
Genetic Diversity of the Black Mangrove Avicennia germinans (L.) Stearn in Northwestern Mexico

Abstract
:
1. Introduction
2. Materials and Methods
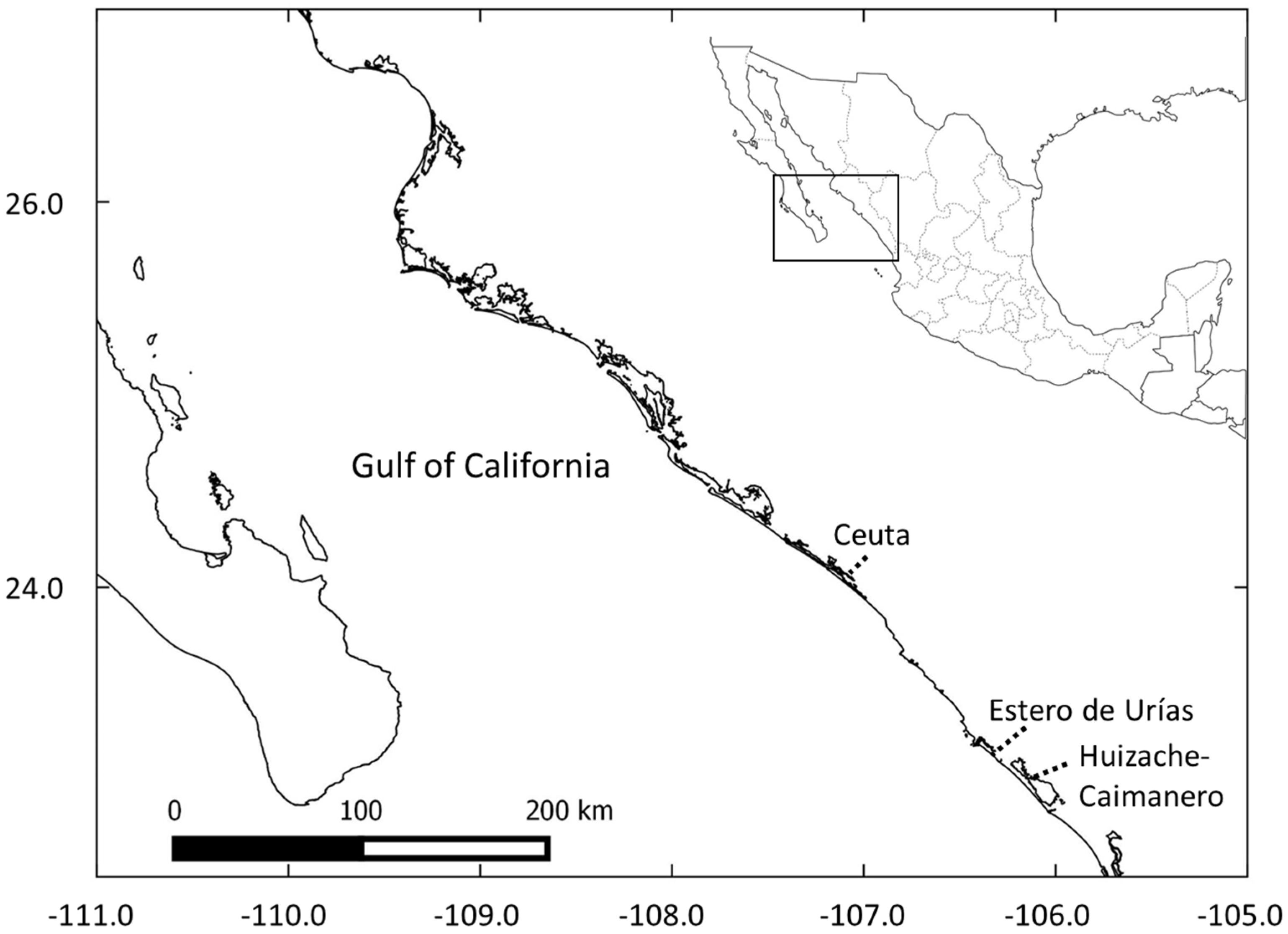
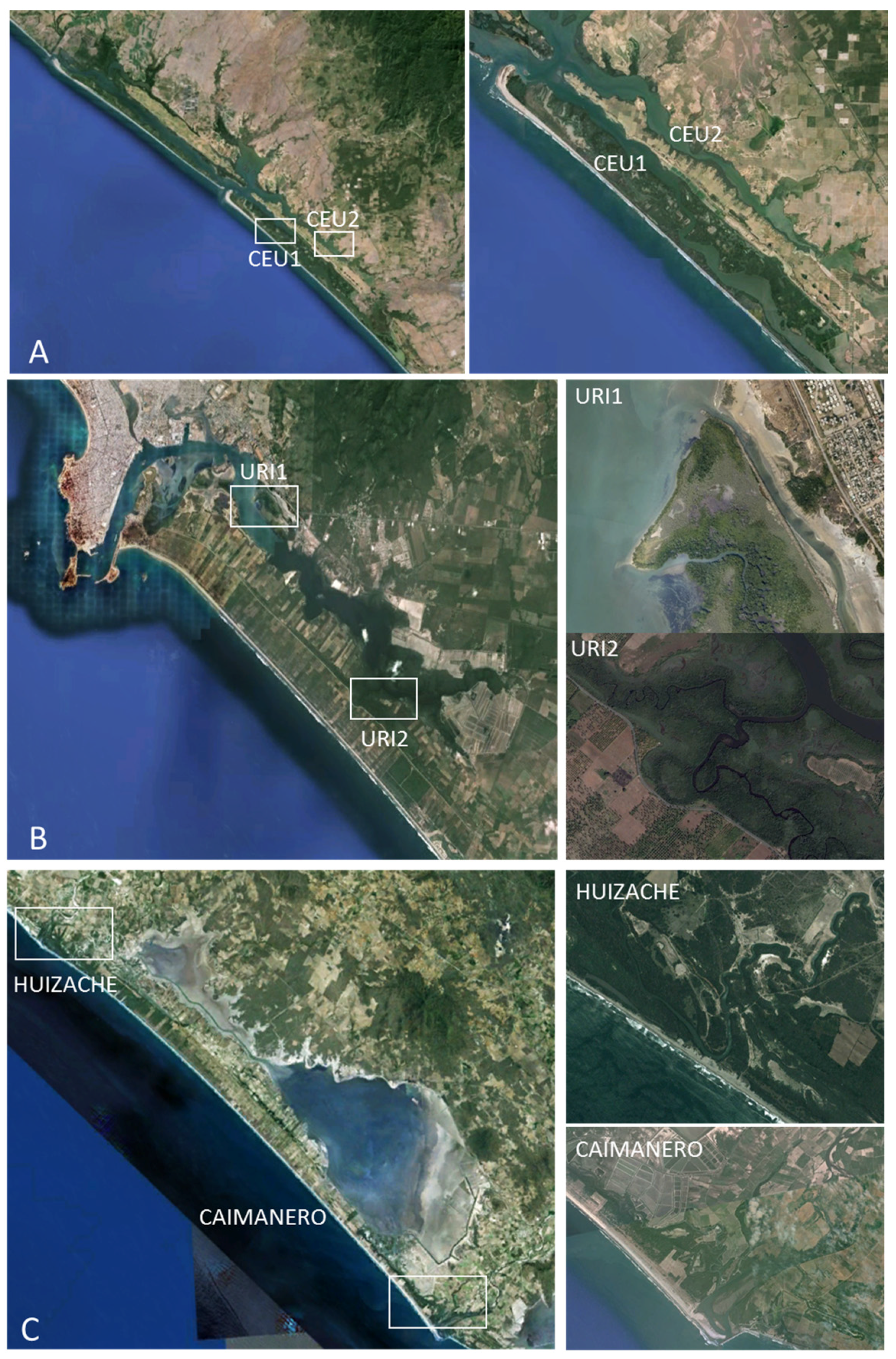
2.1. Study Site Selection
2.2. Sampling Strategy and DNA Extraction
2.3. Microsatellite Analysis
2.4. Statistical Analysis
3. Results
3.1. Genetic Diversity
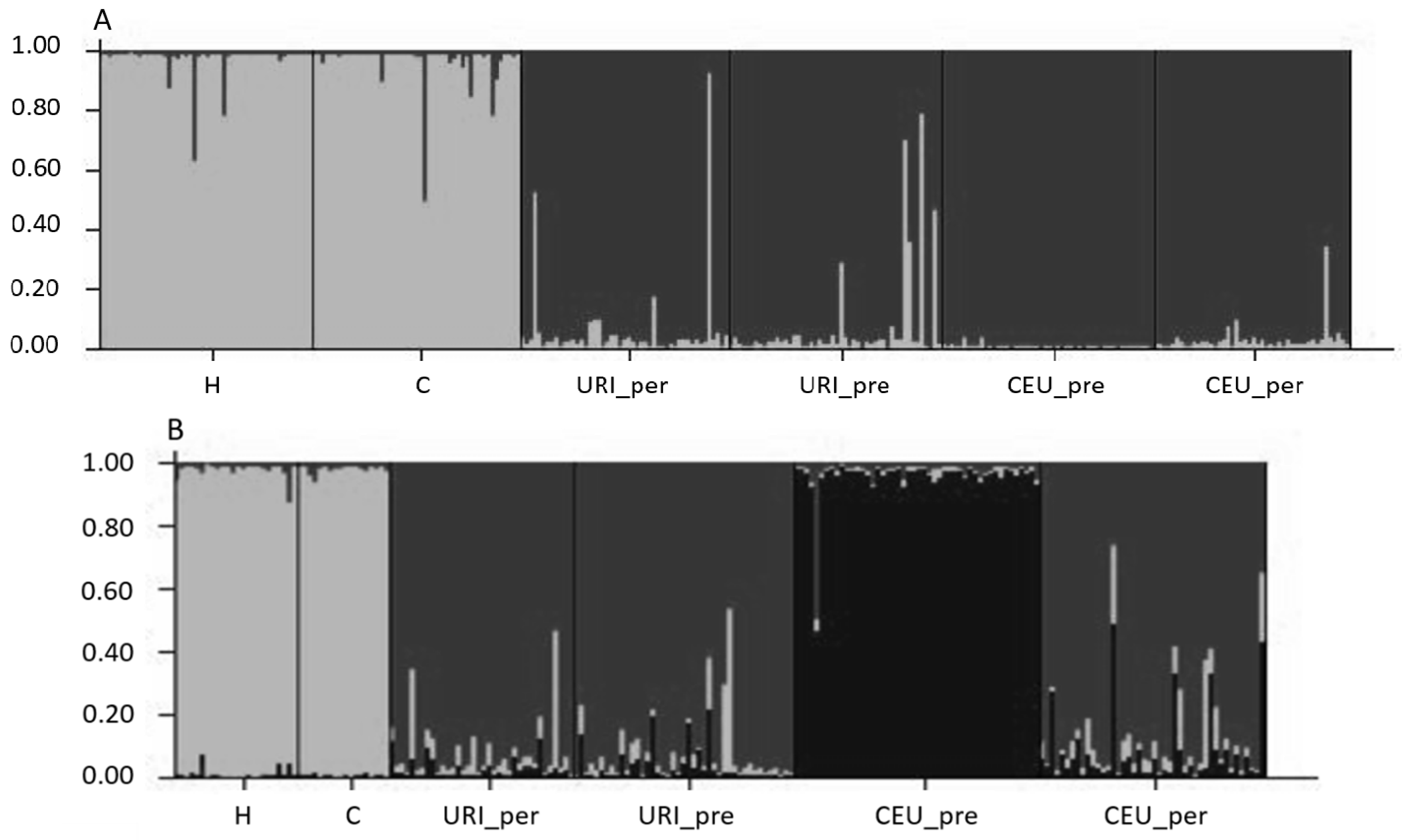
3.2. Population Genetic Structure
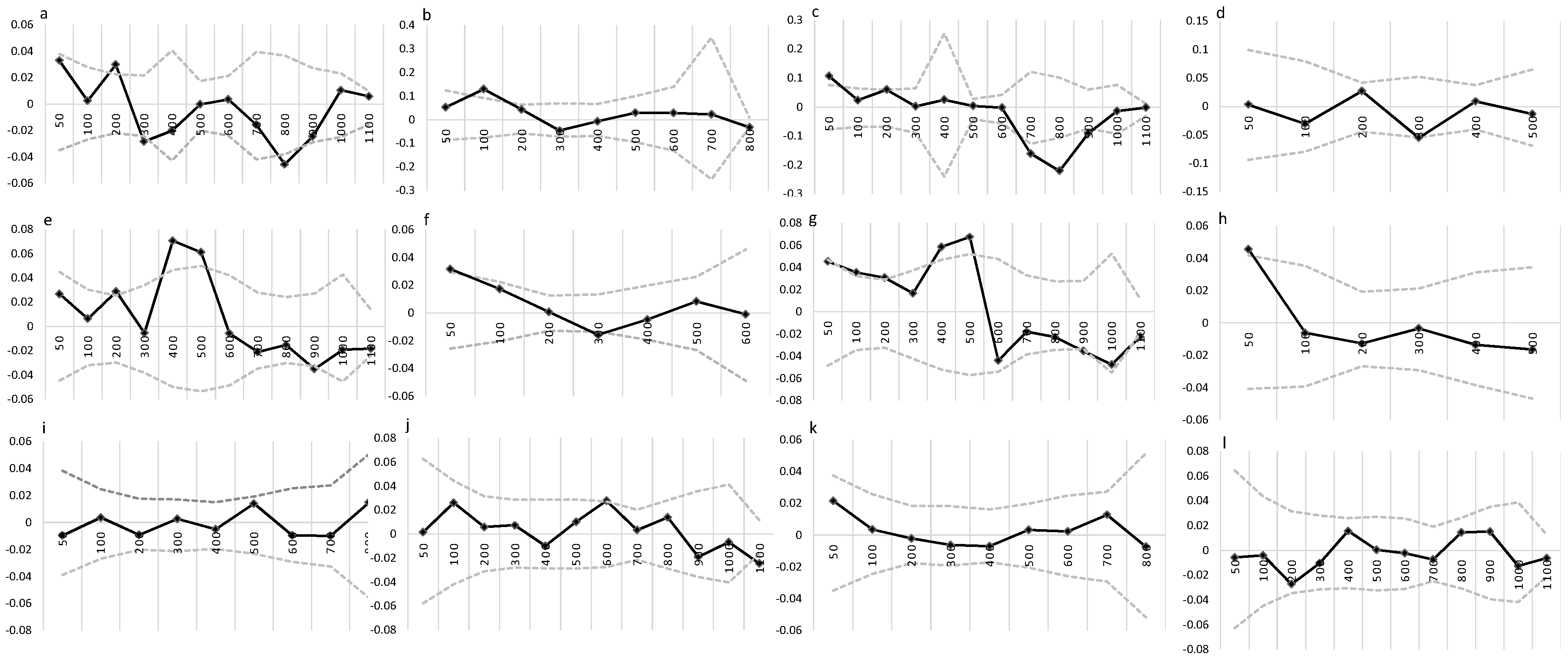
3.3. Spatial Genetic Structure
4. Discussion
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′–3′ | Repeat Motif | Annealing Temperature (°C) | Allele Range (bp) |
|---|---|---|---|---|
| 1 Agerm15 | F: ACT TAC ACA CAA AAT GCA CA | (CA) 4 (AC) 13 | 56.7 | 248 |
| R: CTG AGA GTG CCG ACT GAA TG | ||||
| 1 Agerm16 | F: CCT AAT ACA AAT GAC ACT AAA A | (TG) 9 | 53.4 | 176 |
| R: TGC ATG TCA ATT ATC AGT CT | ||||
| 1 Agerm11 | F: GCC CTC AGC ATG AAA CTT GT | (TG) 10 | 53.4 | 138 |
| R: AAA TAA TAA GGC GCC CGT GT | ||||
| 1 Agerm07 | F: CCT GAC ACT CTG GGA CAT CA | (GT) 9 | 50.5 | 157 |
| R: CCT TTT GAC GCA TTT GTG G | ||||
| 1 Agerm03 | F: CCA TGT TTT TGA CTT TTT ATT TTG | (CA) 9 | 48.2 | 161 |
| R: TTA CGA TAG GGT GGA TTG AGA TTT T | ||||
| 1 Agerm06 | F: GAA TTG GCT GGA ATG AGG AA | (GT) 6 | 63.4 | 175 |
