
Ecofunctional Traits and Biomass Production in Leguminous Tree Species under Fertilization Treatments during Forest Restoration in Amazonia

 ,
,
Abstract
:1. Introduction
2. Experimental Section
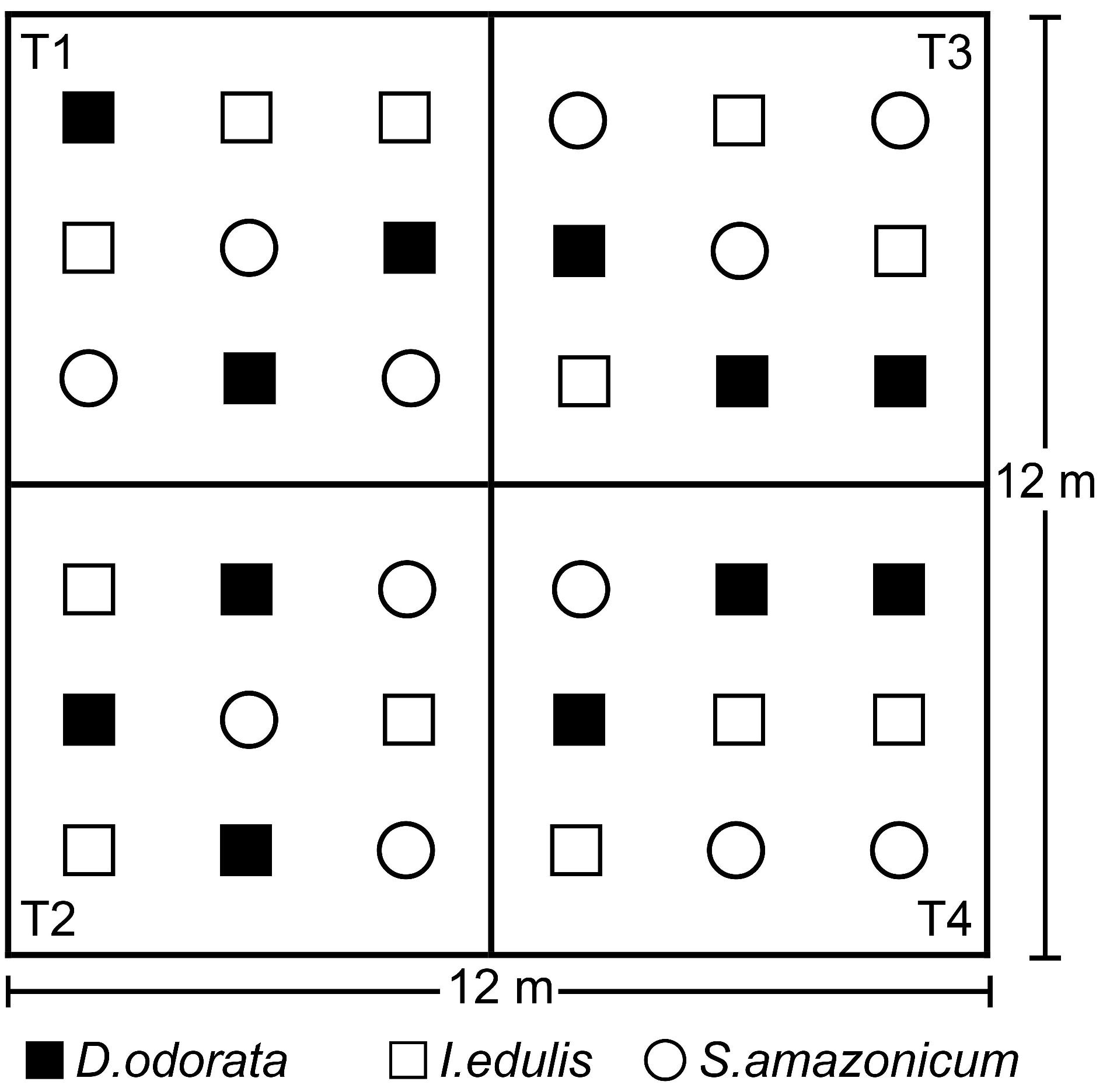
2.1. Study Site, Leguminous Species and Fertilization Treatments
2.2. Soil Fertility Analysis
2.3. Relative Height and Diameter Growth Rates
2.4. Biomass Production and Allocation
2.5. Leaf Nutrient Content
2.6. Photosynthesis and Photosynthetic Nutrient Use Efficiency
