
A Review of the Characteristics of Small-Leaved Lime (Tilia cordata Mill.) and Their Implications for Silviculture in a Changing Climate


Abstract
:1. Introduction
2. Site Requirements
2.1. Temperature
2.2. Water Supply
2.3. Soil Nutrients
3. Life Strategy
3.1. Light Requirements
3.2. Growth and Productivity
3.2.1. Height Growth
3.2.2. Diameter Growth and Crown Dimensions
3.2.3. Volume Growth and Productivity
3.3. Regeneration
3.3.1. Generative Reproduction
3.3.2. Vegetative Reproduction
3.3.3. Hybridization
3.4. Phenology
3.5. Health and Robustness
4. Genetic Resources
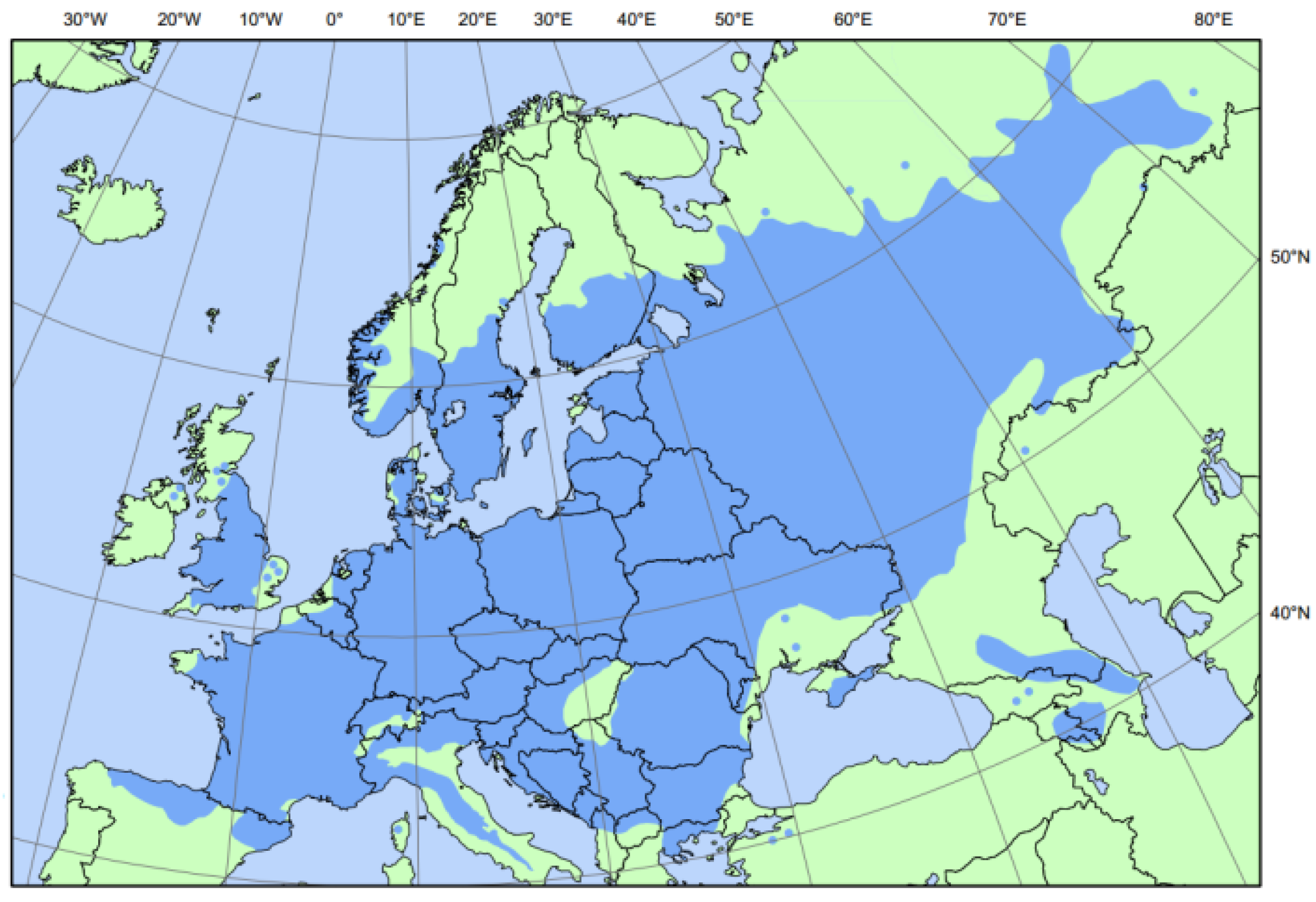
5. Distribution
5.1. Natural Areas
5.2. Local Distribution and Abundance
6. Silviculture
6.1. Mixed Stands and Ecosystem Services
6.2. Timber Production
6.2.1. Basic Wood Properties and Use
6.2.2. Bole Forming
6.2.3. Plantations
7. Tilia cordata and Global Change
7.1. Past Dominance and Decline
7.2. Current Climate Change
7.3. Impact of Climate Change on T. cordata
7.4. Potential Future Distribution
7.5. Opportunities
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bréda, N.; Badeau, V. Forest tree responses to extreme drought and some biotic events: Towards a selection according to hazard tolerance? Comptes Rendus Geosci. 2008, 340, 651–662. [Google Scholar] [CrossRef]
- Landmann, G.; Breda, N.; Houillier, F.; Dreyer, E.; Flot, J.-L. Sécheresse et canicule de l'été 2003: Quelles conséquences pour les forêts françaises? Rev. For. Fr. 2003, 55, 299–308. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Bradshaw, R.H.W.; Holmqvist, B.H.; Cowling, S.A.; Sykes, M.T. The effects of climate change on the distribution and management of Picea abies in southern Scandinavia. Can. J. For. Res. 2000, 30, 1992–1998. [Google Scholar] [CrossRef]
- Landmann, G. Le hêtre face aux changements climatiques. Le hêtre en France en 2100: La portion congrue? Forêt-Entreprise 2008, 180, 28–33. [Google Scholar]
- Leuschner, C.; Köckemann, B.; Buschmann, H. Abundance, niche breadth, and niche occupation of Central European tree species in the centre and at the margin of their distribution range. For. Ecol. Manag. 2009, 258, 1248–1259. [Google Scholar] [CrossRef]
- Randin, C.F.; Engler, R.; Normand, S.; Zappa, M.; Zimmermann, N.E.; Pearman, P.B.; Vittoz, P.; Thuillier, W.; Guisan, A. Climate change and plant distribution: Local models predict high-elevation persistence. Glob. Chang. Biol. 2009, 15, 1557–1569. [Google Scholar] [CrossRef]
- Pigott, D. Lime-trees and Basswoods: A Biological Monograph of the Genus Tilia, 1st ed.; Cambridge University Press: New York, NY, USA, 2012; p. 405. [Google Scholar]
- Radoglou, K.; Dobrowolska, D.; Spyroglou, G.; Nicolescu, V.-N. A review on the ecology and silviculture of limes (Tilia cordata Mill., Tilia platyphyllos Scop. and Tilia tomentosa Moench.) in Europe. Die BodenKult. 2009, 60, 9–20. [Google Scholar]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen, 6th ed.; Ulmer Verlag: Stuttgart, Germany, 2010; p. 1357. [Google Scholar]
- Hemery, E.G.; Clark, R.J.; Aldinger, E.; Claessens, H.; Malvolti, E.M.; O’connor, E.; Raftoyannis, Y.; Savill, S.P.; Brus, R. Growing scattered broadleaved tree species in Europe in a changing climate: A review of risks and opportunities. Forestry 2010, 83, 65–81. [Google Scholar] [CrossRef]