| R: GTG TTT TGG AAG GAG CCT GT | ||||
| 1 Agerm01 | F: CAG TTT GGT GAG AAG GAT GTT | (AC) 15 | 53.4 | 127 |
| R: TTT GAG GTC GGC TCG TTA AG | ||||
| 2 Di6t | F: TAT ATG GAA ACC AAC CAT GC | (ATT) 4 N 7 (GT) 15 | 50 | 207–260 |
| R: GAT GGT GCG GGC TGT C | ||||
| 2 Te4T | F: CAA GAT TTT GAT CAA TTA TG | (CATA) 5 CATG (CATA) 9 | 52 | 79–107 |
| R: ACG TGA CGC ATT GAT CTA CC | ||||
| 2 Di13 | F: TTC ACC TAT ATG AAT GGC GCA C | (CA) 10 | 58 | 297–303 |
| R: GGA GCA CGC GAT AAC AAG AGC | ||||
| 2 Te8 | F: ACA CAA CGC AGA TAA ATC C | (TGTA) 6 | 52 | 104–112 |
| R: AAT GAT CGC CTG TCT CCG TC | ||||
| 1 Agerm20 | F: TAT AAC AAT GCC CTG ACA CTC T | (GT) 9 | 59.6 | 203 |
| R: ACA AAC ATG CCA ACA CAA ATA | ||||
| 1 Agerm14 | F: CCA ATT GTG TCG TCC TTT TA | (CA) 8 (AT) 6 | 59.6 | 159 |
| R: AGC CTT ACT TTT CCT TTG T | ||||
| 3 CTT1 | F: CAT CCA CAT TGC CCT GAT | (CTT) 8 | 55 | 114–141 |
| R: GCC TGA TAA GTT GAG TTG CTG | ||||
| 3 CA1 | F: AAG AGA ATG AGG AGT GGT AGG C | (CA) 13 | 56 | 335–391 |
| R: CAA GCC TCG CAT TAA AGT GG | ||||
| 3 CA2 | F: AGC CCT GGT GAT AGT GAC AG | (CA) 12 | 55 | 310–346 |
| R: ACC AGC AAA ATG TAA AGT GGC ATC | ||||
| 3 CT3 | F: ATT CCG AGC AAG AGC CTA C | (CT) 14 | 55 | 412–516 |
| R: ACT GAG GAT GAG TTG GTT TAT CTC | ||||
| 2 Te7 | F: CTA AGT AGG ACA GTA ATG CGA C | (CAT) 2 (AT) 3 (GTAT) 5 | 50 | 170–198 |