2.7. Data Analysis
3. Results and Discussion
3.1. Soil Fertility
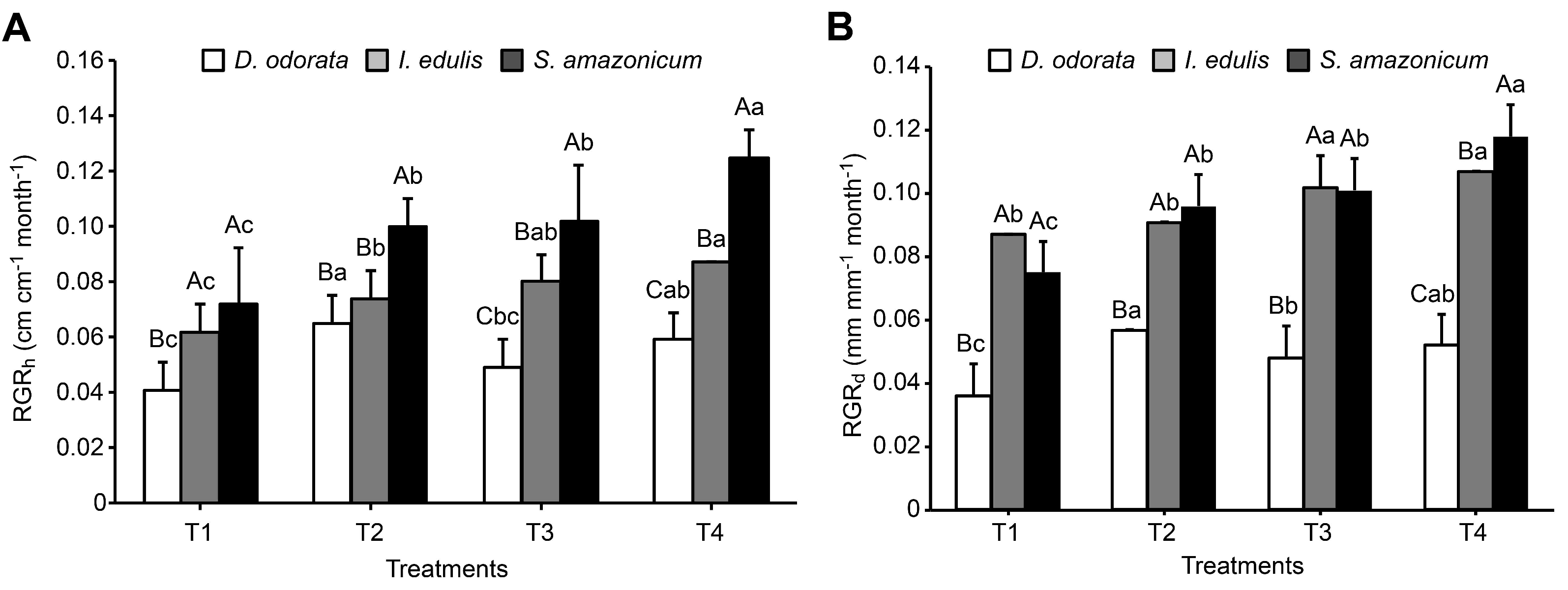
3.2. Plant Growth and Biomass Allocation
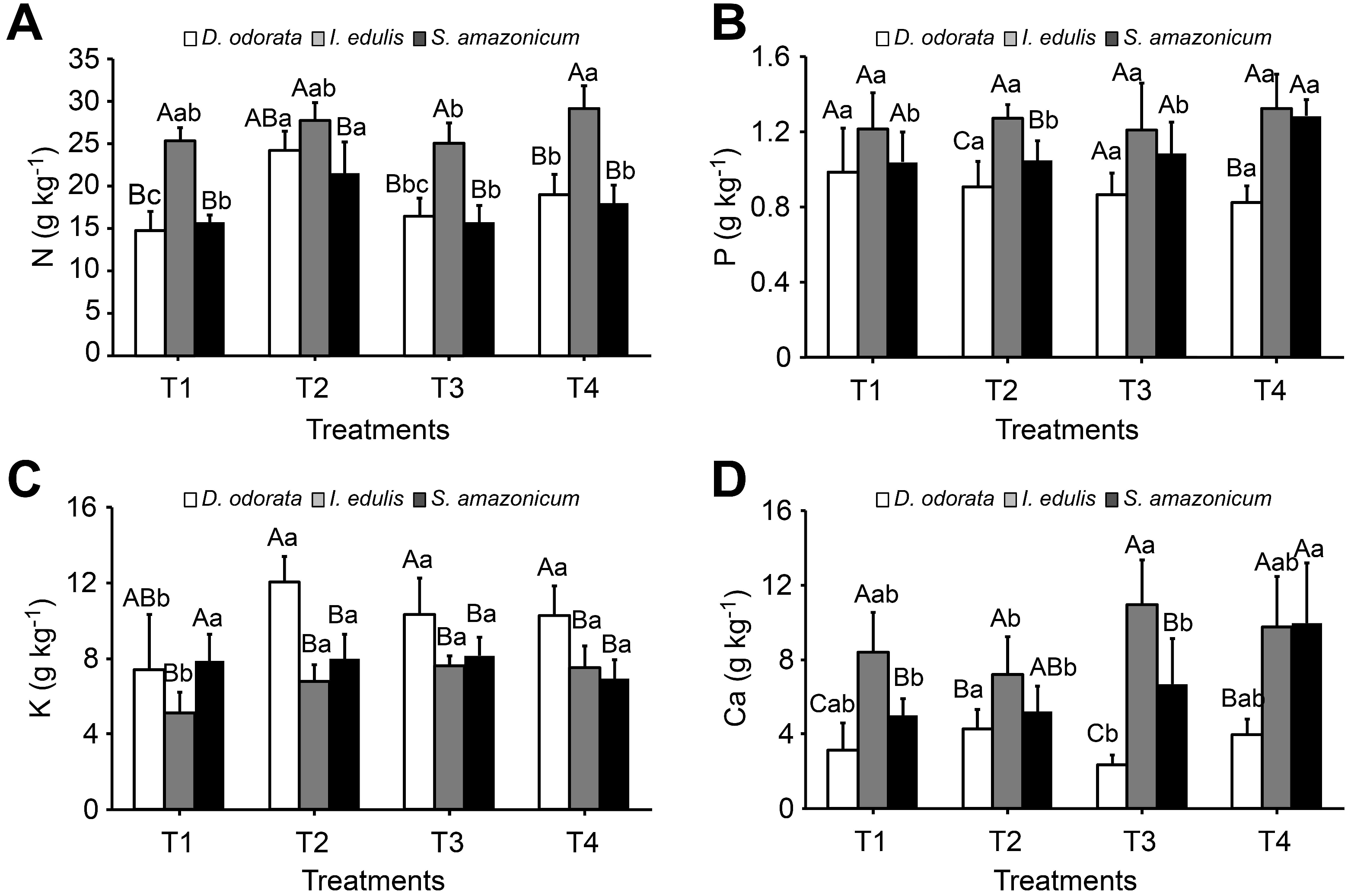
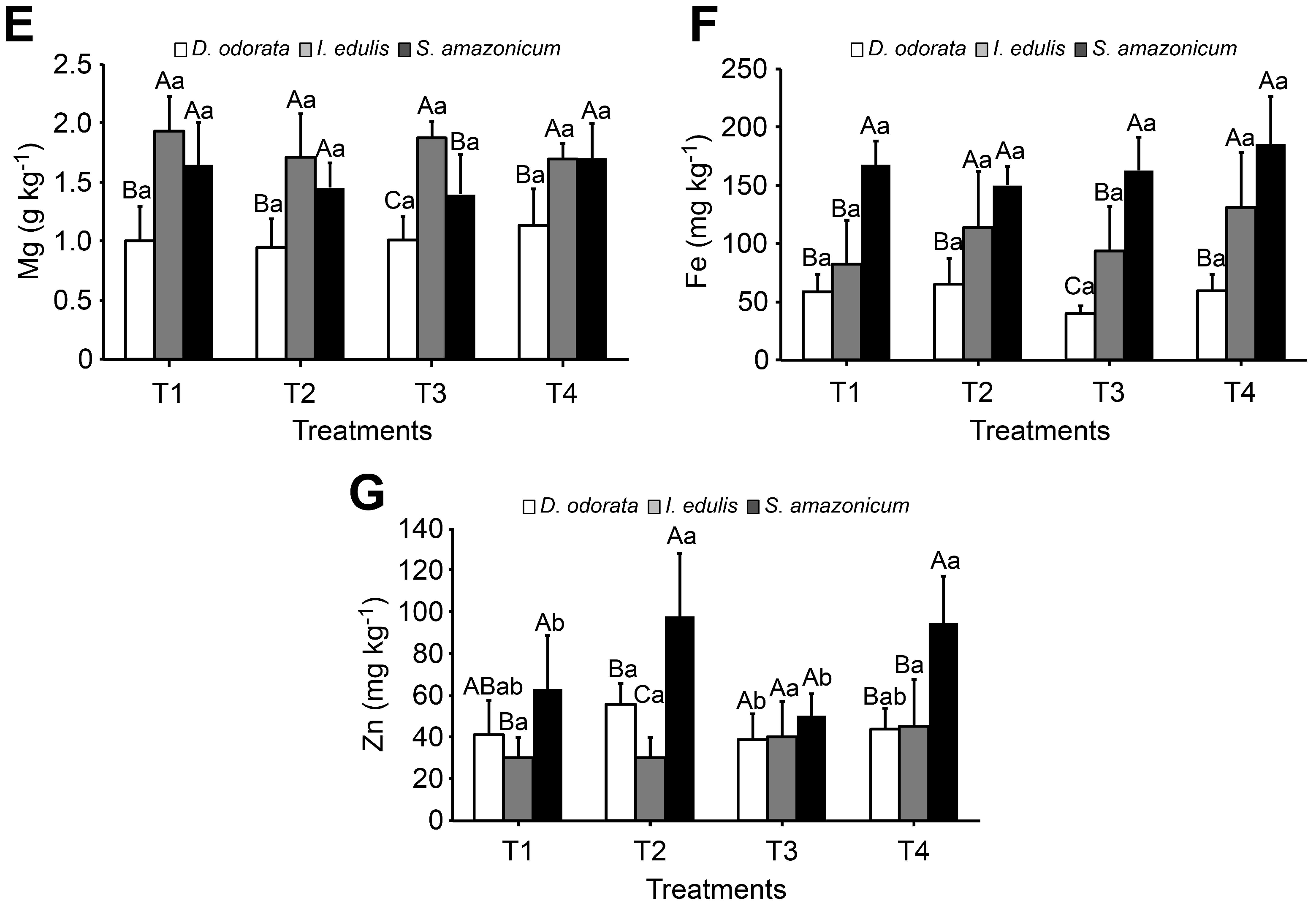
3.3. Leaf Nutrient Content
3.4. Photosynthesis and Photosynthetic Nutrient Use Efficiency
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Souza, C.R.; de Azevedo, C.P.; Lima, R.M.; Rossi, L.M.B. Comportamento de espécies florestais Em plantios a pleno Sol e Em faixas de enriquecimento de Capoeira na Amazônia. Acta Amazonica 2010, 40, 127–134. [Google Scholar] [CrossRef]
- Davidson, E.A.; Araújo, A.C.; Artaxo, P.; Balch, J.K.; Brown, I.F.; Bustamante, M.M.; Coe, M.T.; DeFries, R.S.; Keller, M.; Longo, M.; et al. The Amazon basin in transition. Nature 2012, 481, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Pesquisas Espaciais—INPE. PROJETO PRODES. Available online: http://www.obt.inpe.br/prodes/index.php (accessed on 22 March 2015).