- Lestrade, M.; Gonin, P.; Coello, J. Autécologie du Tilleul à petites feuilles. Forêt-Entreprise 2013, 211, 6–11. [Google Scholar]
- Pigott, C.D.; Huntley, J.P. Factors controlling the distribution of Tilia cordata Mill. at the northern limits of its geographical range. IV. Estimated ages of the trees. III. Nature and cause of seed sterility. New Phytol. 1981, 87, 817–839. [Google Scholar] [CrossRef]
- Till, O. Uber die Frosthärte von Pflanzen sommergrüner Laubwälder. Flora 1956, 143, 499–542. [Google Scholar]
- Pigott, C.D. Tilia cordata Miller Biological Flora of the British Isles. J. Ecol. 1991, 79, 1147–1207. [Google Scholar] [CrossRef]
- Weissen, F.; Baix, P.; Boseret, J.; Bronchart, L.; Lejeune, M.; Maquet, P.; Marchal, D.; Marchal, J.; Masson, C.; Onclinckx, F.; et al. Le Fichier Écologique des Essences. Fasc. 2: Fiches Techniques, 1st ed.; Ministry of the Wallooon Region: Namur, Belgium, 1991; pp. 165–168. [Google Scholar]
- Köcher, P.; Horna, V.; Beckmeyer, I.; Leuschner, C. Hydraulic properties and embolism in small–diameter roots of five temperate broad–leaved tree species with contrasting drought tolerance. Ann. For. Sci. 2012, 69, 693–703. [Google Scholar] [CrossRef]
- Thompson, R.; Clark, R.M. Spatio-temporal modelling and assessment of within-species phenological variability using thermal time methods. Int. J. Biometeorol. 2006, 50, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Rameau, J.-C.; Mansion, D.; Dumé, G. Flore Forestière Française. Guide écologique illustré. Plaines et collines; Institut Développement Forestier: Paris, France, 1989; pp. 667–671. [Google Scholar]
- Lawesson, J.E.; Oksanen, J. Niche characteristics of Danish woody species as derived from coenoclines. J. Veg. Sci. 2002, 13, 279–290. [Google Scholar] [CrossRef]
- Pigott, C.D.; Pigott, S. Water as a determinant of the distribution of trees at the boundary of the Mediterranean zone. J. Ecol. 1993, 81, 557–566. [Google Scholar] [CrossRef]
- Aasamaa, K.; Sõber, A.; Hartung, W.; Niinemets, Ü. Drought acclimation of two deciduous tree species of different layers in a temperate forest canopy. Trees 2004, 18, 93–101. [Google Scholar]
- Pigott, C.D. Survival of mycorrhiza formed by Cenococcum geophilum Fr. in dry soils. New Phytol. 1982, 92, 513–517. [Google Scholar] [CrossRef]
- Kulagin, A.Y.; Shayakhmetov, I.F. Natural under-canopy regeneration and height-age structure of small-leaved linden (Tilia cordata Mill.) undergrowth in water-conservation forests around Pavlovskoe Reservoir, Ufa River. Russ. J. Ecol. 2007, 38, 247–252. [Google Scholar] [CrossRef]
- Lyr, H. Vergleichende untersuchungen zu physiologischen reaktionen auf wurzelanaerobiose bei Fagus sylvatica, Quercus robur und Tilia cordata. Beitr. Forstwirtsch. Landschaftsökologie 1993, 27, 18–23. [Google Scholar]
- Kozlowski, T.T. Responses of woody plants to flooding and salinity. Tree Physiol. Monogr. 1997, 1, 1–29. [Google Scholar] [CrossRef]
- Hauschild, R.; Hein, S. Zur Hochwassertoleranz von laubbäumen nach einem extremen Überflutungsereignis—Eine Fallstudie aus der südlichen Oberrheinaue. Allg. Forst Jagdztg. 2009, 180, 100–109. [Google Scholar]