| R: AAT CAT CAG AAT CCC TCA AGT GC | ||||
| 2 Te9 | F: GAA CCT GAT TTA ATG GTT GC | (CA) 8 (GA) 2 (CAGA) 5 | 50 | 218–238 |
| R: TCA ATT TGC TTG TAG AGG |
References
- Bradshaw, R.H.W. Past anthropogenic influence on European forests and some possible genetic consequences. For. Ecol. Manag. 2004, 197, 203–212. [Google Scholar] [CrossRef]
- Azevedo, V.C.R.; Kanashiro, M.; Ciampi, A.Y.; Grattapaglia, D. Genetic structure and mating system of Manilkara huberi (Ducke) A. Chev., a Heavily Logged Amazonian Timber Species. J. Hered. 2007, 98, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Ratnam, W.; Rajora, O.P.; Finkeldey, R.; Aravanopoulos, F.; Bouvet, J.M.; Vaillancourt, R.E.; Kanashiro, M.; Fady, B.; Tomita, M.; Vinson, C. Genetic effects of forests management practices: Global synthesis and perspectives. For. Ecol. Manag. 2014, 332, 52–65. [Google Scholar] [CrossRef]
- Golladay, S.W.; Martin, K.L.; Vose, J.M.; Wear, D.N.; Covich, A.P.; Hobbs, R.J.; Klepzig, K.D.; Likens, G.E.; Naiman, R.J.; Shearer, A.W. Achievable future conditions as a framework for guiding forest conservation and management. For. Ecol. Manag. 2016, 360, 80–96. [Google Scholar] [CrossRef]
