
Clearcutting and Site Preparation, but Not Planting, Promoted Early Tree Regeneration in Boreal Alaska

Abstract
:1. Introduction
2. Materials and Methods
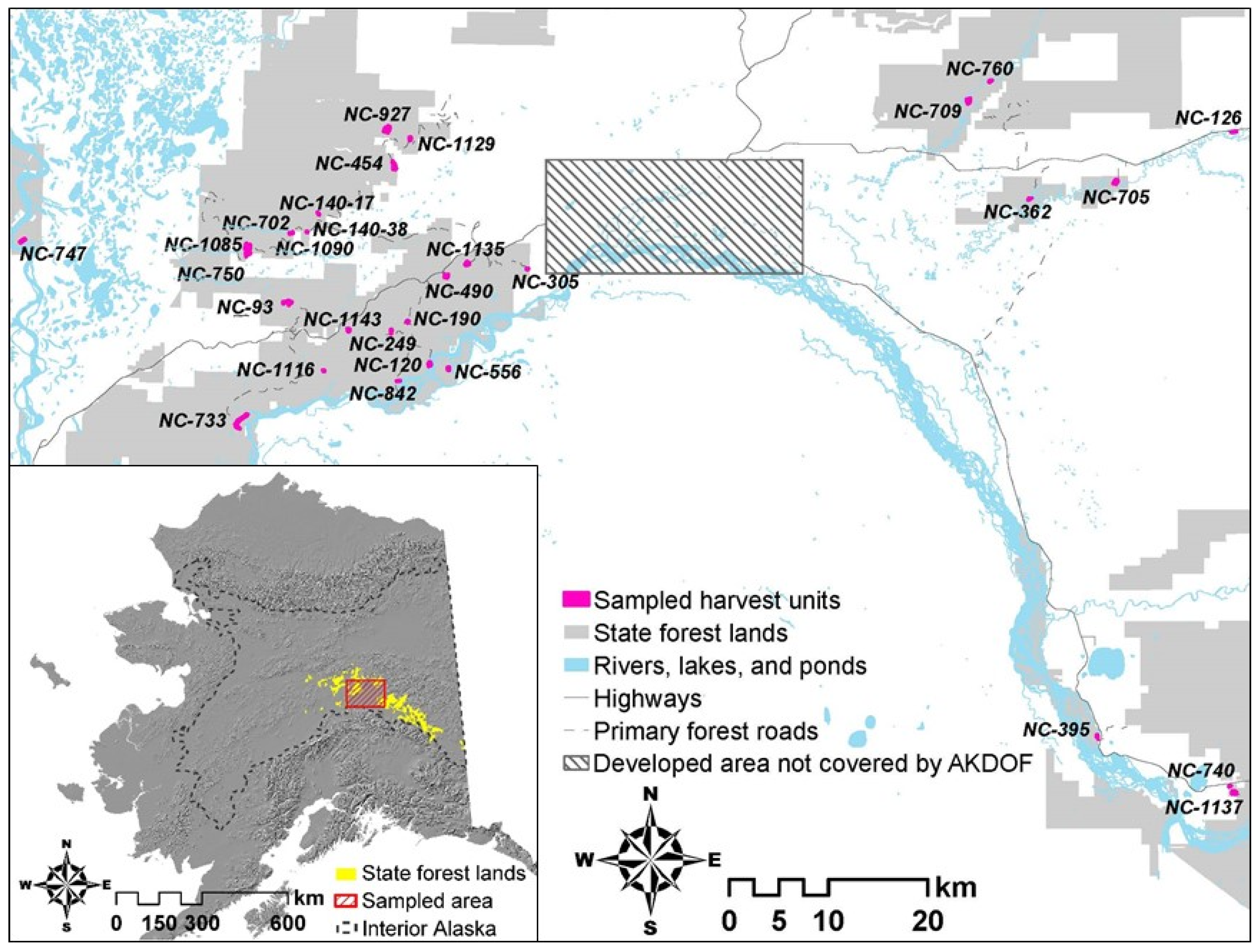
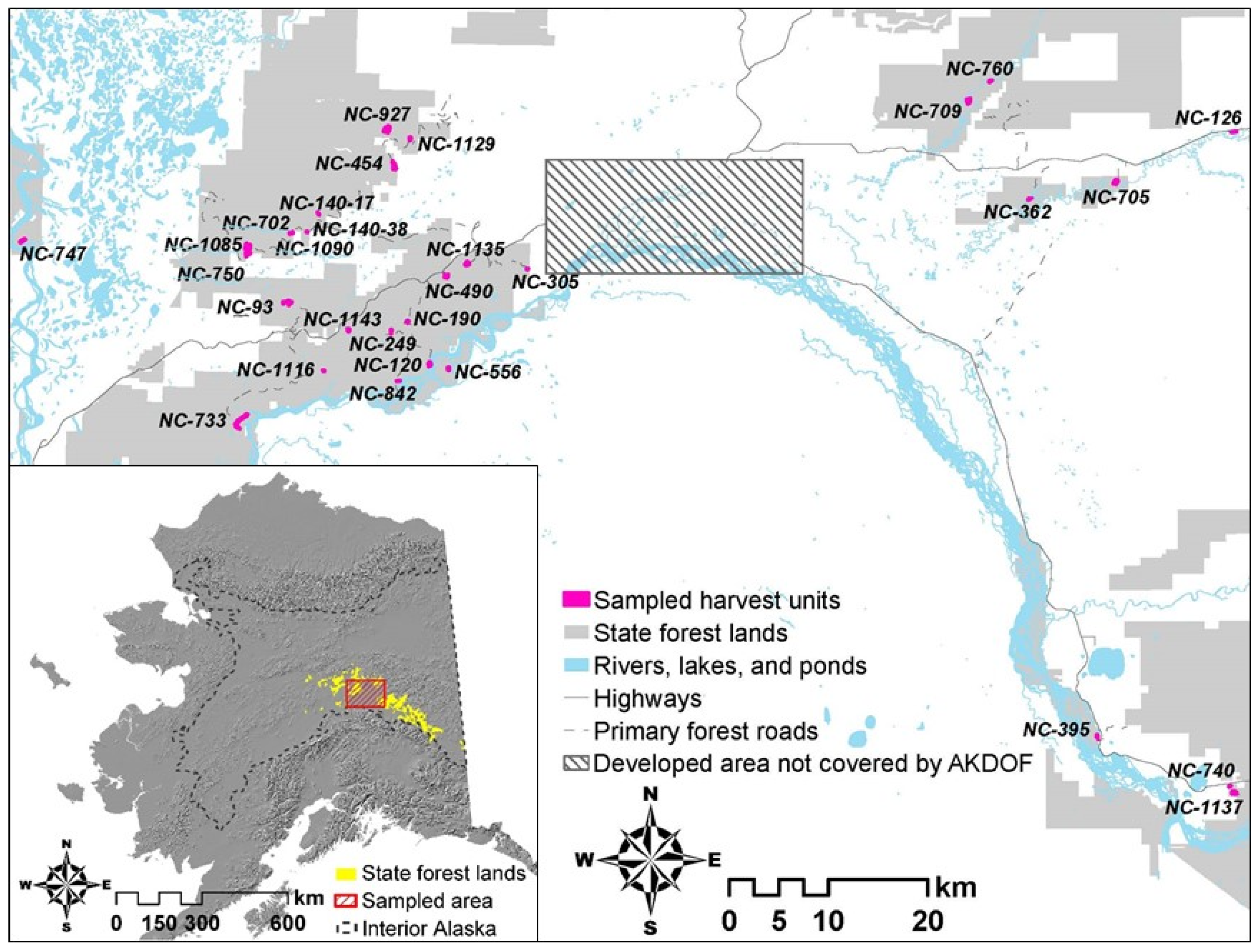
2.1. Study Area
2.2. Silvicultural Systems
2.3. Sampling Design
2.4. Data Collection and Preparation
2.4.1. Response Variables
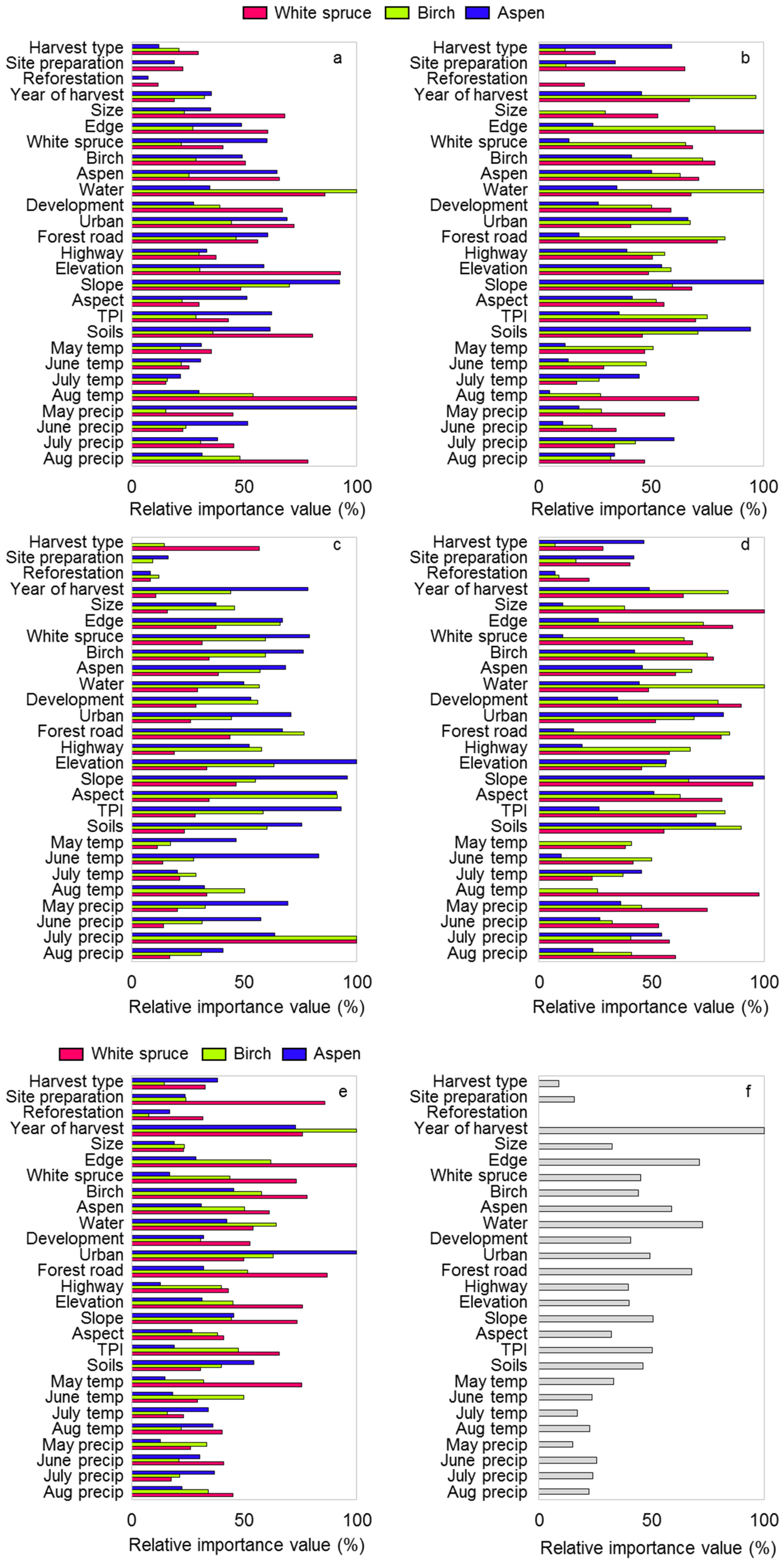
2.4.2. Predictors
2.5. Statistical Analysis
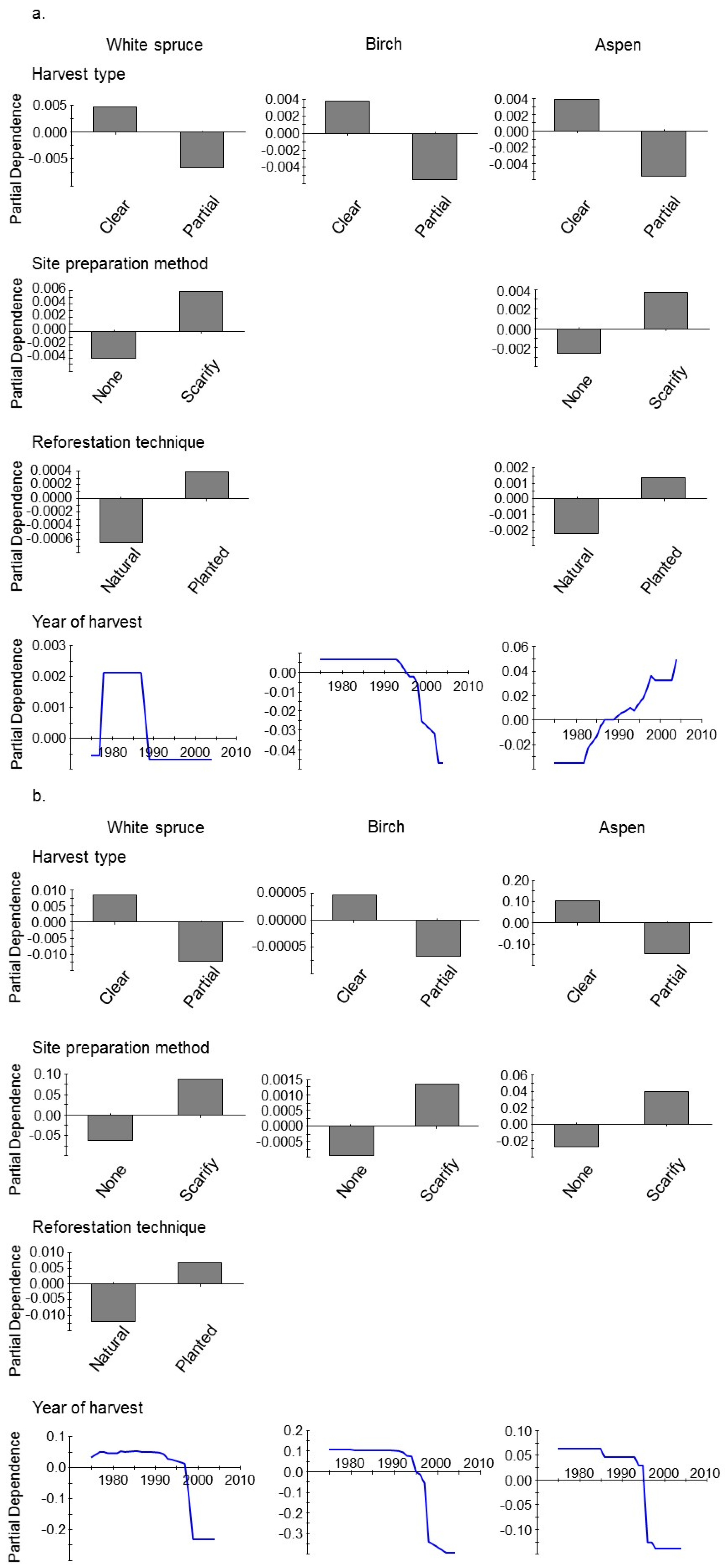
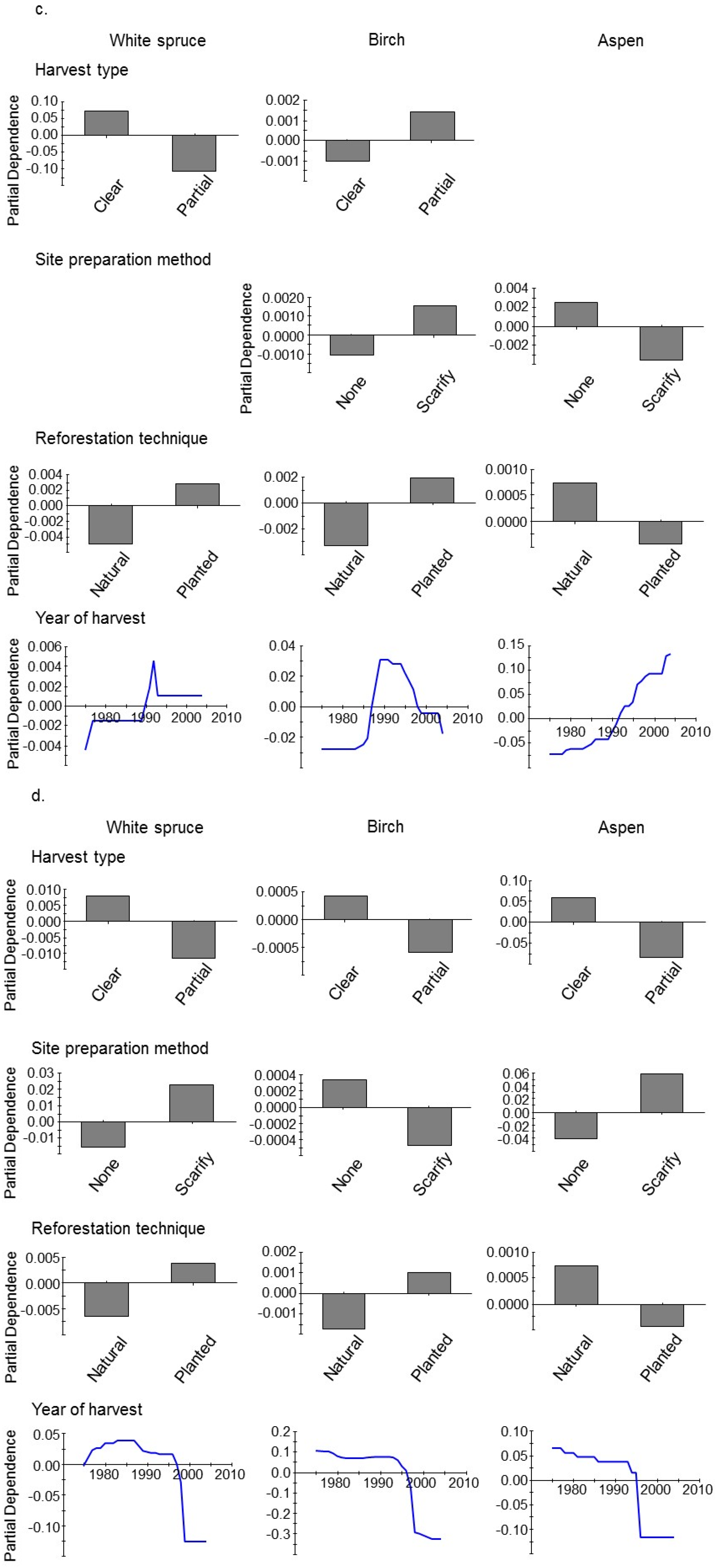
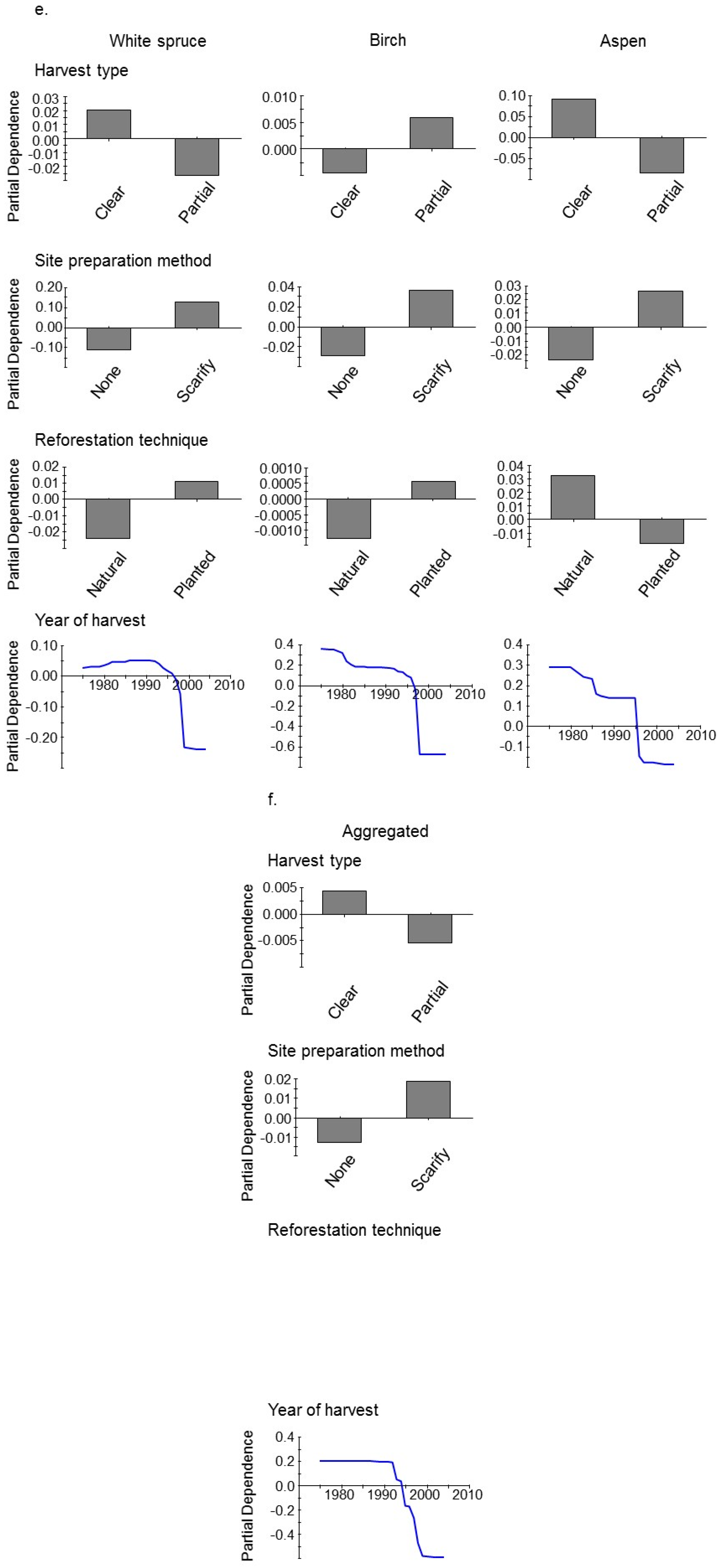
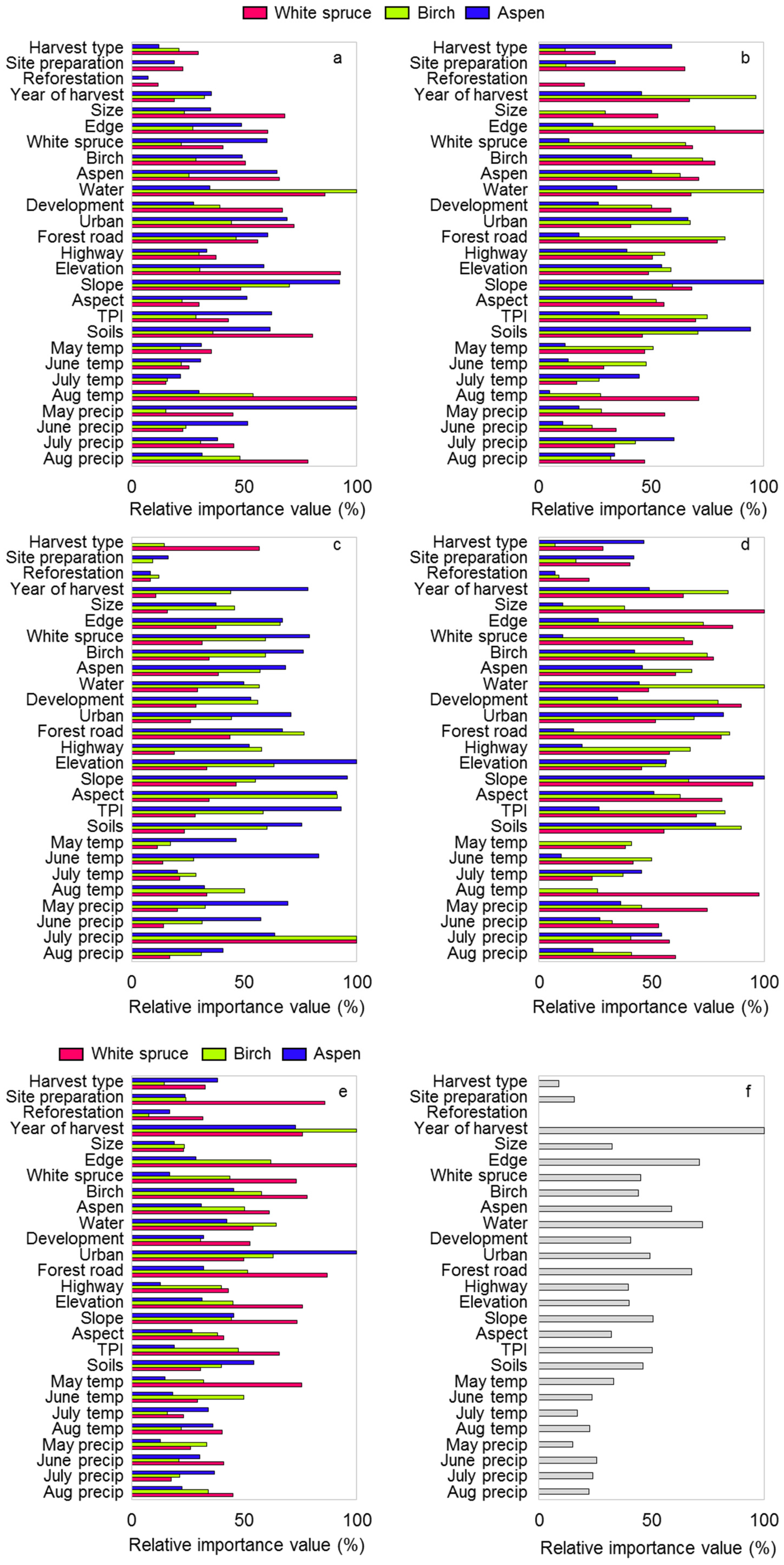
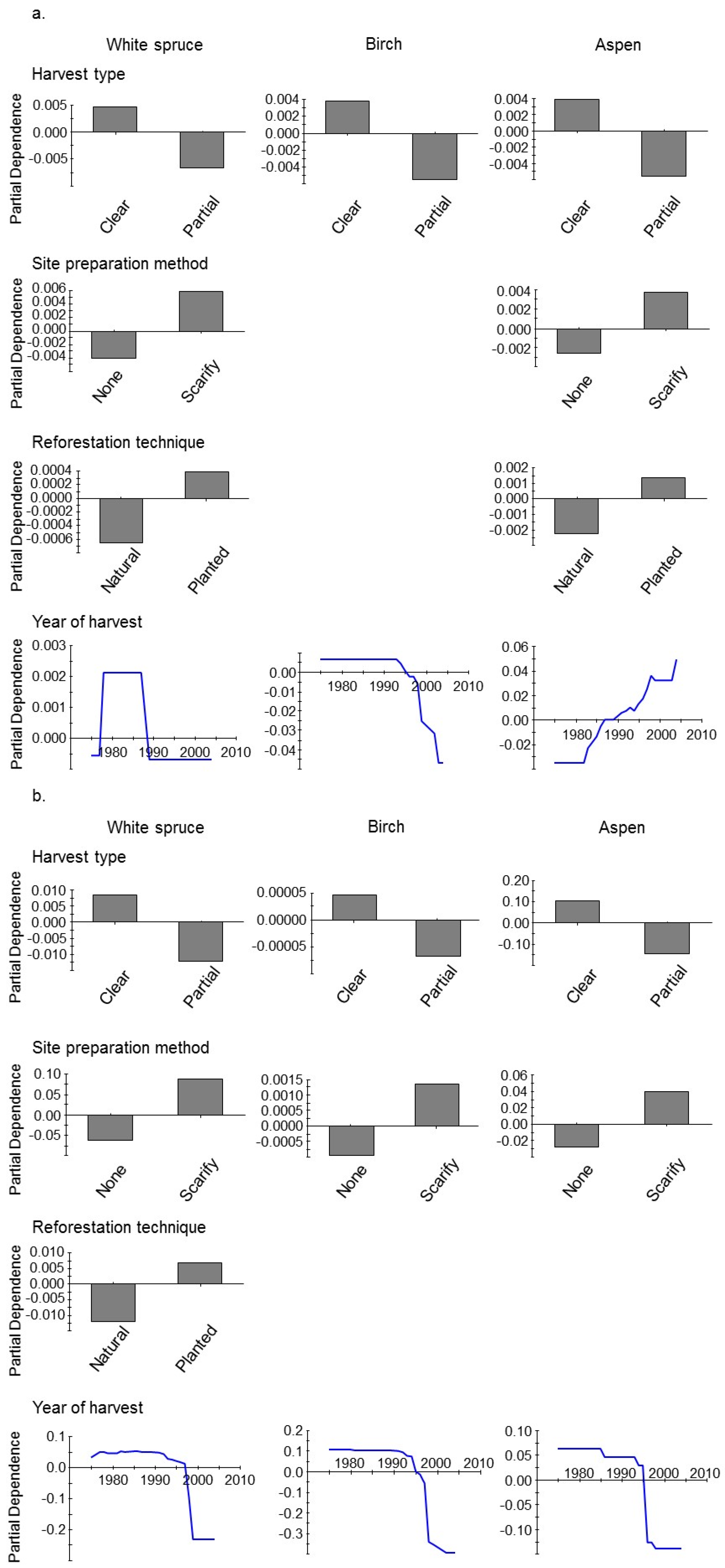
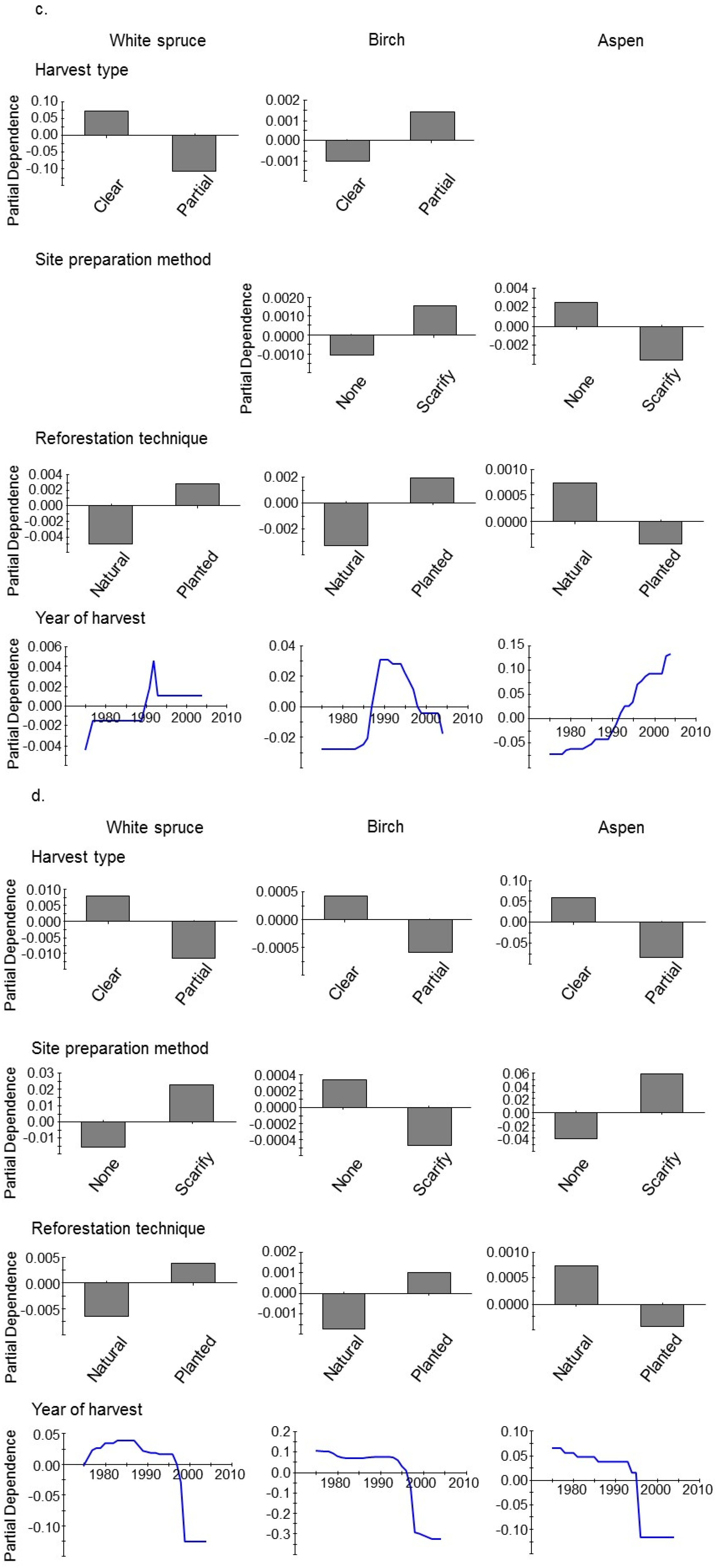
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wurtz, T.L.; Gasbarro, A.F. A brief history of wood use and forest management in Alaska. For. Chron. 1996, 72, 47–50. [Google Scholar] [CrossRef]
- Youngblood, A.; Cole, E.; Newton, M. Survival and growth response of white spruce stock types to site preparation in Alaska. Can. J. For. Res. 2011, 41, 793–809. [Google Scholar] [CrossRef]
- Youngblood, A.P.; Zasada, J.C. White spruce artificial regeneration options on river floodplains in interior Alaska. Can. J. For. Res. 1991, 21, 423–433. [Google Scholar] [CrossRef]