- Schaffrath, J. Auswirkungen des extremen Sommerhochwassers des Jahres 1997 auf die Gehölzvegetation in der Oderaue bei Frankfurt (O.). Naturschutz Landschaftpflege Brandenbg. 2000, 9, 4–13. [Google Scholar]
- Späth, V. Hochwassertoleranz von Waldbäumen in der Rheinaue. AFZ DerWald 2002, 15, 807–810. [Google Scholar]
- Glenz, C.; Schlaepfer, R.; Iorgulescu, I.; Kienast, F. Flooding tolerance of Central European tree and shrub species. For. Ecol. Manag. 2006, 235, 1–13. [Google Scholar] [CrossRef]
- Coello, J.; Becquey, J.; Ortisset, J.-P.; Gonin, P.; Baiges, T.; Piqué, M. Les Tilleuls à Bois (Tilia Platyphyllos et T. Cordata); Centre de la Propietat Forestal: Santa Perpètua de Mogoda, Spain, 2013; pp. 54–61. [Google Scholar]
- Barengo, N.; Rudow, A.; Schwab, P. Projet Favoriser les Essences Rares: Tilleul à Petites Feuilles (Tilia Cordata Mill.) et Tilleul à Grandes Feuilles (Tilia Platyohyllos Scop.); OFEFP—Direction fédérale des forêts et EPFZ—Chaire de sylviculture: Zurich, Switzerland, 2001; pp. 1–8. [Google Scholar]
- Jaworski, A.; Kolodziej, Z.; Bartkowicz, L. Structure and dynamics of stands of primeval character composed of the little-leaf linden (Tilia cordata Mill.) in the “Las lipowy Obrozyska” reserve (southern Poland). J. For. Sci. 2005, 51, 283–304. [Google Scholar]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2008, 76, 521–547. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kull, O.; Tenhunen, J. Variability in leaf morphology and chemical composition as a function of canopy light environment in coexisting deciduous trees. Int. J. Plant Sci. 1999, 160, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Wiley-VCH Verlag: Cambridge, UK, 1988; Volume 101, p. 756. [Google Scholar]
- Diekmann, M. Ecological behaviour of deciduous hardwood trees in boreo-nemoral Sweden in relation to light and soil conditions. For. Ecol. Manag. 1996, 86, 1–14. [Google Scholar] [CrossRef]
- Seppä, H.; Alenius, T.; Bradshaw, R.H.W.; Giesecke, T.; Heikkilä, M.; Muukkonen, P. Invasion of Norway spruce (Picea abies) and the rise of the boreal ecosystem in Fennoscandia. J. Ecol. 2009, 97, 629–640. [Google Scholar] [CrossRef]
- Pigott, C.D. Factors controlling the distribution of Tilia cordata Mill. at the northern limits of its geographical range. IV. Estimated ages of the trees. New Phytol. 1989, 112, 117–121. [Google Scholar] [CrossRef]
- Mayer, H. Waldbau auf Soziologisch-Ökologischer Grundlage, 4th ed.; Fischer Verlag: Stuttgart, Germany, 1992; p. 522. [Google Scholar]
- Wunder, J.; Brzeziecki, B.; Żybura, H.; Reineking, B.; Bigler, C.; Bugmann, H. Growth–mortality relationships as indicators of life-history strategies: A comparison of nine tree species in unmanaged European forests. Oikos 2008, 117, 815–828. [Google Scholar] [CrossRef]
- Böckmann, T. Wachstum und Ertrag der Winterlinde (Tilia cordata Mill.) in Niedersachsen und Nordhessen. Ph.D. Thesis, Universität Göttingen, Göttingen, Germany, 1990. [Google Scholar]
- Magyar, J. Hársaink dendrometriai alapmércéi. Erdögasdaság Faip. (ERFA) 1978, 31, 10–12. [Google Scholar]
