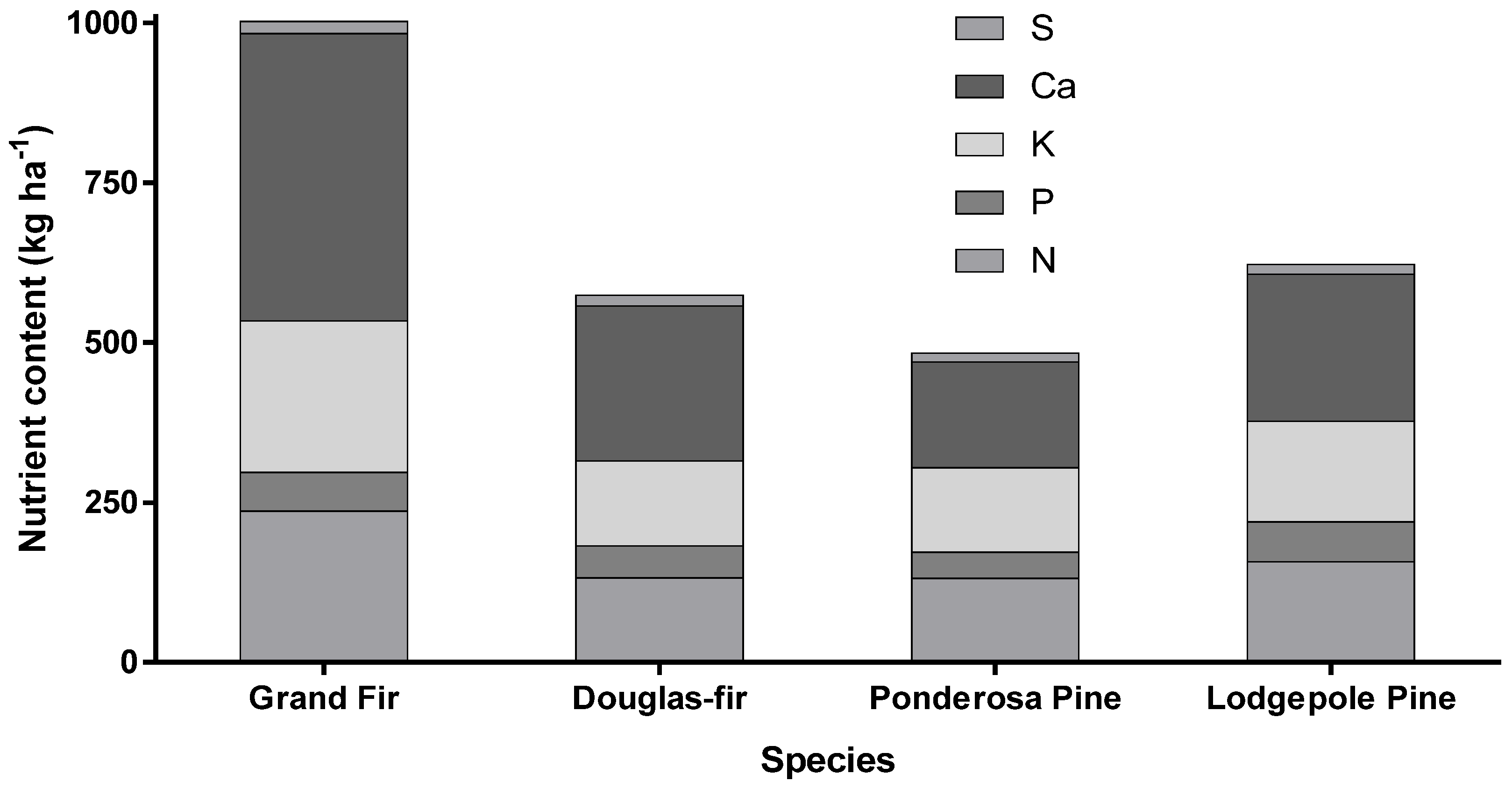
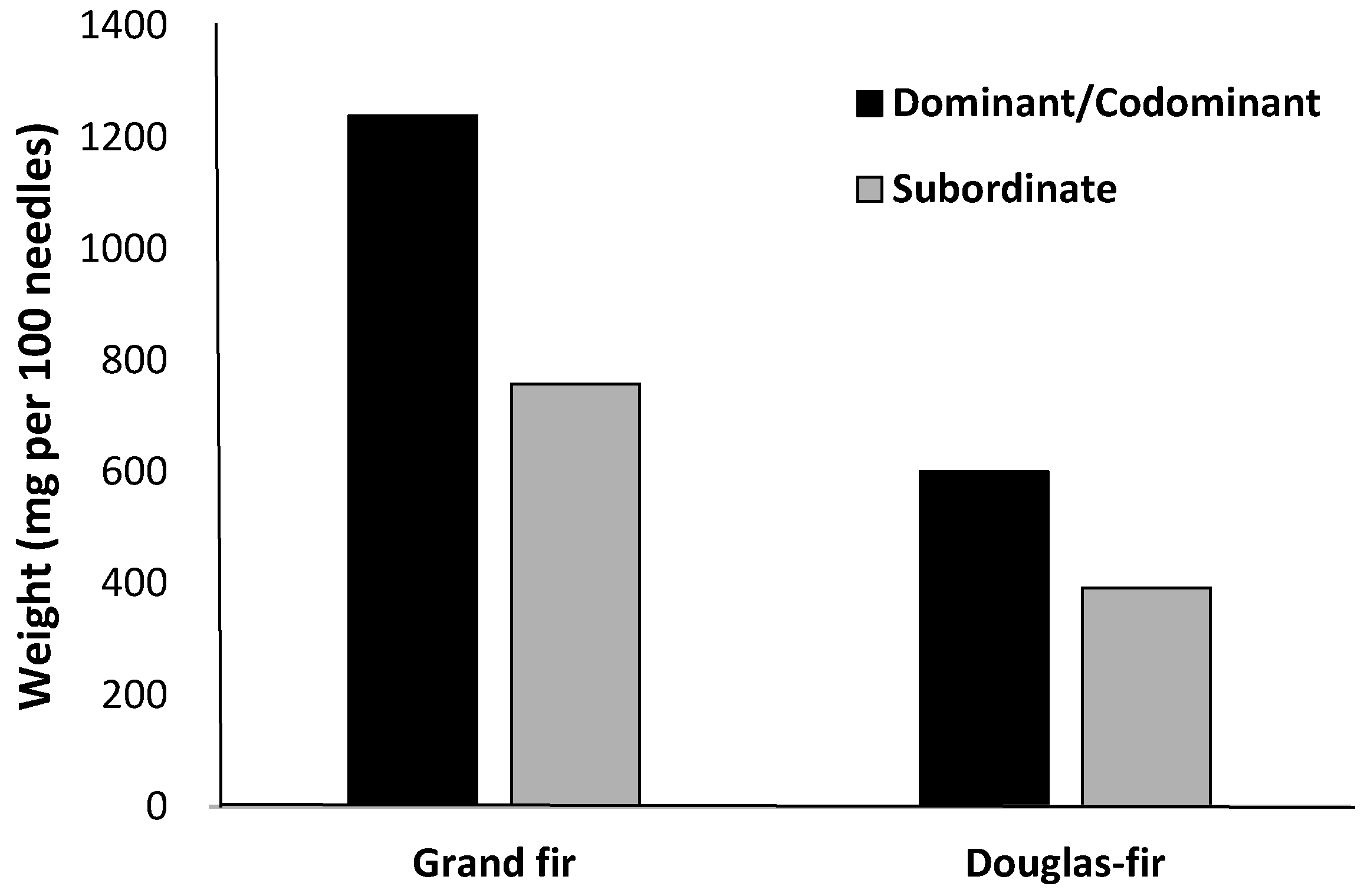
Canopy nutrient content shows that grand fir acquires more nutrients than other conifer species (Figure 3). Grand fir holds twice the canopy foliar biomass as ponderosa pine and lodgepole pine and 1.4 times the foliar mass as Douglas-fir for a given diameter class [15], which is in part due to greater needle weight in comparison to other inland species (Figure 4).
Shade tolerant tree species (such as grand fir) incorporate cations at higher rates than intolerants (Figure 2b,c). Consequently, in mixed-conifer stands, grand fir may be more competitive for nutrients than less tolerant conifers, particularly when nutrient supply is limited [14]. This indicates that grand fir may have greater nutrient demand than associated species, which would affect its competitive ability in mixed stands and its response to nutrient additions.
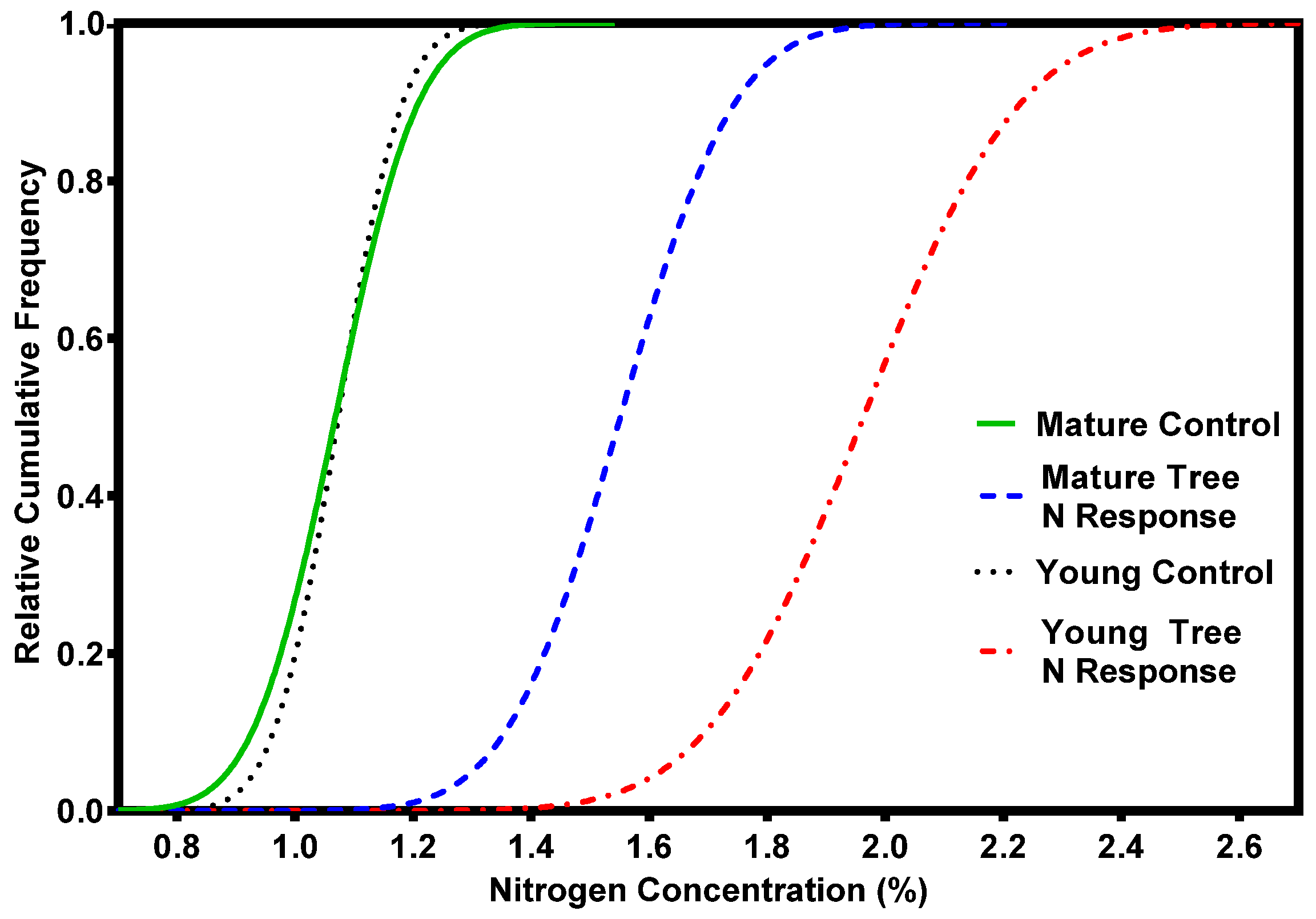
We evaluate nutrient status by comparing foliage nutrient concentrations of field-collected test samples to critical foliar nutrient levels determined from greenhouse-grown seedlings. The critical nutrient level is the concentration above which growth does not respond to nutrient additions [14,18]. Table 2 shows the critical foliar nutrient levels for four Inland Northwest tree species. Seedling-based critical levels are used as reference values in mature stands. The critical level of foliar N in grand fir is 11.5 mg·g−1, 18 percent less than Douglas fir. Critical levels of other selected macro- and micro-nutrients are about the same as for other species, suggesting that grand fir may be able to cope with lower N concentrations than other species due to its larger crown biomass, and relatively low N values (Figure 2a). Although critical levels of N are lower, only 20 percent of unfertilized grand fir has adequate levels of N (Figure 2a) indicating that there still may be considerable opportunity for growth increases from N additions. However, there are important limitations in our knowledge of seedling critical levels relate to fertilizer responses in mature trees.