- Densmore, R.V.; Juday, G.P.; Zasada, J.C. Regeneration alternatives for upland white spruce after burning and logging in interior Alaska. Can. J. For. Res. 1999, 29, 413–423. [Google Scholar] [CrossRef]
- Wurtz, T.L.; Zasada, J.C. An alternative to clear-cutting in the boreal forest of Alaska: A 27-year study of regeneration after shelterwood harvesting. Can. J. For. Res. 2001, 31, 999–1011. [Google Scholar] [CrossRef]
- Chapin, F.; Fastie, C.; Viereck, L.; Ott, R.; Adams, P.; Mann, D.; Van Cleve, K.; Johnstone, J. Successional processes in the Alaskan Boreal Forest. In Alaska’s Changing Boreal Forest; Chapin, F., Oswood, M., Van Cleve, K., Viereck, L., Verbyla, D., Eds.; Oxford University Press: New York, NY, USA, 2006; pp. 100–120. [Google Scholar]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.Z.; Schepaschenko, D.G. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Foote, M.J. Classification, Description, and Dynamics of Plant Communities after Fire in the Taiga of Interior Alaska; U.S. Department of Agriculture, Pacific Northwest Forest and Range Experiment Station, Ed.; U.S. Department of Agriculture, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1983.
- Burton, P.J.; Parisien, M.-A.; Hicke, J.A.; Hall, R.J.; Freeburn, J.T. Large fires as agents of ecological diversity in the north american boreal forest. Int. J. Wildland Fire 2008, 17, 754–767. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Lovecraft, A.L.; Zavaleta, E.S.; Nelson, J.; Robards, M.D.; Kofinas, G.P.; Trainor, S.F.; Peterson, G.D.; Huntington, H.P.; Naylor, R.L. Policy strategies to address sustainability of alaskan boreal forests in response to a directionally changing climate. Proc. Natl. Acad. Sci. USA 2006, 103, 16637–16643. [Google Scholar] [CrossRef] [PubMed]
- Calef, M.P.; Varvak, A.; McGuire, A.D.; Chapin, F.S., III; Reinhold, K.B. Recent changes in annual area burned in interior Alaska: The impact of fire management. Earth Interact. 2015, 19, 1–17. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Hollingsworth, T.N.; Chapin, F.S., III; Mack, M.C. Changes in fire regime break the legacy lock on successional trajectories in Alaskan boreal forest. Glob. Chang. Biol. 2010, 16, 1281–1295. [Google Scholar] [CrossRef]
- Hollingsworth, T.N.; Johnstone, J.F.; Bernhardt, E.L.; Chapin, F.S., III. Fire severity filters regeneration traits to shape community assembly in Alaska’s boreal forest. PLoS ONE 2013, 8, e56033. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, A.; Johnstone, J.F.; Kasischke, E.S.; Kielland, K. Persistent effects of fire severity on early successional forests in interior alaska. For. Ecol. Manag. 2011, 261, 381–390. [Google Scholar] [CrossRef]
- Valentine, D.W.; Kielland, K.; Chapin, F.S., III; McCuire, A.D.; Van Cleve, K. Patterns of biogeochemistry in Alaskan boreal forests. In Alaska’s Changing Boreal Forest; Oxford University Press: New York, NY, USA, 2006. [Google Scholar]
- Johnstone, J.; Chapin, F. Effects of soil burn severity on post-fire tree recruitment in boreal forest. Ecosystems 2006, 9, 14–31. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Kasischke, E.S. Stand-level effects of soil burn severity on postfire regeneration in a recently burned black spruce forest. Can. J. For. Res. 2005, 35, 2151–2163. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Chapin, F.S.; Foote, J.; Kemmett, S.; Price, K.; Viereck, L. Decadal observations of tree regeneration following fire in boreal forests. Can. J. For. Res. 2004, 34, 267–273. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Rupp, T.S.; Olson, M.; Verbyla, D. Modeling impacts of fire severity on successional trajectories and future fire behavior in Alaskan boreal forests. Landsc. Ecol. 2011, 26, 487–500. [Google Scholar] [CrossRef]