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of degraded tropical forest Landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Kelty, M.J. The role of species mixtures in Plantation Forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Lamb, D. Forest restoration—The third big Silvicultural Challenge. J. Trop. For. Sci. 2012, 24, 295–299. [Google Scholar]
- Van Haren, J.; de Oliveira, R.C.; Beldini, P.T.; de Camargo, P.B.; Keller, M.; Saleska, S. Tree species effects on soil Properties and Greenhouse Gas Fluxes in East-Central Amazonia: Comparison between Monoculture and Diverse Forest. Biotropica 2013, 45, 709–718. [Google Scholar] [CrossRef]
- Lima, T.A.; Vieira, G. High plant species richness in Monospecific Tree Plantations in the Central Amazon. For. Ecol. Manag. 2013, 295, 77–86. [Google Scholar] [CrossRef]
- Marenco, R.A.; Gonçalves, J.F.C.; Vieira, G. Photosynthesis and leaf nutrient contents in Ochroma Pyramidale. Photosynthetica 2001, 39, 539–543. [Google Scholar] [CrossRef]
- Gonçalves, J.F.C.; Santos Junior, U.M. Utilization of the chlorophyll a fluorescence technique as a Tool for Selecting Tolerant Species to Environments of High Irradiance. Braz. J. Plant Physiol. 2005, 17, 307–313. [Google Scholar] [CrossRef]
- Santos Junior, U.M.; Gonçalves, J.F.C.; Feldpausch, T.R. Growth, leaf nutrient concentration and Photosynthetic Nutrient Use Efficiency in Tropical Tree Species Planted in Degraded Areas in Central Amazonia. For. Ecol. Manag. 2006, 226, 299–309. [Google Scholar] [CrossRef]
- Quesada, C.A.; Phillips, O.L.; Schwarz, M.; Czimczik, C.I.; Baker, T.R.; Patiño, S.; Fyllas, N.M.; Hodnett, M.G.; Herrera, R.; Almeida, S.; et al. Basin-wide variations in Amazon Forest Structure and Function Are Mediated by Both Soils and Climate. Biogeosciences 2012, 9, 2203–2246. [Google Scholar] [CrossRef] [Green Version]
- Fageria, N.K.; Baligar, V.C.; Li, Y.C. The role of nutrient efficient plants in Improving Crop Yields in the Twenty First Century. J. Plant Nutr. 2008, 31, 1121–1157. [Google Scholar] [CrossRef]
- Denich, M.; Vlek, P.; Deabreusa, T.; Vielhauer, K.; Lucke, W. A concept for the development of fire-free Fallow Management in the Eastern Amazon, Brazil. Agric. Ecosyst. Environ. 2005, 110, 43–58. [Google Scholar] [CrossRef]
- Siddique, I.; Engel, V.L.; Parrotta, J.A.; Lamb, D.; Nardoto, G.B.; Ometto, J.P.; Martinelli, L.A.; Schmidt, S. Dominance of legume trees alters nutrient Relations in Mixed Species Forest Restoration Plantings within Seven Years. Biogeochemistry 2008, 88, 89–101. [Google Scholar] [CrossRef]
- De Faria, S.M.; Diedhiou, A.G.; de Lima, H.C.; Ribeiro, R.D.; Galiana, A.; Castilho, A.F.; Henriques, J.C. Evaluating the nodulation status of leguminous species from the Amazonian Forest of Brazil. J. Exp. Bot. 2010, 61, 3119–3127. [Google Scholar] [CrossRef] [PubMed]
- Brienza Junior, J.S.; Oliveira, R.P.; Denich, M.; Vlek, P.L.G. Plantio de Árvores de Crescimento Rápido para Recuperação de Áreas Agrícolas na Amazônia Oriental Brasileira: Estudo de Caso Com Produção de Milho e Mandioca. Braz. J. For. Res. 2011, 31, 347–353. [Google Scholar] [CrossRef]
- Joslin, A.H.; Markewitz, D.; Morris, L.A.; DeAssis Oliveira, F.; Figueiredo, R.O.; Kato, O.R. Five native tree species and Manioc under Slash-And-Mulch Agroforestry in the Eastern Amazon of Brazil: Plant Growth and Soil Responses. Agrofor. Syst. 2011, 81, 1–14. [Google Scholar] [CrossRef]
- Franco, A.A.; de Faria, S.M. The contribution of n2-fixing tree Legumes to Land Reclamation and Sustainability in the Tropics. Soil Biol. Biochem. 1997, 29, 897–903. [Google Scholar] [CrossRef]
- Sprent, J.I.; Parsons, R. Nitrogen fixation in legume and Non-Legume Trees. Field Crops Res. 2000, 65, 183–196. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Mitchell, P.A.; Brockwell, J. Productivity of three young Mixed-Species Plantations Containing N2-Fixing Acacia and Non-N2-Fixing Eucalyptus And Pinus Trees in Southeastern Australia. For. Sci. 2007, 53, 426–434. [Google Scholar]