Grand fir’s shade tolerant ecological status may help it survive and grow in a subordinate position in mixed stands. The lower critical N levels compared to Douglas-fir and lodgepole pine and foliar N levels in untreated stands indicate that grand fir is able to use N conservatively when in low supply. Conversely, when N is available, grand fir may utilize N more aggressively. Higher foliar concentrations of other nutrients such as K and B in untreated grand fir stands suggest that it is commonly in good nutritional balance [14].
Nutrient balance is an important consideration in tree nutrition. Plants require nutrients in specific proportions, which Ingestad [19,20] expressed as optimal nutrient ratios calculated relative to N. Nutrient ratios are consistent among plant taxa due to metabolic requirements, and therefore, are important indicators of nutrient balance (Table 2). As with critical nutrient concentrations, optimal nutrient ratios are based on seedling studies. We assume they are consistent in mature trees because there is important stoichiometric balance among all taxa [21], but we are not aware of studies that consider nutrient balance in mature trees. Grand fir nutrient ratios across three studies found more than 80% of trees measured had K, S, B, and Cu ratios above the Ingestad ratios listed in Table 2. Phosphorous ratios of grand fir averaged near Ingestad’s values. Grand fir has relatively high nutrient ratios compared with inland Douglas-fir [22] and perhaps other inland northwest conifers because of the relatively low N concentrations and high concentrations of other nutrients in tree foliage (Figure 2). The resulting good nutritional balance of grand fir further suggests that it may have a competitive advantage.
The previous data show that grand fir has lower N concentrations and N critical levels than Douglas-fir and other inland conifer species. However, the concentrations and ratios of other nutrients are relativity high in grand fir. Grand fir may be better able to utilize available site N than other inland conifer species because of this favorable nutrient balance. Furthermore, grand fir maintains relatively high foliage biomass and therefore stores site nutrients for its own use and out of reach from competitors. These unique features may be partially explained by grand fir’s physiology as a shade tolerant species [23]. Perhaps grand fir can tolerate low levels of soil N by sequestering site N in foliage while maintaining a favorable nutrient balance. It is then well-prepared for aggressive growth when opportunity arises.
3. Response to Fertilization
Fertilization is an important tool in nutrient management of conifer stands containing grand fir. The commonly deficient N levels along with subordinate canopy position and shade-tolerance of grand fir suggest that it may have strong foliar and growth response to N additions. However, several complicating factors must be accounted for when considering foliar nutrient and stand growth response to fertilizer. These factors include stand density, the relationships with site quality, and the fact that N fertilizer growth response can be offset by an increase in mortality. We consider each of these factors in the following sections.
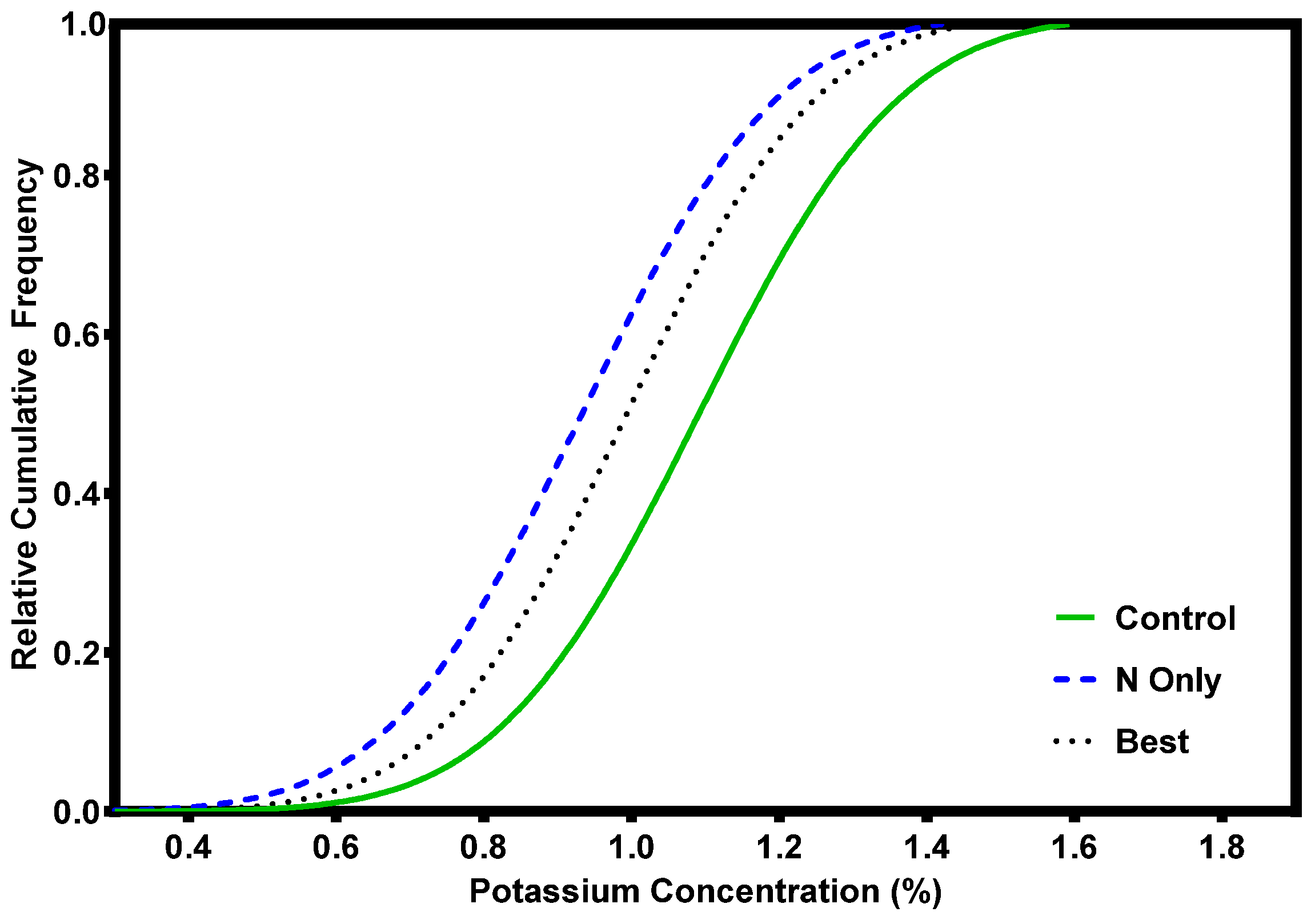
Foliar data for grand fir reveals a strong response to N additions, both in young stands at crown closure and mature stands at the end of the rotation (Figure 5). There was also good foliar response to additions of S and B (data not shown). Foliage did not respond positively to K additions (Figure 6) perhaps because all the trees were above critical K levels before fertilization. It is also possible that the amount of added K was insufficient relative to added N, or that the results simply reflect the dilution effect of expanding foliage in response to the N. Limited trials with phosphorous (P) in the Inland Northwest show no response in either foliar nutrient or growth responses [24]. Consequently, P has receive little attention as a limiting nutrient in the Inland Northwest. There is some evidence that mature trees respond to P amendments in more alkaline, N-rich soils where additional research is warranted [25].