- Viereck, L.A.; Schandelmeier, L.A. Effects of Fire in Alaska and Adjacent Canada: A Literature Review; U.S. Department of the Interior, Bureau of Land Management, Alaska State Office: Anchorage, AK, USA, 1980.
- Purdy, B.G.; Macdonald, S.E.; Dale, M.R.T. The regeneration niche of white spruce following fire in the mixedwood boreal forest. Silva Fenn. 2002, 36, 289–306. [Google Scholar] [CrossRef]
- Boateng, J.O.; Heineman, J.; Bedford, L.; Harper, G.; Nemec, A.F.L. Long-term effects of site preparation and postplanting vegetation control on picea glauca survival, growth and predicted yield in boreal british columbia. Scand. J. For. Res. 2009, 24, 111–129. [Google Scholar] [CrossRef]
- Alaska Division of Forestry. Forest management database. Data obtained from Alaska Division of Forestry, Fairbanks, Alaska; 2013. [Google Scholar]
- Wurtz, T.; Ott, R.; Maishc, J. Timber harvest in interior Alaska. In Alaska’s Changing Boreal Forest; Chapin, F., Oswood, M., Van Cleve, K., Viereck, L., Verbyla, D., Eds.; Oxford University Press: New York, NY, USA, 2006; pp. 302–308. [Google Scholar]
- Timoney, K.P.; Peterson, G. Failure of natural regeneration after clearcut logging in wood buffalo national park, Canada. For. Ecol. Manag. 1996, 87, 89–105. [Google Scholar] [CrossRef]