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient use efficiency in plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Nichols, J.D.; Carpenter, F.L. Interplanting Inga Edulis yields nitrogen benefits to Terminalia Amazonia. For. Ecol. Manag. 2006, 233, 344–351. [Google Scholar] [CrossRef]
- Jaquetti, R.K.; Gonçalves, J.F.C.; Ferraz, J.B.S.; Ferreira, M.J.; Santos Junior, U.M.; Lacerda, C.F. Green fertilization enhances the photosynthetic performance and the Growth of Leguminous Trees for Restoration Plantation in Central Amazon. Am. J. Plant Sci. 2014, 5, 2497–2508. [Google Scholar] [CrossRef]
- Silva, A.K.L.; Vasconcelos, S.S.; de Carvalho, C.J.R.; Cordeiro, I.M.C.C. Litter dynamics and fine root Production in Schizolobium Parahyba Var. Amazonicum Plantations and Regrowth Forest in Eastern Amazon. Plant Soil 2011, 347, 377–386. [Google Scholar]
- Siviero, M.A.; Motta, A.M.; dos Lima, D.S.; Birolli, R.R.; Huh, S.Y.; Santinoni, I.A.; Murate, L.S.; de Castro, C.M.A.; Miyauchi, M.Y.H.; Zangaro, W.; et al. Interaction among N-fixing bacteria and am fungi in Amazonian Legume Tree (Schizolobium Amazonicum) in Field Conditions. Appl. Soil Ecol. 2008, 39, 144–152. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis, Part 3: Chemical Methods; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Miyazawa, M.; Pavan, M.A.; Muraoka, T.; Carmo, F.S.; Mello, W.J. Análise química de tecidos vegetais. In Manual de Análise Química de Solos, Plantas e Fertilizantes; Silva, F.C., Ed.; EMBRAPA: Brasília, Brazil, 2009. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution reagent for the Determination of Phosphate in Natural Waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Hunt, R. Basic Growth Analysis; Edward Arnold: London, UK, 1990; p. 112. [Google Scholar]
- Zhao, D.; Oosterhuis, D.M.; Bednarz, C.W. Influence of potassium deficiency on Photosynthesis, Chlorophyll Content, and Chloroplast Ultrastructure of Cotton Plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Schroth, G.; D’Angelo, S.A.; Teixeira, W.G.; Haag, D.; Lieberei, R. Conversion of secondary Forest into Agroforestry and Monoculture Plantations in Amazonia: Consequences for Biomass, Litter and Soil Carbon Stocks after 7 Years. For. Ecol. Manag. 2002, 163, 131–150. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to Leaves, Stems and Roots: Meta-Analyses of Interspecific Variation and Environmental Control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Vitti, G.C.; Ferreira, A.C. Síntese de Análises Químicas em Tecido Vegetal; Escola Superior de Agricultura Luiz de Queiroz: Piracicaba, Brazil, 1997. [Google Scholar]
- EMBRAPA. Manual de Análises Químicas de Solos, Plantas e Fertilizantes; EMBRAPA: Brasília, Brazil, 2009; p. 370. [Google Scholar]
- Santos Junior, U.M.; Gonçalves, J.F.C.; Fearnside, P.M. Measuring the impact of Flooding on Amazonian Trees: Photosynthetic Response Models for Ten Species Flooded by Hydroelectric Dams. Trees 2013, 27, 193–210. [Google Scholar] [CrossRef]
- Shepherd, M.A.; Harrison, R.; Webb, J. Managing soil Organic Matter—Implications for Soil Structure on Organic Farms. Soil Use Manag. 2002, 18, 284–292. [Google Scholar] [CrossRef]
- Lal, R. Challenges and opportunities in Soil Organic Matter Research. Eur. J. Soil Sci. 2009, 60, 158–169. [Google Scholar] [CrossRef]
- Maathuis, F.J. Physiological Functions of Mineral Macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.J.; Gonçalves, J.F.C.; Ferraz, J.B.S. Crescimento e Eficiência do Uso da Água de Plantas Jovens de castanheira-da-amazônia em Área Degradada e Submetidas à Adubação. Ciênc. Florest. 2012, 22, 393–401. [Google Scholar] [CrossRef]