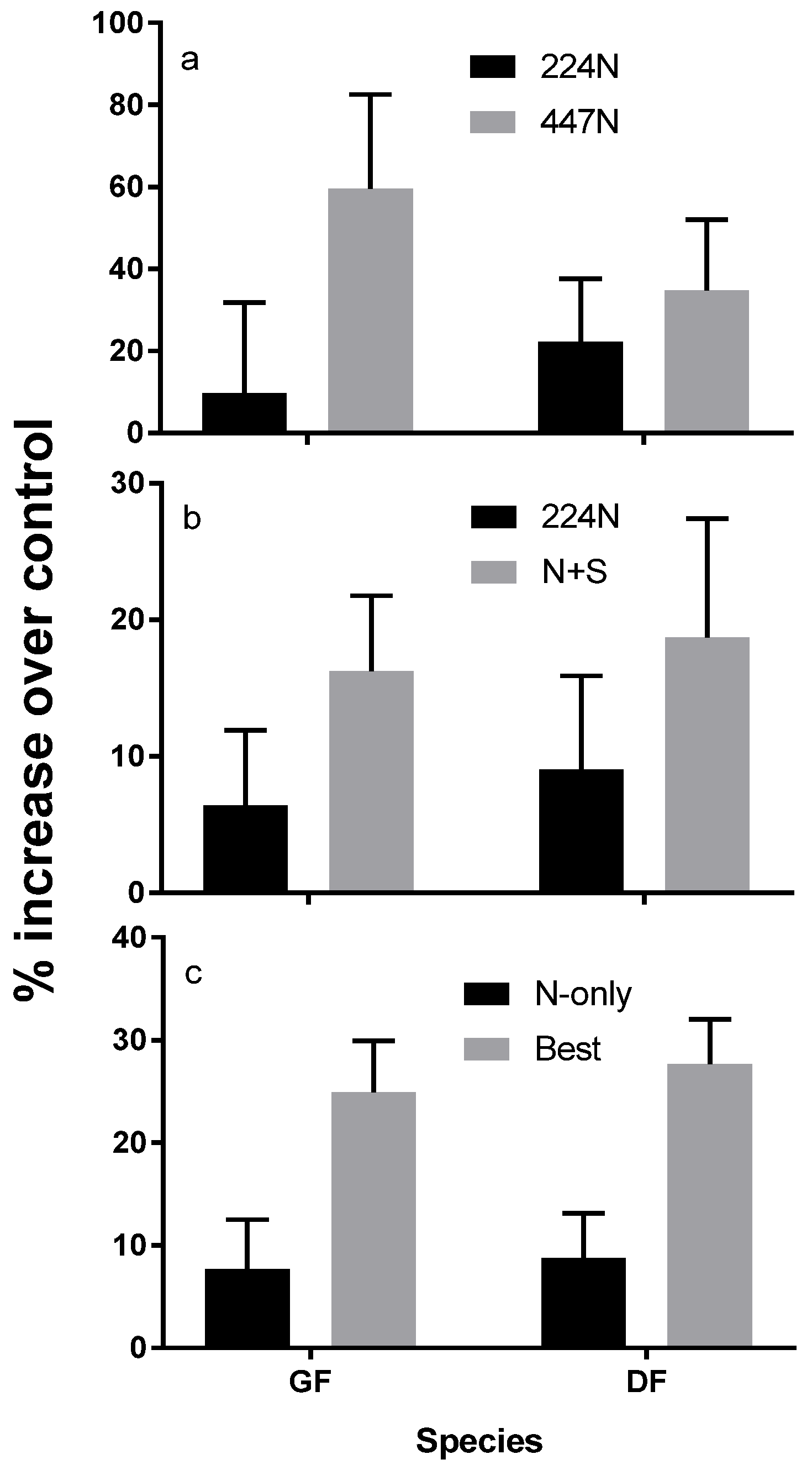
Reflecting foliar responses, N fertilization trials generally show a strong response in both tree diameter, and height [26]. Various trials show consistent growth responses to N rates or combinations of N with other nutrients [16,27] (Figure 7). Growth responses over 20 percent are within the realm of economic feasibility. Additional macro- and micronutrients have resulted in a response in some, but not all instances. For example, multi-nutrient trials showed that grand fir responds well to S and B applications but not necessarily K [16,27,28]. Limited responses to K are also observed in Douglas-fir, even though K is suspected of protecting trees from mortality following N-only fertilization [29].
Response trials have shown that the length of the growth response is 4–6 years for unthinned stands [23,32] and up to 14 years in properly thinned stands [32,33]. Thinning concentrates the growth on fewer trees allowing them to take advantage of other site resources and sustain the response for a longer period.
Repeating a fertilization application can have differing effects depending on the site [27]. For example, grand fir showed a strong repeat response with an additional 90 kg N ha−1 treatment on the drier sites, but did not respond to higher rates. Grand fir also showed a good repeat response to 89 and 180 kg N ha−1 on the wetter sites. However, Douglas fir did not respond to these repeat treatments. Repeat fertilizations of grand fir on moist sites at rates of 90 to 180 kg N ha−1 are expected to result in a significant basal area growth response compared to a single fertilization. However, a significant response of grand fir is not likely on drier sites.
Differences in site quality can partly explain the range in fertilizer response. Important factors affecting fertilizer response to site quality include soil parent material, and soil moisture as indicated by climax vegetation series. Soil types modified by bedrock geology can influence growth and response to nutrient additions. One of the earliest investigations involving geologic rock types found no differentiation between responses on three different rock types [34]. More precise experimental designs have since detected parent material effects by using individual tree screening trials. Screening trials include different treatments of replicated trees across a uniform site. Sites with various parent materials then contain replicates of that design.
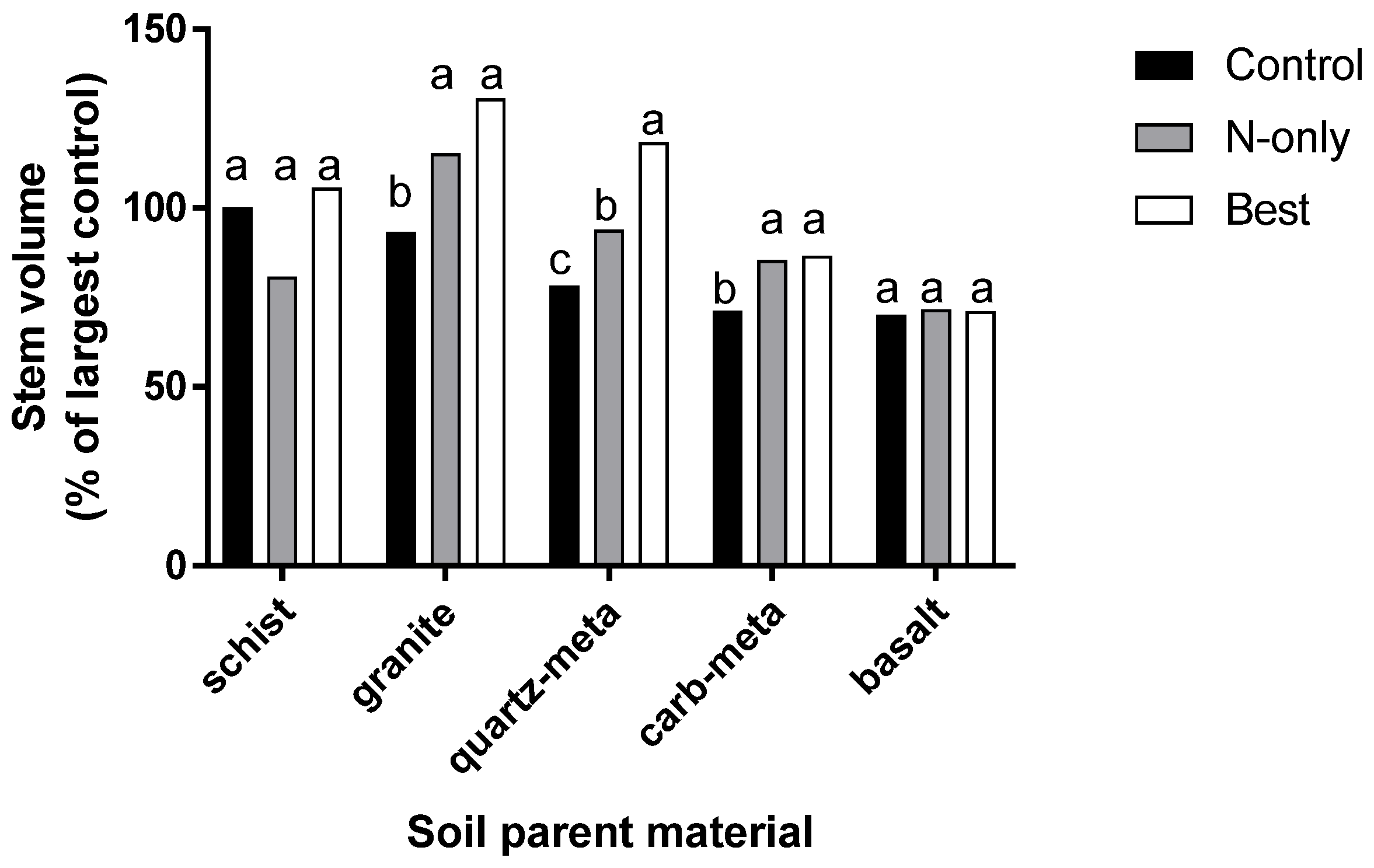
Rock types divide into three categories when unfertilized controls are compared using N-only fertilization, and a multi-nutrient treatment that included N, K, S, and B (Figure 8). Grand fir does not respond to fertilizer additions on soils derived from some parent material types (schist and basalt), indicating they are not deficient in any of the applied nutrients. Sites with soils from other parent materials respond only to N additions, but not multi-nutrients (granite, carbonate bearing metasedimentary, indicating they are N deficient, but other applied nutrients are not limiting. Heavily weathered quartz metasedimentary rocks have the greatest response to multiple nutrients to achieve optimal growth, indicating more than N is deficient. Productivity of grand fir on quartz-meta parent material is equal to the highest growth observed among the screening trials when treated with multi-nutrient fertilizer.
Vegetation series as described by habitat type [2] also affect growth response due to soil moisture variations. Habitat types containing more available soil moisture or higher amounts of organic matter would tend to have better response. Soils containing volcanic ash display increased productivity and N response for all conifer species due to the increase in water-holding capacity [3]. Yet, only half of the sites where grand fir is the climax vegetation contain significant ash deposits. Higher elevation vegetation series are associated with greater moisture availability and ash depth, and grand fir is capable of higher productivity on these sites due to increased moisture availability during the growing season. The limited ash deposits and dryer site conditions for the grand fir habitat type suggest that grand fir is tolerant of dryer conditions. However, when grown on sites with abundant moisture grand fir is capable of better growth than on drier sites. The above results demonstrate that there are differences in nutrient availability and fertilization response among soils, bedrock types, and vegetation series but these differences interact with moisture availability and will require focused research effort to discover mechanisms responsible for distinctions among bedrock geology, surficial deposits, and moisture availability.
The development stage of a forest stand explains some of the response variability observed in previous experiments with soils and site types. A good understanding of these stages and their effect on nutrient requirements helps determine when nutrient additions will provide a growth response. Stands of trees have differing nutritional needs depending on where they are in the successional pathway of development [35].
At the seedling and sapling stage, the trees are unrestricted in their quest for nutrients, especially if artificially planted with competition control. Consequently, during the seedling stage of development, sites meet seedling nutrient requirements, and seedlings can explore the soil for available nutrients reducing the need for additions. Furthermore, they can take advantage of the nutrient flush that often occurs after timber harvest. Grand fir plantations are rare because the species abundantly regenerates in association with planted seedlings of higher-value species. Consequently, we are not aware of seedling fertilizer tests specifically for grand fir. Nutrition of natural grand fir seedling development is a potential area of research. Tests using planted seedling of other inland conifer species on multiple sites show few consistent responses to broadcast applied nutrients [36]. During one seedling trial where fertilizer was applied into the planting hole, Douglas-fir and ponderosa pine exhibited increased height and diameter growth after outplanting, but also had higher mortality, especially at greater N application rates [37].
As stands develop through intermediate ages, nutrient demands increase until they reach the self-thinning stage when nutrient demands are greatest [35]. At this stage, the ability to respond to treatments is high because of nutrient limitations and high stand vigor. This stage is not necessarily age-related because stand age at self-thinning can vary depending on the size-density relationship, species, site quality, and stand establishment practices [38,39]. One would expect during self-thinning that nutrient additions would increase growth. Unfortunately, many of the recorded grand fir experiments do not specifically indicate the stage of stand development or measures of density.