- Franklin, J.F.; Berg, D.R.; Thornburgh, D.A.; Tappeiner, J.C. Alternative silvicultural approaches to timber harvesting: Variable retention harvest systems. In Creating a Forestry for the 21st Century: The Science of Ecosystem Management; Kohm, K.A., Franklin, J.F., Eds.; Island Press: Washington, DC, USA, 1997; pp. 111–139. [Google Scholar]
- Roland, C.A.; Schmidt, J.H.; Johnstone, J.F. Climate sensitivity of reproduction in a mast-seeding boreal conifer across its distributional range from lowland to treeline forests. Oecologia 2014, 174, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Juday, G.P.; Barber, V.; Rupp, S.; Zasada, J.; Wilmking, M. A 200-year perspective of climate variability and the response of white spruce in Interior Alaska. In Climate Variability and Ecosystem Response at Long-Term Ecological RESEARCH Sites; Greenland, D., Goodin, D.G., Smith, R.C., Eds.; Oxford University Press: New York, NY, USA, 2003. [Google Scholar]
- Zasada, J.C.; Gregory, R.A. Regeneration of White Spruce With Reference to Interior Alaska: A Literature Review; Research Paper PNW-79; Institute of Northern Forestry, U.S. Forest Service: Washington, DC, USA, 1969; p. 37.