- Czuraj, M. Tablice Zasobności i Przyrostu Drzewostanów; Państwowe Wydawnictwo Rolnicze i Leśne: Warzawa, Poland, 1990; Volume 165, pp. 120–123. [Google Scholar]
- Tyurin, A.V.; Naumenko, I.M.; Voropanov, P.V. Lesnaya Vspomogatel’naya Knizhka (po Taksatii Lesa); Goslesbumizdat: Moscow, Russia, 1956; p. 532. [Google Scholar]
- Shvydenko, A.S.; Strochynskyy, A.A.; Savych, Y.N. Normativno-Spravochnyje Materialy dla Taksaziji Lesov Ukrainy i Moldavii; Urozhaj Verlag: Kiev, Ukraine, 1987; pp. 152–153. [Google Scholar]
- Hein, S.; Collet, C.; Ammer, C.; Goff, N.L.; Skovsgaard, J.P.; Savill, P. A review of growth and stand dynamics of Acer pseudoplatanus L. in Europe: Implications for silviculture. Forestry 2009, 82, 361–385. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Hein, S.; Oosterbaan, A.; Wagner, S.; Clark, J.; Skovsgaard, J.P. A review of European ash (Fraxinus excelsior L.): Implications for silviculture. Forestry 2011, 84, 133–148. [Google Scholar] [CrossRef]
- Claessens, H.; Oosterbaan, A.; Savill, P.; Rondeux, J. A review of the characteristics of black alder (Alnus glutinosa (L.) Gaertn.) and their implications for silvicultural practices. Forestry 2010, 83, 163–175. [Google Scholar] [CrossRef]
- Röös, M. Zum Wachstum der Vogelkirsche (Prunus avium L.) in Nordrhein-Westfalen und angrenzenden Gebieten. Ph.D. Thesis, Universität Göttingen, Göttingen, Germany, 1990. [Google Scholar]
- Hein, S.; Spiecker, H. Controlling diameter growth of common ash, sycamore and wild cherry. In Valuable Broadleaved Forests in Europe; European Forest Research Institute: Freiburg, Germany, 2009; Volume 22, pp. 123–147. [Google Scholar]
- Semaškienė, L. Small-Leaved Lime (Tilia Cordata Mill.) in Lithuania: Phenotypical Diversity and Productivity of Modal Stands. Ph.D. Thesis, Lithuanian University of Agriculture, Kaunas, Lithuania, 2006. [Google Scholar]
- Schober, R. Ertragstafeln Wichtiger Baumarten bei Verschiedener Durchforstung; J.D. Sauerländer’s Verlag: Frankfurt, Germany, 1995; p. 166. [Google Scholar]
- Glavac, V. Über höhenwuchsleistung und wachstungoptimum der schwarzerle auf vergleichbaren standorten in nord-, mittel- und südeuropa. Schriftenreihe forstl. Fak. Univ. Göttingen 1972, 45, 1–61. [Google Scholar]
- Rossi, R. Ricerca sulla crescita e l’attitudine selvicolturale di piantagionii di tiglio (Tilia cordata Mill.) e tiglio nostrano (Tiglia platyphyllos Scop.). Schweiz. Z. Forstwes. 1993, 144, 627–637. [Google Scholar]
- Trauboth, V. Anbau der winterlinde (Tilia cordata Mill.) im vorland des nordwestlichen thüringer waldes. Forst Holz 2005, 60, 329–334. [Google Scholar]
- Erteld, W. Über die wachstumsentwicklung der linde. Arch. Forstwes. 1963, 12, 1152–1158. [Google Scholar]
- Koop, H. Forest Dynamics. SILVI-STAR: A Comprehensive Monitoring System, 1st ed.; Springer Verlag: Berlin, Germany, 1989; p. 229. [Google Scholar]
- Hein, S.; Ehring, A.; Kohnle, U. Wachstumskundliche Grundlagen für die Wertholzproduktion mit der edelkastanie (Castanea sativa Mill.). Allg. Forst Jagdztg. 2014, 185, 1–16. [Google Scholar]