- Alongi, D.M. Present state and future of the world’s mangrove forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Ruiz-Luna, A.; Berlanga-Robles, C.A. Modifications in Coverage Patterns and Land Use around the Huizache-Caimanero Lagoon System, Sinaloa, Mexico: A Multi-temporal Analysis using LANDSAT images. Estuar. Coast. Shelf Sci. 1999, 49, 37–44. [Google Scholar] [CrossRef]
- Sjölund, M.J.; Jump, A.S. Coppice management of forest impacts spatial genetic structure but not genetic diversity in European beech (Fagus sylvatica L.). For. Ecol. Manag. 2015, 336, 65–71. [Google Scholar] [CrossRef]
- Piotti, A.; Leonardi, S.; Heuertz, M.; Buiteveld, J.; Geburek, T.; Gerber, S.; Kramer, K.; Vettori, C.; Vendramin, G.G. Within-Population Genetic Structure in Beech (Fagus sylvatica L.) Stands Characterized by Different Disturbance Histories: Does Forest Management Simplify Population Substructure? PLoS ONE 2013, 8, e73391. [Google Scholar] [CrossRef] [PubMed]
- Loveless, M.D.; Hamrick, J.L. Ecological Determinants of Genetic Structure in Plants Populations. Annu. Rev. Ecol. Syst. 1984, 15, 65–95. [Google Scholar] [CrossRef]
- Jump, A.S.; Marchant, R.; Peñuelas, J. Environmental change and the option value of genetic diversity. Trends Plant Sci. 2009, 14, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Jennerjahn, T.C.; Itekkot, V. Revelance of mangroves for the production and deposition of organic matter along tropical continental margins. Sci. Nat. 2002, 89, 23–30. [Google Scholar] [CrossRef]