- Lawrence, D. The Response of Tropical Tree Seedlings to Nutrient Supply: Meta-Analysis for Understanding a Changing Tropical Landscape. J. Trop. Ecol. 2003, 19, 239–250. [Google Scholar] [CrossRef]
- Resende, A.V.; Furtini Neto, A.E.; Curi, N. Mineral Nutrition and Fertilization of Native Tree Species in Brazil: Research Progress and Suggestions for Management. J. Sustain. For. 2005, 20, 45–81. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Lima, R.A.F.; Gandolfi, S.; Nave, A.G. On the Restoration of High Diversity Forests: 30 Years of Experience in the Brazilian Atlantic Forest. Biol. Conserv. 2009, 142, 1242–1251. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Harris, J.A. Restoration Ecology: Repairing the Earth’s Ecosystems in the New Millennium. Restor. Ecol. 2001, 9, 239–246. [Google Scholar] [CrossRef]
- Viana, R.M.; Ferraz, J.B.S.; Neves, A.F.; Vieira, G.; Pereira, B.F.F. Soil Quality Indicators for Different Restoration Stages on Amazon Rainforest. Soil Tillage Res. 2014, 140, 1–7. [Google Scholar] [CrossRef]
- Feldpausch, T.R.; Rondon, M.A.; Fernandes, E.C.M.; Riha, S.J.; Wandelli, E. Carbon and Nutrient Accumulation in Secondary Forests Regenerating on Pastures in Central Amazonia. Ecol. Appl. 2004, 14, S164–S176. [Google Scholar] [CrossRef] [Green Version]
- Lojka, B.; Preininger, D.; van Damme, P.; Rollo, A.; Banout, J. Use of the Amazonian Tree Species Inga Edulis for Soil Regeneration and Weed Control. J. Trop. For. Sci. 2012, 24, 89–101. [Google Scholar]
- Gama, B.; Rocha, R.B.; Salman, A.K.D.; Mendes, Â.M.; Figueiró, M.R. Reforestation Feasibility in Area Formerly Used for Cattle Rasing in the State of Rondônia, Northwest Brazilian Amazon. Rev. Árvore 2013, 37, 1001–1010. [Google Scholar] [CrossRef]
- Monteiro, K.F.G.; Kern, D.C.; Ruivo, M.L.P.; Rodrigues, T.E.; Cometti, J.L.S. Uso de Resíduos de Madeira Como Alternativa de Melhorar as Condições Ambientais em Sistema de Reflorestamento. Acta Amazonica 2010, 40, 409–414. [Google Scholar]
- Marques, T.C.; Carvalho, J.G.; Lacerda, M.P.C.; Mota, P.E.F. Initial Growthing of Paricá (Schizolobium Amazonicum) under Nutrient Omission and Sodium in Nutritive Solution. Cerne 2004, 10, 184–195. [Google Scholar]
- Caione, G.; Lange, A.; Schoninger, E.L. Growth of Seedlings Schizolobium Amazonicum (Huber Ex Ducke) on Substrate Fertilized with Nitrogen, Phosphorus and Potassium. Sci. For. 2012, 40, 213–221. [Google Scholar]
- Chen, Y.; Liu, Z.; Rao, X.; Wang, X.; Liang, C.; Lin, Y.; Zhou, L.; Cai, X.; Fu, S. Carbon Storage and Allocation Pattern in Plant Biomass among Different Forest Plantation Stands in Guangdong, China. Forests 2015, 6, 794–808. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The Role of Dry Mass Allocation in the Growth Response of Plants to Different Levels of Light, CO2, Nutrients and Water: A Quantitative Review. Aust. J. Plant Physiol. 2000, 27, 595–607. [Google Scholar] [CrossRef]
- Cheng, X.; Yu, M.; Wang, G.G.; Wu, T.; Zhang, C. Growth, Morphology and Biomass Allocation in Response to Light Gradient in Five Subtropical Evergreen Broadleaved Tree Seedlings. J. Trop. For. Sci. 2013, 25, 537–546. [Google Scholar]
- Ferreira, M.J.; Gonçalves, J.F.C.; Ferraz, J.B.S. Photosynthetic Parameters of Young Brazil Nut (Bertholletia Excelsa H.B.) Plants Subjected to Fertilization in a Degraded Area in Central Amazonia. Photosynthetica 2009, 47, 616–620. [Google Scholar] [CrossRef]
- Cakmak, I. The Role of Potassium in Alleviating Detrimental Effects of Abiotic Stresses in Plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Silva, C.E.M.; Gonçalves, J.F.C.; Feldpausch, T.R.; Luizão, F.J.; Morais, R.R.; Ribeiro, G.O. Eficiência no Uso dos Nutrientes Por Espécies Pioneiras Crescidas em Pastagens Degradadas na Amazônia Central. Acta Amazonica 2006, 36, 503–512. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Brown, S.; Niles, J.O.; Foley, J.A. Monitoring and Estimating Tropical Forest Carbon Stocks: Making REDD a Reality. Environ. Res. Lett. 2007, 2, 1–13. [Google Scholar] [CrossRef]