- Larsen, F.K.; Kristoffersen, P. Tilia’s physical diemensions over time. J. Arboric. 2002, 28, 209–213. [Google Scholar]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Rötzer, T.; Caldentey, J.; Koike, T.; van Con, T.; Chavanne, A.; Seifert, T.; et al. Crown size and growing space requirement of common tree species in urban centres, parks, and forests. Urban For. Urban Green. 2015, 14, 466–479. [Google Scholar] [CrossRef]
- Sæbø, A.; Benedikz, T.; Randrup, T.B. Selection of trees for urban forestry in the Nordic countries. Urban For. Urban Green. 2003, 2, 101–114. [Google Scholar] [CrossRef]
- Spiecker, H.; Hein, S.; Makkonen-Spiecker, K.; Thies, M. Valuable Broadleaved Forests in Europe. European Forest Institute Research Reports; BRILL: Leiden, The Netherlands, 2009; Volume 22, pp. 123–147. [Google Scholar]
- Savill, S.P. The Silviculture of Trees Used in British Forestry; CAB International: Wallingford, UK, 1991; p. 143. [Google Scholar]
- Tal, O. Comparative Flowering Ecology of Fraxinus Excelsior, Acer Platanoides, Acer Pseudoplatanus and Tilia cordata in the Canopy of Leipzig’s Floodplain Forest. Ph.D. Thesis, University of Leipzig, Leipzig, Germany, 2006. [Google Scholar]
- Anderson, G.J. The pollination biology of Tilia. Am. J. Bot. 1976, 63, 1203–1212. [Google Scholar] [CrossRef]
- Kelly, D.; Sork, V.L. Mast seeding in perennial plants: Why, how, where? Annu. Rev. Ecol. Syst. 2002, 33, 427–447. [Google Scholar] [CrossRef]
- Gellini, R.; Grossoni, P. Botanica Forestale—II: Angiosperme, 2nd ed.; CEDAM: Padova, Italy, 1998; pp. 208–210. [Google Scholar]
- Suszka, B.; Muller, C.; Bonnet-Masimbert, M. Seeds of Forest Broadleaves from Harvest to Sowing; Wydawnictwo naukowe: Warszawa-Poznan, Poland, 1994; p. 299. [Google Scholar]
- Browse, P.D.A.M. Hardy, Woody Plants from Seed; Grower Books: London, UK, 1979; p. 163. [Google Scholar]
- Heit, C. Propagation from seed: 7. Successful propagation of six hardseeded group species. Am. Nurserym. 1967, 125, 10–45. [Google Scholar]
- Cistyakova, A.A. Bolsoj ziznennyj cikl Tilia cordata Mill. [High life cycle of Tilia cordata Mill.]. Mosk. Obsc. Ispyt. Prir. Otd. Biol. 1979, 84, 85–98. [Google Scholar]
- Cistyakova, A.A. Biologiceskije osobennosti vegetativnogo vozobnovienia osnovnych porod v sirokolistvennych lesach [Biological features of vegetative reproduction of basic rocks in deciduous forests]. Lesovedenie 1982, 2, 11–17. [Google Scholar]
- Pawlaczyk, P. Wegetatywne odnowienie lipy drobnolistnej (Tilia cordata Mill.) i jego znaczenie ekologiczne w grądzie w Białowieskim Parku Narodowym. Phytocoen. Semin. Geobot. 1991, 3, 161–171. [Google Scholar]
- Koop, H. Vegetative reproduction of trees in some European natural forests. Vegetatio 1987, 72, 103–110. [Google Scholar]
- Schieber, B.; Janík, R.; Snopková, Z. Phenology of four broad-leaved forest trees in a submountain beech forest. J. For. Sci. 2009, 55, 15–22. [Google Scholar]
- Wesołowski, T.; Rowiński, P. Timing of bud burst and tree-leaf development in a multispecies temperate forest. Forest Ecol. Manag. 2006, 237, 387–393. [Google Scholar] [CrossRef]
- Kramer, K. A modelling analysis of the effects of climatic warming on the probability of spring frost damage to tree species in The Netherlands and Germany. Plant Cell Environ. 1994, 17, 367–377. [Google Scholar] [CrossRef]