Experiments in relatively young (10–30 years old), high-density stands showed strong response to fertilization [26,40,41,42]. As stands mature, nutrient demands continue and stand density becomes a controlling influence. If artificial fertilization occurs during this time, grand fir should have a strong response in comparison with associated species. Tree spacing control during this period is important to allow sufficient non-nutrient resources to support fertilizer response. As stands reach mature and old growth conditions, nutrient demands depend more on site factors. Grand fir’s physiology as a late-successional, shade-tolerant tree may affect its response by allowing it to conserve N when necessary but be more demanding during competitive growth phases such as that which would occur during crown closure [23]. Focused study is required on the ability of grand fir to compete for nutrients during between crown closure and self-thinning stages of stand development.
During the crown closure through self-thinning stages of development, additional growth response is possible when thinning and fertilization are combined as demonstrated from numerous experiments [32,34,43,44,45]. After thinning a crowded stand, light, water, and temperature become more favorable for growth [46]. Fertilization within a decade after thinning, or in properly spaced stands allows the trees to take advantage of the additional resources [30,47]. Thinning and then nutrient additions are beneficial, especially if relative densities prior to fertilization allow adequate space for increased growth. Research trials show that combined thinning and fertilization are additive compared to the individual treatments [47,48]. In some instances the total growth response of the crop trees is greater than the sum of either fertilization or thinning effects alone [32].
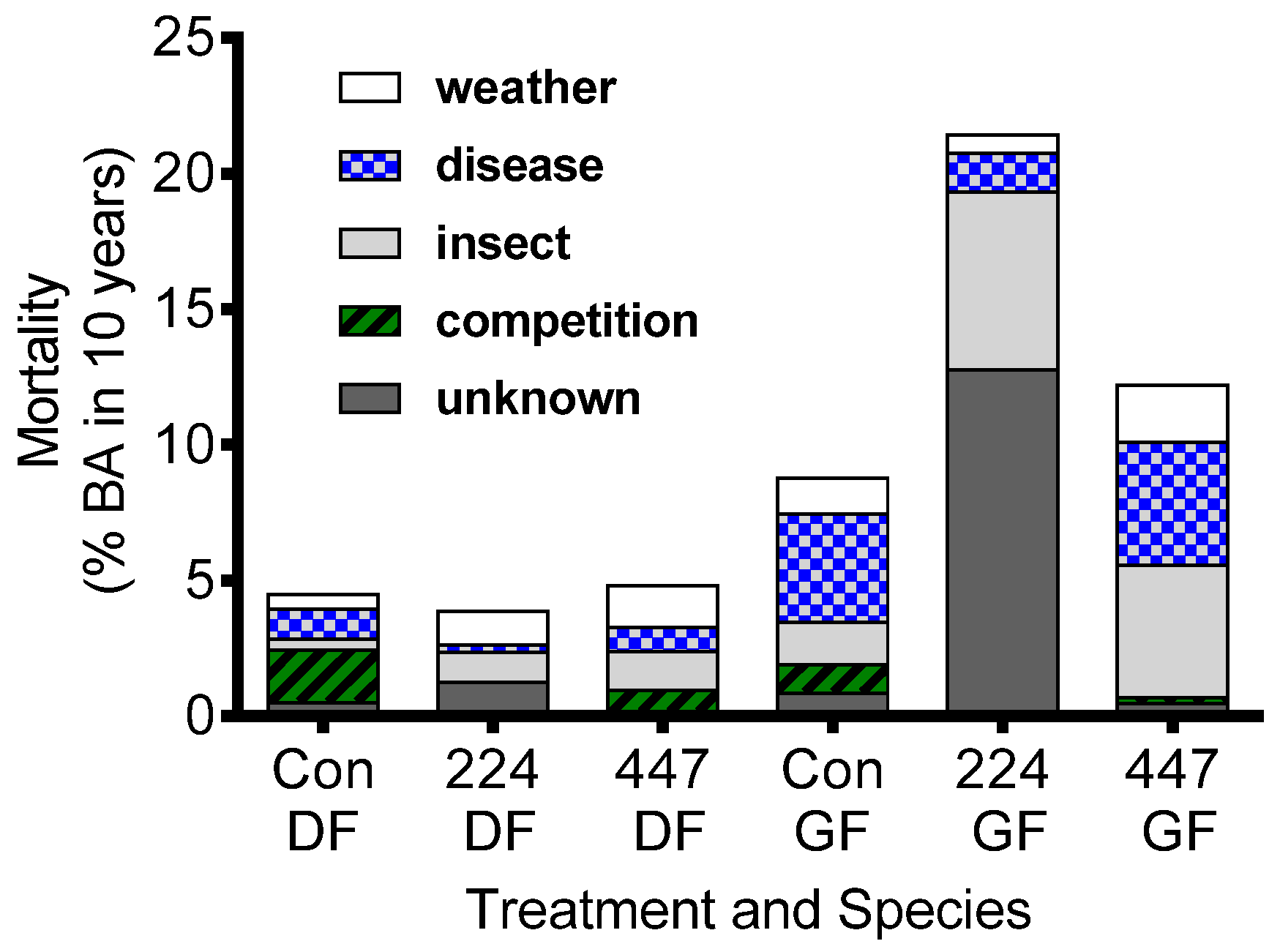
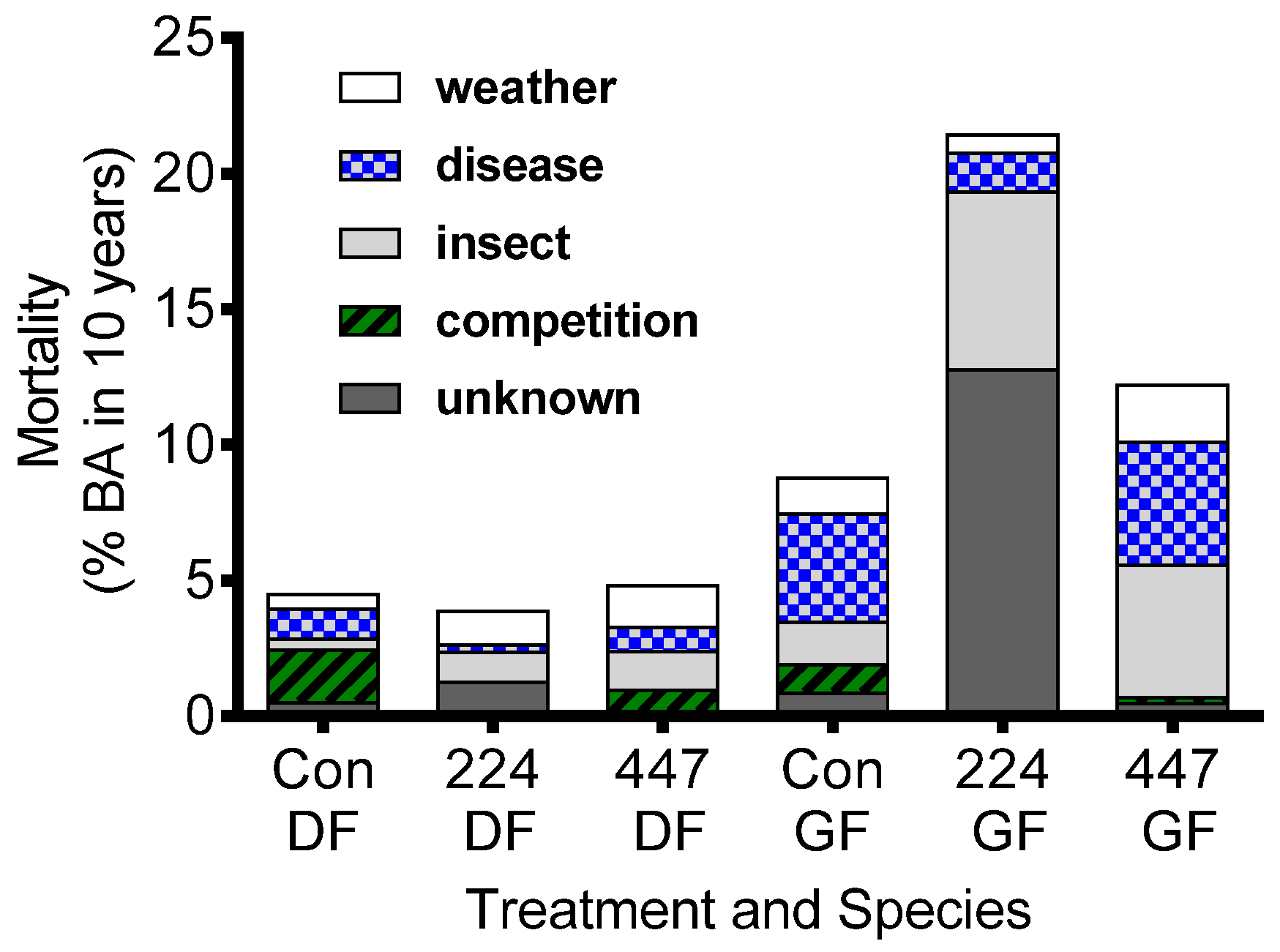
Grand fir exhibits higher mortality after fertilization, especially with N-only amendments. Its mortality is naturally greater than that of Douglas-fir, and N fertilization only increases that trend (Figure 9) [49]. Of the known reasons for mortality, N fertilization increases insect-caused tree mortality more than other causes. This is commonly observed in trees and is attributed to improved foliage nutritional quality [50]. Addition of other macro- and micronutrients tended to reduce mortality but not significantly [16]. There is critical need to learn more about the causes of increase mortality to N-only fertilizer applications.
Nitrogen fertilization can potentially stimulate pests and disease. Insects such as spruce budworm (Choristoneura occidentalis Freeman) reduce current growth, and can diminish fertilizer response [48]. Grand fir is more susceptible to Douglas-fir tussock moth (Orgyia pseudotsugata McDunnough) than other inland conifers, but experience confirms that susceptibility tends to decreases with soil volcanic ash content [51] which, as described above, is expected to interact with soil moisture, tree nutrition and fertilizer application. The effects of Armillaria root disease (Armillaria ostoyae Romagnesi) can substantially reduce net response to fertilization. Therefore, stand-level responses to fertilization may be lower than expected if root disease is present in the stand. Although there were initial experiments implying that fertilization (especially with K additions) [52] may reduce incidence of root disease, a 10-year study in a diseased stand found no treatment differences due to fertilization [27]. Moore et al. [29] suggested that tree nutrition (K/N ratio) controls production of phenolic defense compounds and resistance to the root rot disease Armillaria in Douglas-fir. If true, grand fir should exhibit a similar relationship because it too is susceptible to Armillaria infection. While the relationship between favorable K/N ratios and phenolic concentrations was confirmed in seedlings [53,54] they could not demonstrate an effect of stand-level N and K fertilization on root disease or mortality after ten years. The potential negative effects of pests and diseases following fertilization are important to recognize [55] and managers should avoid fertilization in stands that display these properties. Increased effort is needed to elucidate the relationship between nutrient balance, defense compounds, and root disease.