- Day, J.W., Jr.; Hall, C.A.S.; Kemp, W.M.; Yanez-Arancibia, A. Estuarine Ecology; John Wiley & Sons, Inc.: New York, NY, USA, 1989; p. 543. [Google Scholar]
- Aburto-Oropeza, O.; Ezcurra, E.; Danemann, G.; Valdez, V.; Murray, J.; Sala, E. Mangroves in the Gulf of California increase fishery yields. Environ. Sci. 2008, 105, 10456–10459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valiela, I.; Bowen, J.L.; York, J.K. Mangrove forests: One of the world’s threatened major tropical environments. Bioscience 2001, 51, 807–815. [Google Scholar] [CrossRef]
- Gunderson, L.H.; Holling, C.S. Panarchy: Understanding Transformations in Human and Natural Systems; Island Press: Washington, DC, USA, 2002. [Google Scholar]
- Duque-Estrada, G.C.; Gomes-Soares, M.L.; Oliveira-Chaves, F.D.; Fernandez-Cavalcanti, V. Analysis of the structural variability of mangrove forests through the physiographic types approach. Aquat. Bot. 2013, 111, 135–143. [Google Scholar] [CrossRef]
- Tomlinson, B.P. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Millán-Aguilar, O.; Nettel-Hernanz, A.; Hurtado-Oliva, M.; Flores-Cárdenas, F.; Díaz-Lara, L.; Manzano-Sarabia, M. Landscape metrics and conservation status of five mangrove wetlands in the Eastern Gulf of California margin. Sci. Total Environ. submitted for publication. 2016. [Google Scholar]