- Caffarra, A.; Donnelly, A. The ecological significance of phenology in four different tree species: Effects of light and temperature on bud burst. Int. J. Biometeorol. 2011, 55, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Spano, D.; Cesaraccio, C.; Duce, P.; Snyder, R.L.; Botarelli, L.; Pratizzoli, W.; Sacchetti, V. Observations and prediction of phenological trends in two Italian phenological gardens. Ital. J. Agrometeorol. 2007, 12, 13–18. [Google Scholar]
- Differt, J. Phénologie des Espèces Arborées. Synthèse Bibliographique. Analyse des Données du Réseau National de Suivi à long Terme des Ecosystèmes Forestiers (RENECOFOR). Rapport Scientifique; UMR ENGREF- INRA-LERFORB: Nancy, France, 2001; p. 97. [Google Scholar]
- Noirfalise, A. Forêts et Stations Forestières en Belgique, 1st ed.; Les Presses Agronomiques de Gembloux: Gembloux, Belgium, 1984; p. 236. [Google Scholar]
- Kirichenko, N.I. The lime leafminer Phyllonorycter issikii in Western Siberia: Some ecological characteristics of the population of the recent invader. Contemp. Probl. Ecol. 2014, 7, 114–121. [Google Scholar] [CrossRef]
- Brasier, C. Phytophthora Pathogens of Trees: Their Rising Profile in Europe; Forestry Commission: Edinburgh, UK, 1999. [Google Scholar]
- Van Lerberghe, P.; Balleux, P. Lutter contre les dégats du gibier dans les plantations forestières. Les types de dégats et leurs conséquences. Forêt Wallonne 1999, 42, 8–13. [Google Scholar]
- Pigott, C.D. Natural regeneration of Tilia cordata in relation to forest-structure in the forest of Bialowieza, Poland. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1975, 270, 151–179. [Google Scholar] [CrossRef]
- Gill, R.M.A. A review of damage by mammals in north temperate forests: 1. Deer. Forestry 1992, 65, 145–169. [Google Scholar] [CrossRef]
- Svejgaard Jensen, J. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Lime (Tilia spp.); International Plant Genetic Resources Institute: Rome, Italy, 2003; p. 6. [Google Scholar]
- Logan, S.A.; Phuekvilai, P.; Wolff, K. Ancient woodlands in the limelight: Delineation and genetic structure of ancient woodland species Tilia cordata and Tilia platyphyllos (Tiliaceae) in the UK. Tree Genet. Genomes 2015, 11, 1–12. [Google Scholar] [CrossRef]
- Phuekvilai, P. Relicts, Refugia, and Reticulation: A Study of Population History, Hybrids and Phylogeny in the Long-Lived Flowering Tree Genus Tilia. Ph.D. Thesis, Newcastle University, Newcastle, UK, 2014. [Google Scholar]
- Myking, T. Evaluating genetic resources of forest trees by means of life history traits—A Norwegian example. Biodivers. Conserv. 2002, 11, 1681–1696. [Google Scholar] [CrossRef]
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen maps for Europe: 0–13000 BP; Cambridge University Press: Cambridge, UK, 1983; p. 667. [Google Scholar]
- Turok, J.; Erikson, G.; Kleinschmit, J.; Canger, S. Report of the First Meeting; International Plant Genetic Resources Institute: Rome, Italy, 1996; p. 172. [Google Scholar]
- Hemery, G.; Spiecker, H.; Aldinger, E.; Kerr, G.; Collet, C.; Bell, S. COST Action E42: Growing Valuable Broadleaved Tree Species. Final Report. Available online: http://www.valbro.uni-freiburg.de/pdf/COST_E42_Final_Report_2008.pdf (accessed on 28 January 2016).