Vegetation control using herbicides is expanding in the Inland Northwest. Herbicide application is cheaper, more effective, and protects the soil surface from compaction and displacement by heavy site preparation machinery. Grand fir is generally more tolerant to herbicides than associated species [56] so prudent use presents few difficulties. Specific effects of herbicide application in combination with fertilization are not well understood in the Inland Northwest Region. However, in the southeastern U.S., research confirms the positive combined effects of herbicide treatments and nutrient additions [57]. There is evidence that vegetation control alone may provide additional nutrients to trees by eliminating competition and causing faster breakdown and mineralization of organic matter [55].
In summary, these studies indicate that grand fir growing under the right conditions can respond to N fertilization with volume growth ranging from 15% to 50%. Grand fir generally has a longer response duration than other inland conifers, the greatest response occurring during 3–5 years, tapering off during 5–8 years, with some response lasting through 10–12 years under optimum conditions. The response can be greater after well-timed thinning. Repeated fertilizer applications have the potential to maintain growth response depending on the site and the stage of stand development. It is important to recognize site types and their interaction on fertilizer response. Grand fir is sensitive to higher mortality rates after fertilization so when seeking locations for effective treatments, it is critical to avoid sites with heavily weathered parent material where nutrient imbalance may lead to high pest and disease susceptibility.
4. Economics of Grand Fir Fertilization
Large land-holding companies in the Inland Northwest are interested in the economics of fertilization. As a financial investment, forest fertilization must consider the time value of money. Success is very sensitive to the time between the initial outlay and the final return on investment; thus, fertilization in older stands is considered to be more profitable because the volume gains can be harvested sooner [58] and investment returns increase [59]. Late rotation fertilization has a short payback period with relatively low investment costs relative to other forestry investments [60]. Repeated fertilization can extend the response duration, however a second amendment doubles the cost and the response is not as strong as the first application. These economic principles have led foresters in the region to believe that a one-time fertilization late in the rotation can be a sound forestry investment. A recent Internal Revenue Service (IRS) tax law change that allowed expensing of fertilization costs further increased the investment potential [61].
Since most grand fir in the Inland Northwest grows in mixture with Douglas-fir and other species, fertilization experiments require additional analysis and assumptions to confine the results to just grand fir. Because of this, grand fir response, by itself, is usually inferred using data from mixed stands.
Properly spaced, 60–90 years old mature stands containing grand fir may respond well to N fertilization with responses similar to those obtained for Douglas-fir stands in western Washington [58]. The treatment generally involves application of 150–300 kg ha−1 of urea N followed by timber harvest in 8–12 years. This results in a basal area growth response of 20%–80% with comparable volume responses. Addition of multi-nutrients may improve protection against mortality but does not significantly improve individual tree growth response [29], so the added cost does not increase the economic benefit.
Recent operational fertilization treatments in mixed stands including grand fir confirm that investment returns are very good, often surpassing those of other forestry investments, if stands are carefully selected [62]. It is essential to identify stand conditions that will provide an acceptable economic response [63]. Incorporation of a carefully selected fertilization treatment into a general forest management plan can be economically warranted. These growth responses can be used for more sophisticated economic analysis for more accurate projections depending on management requirements. However, such growth responses may require adjustments in coming decades with increased atmospheric carbon dioxide concentrations and accompanying changes in climate.
5. Management Recommendations
We recommend that a grand fir nutrient management and fertilization program consider the following concepts.
1. The greatest response from added nutrients seems to occur just after crown closure when competitive stresses are high.
2. Grand fir is expected to best respond to nutrient additions when it is maintained in a healthy and vigorous condition through stand density management.
3. To conserve site nutrients, soil disturbance should be minimized during timber harvest and stand regeneration activities. Heavy-handed site preparation and slash cleanup may have a negative impact on the site’s nutrient resources, especially low-quality sites [14]. Some alternate possibilities to consider include light broadcast burning; leaving small limbs, foliage, and larger material scattered across the site; and the use of herbicides.
4. Carefully analyze timber stands to determine tree and site conditions, species mix, stand density, soil parent material, and harvest plans before making fertilization decisions to assure effective results. Consult the latest fertilization research results to guide grand fir nutrient management.
6. Conclusions
Grand fir is ubiquitous in Inland Northwest forests growing in mixture with other species, mainly Douglas-fir. It often contains higher crown nutrient content than associate species [14]. Grand fir has good growth response to N fertilizer and possibly K. The effect of nutrition, especially K, on root disease potential remains unconfirmed. Good investment returns are possible from fertilization treatments after careful site evaluation. With good management, and a solid understanding of nutrient dynamics, grand fir will remain an important species with solid economic values and a positive response to fertilizer under the right conditions in Inland Northwest forests.
Acknowledgments
Funding for analysis of data included in this report and preparation of the manuscript was supplied by the Intermountain Forestry Cooperative, formerly Intermountain Forest Tree Nutrition Cooperative (IFTNC). We wish to thank the following people who assisted with the preparation and review of this manuscript: Mark J. Kimsey, Research Assistant Professor, IFTNC/University of Idaho, Terry M. Shaw Research Scientist, IFTNC/University of Idaho.
Author Contributions
Dennis Parent and Mark D. Coleman preformed equally in drafting, editing and revising the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Foiles, M.W.; Graham, R.T.; Olson, D.F. Grand fir. In Silvics of Forest Trees of the United States, Conifers; Burns, R.M., Honkala, B.H., Eds.; USFS: Washington, DC, USA, 1990; Volume 1, pp. 52–59. [Google Scholar]
- Cooper, S.V.; Neiman, K.E.; Roberts, D.W. Forest Habitat Types of Northern Idaho: A Second Approximation; USFS, Intermountain Research Station: Ogden, UT, USA, 1991.
- Garrison-Johnston, M.T.; Mika, P.G.; Miller, D.L.; Cannon, P.; Johnson, L.R. Ash cap influences on site productivity and fertilizer response in forests of the Inland Northwest. In Volcanic-ash-derived Forest Soils of the Inland Northwest: Properties and implications for management and restoration; USFS, RMRS: Coeur d’Alene, ID, USA, 2007; pp. 137–163. [Google Scholar]
- Pfister, R.D.; Kovalchik, B.L.; Arno, S.F.; Presby, R.C. Forest Habitat Types of Montana; USFS, INT Station: Ogden, UT, USA, 1977.
- Williams, C.K.; Kelley, B.F.; Smith, B.; Lillybridge, T.R. Forested Plant Associations of the Colville National Forest; Pacific Northwest Research Station: Portland, OR, USA, 1995.
- USGS; Little, E.L., Jr. Digital representations of “Atlas of United States Trees”. Available online: http://esp.cr.usgs.gov/data/atlas/little/abiegran.pdf (accessed on 31 August 2016).
- Deitschman, G.H.; Green, A.W. Relations between Western White Pine Site Index and Tree Height of Several Associated Species; USFS: Ogden, UT, USA, 1965.
- Haig, I.T. Second-growth Yield, Stand, and Volume Tables for the Western White Pine Type; USFS, Northern Rocky Mountain Forest Experiment Station: Washington, DC, USA, 1932.
- Stage, A.R.; Renner, D.L.; Chapman, R.C. Selected Yield Tables for Plantations and Natural Stands in the Inland Northwest forests; USFS, INT Research Station: Ogden, UT, USA, 1988.
- McArdle, R.E.; Meyer, W.H.; Bruce, D. The Yield of Douglas-Fir in the Pacific Northwest; USFS: Washington, DC, USA, 1961.
- USFS. Forest Inventory Data Online (FIDO). Available online: http://apps.fs.fed.us/fia/fido/index.html (accessed on 2 Jaunary 2016).
- Simmons, E.A.; Hayes, S.W.; Morgan, T.A.; Keegan, C.E., III; Witt, C. Idaho’s Forest Products Industry and Timber Harvest 2011 with Trends through 2013; USFS: Fort Collins, CO, USA, 2013.
- Moore, J.A.; Mika, P.G.; Shaw, T.M.; Garrison-Johnston, M. Foliar nutrient characteristics of four conifer species in the interior Northwest United States. West. J. Appl. For. 2004, 19, 13–24. [Google Scholar]
- Garrison, M.T.; Moore, J.A. Nutrient Management: A Summary and Review. Available online: http://digital.lib.uidaho.edu/cdm/singleitem/collection/iftnc/id/3357/rec/40 (accessed on 29 September 2016).
- Brown, J.K. Weight and Density of Crowns of Rocky Mountain Conifers; USFS, INT Forest & Range Experiment Station: Ogden, UT, USA, 1978.
- Mika, P.G. Grand Fir: Nutrient Ecology and Response to Fertilization. Available online: http://digital.lib.uidaho.edu/cdm/singleitem/collection/iftnc/id/3446/rec/6 (accessed on 29 September 2016).
- Garrison-Johnston, M. Nutrient Cycling in a North Idaho Conifer Stand. Ph.D. Thesis, University of Idaho, Moscow, ID, USA, 2003. [Google Scholar]
- Walker, R.B.; Gessel, S.P. Mineral Deficiencies of Coastal Northwest Conifers; College of Forest Resources, University of Washington: Seattle, WA, USA, 1991. [Google Scholar]
- Ingestad, T. Mineral nutrient requirements of Pinus silvestris and Picea abies seedlings. Physiol. Plant. 1979, 45, 373–380. [Google Scholar] [CrossRef]
- Ingestad, T. A definition of optimum nutrient requirements in birch seedlings. II. Physiol. Plant. 1971, 24, 118–125. [Google Scholar] [CrossRef]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef]
- Coleman, M.D.; Shaw, T.M.; Kimsey, M.J.; Moore, J.A. Nutrition of Douglas-fir in the Inland Northwest. Soil Sci. Soc. Am. J. 2014, 78, 11–22. [Google Scholar] [CrossRef]
- Graham, R.T.; Tonn, J.R. Ten-Year Results of Fertilizing Grand Fir, Western Hemlock, Western Larch, and Douglas-Fir with Nitrogen in Northern Idaho; USFS, INT Station: Ogden, UT, USA, 1985.