- Ramsar. Available online: http://www.ramsar.org/wetland/mexico (accessed on 1 September 2016).
- Cullings, K.W. Design and testing of a plant-specific PCR primer for ecological and evolutionary studies. Mol. Ecol. 1992, 1, 233–240. [Google Scholar] [CrossRef]
- Nettel, A.; Rafii, F.; Dodd, S. Characterizacion of microsatellite markers for the mangrove tree Avicennia germinans L. Avicenniaceae. Mol. Ecol. Resour. 2005, 5, 103–105. [Google Scholar] [CrossRef]
- Ceron-Souza, I.; Rivera-Ocasio, E.; Funk, M.S.; McMillan, O. Development of six microsatellite loci for black mangrove (Avicennia germinans). Mol. Ecol. 2006, 6, 692–694. [Google Scholar] [CrossRef]
- Mori, G.M.; Zucchi, M.I.; Sampaio, I.; Souza, A.P. Microsatellites for the mangrove tree Avicennia germinans. Acanthaceae: Tools for hybridization and mating system studies. Am. J. Bot. 2010, 97, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Van Oosterhout, C.V.; Hutchison, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Resour. 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer-Heidi, E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices; Institute of Ecology Biology Building, UNIL: Lausanne, Switzerland, 2002. [Google Scholar]
- GENEPOP software. Available online: http://genepop.curtin.edu.au (accessed on 1 September 2016).