- Youngblood, A.; Max, T.A. Dispersal of White Spruce Seed on Willow Island in Interior Alaska; USDA Forest Service Pacific Northwest Research Station Research Paper; USDA: Washington, DC, USA, 1992; pp. U1–U17.
- Youngblood, A. Regenerating White Spruce in Boreal Forests of Alaska. Land and Ecosystem Management, 2012. Available online: http://www.fs.fed.us/pnw/lwm/lem/projects/youngblood_alaska.shtml (accessed on 2 May 2013).
- Lieffers, V.I.; Macdonald, S.E.; Hogg, E.H. Ecology of and control strategies for Calamagrostis Canadensis in boreal forest sites. Can. J. For. Res. 1993, 23, 2070–2077. [Google Scholar] [CrossRef]
- Alaska Division of Forestry. Reforestation Handbook; Alaska Department of Natural Resources Division of Forestry: Fairbanks, AK, USA, 2008.
- Fresco, N.; Chapin, F.S., III. Assessing the Potential for Conversion to Biomass Fuels in Interior Alaska; U S Forest Service Pacific Northwest Research Station Research Paper PNW-RP; US Forest Service: Washington, DC, USA, 2009; pp. 1–56.
- Alaska Energy Authority. Alaska Wood Energy Development Task Force Project Status; AWEDTG_Status_081315_8.5×11; AEA: Anchorage, AK, USA, 2015. [Google Scholar]
- Janowiak, M.K.; Webster, C.R. Promoting ecological sustainability in woody biomass harvesting. J. For. 2010, 108, 16–23. [Google Scholar]
- Burton, P.J.; Messier, C.; Adamowicz, W.L.; Kuuluvainen, T. Sustainable management of Canada’s boreal forests: Progress and prospects. Ecoscience 2006, 13, 234–248. [Google Scholar] [CrossRef]
- Larsson, S.; Danell, K. Science and the management of boreal forest biodiversity. Scand. J. For. Res. 2001, 16, 5–9. [Google Scholar] [CrossRef]
- Roessler, J.S.; Packee, E.C. Disturbance history of the Tanana River Basin in Alaska: Management implications. In Proceedings of the Annual Tall Timbers Fire Ecology Conference. Fire and Forest Ecology: Innovative Silviculture and Vegetation Management, Tallahassee, FL, USA, 2000; pp. 46–57.
- Youngblood, A. Regenerating white spruce in boreal forests of Alaska. Available online: http://www.fs.fed.us/pnw/lwm/lem/projects/youngblood_alaska.shtml (accessed on 14 May 2016).
- Labau, V.J.; van Hees, W. An inventory of Alaska’s boreal forests: Their extent, condition, and potential use. In Proceedings of the International Symposium on Boreal Forests: Condition, Dynamics, Anthropogenic Effects, Archangelsk, Russia, 16–26 July 1990.
- Hanson, D. Timber Inventory of State Forest Lands in the Tanana Valley 2013; Alaska Department of Natural Resources Division of Forestry: Fairbanks, AK, USA, 2013.
- Viereck, L.A.; Dyrness, C.T.; Van Cleve, K.; Foote, M.J. Vegetation, soils, and forest productivity in selected forest types in interior Alaska. Canadian Journal of Forest Research 1983, 13, 703–720. [Google Scholar] [CrossRef]
- Ping, C.; Boone, R.; Clark, M.; Packee, E.; Swanson, D. State factor control of soil formation in interior alaska. In Alaska’s Changing Boreal Forest; Chapin, F., Oswood, M., Van Cleve, K., Viereck, L., Verbyla, D., Eds.; Oxford University Press, Inc.: New York, NY, USA, 2006; pp. 21–38. [Google Scholar]
- Shulski, M.; Wendler, G. The Climate of Alaska; University of Alaska Press: Fairbanks, AK, USA, 2007. [Google Scholar]
- Wendler, G.; Shulski, M. A century of climate change for Fairbanks, Alaska. Arctic 2009, 62, 295–300. [Google Scholar] [CrossRef]
- Nowacki, G.; Spencer, P.; Fleming, M.; Brock, T.; Jorgenson, T. Ecoregions of Alaska: 2001; U.S. Geological Survey Open-File Report 02-297 (map); U.S. Geological Survey: Reston, VA, USA, 2001.
- The Society of American Foresters. Dictionary of Forestry; The Society of American Foresters: Bethesda, MD, USA, 1994. [Google Scholar]
- Alaska Division of Forestry. Annual Report; Alaska Department of Natural Resources Division of Forestry: Anchorage, AK, USA, 2000.
- Juday, G.P. Monitoring hectare-scale forest reference stands at Bonanza Creek Experimental Forest LTER. In Long-Term Silvicultural & Ecological Studies: Results for Science and Management; Camp, A.E., Irland, L.C., Carroll, C.J.W., Eds.; Global Institute of Sustainable Forestry, School of Forestry & Environmental Studies, Yale University: New Haven, CT, USA, 2012; pp. 31–48. [Google Scholar]
- Bonanza Creek LTER. Available online: http://www.Lter.Uaf.Edu/ (accessed on 13 September 2016).
- Yarie, J.; Kane, E.; Mack, M.C. Aboveground biomass equations for trees of Interior Alaska. In Agricultural and Forestry Experiment Station Bulletin; Univesity of Alaska Fairbanks: Fairbanks, Alaska, USA, 2007. [Google Scholar]
- Jenness, J.; Brost, B.; Beier, P. Land Facet Corridor Designer: Extension for ArcGIS. 2013. Available online: http://www.jennessent.com/arcgis/land_facets.htm (accessed on 20 May 2014).