- Shaw, T.M.; Kimsey, M. Screening trial data. Intermountain Forestry Cooperative, University of Idaho: Moscow, ID, USA, Unpublished data. 2016. [Google Scholar]
- Mainwaring, D.B.; Maguire, D.A.; Perakis, S.S. Three-year growth response of young Douglas-fir to nitrogen, calcium, phosphorus, and blended fertilizers in Oregon and Washington. For. Ecol. Manag. 2014, 327, 178–188. [Google Scholar] [CrossRef]
- Loewenstein, H.; Pitkin, F.H. Growth Responses and Nutrient Relations of Fertilized and Unfertilized Grand Fir; University of Idaho, Forestry, Wildlife, and Range Experiment Station: Moscow, ID, USA, 1971. [Google Scholar]
- Shaw, T.; Coleman, M.; Kimsey, M.; Mika, P. Forest Health and Nutrition Ten-year Growth, Mortality and Foliar Nutrition; Intermountain Forest Tree Nutrition Cooperative: Moscow, ID, USA, 2014. [Google Scholar]
- Garrison, M.T.; Moore, J.A.; Shaw, T.M.; Mika, P.G. Foliar nutrient and tree growth response of mixed-conifer stands to three fertilization treatments in northeast Oregon and north central Washington. For. Ecol. Manag. 2000, 132, 183–198. [Google Scholar] [CrossRef]
- Moore, J.A.; Mika, P.G.; Schwandt, J.W.; Shaw, T.M. Nutrition and forest health. In Interior Cedar-Hemlock-White Pine Forests: Ecology and Management; Washington State University: Spokane, WA, USA, 1994; pp. 173–176. [Google Scholar]
- Moore, J.A.; Mika, P.G.; Vanderploeg, J.L. Nitrogen fertilizer response of Rocky Mountain Douglas-fir by geographic area across the Inland Northwest. West. J. Appl. For. 1991, 6, 94–98. [Google Scholar]
- Garrison, M.T.; Moore, J.A.; Mika, P.G.; Shaw, T.M. Six-Year Growth Response of the Umatilla and Four-Year Growth Response of the Okanogan Mixed Conifer Stands to N, N + K and N + S Fertilization. Available online: http://digital.lib.uidaho.edu/cdm/ref/collection/iftnc/id/3358 (accessed on 3 October 2016).
- Shafii, B.; Moore, J.A.; Olson, J.R. Effects of nitrogen fertilization on growth of grand fir and Douglas-fir stands in northern Idaho. West. J. Appl. For. 1989, 4, 54–57. [Google Scholar]
- Scanlin, D.C.; Lowenstein, H. Forest Fertilization in the Intermountain Region-six Year Results; University of Idaho, Forestry, Wildlife and Range Experiment Station: Moscow, ID, USA, 1980. [Google Scholar]
- Scanlin, D.C.; Loewenstein, H.; Pitkin, F.H. Two-year Response of North Idaho Stands of Douglas-Fir and Grand Fir to Urea Fertilizer and Thinning; University of Idaho: Moscow, ID, USA, 1976. [Google Scholar]
- Kimmins, J.P. Forest Ecology: A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Shaw, T.M.; Johnson, L.R. Two-Year Growth Response of Douglas-fir and Ponderosa Pine Seedlings to Boron and Multi-Nutrient Fertilization in Northeast Oregon. Available online: http://digital.lib.uidaho.edu/cdm/ref/collection/iftnc/id/4860 (accessed on 4 October 2016).
- Fan, Z.; Moore, J.A.; Osborne, H.L. Three-year response of ponderosa pine seedlings to controlled-release fertilizer applied at planting. West. J. Appl. For. 2002, 17, 154–164. [Google Scholar]
- Oliver, C.D.; O’Hara, K.L.; McFadden, G.; Nagame, L. Concepts of thinning regimes. In Douglas-Fir: Stand Management for the Future; Oliver, C.D., Hanley, D.P., Johnson, J.A., Eds.; Institute of Forest Resources: Seattle, WA, USA, 1986; pp. 246–257. [Google Scholar]
- Weiskittel, A.; Gould, P.; Temesgen, H. Sources of variation in the self-thinning boundary line for three species with varying levels of shade tolerance. For. Sci. 2009, 55, 84–93. [Google Scholar]
- Loewenstein, H.; Pitkin, F.H. Response of grand fir and western white pine to fertilizer applications. Northwest Sci. 1963, 37, 23–30. [Google Scholar]
- Graham, R.T.; Tonn, J.R. Response of Grand Fir, Western Hemlock, Western White Pine, Western Larch, and Douglas-Fir to Nitrogen Fertilizer in Northern Idaho; USFS, INT: Ogden, UT, USA, 1979.
- Powers, R.F. Response of California true fir to fertilization. In Forest Fertilization Conference; Gessel, S.P., Ed.; University of Washington: Union, WA, USA, 1979; pp. 95–101. [Google Scholar]
- Olson, J. Four-Year Fertilization and Thinning Response Analysis of Douglas-fir and Grand Fir Stands in Northern Idaho; Potlatch Corporation: Lewiston, ID, USA, 1979. [Google Scholar]
- Olson, J. Response of Intermountain Grand Fir and Douglas-Fir Stand Types to Nitrogen Fertilization and Thinning; Potlatch Corporation: Lewiston, ID, USA, 1981. [Google Scholar]
- Olson, J.; Hatch, C.R. Volume response of intermountain grand fir stand types to nitrogen fertilizer and thinning treatments. In The Biology and Management of True Fir in the Pacific Northwest; University of Washington: Seattle, WA, USA, 1981; pp. 247–250. [Google Scholar]
- Chase, C.W.; Kimsey, M.J.; Shaw, T.M.; Coleman, M.D. The response of light, water, and nutrient availability to pre-commercial thinning in dry inland Douglas-fir forests. For. Ecol. Manag. 2016, 363, 98–109. [Google Scholar] [CrossRef]
- Scanlin, D.C.; Loewenstein, H. Response of inland Douglas-fir and grand-fir to thinning and nitrogen fertilization in northern Idaho. In Forest Fertilization; Gessel, S.P., Kenady, R.M., Atkinson, W.A., Eds.; University of Washington: Alderbrook Inn, Union, WA, USA, 1979; pp. 82–88. [Google Scholar]
- Cochran, P.H. Response of Thinned White Fir Stands to Fertilization with Nitrogen Plus Sulfur; USFS, PNW Research Station: Portland, OR, USA, 1991.
- Shen, G.; Moore, J.A.; Hatch, C.R. The effect of nitrogen fertilization, rock type, and habitat type on individual tree mortality. For. Sci. 2001, 47, 203–213. [Google Scholar]
- Herms, D.A. Effects of fertilization on insect resistance of woody ornamental plants: Reassessing an entrenched paradigm. Environ. Entomol. 2002, 31, 923–933. [Google Scholar] [CrossRef]
- Stoszek, K.J.; Mika, P.J. Douglas-fir tussock moth outbreaks related to stand conditions and pest management. In 1980 SAF Convention: Land Use Allocation; Society of American Foresters: Bethesda, MD, USA, 1981; pp. 134–137. [Google Scholar]
- Mika, P.G.; Moore, J.A. Foliar potassium status explains Douglas-fir response to nitrogen fertilization in the inland Northwest, USA. Water Air Soil Pollut. 1990, 54, 477–491. [Google Scholar] [CrossRef]
- Schwandt, J.W. Fertilization effects on root disease in two Douglas-fir stands in northern Idaho. In Proceedings of the Fiftieth Western International Forest Disease Work Conference; USFS: Powell River, BC, Canada, 2002; pp. 1–6. [Google Scholar]
- Shaw, T.M.; Moore, J.A.; Marshall, J.D. Root chemistry of Douglas-fir seedlings grown under different nitrogen and potassium regimes. Can. Jour. For. Res. 1998, 28, 1566–1573. [Google Scholar] [CrossRef]
- Mandzak, J.M.; Moore, J.A. The role of nutrition in the health of the inland western forest. J. Sustain. For. 1994, 2, 191–210. [Google Scholar] [CrossRef]
- Miller, D.L.; Gravelle, P.J. Species Selection Guidelines for Planting, Natural Regeneration and Crop Tree Selection on Potlatch Land in Northern Idaho; Potlatch Corporation: Lewiston, ID, USA, 2009. [Google Scholar]
- Will, R.E.; Munger, G.T.; Zhang, Y.; Borders, B.E. Effects of annual fertilization and complete competition control on current annual increment, foliar development, and growth efficiency of different aged Pinus taeda stands. Can. J. For. Res. 2002, 32, 1728–1740. [Google Scholar] [CrossRef]
- Miller, R.E.; Webster, S.R. Fertilizer response in mature stands of Douglas-fir. In Forest Fertilization; University of Washington: Union, WA, USA, 1979; pp. 126–132. [Google Scholar]
- Miller, R.E.; Fight, R.D. Fertilizing Douglas-fir Forests; USFS, PNW Research Station: Portland, OR, USA, 1979.
- Olson, J.R. Experience with nitrogen fertilization in northern Idaho. In Forest Fertilization: Sustaining and Improving Nutrition and Growth in Western Forests; Chappell, H.N., Weetman, G.F., Miller, R.E., Eds.; University of Washington: Seattle, WA, USA, 1992; pp. 232–236. [Google Scholar]
- National Timber Tax Website. Timber Fertilization. Available online: http://www.timbertax.org/research/revenuerulings/bda/04-62/ (accessed on 14 January 2015).
- Shaw, T.M.; Kimsey, M.; Parent, D.R. Growth Response of an Operationally Fertilized Stand at Brickel Creek, North Idaho; IFTNC, University of Idaho: Moscow, ID, USA, 2010. [Google Scholar]
- Olson, J. The fertilization decision: Using what we have learned to evaluate a nitrogen fertilization program. In IFTNC Fertilization Workshop; IFTNC: Moscow, ID, USA, 1988; pp. 1–15. [Google Scholar]
Figure 1.
Range map of grand fir in the Pacific Northwest [6].
Figure 1.
Range map of grand fir in the Pacific Northwest [6].
Figure 2.
Relative cumulative frequency graphs of foliar N (a), K (b), and Ca (c) concentrations for grand fir (GF), inland Douglas-fir (DF), lodgepole pine (LP), and ponderosa pine (PP). Y-axis shows the proportion of trees sampled that are below the nutrient concentration on the horizontal axis. Foliage was collected from over 160 mixed conifer stands that were randomly selected within categories representing geographic region, habitat type, stand density, and rock type to assure that differences in foliar nutrient characteristics reflect species and not sample site differences [13].
Figure 2.
Relative cumulative frequency graphs of foliar N (a), K (b), and Ca (c) concentrations for grand fir (GF), inland Douglas-fir (DF), lodgepole pine (LP), and ponderosa pine (PP). Y-axis shows the proportion of trees sampled that are below the nutrient concentration on the horizontal axis. Foliage was collected from over 160 mixed conifer stands that were randomly selected within categories representing geographic region, habitat type, stand density, and rock type to assure that differences in foliar nutrient characteristics reflect species and not sample site differences [13].