- Wortley, L.; Hero, J.; Howes, M. Evaluating Ecological Restoration Success: A Review of the Literature. Restor. Ecol. 2013, 21, 537–543. [Google Scholar] [CrossRef]
- Guariguata, M.; Brancalion, P. Current Challenges and Perspectives for Governing Forest Restoration. Forests 2014, 5, 3022–3030. [Google Scholar] [CrossRef]
- Bullock, J.M.; Aronson, J.; Newton, A.C.; Pywell, R.F.; Rey-Benayas, J.M. Restoration of Ecosystem Services and Biodiversity: Conflicts and Opportunities. Trends Ecol. Evol. 2011, 26, 541–549. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Fertilization | Depth | D.F. Fert. | D.F. Depth |
|---|---|---|---|---|
| C | 23.4 ** | 11.8 ** | 3 | 3 |
| SOM | 23.4 ** | 11.8 ** | 3 | 3 |
| N | 18.8 ** | 18.9 ** | 3 | 3 |
| P | 49.1 ** | 22.2 ** | 3 | 3 |
| K | 68.0 ** | 5.9 ** | 3 | 3 |
| Ca | 36.3 ** | 32.3 ** | 3 | 3 |
| Mg | 79.0 ** | 85.1 ** | 3 | 3 |
| Depth (cm) | Fertilization | Parameter | ||||||
|---|---|---|---|---|---|---|---|---|
| C | SOM | N | P | K | Ca | Mg | ||
| (g·kg−1) | (mg·kg−1) | (cmol·kg−1) | ||||||
| 0–2.5 | T1 | 6.9 ± 1.3 Ba | 11.9 ± 2.2 Ba | 0.4 ± 0.1 Ba | 0.2 ± 0.1 Da | 6.3 ± 1.1 Ba | 0.2 ± 0.1 Ca | 0.2 ± 0.1 Ca |
| 2.5–7.5 | 6.8 ± 1.4 Ba | 11.7 ± 2.4 Ba | 0.4 ± 0.1 Ca | 0.2 ± 0.1 Ca | 6.2 ± 0.9 Ca | 0.1 ± 0.0 Ca | 0.1 ± 0.0 Ca | |
| 7.5–20 | 6.7 ± 1.1 Aa | 11.5 ± 1.8 Aa | 0.3 ± 0.1 Aa | 0.2 ± 0.1 Ca | 5.5 ± 1.6 Ca | 0.1 ± 0.1 Ca | 0.1 ± 0.0 Ba | |
| 20–40 | 6.4 ± 1.0 Ba | 11.1 ± 1.9 Ba | 0.3 ± 0.1 Aa | 0.1 ± 0.1 Ba | 4.9 ± 1.6 Ca | 0.1 ± 0.0 Ba | 0.1 ± 0.1 Ca | |
| 0–2.5 | T2 | 7.5 ± 1.8 Ba | 13.0 ± 3.2 Ba | 0.4 ± 0.1 Ba | 442.0 ± 129.0 Aa | 60.0 ± 15.6 Aa | 2.2 ± 0.8 Aa | 0.7 ± 0.1 Aa |
| 2.5–7.5 | 7.3 ± 1.6 Ba | 12.5 ± 2.8 Ba | 0.4 ± 0.1 BCa | 99.2 ± 55.0 Ab | 63.7 ± 21.6 Aa | 1.1 ± 0.2 Ab | 0.4 ± 0.1 Ab | |
| 7.5–20 | 7.3 ± 1.7 Aa | 12.5 ± 2.9 Aa | 0.4 ± 0.1 Aa | 114.5 ± 102.5 Ab | 62.2 ± 14.6 Aa | 0.7 ± 0.3 Ab | 0.4 ± 0.1 Ab | |
| 20–40 | 7.0 ± 1.2 Ba | 12.0 ± 2.1 Ba | 0.4 ± 0.1 Aa | 69.9 ± 57.4 Ab | 62.2 ± 18.9 Aa | 0.7 ± 0.3 Ab | 0.4 ± 0.1 Ab | |
| 0–2.5 | T3 | 12.4 ± 3.0 Aa | 21.3 ± 5.1 Aa | 0.7 ± 0.1 Aa | 13.0 ± 7.2 Ca | 53.3 ± 14.6 Aa | 1.1 ± 0.1 Ba | 0.5 ± 0.1 Ba |
| 2.5–7.5 | 11.1 ± 2.6 Aa | 19.0 ± 4.5 Aa | 0.5 ± 0.1 Ab | 13.1 ± 13.0 Ba | 32.3 ± 9.0 Bb | 0.7 ± 0.3 Ba | 0.2 ± 0.1 Bb | |
| 7.5–20 | 7.8 ± 1.8 Ab | 13.3 ± 3.0 Ab | 0.4 ± 0.1 Ac | 18.4 ± 11.3 Ba | 23.1 ± 8.6 Bb | 0.9 ± 0.3 Aa | 0.1 ± 0.0 Bb | |
| 20–40 | 7.3 ± 0.9 Bb | 12.6 ± 1.6 Bb | 0.4 ± 0.1 Ac | 7.5 ± 6.2 Aa | 20.4 ± 14.6 BCb | 0.7 ± 0.4 Aa | 0.1 ± 0.0 BCb | |
| 0–2.5 | T4 | 15.0 ± 2.1 Aa | 25.9 ± 3.7 Aa | 0.7 ± 0.1 Aa | 124.6 ± 55.1 Ba | 64.0 ± 17.1 Aa | 2.5 ± 0.8 Aa | 0.9 ± 0.2 Aa |
| 2.5–7.5 | 10.4 ± 2.2 Ab | 17.9 ± 3.9 Ab | 0.5 ± 0.1 ABb | 93.1 ± 86.4 Aa | 45.0 ± 15.3 ABb | 0.9 ± 0.4 ABb | 0.4 ± 0.1 Ab | |
| 7.5–20 | 9.1 ± 2.2 Ab | 15.6 ± 3.8 Ab | 0.4 ± 0.1 Abc | 10.5 ± 7.2 Bb | 32.6 ± 8.8 Bb | 0.6 ± 0.2 Ab | 0.1 ± 0.1 Bc | |
| 20–40 | 8.9 ± 1.4 Ab | 15.2 ± 2.3 Ab | 0.4 ± 0.1 Ac | 10.4 ± 9.8 Ab | 37.9 ± 13.3 Bb | 0.6 ± 0.2 Ab | 0.2 ± 0.0 Bc | |
| Parameter | Fertilization | Species | Sp. X Fert. | Block | D.F. Fert. | D.F. Sp. |
|---|---|---|---|---|---|---|
| RGRheight | 72.7 ** | 127.2 ** | 3.0 * | 2.0 ns | 3 | 2 |
| RGRdiameter | 85.9 ** | 212.9 ** | 6.0 ** | 1.6 ns | 3 | 2 |
| AGRleaves | 88.0 ** | 109.9 ** | 38.3 ** | 0.5 ns | 3 | 2 |
| AGRstems | 52.5 ** | 67.6 ** | 23.3 ** | 0.3 ns | 3 | 2 |
| AGRroots | 113.1 ** | 222.9 ** | 57.2 ** | 0.2 ns | 3 | 2 |
| LMF | 19.0 ** | 14.5 ** | 2.0 ns | 1.2 ns | 3 | 2 |
| SMF | 1.1 ns | 6.6 ** | 2.7 * | 0.4 ns | 3 | 2 |
| RMF | 8.7 ** | 14.7 ** | 4.0 ** | 0.3 ns | 3 | 2 |
| Shoot/Root ratio | 9.6 ** | 15.8 ** | 4.9 ** | 0.7 ns | 3 | 2 |