- Koss, H.; Fricke, O. Die Entwicklung von linden als unterbau in stieleichenbeständen [Development of Tilia cordata understorey in oak stands.]. Forstarchiv 1982, 53, 60–66. [Google Scholar]
- Hagen-Thorn, A.; Armolaitis, K.; Callesen, I.; Stjernquist, I. Macronutrients in tree stems and foliage: A comparative study of six temperate forest species planted at the same sites. Ann. For. Sci. 2004, 61, 489–498. [Google Scholar] [CrossRef]
- Langenbruch, C.; Helfrich, M.; Flessa, H. Effects of beech (Fagus sylvatica), ash (Fraxinus excelsior) and lime (Tilia spec.) on soil chemical properties in a mixed deciduous forest. Plant Soil 2012, 352, 389–403. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Callesen, I.; Armolaitis, K.; Nihlgård, B. The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For. Ecol. Manag. 2004, 195, 373–384. [Google Scholar] [CrossRef]
- Tokár, F.; Bolvanský, M. Effects of Thinning on Growth and Productivity of Pure and Mixed European Chestnut Stands [Vpliv Redcenj na Rast in Produktivnost Cistih ter Mesanih Sestojev Pravega Kostanja]; Zbornik Gozdarstva in Lesarstva: Ljubljana, Slovenia, 2002; pp. 97–111. [Google Scholar]
- Ziemer, R.R. The Role of Vegetation in the Stability of Forested Slopes. In Proceedings of the International Union of Forestry Research Organizations, XVII World Congress, Kyoto, Japan, 6–17 September 1981; IUFRO: Kyoto, Japan, 1981; pp. 297–308. [Google Scholar]
- Rowe, D.B.; Blazich, F.A.; Tilia, L. Linden or basswood. In The Woody Plant Seed Manual; U.S. Department of Agriculture: Washington, DC, USA, 2008; pp. 113–118. [Google Scholar]
- Popescu, M.C.; Lisa, G.; Froidevaux, J.; Navi, P.; Popescu, C.M. Evaluation of the thermal stability and set recovery of thermo-hydro-mechanically treated lime (Tilia cordata) wood. Wood Sci. Technol. 2014, 48, 85–97. [Google Scholar] [CrossRef]
- Löf, M.; Karlsson, M.; Sonesson, K.; Welander, T.N.; Collet, C. Growth and mortality in underplanted tree seedlings in response to variations in canopy closure of Norway spruce stands. Forestry 2007, 80, 371–383. [Google Scholar] [CrossRef]
- Latałowa, M.; Pedziszewska, A.; Maciejewska, E.; Świeta-Musznicka, J. Tilia forest dynamics, Kretzschmaria deusta attack, and mire hydrology as palaeoecological proxies for mid-Holocene climate reconstruction in the Kashubian Lake District (N Poland). Holocene 2013, 23, 667–677. [Google Scholar] [CrossRef]
- Giesecke, T. Holocene dynamics of the southern boreal forest in Sweden. Holocene 2005, 15, 858–872. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2014: Synthesis Report of the IPCC Fifth Assessment Report (AR5); Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; p. 1132. [Google Scholar]
- Latte, N.; Lebourgeois, F.; Claessens, H. Increased tree-growth synchronization of beech (Fagus sylvatica L.) in response to climate change in northwestern Europe. Dendrochronologia 2015, 33, 69–77. [Google Scholar] [CrossRef]
- Walther, G.-R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.H.; Carey, P.D.; Combes, J. First leafing dates of trees in Surrey between 1947 and 1996. Lond. Nat. 1997, 76, 15–20. [Google Scholar]
- Juknys, R.; Sujetoviene, G.; Žeimavičius, K.; Šveikauskaite, I. Comparison of climate warming induced changes in silver birch (Betula pendula Roth) and lime (Tilia cordata Mill.) phenology. Balt. For. 2012, 18, 25–32. [Google Scholar]
- Donnelly, A.; Salamin, N.; Jones, M. Changes in tree phenology: An indicator of spring warming in Ireland? Biol. Environ. Proc. R. Ir. Acad. 2006, 106, 49–56. [Google Scholar] [CrossRef]
- Ahas, R.; Aasa, A. The effects of climate change on the phenology of selected Estonian plant, bird and fish populations. Int. J. Biometeorol. 2006, 51, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Veriankaitė, L.; Šaulienė, I.; Bukantis, A. The modelling of climate change influence on plant flowering shift in Lithuania. Žemdirbystė (Agriculture) 2010, 97, 41–48. [Google Scholar]