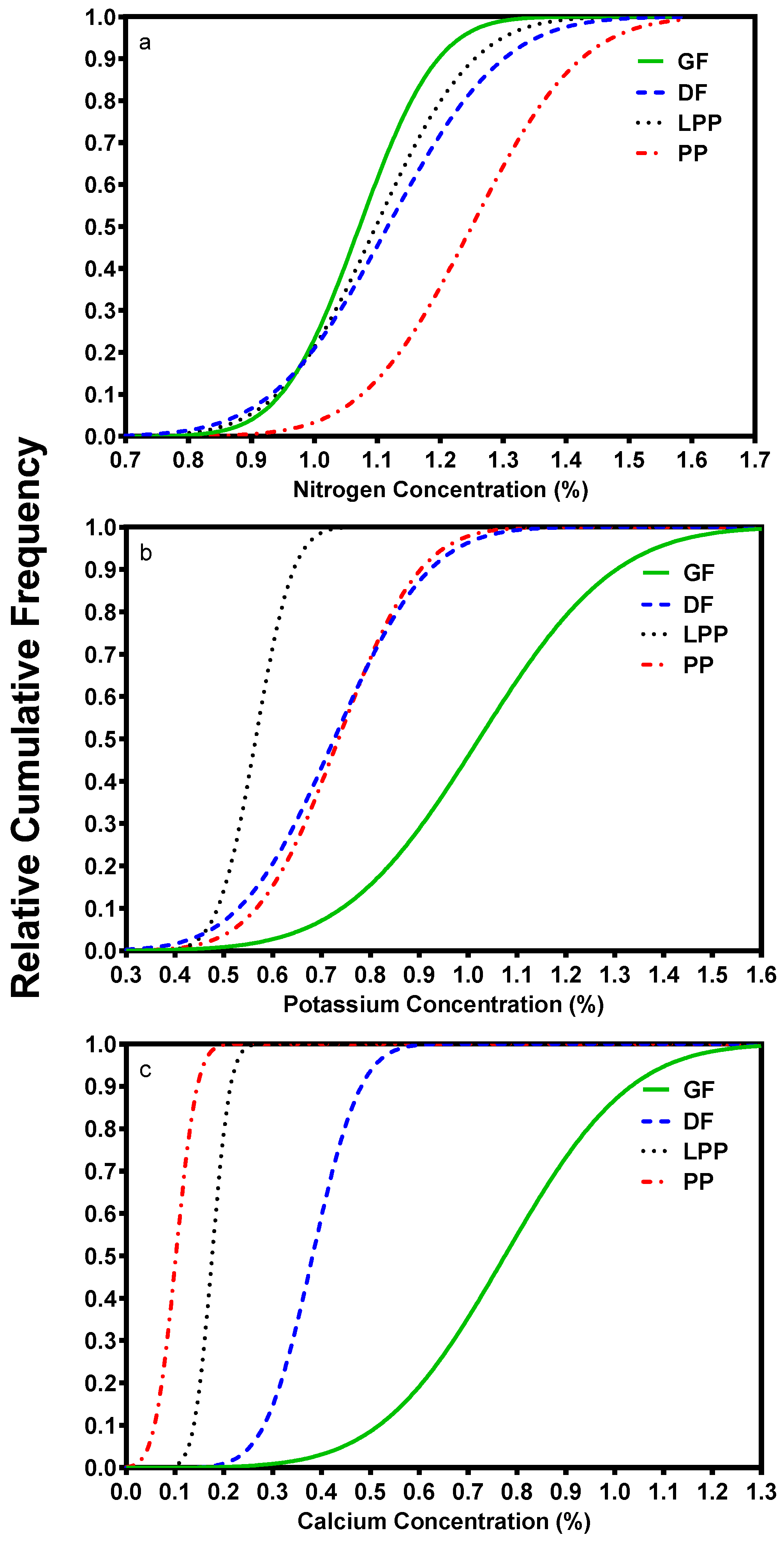
Figure 3.
Total nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), and sulfur (S) content of aboveground portion of four inland tree species: grand fir, Douglas-fir, ponderosa pine, and lodgepole pine [16].
Figure 3.
Total nitrogen (N), phosphorus (P), potassium (K), calcium (Ca), and sulfur (S) content of aboveground portion of four inland tree species: grand fir, Douglas-fir, ponderosa pine, and lodgepole pine [16].

Figure 4.
Foliage weight comparison between crown classes of grand fir and Douglas-fir [17].
Figure 4.
Foliage weight comparison between crown classes of grand fir and Douglas-fir [17].

Figure 5.
Grand fir foliar N response additions for young (~15 year-old) and mature (~40 year-old) stand ages [16]. (Foliage samples were collected one-year after fertilization with 224–336 kg N ha−1 as urea).
Figure 5.
Grand fir foliar N response additions for young (~15 year-old) and mature (~40 year-old) stand ages [16]. (Foliage samples were collected one-year after fertilization with 224–336 kg N ha−1 as urea).

Figure 6.
Grand fir foliar K response after fertilization with 224 kg N ha−1 (N-only) or a blend of 224 kg N ha−1, 170 kg K ha−1 + 73 kg S ha−1 + 2.7 kg B ha−1 (Best) [16].
Figure 6.
Grand fir foliar K response after fertilization with 224 kg N ha−1 (N-only) or a blend of 224 kg N ha−1, 170 kg K ha−1 + 73 kg S ha−1 + 2.7 kg B ha−1 (Best) [16].
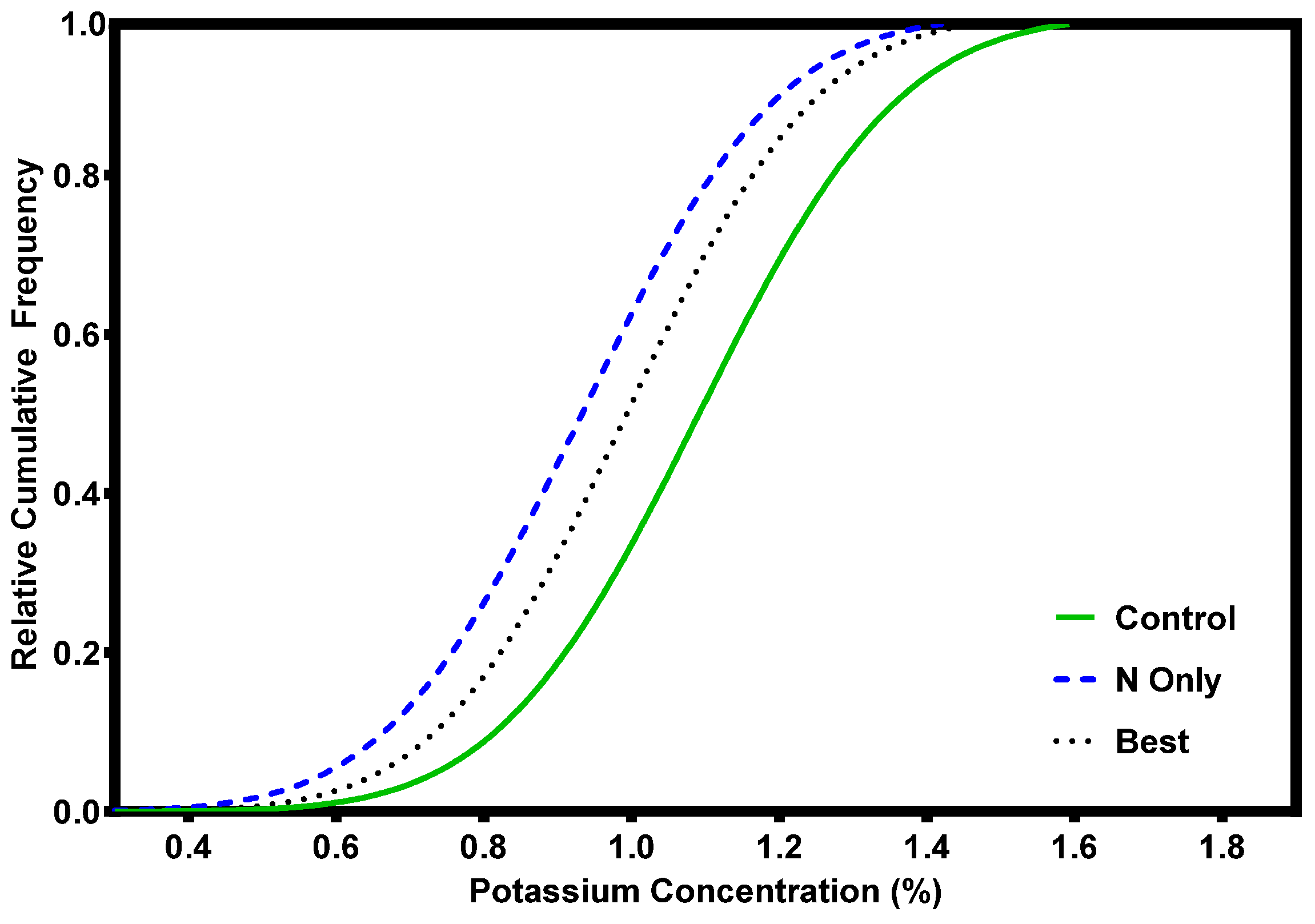
Figure 7.
Grand fir (GF) and Douglas-fir (DF) volume increment response in three different fertilization trials at various locations. (a) Average annual ten-year response to two levels of N-only fertilization at 224 and 447 kg N ha−1 in 30 Douglas-fir dominated (80% basal area) stands in Idaho, Montana, Oregon and Washington including five parent material classes and three vegetation series representing the range of regional productivity [30]; (b) Average annual eight-year response to either 224 kg N ha−1 or 224 kg N ha−1 + 73 kg S ha−1) in seven mixed conifer stands growing on basalt parent materials at 1370 to 1680 m in Abies grandis and Abies lasiocarpa vegetation series [31]; (c) Average annual two-year response to individual-tree treatments of 224 kg N ha−1 (N-only) or a blend of 224 kg N ha−1, 190 kg K ha−1 + 73 kg S ha−1 + 2.7 kg B ha−1 (Best) in eight young (5–10 cm average diameter) stands all on metasedimentary parent material [16].
Figure 7.
Grand fir (GF) and Douglas-fir (DF) volume increment response in three different fertilization trials at various locations. (a) Average annual ten-year response to two levels of N-only fertilization at 224 and 447 kg N ha−1 in 30 Douglas-fir dominated (80% basal area) stands in Idaho, Montana, Oregon and Washington including five parent material classes and three vegetation series representing the range of regional productivity [30]; (b) Average annual eight-year response to either 224 kg N ha−1 or 224 kg N ha−1 + 73 kg S ha−1) in seven mixed conifer stands growing on basalt parent materials at 1370 to 1680 m in Abies grandis and Abies lasiocarpa vegetation series [31]; (c) Average annual two-year response to individual-tree treatments of 224 kg N ha−1 (N-only) or a blend of 224 kg N ha−1, 190 kg K ha−1 + 73 kg S ha−1 + 2.7 kg B ha−1 (Best) in eight young (5–10 cm average diameter) stands all on metasedimentary parent material [16].