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes—Application to human mitochondrial—DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Pritchard, D.W.; Stephens, M.; Donnelly, P.J. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Dent, E.A.; VonHoldt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Hardy, O.J.; Vekemans, X. SPAGEDI: A versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol. Ecol. 2002, 2, 618–620. [Google Scholar] [CrossRef]
- Loiselle, B.A.; Sork, V.L.; Nason, J.; Graham, C. Spatial genetic-structure of a tropical understory shrub, Sychotria officinalis (Rubiaceae). Am. J. Bot. 1995, 82, 1420–1425. [Google Scholar] [CrossRef]
- Vekemans, X.; Hardy, O.J. New insights from fine-scale spatial genetic structure analyses in plant populations. Mol. Ecol. 2004, 13, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Nettel, A.; Dodd, R.S. Drifting propagules and receding swamps: Genetic footprints of mangrove recolonization and dispersal along tropical coast. Journal compilation. Int. J. Org. Evol. 2007, 61, 958–971. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Castro, E.; Dodd, R.S.; Riosmena-Rodríguez, R.; Enriquez-Paredes, L.M.; Tovilla-Hernández, C.; López-Vivas, J.M.; Aguilar-May, B.; Muñiz-Salazar, R. Post-Glacial expansión and population genetic divergence of mangrove species Avicennia germinans and Rhizophora mangle L. along the Mexican coast. PLoS ONE 2014, 9, e93358. [Google Scholar] [CrossRef] [PubMed]
- Cerón-Souza, I.; Bermingham, E.; McMillan, O.W.; Jones, F.A. Comparative genetic structure of two mangrove species in Caribbean and Pacific estuaries of Panama. BMC Evol. Biol. 2010, 12, 205. [Google Scholar] [CrossRef] [PubMed]
- Cresswell, J.E. Spatial heterogeneity, pollinator behavior and pollinator-mediated gene flow: Bumblebee movements in variously aggregated rows of oil-seed rape. Oikos 1997, 78, 546–556. [Google Scholar] [CrossRef]
- Goverde, M.; Schweizer, K.; Baur, B.; Erhardt, A. Small-scale habitat fragmentation effects on pollinator behavior: Experimental evidence from the bumblebee Bombus veteranus on calcareous grasslands. Biol. Conserv. 2002, 104, 293–299. [Google Scholar] [CrossRef]
- Vranckx, G.; Jacquemyn, H.; Muys, B.; Honnay, O. Meta-Analysis of susceptibility of woody plants to loss of genetic diversity through habitat fragmentation. Conserv. Biol. 2011, 26, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Salas-Leiva, D.E.; Mayor-Durán, V.M.; Toro-Perea, N. Genetic diversity of the black mangrove. Avicennia germinans L. in Colombia. Aquat. Bot. 2009, 91, 187–193. [Google Scholar] [CrossRef]
- Flores-Cárdenas, F.; Hurtado-Oliva, M.A.; Doyle, T.W.; Nieves-Soto, M.; Díaz-Castro, S.; Manzano-Sarabia, M. Litterfall production of mangroves in Huizache-Caimanero lagoon system, México. J. Coast. Res. 2016. [Google Scholar] [CrossRef]
- Ramsar. Ramsar Sites Information System. Available online: https://rsis.ramsar.org/ris/1689 (accessed on 31 August 2016).




| Age Groups | Site | N | He | Ho | Allelic Richness | FIS | HW.eq |
|---|---|---|---|---|---|---|---|
| PRESERVED | |||||||
| Adults | CEU | 50 | 0.226 | 0.192 | 1.89 | 0.155 | 0.000 |
| URI | 50 | 0.221 | 0.182 | 1.84 | 0.175 | 0.000 | |
| HUIZACHE | 50 | 0.165 | 0.151 | 1.55 | 0.090 | 0.017 | |
| Total/Mean | 150 | 0.204 | 0.175 | 1.76 | 0.140 | ||
| Saplings | CEU | 48 | 0.234 | 0.232 | 1.78 | 0.007 | 0.195 |
| URI | 43 | 0.237 | 0.210 | 1.90 | 0.115 | 0.000 | |
| HUIZACHE | 24 | 0.194 | 0.156 | 1.75 | 0.200 | 0.001 | |