- Chmielewski, F.-M.; Rötzer, T. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavska, O.G.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Minin, A.A.; Voskova, A.V. Homeostatic responses of plants to modern climate change: Spatial and phenological aspects. Russ. J. Dev. Biol. 2014, 45, 127–133. [Google Scholar] [CrossRef]
- Faliński, J.B. Vegetation Dynamics in Temperate Lowland Primeval Forests. Ecological Studies in Białowieża Forest; Junk Publishers: Dordrecht, The Netherlands, 1986; Volume 8, p. 537. [Google Scholar]
- Koca, D.; Smith, B.; Sykes, M. Modelling regional climate change effects on potential natural ecosystems in Sweden. Clim. Chang. 2006, 78, 381–406. [Google Scholar] [CrossRef]
- Sykes, M.T.; Prentice, I.C.; Cramer, W. A bioclimatic model for the potential distributions of north European tree species under present and future climates. J. Biogeogr. 1996, 23, 203–233. [Google Scholar]
- Löf, M.; Boman, M.; Brunet, J.; Hannerz, M.; Mattsson, L.; Nylinder, M. Broadleaved forest management for multiple goals in southern Sweden—An overview including future research prospects. Ecol. Bull. 2010, 53, 235–245. [Google Scholar]
- Attorre, F.; Alfò, M.; de Sanctis, M.; Francesconi, F.; Valenti, R.; Vitale, M.; Bruno, F. Evaluating the effects of climate change on tree species abundance and distribution in the Italian peninsula. Appl. Veg. Sci. 2011, 14, 242–255. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Bachand, R.R. Lodgepole pine (Pinus contorta) ecotones in rocky mountain national park, Colorado, USA. Ecology 1997, 78, 632–641. [Google Scholar] [CrossRef]
- Thompson, I.; Mackey, B.; McNulty, S.; Mosseler, A. Forest Resilience, Biodiversity and Climate Change. A Synthesis of the Biodiversity/Resilience/Stability Relationship in Forest Ecosystems; Secretariat of the Convention on Biological Diversity: Montreal, Canada, 2009; pp. 1–67. [Google Scholar]
- Ray, D.; Morison, J.; Broadmeadow, M. Climate Change: Impacts and Adaptation in England’s Woodlands; Forestry Commission England: Edinburg, UK, 2010; p. 16. [Google Scholar]
- Lindner, M.; Garcia-Gonzalo, J.; Kolström, M.; Green, T.; Reguera, R. Impacts of Climate Change on European Forests and Options for Adaptation; European Forest Institute: Joensuu, Finland, 2008. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Yield Tables | ||
| Country [Reference] | Parameter | Range of Value |
| Germany [45] | Range of max CAVI and corresponding ages | 11.9 to 20.2 m3·ha−1·years−1 30 to 45 years |
| Range of max MAVI and corresponding ages | 10 to 11.5 m3·ha−1·years−1 60 to 100 years | |
| Poland [47] | Max CAVI and corresponding age | 6.2 m3·ha−1·years−1 at 35 years |
| Max MAVI and corresponding age | 4.7 m3·ha−1·years−1 at 35 years | |
| Local Observations | ||
| Country | Parameter | Range of Value |
| Sweden, Lithuania and Denmark [67] | CAVI and corresponding age | 10 m3·ha−1·years−1 at 35 years |
| Slovakia [68] | CAVI and corresponding age | 5–11 m3·ha−1·years−1 at 30 years |
| Poland, unmanaged forest [36] | Standing volume | 760–860 m3·ha−1 |
| Russia [48] | Standing volume | 594–810 m3·ha−1 at 100–200 years |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Jaegere, T.; Hein, S.; Claessens, H. A Review of the Characteristics of Small-Leaved Lime (Tilia cordata Mill.) and Their Implications for Silviculture in a Changing Climate. Forests 2016, 7, 56. https://doi.org/10.3390/f7030056
De Jaegere T, Hein S, Claessens H. A Review of the Characteristics of Small-Leaved Lime (Tilia cordata Mill.) and Their Implications for Silviculture in a Changing Climate. Forests. 2016; 7(3):56. https://doi.org/10.3390/f7030056
Chicago/Turabian StyleDe Jaegere, Tanguy, Sebastian Hein, and Hugues Claessens. 2016. "A Review of the Characteristics of Small-Leaved Lime (Tilia cordata Mill.) and Their Implications for Silviculture in a Changing Climate" Forests 7, no. 3: 56. https://doi.org/10.3390/f7030056