| N | 21.5 ** | 96.0 ** | 2.7** | 1.8 ns | 3 | 2 |
| P | 0.2 ns | 6.6 ** | 0.7 ns | 0.9 ns | 3 | 2 |
| K | 6.3 ** | 26.5 ** | 3.0 ** | 1.7 ns | 3 | 2 |
| Ca | 5.0 ** | 42.8 ** | 3.2 ** | 2.3 ns | 3 | 2 |
| Mg | 1.1 ns | 41.6 ** | 0.8 ns | 2.4 * | 3 | 2 |
| Fe | 1.2 ns | 47.2 ** | 0.5 ns | 4.2 ** | 3 | 2 |
| Zn | 4.4 ** | 22.7 ** | 2.7 * | 1.7 ns | 3 | 2 |
| Pnmass | 1.9 ns | 31.2 ** | 0.8 ns | 1.2 ns | 3 | 2 |
| PNUE | 3.4 ** | 33.6 ** | 1.0 ns | 0.7 ns | 3 | 2 |
| PPUE | 1.3 ns | 3.5 * | 0.9 ns | 0.7 ns | 3 | 2 |
| PKUE | 2.2 ns | 24.2 ** | 1.3 ns | 0.9 ns | 3 | 2 |
| PCaUE | 1.4 ns | 9.4 ** | 3.2 ** | 0.7 ns | 3 | 2 |
| PMgUE | 0.7 ns | 2.0 ns | 0.2 ns | 2.8 * | 3 | 2 |
| PFeUE | 2.9 * | 18.9 ** | 2.6 * | 1.6 ns | 3 | 2 |
| PZnUE | 0.9 ns | 46.7 ** | 0.9 ns | 2.8 * | 3 | 2 |
| Fertilization | Species | Variables | ||
|---|---|---|---|---|
| AGRleaves | AGRstems | AGRroots | ||
| (g·plant−1·month−1) | ||||
| T1 | D. odorata | 0.59 ± 0.3 Bb | 0.57 ± 0.1 Bb | 0.66 ± 0.3 Bb |
| I. edulis | 3.12 ± 1.8 Ac | 4.95 ± 2.2 Ac | 5.70 ± 2.9 Ac | |
| S. amazonicum | 2.59 ± 1.0 Ac | 6.17 ± 3.3 Ac | 3.97. ± 2.9 Ac | |
| T2 | D. odorata | 17.10 ± 5.1 Ba | 15.15 ± 5.3 Ba | 8.74 ± 1.5 Ba |
| I. edulis | 104.71 ± 20.1 Ab | 116.33 ± 31.1 Ab | 131.78 ± 30.8 Ab | |
| S. amazonicum | 15.82 ± 6.7 Bb | 16.92 ± 8.7 Bb | 8.54 ± 4.9 Bbc | |
| T3 | D. odorata | 6.95 ± 2.5 Bb | 6.47 ± 2.7 Bb | 3.10 ± 1.2 Bb |
| I. edulis | 134.19 ± 61.2 Ab | 139.61 ± 34.1 Ab | 118.42 ± 39.4 Ab | |
| S. amazonicum | 14.76 ± 6.8 Bb | 24.12 ± 15.5 Bb | 19.12 ± 10.9 Bb | |
| T4 | D. odorata | 16.97 ± 4.8 Ba | 15.37 ± 4.4 Ba | 7.63 ± 3.3 Ca |
| I. edulis | 224.98 ± 54.1 Aa | 252.10 ± 75.6 Aa | 190.58 ± 25.1 Aa | |
| S. amazonicum | 54.32 ± 12.9 Ba | 65.88 ± 24.9 Ba | 50.27 ± 16.2 Ba | |
| Species | Fertilization | Parameter | |||
|---|---|---|---|---|---|
| LMF | SMF | RMF | Shoot/Root Ratio | ||
| T1 | D. odorata | 0.33 ± 0.06 Ab | 0.34 ± 0.04 Aa | 0.34 ± 0.07 ABa | 2.1 ± 0.8 ABb |
| I. edulis | 0.23 ± 0.06 Bb | 0.37 ± 0.04 Aa | 0.40 ± 0.09 Aa | 1.6 ± 0.6 Ab | |
| S. amazonicum | 0.24 ± 0.06 Bc | 0.48 ± 0.05 Ba | 0.28 ± 0.06 Ba | 2.7 ± 0.8 Bb | |
| T2 | D. odorata | 0.42 ± 0.04 Aa | 0.36 ± 0.03 Aa | 0.22 ± 0.03 Bb | 3.6 ± 0.6 Aa |
| I. edulis | 0.30 ± 0.04 BAb | 0.33 ± 0.08 Aa | 0.37 ± 0.07 Aab | 1.8 ± 0.6 Bab | |
| S. amazonicum | 0.40 ± 0.07 Aa | 0.40 ± 0.06 Ab | 0.20 ± 0.03 Bb | 4.3 ±0.8 Aa | |
| T3 | D. odorata | 0.42 ± 0.04 Aa | 0.39 ± 0.07 ABa | 0.18 ± 0.03 Bb | 4.4 ± 1.4 Aa |
| I. edulis | 0.34 ± 0.04 Ba | 0.36 ± 0.03 Ba | 0.30 ± 0.06 Ab | 2.3 ± 0.2 ABb | |
| S. amazonicum | 0.25 ± 0.05 Cc | 0.42 ± 0.05 Aab | 0.33 ± 0.04 Aa | 2.6 ± 1.1 Bb | |
| T4 | D. odorata | 0.42 ± 0.03 Aa | 0.38 ± 0.06 Aa | 0.19 ± 0.04 Bb | 4.5 ± 1.0 Aa |
| I. edulis | 0.33 ± 0.04 Ba | 0.37 ± 0.04 Aa | 0.30 ± 0.07 Ab | 2.5 ± 0.6 Ba | |
| S. amazonicum | 0.33 ± 0.06 Bb | 0.38 ± 0.06 Ab | 0.29 ± 0.05 Aa | 2.5 ± 0.5 Bb | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaquetti, R.K.; Gonçalves, J.F.d.C.; Ferraz, J.B.S.; Ferreira, M.J.; Santos Junior, U.M.d. Ecofunctional Traits and Biomass Production in Leguminous Tree Species under Fertilization Treatments during Forest Restoration in Amazonia. Forests 2016, 7, 76. https://doi.org/10.3390/f7040076
Jaquetti RK, Gonçalves JFdC, Ferraz JBS, Ferreira MJ, Santos Junior UMd. Ecofunctional Traits and Biomass Production in Leguminous Tree Species under Fertilization Treatments during Forest Restoration in Amazonia. Forests. 2016; 7(4):76. https://doi.org/10.3390/f7040076
Chicago/Turabian StyleJaquetti, Roberto K., José Francisco de C. Gonçalves, João Baptista S. Ferraz, Marciel J. Ferreira, and Ulysses M. dos Santos Junior. 2016. "Ecofunctional Traits and Biomass Production in Leguminous Tree Species under Fertilization Treatments during Forest Restoration in Amazonia" Forests 7, no. 4: 76. https://doi.org/10.3390/f7040076