| Total/Mean | 115 | 0.222 | 0.199 | 1.81 | 0.107 | ||
| PERTURBED | |||||||
| Adults | CEU | 46 | 0.202 | 0.198 | 1.71 | 0.020 | 0.035 |
| URI | 49 | 0.193 | 0.199 | 1.73 | −0.061 | 0.179 | |
| CAIMANERO | 49 | 0.188 | 0.185 | 1.69 | 0.021 | 0.486 | |
| Total/Mean | 144 | 0.195 | 0.194 | 1.71 | −0.007 | ||
| Saplings | CEU | 44 | 0.184 | 0.158 | 1.86 | 0.138 | 0.000 |
| URI | 36 | 0.219 | 0.189 | 1.77 | 0.140 | 0.001 | |
| CAIMANERO | 18 | 0.244 | 0.220 | 1.66 | 0.104 | 0.098 | |
| Total/Mean | 98 | 0.220 | 0.190 | 1.76 | 0.13 | ||
| Source of Variation | d.f. | Variance Component | % Total Variance | p |
|---|---|---|---|---|
| Coastal lagoon system | ||||
| Among regions | 2 | 0.102 | 7.33 | 0.013 |
| Within regions | 9 | 0.134 | 9.57 | <0.01 |
| Within populations | 1002 | 1.160 | 83.1 | <0.01 |
| Status: Preserved vs. Perturbed (saplings and adults) | ||||
| Among regions | 1 | 0.046 | 3.31 | 0.169 |
| Within regions | 10 | 0.183 | 13.16 | <0.01 |
| Within populations | 1002 | 1.160 | 83.54 | <0.01 |
| Age: Saplings vs. Adults (preserved and perturbed) | ||||
| Among regions | 1 | −0.036 | −2.68 | 0.804 |
| Within regions | 10 | 0.227 | 16.82 | <0.01 |
| Within populations | 1002 | 1.160 | 85.86 | <0.01 |
| Status and age | ||||
| Among regions | 3 | −0.024 | −1.73 | 0.701 |
| Within regions | 8 | 0.227 | 16.65 | <0.01 |
| Within populations | 1002 | 1.160 | 85.08 | <0.01 |
| Preserved and perturbed saplings | ||||
| Among regions | 1 | −0.071 | −4.71 | 0.801 |
| Within regions | 4 | 0.484 | 32.08 | 0.00 |
| Within populations | 420 | 1.095 | 72.62 | 0.00 |
| Preserved and perturbed adults | ||||
| Among regions | 1 | −0.013 | −0.99 | 0.900 |
| Within regions | 4 | 0.212 | 15.81 | <0.01 |
| Within populations | 582 | 1.143 | 85.18 | <0.01 |
| Sites | Age Group | Condition | F1 | b-log | Sp | SE |
|---|---|---|---|---|---|---|
| HC | Adults | Preserved | 0.033 * | −0.005 | 0.005 | 0.003 |
| Perturbed | 0.026 | −0.005 * | 0.005 | 0.003 | ||
| Saplings | Preserved | 0.106 ** | −0.031 ** | 0.035 | 0.008 | |
| Perturbed | 0.003 | 0.0002 | −0.0002 | 0.01 | ||
| URI | Adults | Preserved | 0.026 | −0.015 ** | 0.015 | 0.004 |
| Perturbed | 0.031 * | −0.013 * | 0.013 | 0.005 | ||
| Saplings | Preserved | 0.045 * | −0.023 *** | 0.024 | 0.005 | |
| Perturbed | 0.046 * | 0.006 | −0.006 | 0.009 | ||
| CEU | Adults | Preserved | 0.009 | 0.002 | −0.002 | 0.005 |
| Perturbed | 0.002 | −0.011 * | 0.011 | 0.005 | ||
| Saplings | Preserved | 0.021 | −0.002 | 0.002 | 0.005 | |
| Perturbed | −0.005 | 0.003 | −0.003 | 0.005 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Millán-Aguilar, O.; Manzano-Sarabia, M.; Nettel-Hernanz, A.; Dodd, R.S.; Hurtado-Oliva, M.Á.; Velázquez-Velázquez, E. Genetic Diversity of the Black Mangrove Avicennia germinans (L.) Stearn in Northwestern Mexico. Forests 2016, 7, 197. https://doi.org/10.3390/f7090197
Millán-Aguilar O, Manzano-Sarabia M, Nettel-Hernanz A, Dodd RS, Hurtado-Oliva MÁ, Velázquez-Velázquez E. Genetic Diversity of the Black Mangrove Avicennia germinans (L.) Stearn in Northwestern Mexico. Forests. 2016; 7(9):197. https://doi.org/10.3390/f7090197
Chicago/Turabian StyleMillán-Aguilar, Olivia, Marlenne Manzano-Sarabia, Alejandro Nettel-Hernanz, Richard S. Dodd, Miguel Ángel Hurtado-Oliva, and Ernesto Velázquez-Velázquez. 2016. "Genetic Diversity of the Black Mangrove Avicennia germinans (L.) Stearn in Northwestern Mexico" Forests 7, no. 9: 197. https://doi.org/10.3390/f7090197