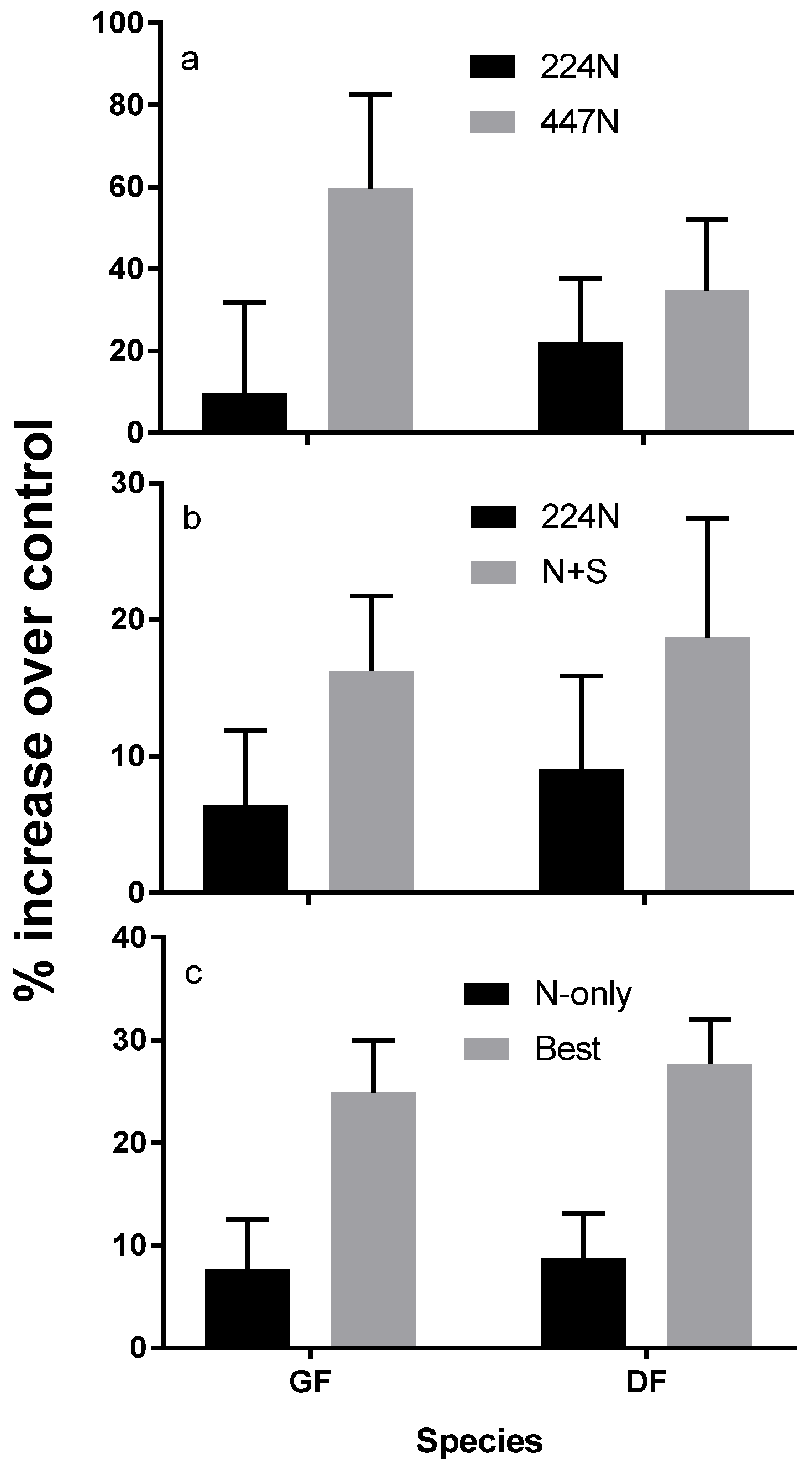
Figure 8.
Fertilizer treatment effects on 2-year stem volume by soil parent material. Stem volume is expressed as a percentage of untreated trees growing on schist soils. Fertilizer treatments included 224 kg N ha−1 (N-only), or a blend of 224 kg N ha−1, 190 kg K ha−1 + 73 kg S ha−1 + 2.7 kg B ha−1 (Best). Two metasedimentary parent materials include a quartzite (quartz-meta) and carbonate (carb-meta) [24].
Figure 8.
Fertilizer treatment effects on 2-year stem volume by soil parent material. Stem volume is expressed as a percentage of untreated trees growing on schist soils. Fertilizer treatments included 224 kg N ha−1 (N-only), or a blend of 224 kg N ha−1, 190 kg K ha−1 + 73 kg S ha−1 + 2.7 kg B ha−1 (Best). Two metasedimentary parent materials include a quartzite (quartz-meta) and carbonate (carb-meta) [24].
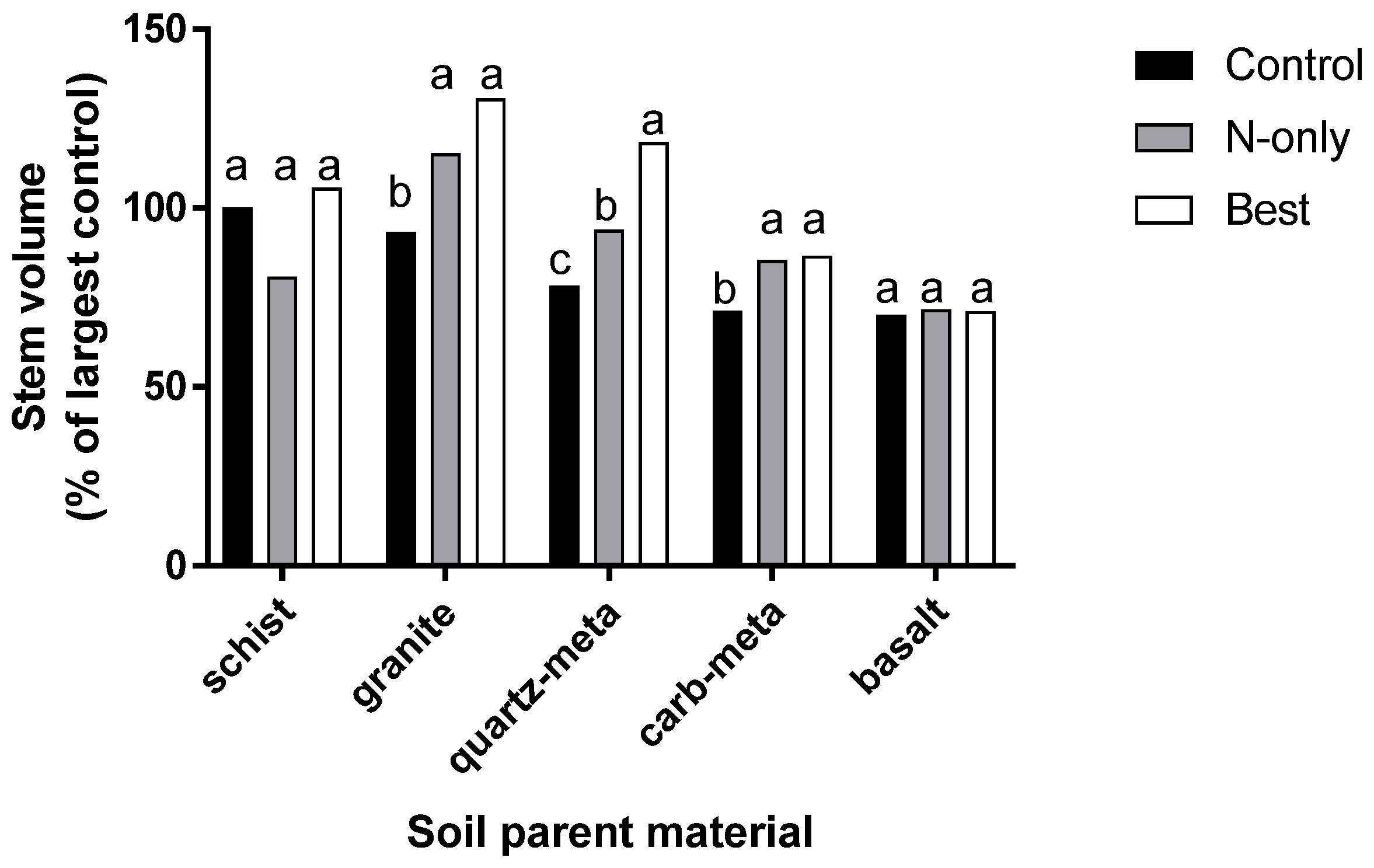
Figure 9.
The causes of ten-year basal area mortality of Douglas-fir and grand fir treated with either 224 or 447 kg N ha−1 and compared with unfertilized controls. Data are from 30 mature (average 65 year-old) Douglas-fir dominated (80% basal area) stands in Idaho, Montana, Oregon and Washington including five parent material classes and three vegetation series representing the range of regional productivity. All stands were recently thinned or were naturally well spaced. Initial basal area averaged 36 ± 10 m2 ha−1 [30].
Figure 9.
The causes of ten-year basal area mortality of Douglas-fir and grand fir treated with either 224 or 447 kg N ha−1 and compared with unfertilized controls. Data are from 30 mature (average 65 year-old) Douglas-fir dominated (80% basal area) stands in Idaho, Montana, Oregon and Washington including five parent material classes and three vegetation series representing the range of regional productivity. All stands were recently thinned or were naturally well spaced. Initial basal area averaged 36 ± 10 m2 ha−1 [30].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Grand fir height and total volume yield compared to associated species in the US Inland Northwest. All yields are from yield tables of fully stocked stands.
| Stand Age | Grand Fir | Western White Pine | Western Larch | Inland Douglas-Fir | Coastal Douglas-Fir 1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Height (m) 2 | Volume (m3) 3 | Height (m) 2 | Volume (m3) 4 | Height (m) 2 | Volume (m3) 5 | Height (m) 2 | Volume (m3) 6 | Height (m) 7 | Volume (m3) 8 | |
| 50 | 20.6 | 74.1 | 21.5 | 140.3 | 22.8 | 37.9 | 20.0 | 73.4 | 23.7 | 110.0 |
| 100 | 36.3 | 367.1 | 42.4 | 393.3 | 37.8 | 141.2 | 35.4 | 206.6 | 33.8 | 260.1 |
Table 2.
Critical nutrient concentrations for inland conifer species: grand fir (GF), Douglas-fir (DF), lodgepole pine (LP), and ponderosa pine (PP) [14].
| Foliar Concentration | GF | DF | LP | PP | Ingestad Ratios * |
|---|---|---|---|---|---|
| N (mg·g−1) | 11.5 | 14.0 | 12.0 | 11.0 | 100 |
| P (mg·g−1) | 1.5 | 1.2 | 1.2 | 0.8 | 16 |
| K (mg·g−1) | 5.8 | 6.0 | 5.0 | 4.8 | 50 |
| S (mg·g−1)) | 0.8 | 1.1 | 0.9 | 0.8 | 7 |
| Cu (µg·g−1) | 3 | 2 | 3 | 3 | 0.03 |
| B (µg·g−1) | 10 | 10 | 4.3 | 20 | 0.2 |
* Optimum nutrient ratios, R = 100 (M/N), where M is the concentration of the element of interest and N is the concentration of N ([19] except S/N).
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Parent, D.R.; Coleman, M.D. Grand Fir Nutrient Management in the Inland Northwestern USA. Forests 2016, 7, 261. https://doi.org/10.3390/f7110261
AMA Style
Parent DR, Coleman MD. Grand Fir Nutrient Management in the Inland Northwestern USA. Forests. 2016; 7(11):261. https://doi.org/10.3390/f7110261
Chicago/Turabian StyleParent, Dennis R., and Mark D. Coleman. 2016. "Grand Fir Nutrient Management in the Inland Northwestern USA" Forests 7, no. 11: 261. https://doi.org/10.3390/f7110261
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.