- Alaska Division of Forestry. Vegetation and Community Mapping of the Tanana Valley; Alaska Department of Natural Resources Division of Forestry Northern Region: Fairbanks, AK, USA, 2013.
- Web soil survey. Available online: http://websoilsurvey.sc.egov.usda.gov/App/HomePage.htm (accessed on 23 August 2016).
- Snap data. Available online: http://ckan.snap.uaf.edu/dataset (accessed on 24 May 2016).
- Wilmking, M.; Juday, G.P.; Barber, V.A.; Zald, H.S.J. Recent climate warming forces contrasting growth responses of white spruce at treeline in Alaska through temperature thresholds. Glob. Chang. Biol. 2004, 10, 1724–1736. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Duffy, P.A.; Mann, D.H. Nonlinear responses of white spruce growth to climate variability in interior Alaska. Can. J. For. Res. 2013, 43, 331–343. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Juday, G.P.; Alix, C.; Barber, V.A.; Winslow, S.E.; Sousa, E.E.; Heiser, P.; Herriges, J.D.; Goetz, S.J. Changes in forest productivity across alaska consistent with biome shift. Ecol. Lett. 2011, 14, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Juday, G.P.; Alix, C. Consistent negative temperature sensitivity and positive influence of precipitation on growth of floodplain Picea glauca in Interior Alaska. Can. J. For. Res. 2012, 42, 561–573. [Google Scholar] [CrossRef]
- Van Cleve, K.; Viereck, L.A.; Dyrness, C.T. State factor control of soils and forest succession along the Tanana River in interior Alaska, USA. Arct. Alp. Res. 1996, 28, 388–400. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Additive logistic regression: A statistical view of boosting. Ann. Stat. 2000, 28, 337–374. [Google Scholar] [CrossRef]
- Breiman, L. Statistical modeling: The two cultures. Stat. Sci. 2001, 16, 199–215. [Google Scholar] [CrossRef]
- Betts, M.G.; Ganio, L.M.; Huso, M.M.P.; Som, N.A.; Huettmann, F.; Bowman, J.; Wintle, B.A. Comment on "Methods to account for spatial autocorrelation in the analysis of species distributional data: A review". Ecography 2009, 32, 374–378. [Google Scholar] [CrossRef]
- Ohse, B.; Huettmann, F.; Ickert-Bond, S.M.; Juday, G.P. Modeling the distribution of white spruce (Picea glauca) for Alaska with high accuracy: An open access role-model for predicting tree species in last remaining wilderness areas. Polar Biol. 2009, 32, 1717–1729. [Google Scholar] [CrossRef]
- Drew, C.A.; Wiersma, Y.; Huettmann, F. Predictive Species and Habitat Modeling in Landscape Ecology: Concepts and Applications; Springer Science & Business Media: New York, NY, USA, 2010. [Google Scholar]
- Salford Systems. Introducing Treenet; Salford Systems, Ed.; Salford Systems: San Diego, CA, USA, 2013. [Google Scholar]
- Metz, C.E. Basic principles of ROC analysis. Sem. Nucl. Med. 1978, 8, 283–298. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference and Prediction; Springer: New York, NY, USA, 2009. [Google Scholar]
- Nienstaedt, H.; Zasada, J.C. Picea glauca (moench) voss, white spruce. In Silvics of North America: Volume 1. Conifers. Agriculture handbook 654; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; pp. 204–226. [Google Scholar]
- Perala, D.A. Populus tremuloides Michx. Quaking Aspen. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; pp. 1082–1115. [Google Scholar]
- Solarik, K.A.; Lieffers, V.J.; Volney, W.J.A.; Pelletier, R.; Spence, J.R. Seed tree density, variable retention, and stand composition influence recruitment of white spruce in boreal mixedwood forests. Can. J. For. Res. 2010, 40, 1821–1832. [Google Scholar] [CrossRef]
- Safford, L.O.; Bjorkbom, J.C.; Zasada, J.C. Paper birch. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; Forest Service, United States Department of Agriculture: Washington, DC, USA, 1990. [Google Scholar]
- Marquis, D.A.; Bjorkborn, J.C.; Yelenosky, G. Effect of seedbed condition and light exposure on paper birch regeneration. J. For. 1964, 62, 876–881. [Google Scholar]
- Perala, D.A.; Alm, A.A. Regeneration silviculture of birch: A review. For. Ecol. Manag. 1990, 32, 39–77. [Google Scholar] [CrossRef]
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Keenan, R.J.; Kimmins, J.P. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Gartner, S.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and management of natural regeneration of white spruce in the boreal forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]
- Cole, E.; Youngblood, A.; Newton, M. Effects of competing vegetation on juvenile white spruce (Picea Glauca (Moench) Voss) growth in Alaska. Ann. For. Sci. 2003, 60, 573–583. [Google Scholar] [CrossRef]
- Calogeropoulos, C.; Greene, D.F.; Messier, C.; Brais, S. The effects of harvest intensity and seedbed type on germination and cumulative survivorship of white spruce and balsam fir in northwestern Quebec. Can. J. For. Res. 2004, 34, 1467–1476. [Google Scholar] [CrossRef] [Green Version]
- Lamontagne, J.M.; Boutin, S. Local-scale synchrony and variability in mast seed production patterns of Picea glauca. J. Ecol. 2007, 95, 991–1000. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unit | Size (ha) | Number Plots Sampled | Logged Year | Harvest Type | Site Preparation | Reforestation |
|---|---|---|---|---|---|---|
| NC-120 | 10.4 | 41 | 1975 | Partial cut | None | Plant |
| NC-93 | 17.9 | 35 | 1975 | Partial cut | None | Natural |
| NC-190 | 5.1 | 22 | 1977 | Clearcut | Scarify | Natural |
| NC-126 | 5.7 | 22 | 1978 | Partial cut | None | Natural |
| NC-140-17 | 2.5 | 8 | 1979 | Clearcut | None | Natural |
| NC-249 | 5.0 | 22 | 1980 | Clearcut | Scarify | Natural |
| NC-362 | 4.4 | 15 | 1981 | Partial cut | None | Natural |
| NC-140-38 | 1.5 | 7 | 1982 | Clearcut | Scarify | Natural |
| NC-395 | 5.1 | 21 | 1983 | Clearcut | None | Natural |
| NC-490 | 8.4 | 32 | 1985 | Clearcut | None | Natural |
| NC-556 | 6.6 | 26 | 1986 | Clearcut | None | Plant |
| NC-305 | 3.5 | 11 | 1987 | Partial cut | Scarify | Plant |
| NC-705 | 11.0 | 44 | 1989 | Clearcut | Scarify | Plant |
| NC-454 | 20.4 | 44 | 1991 | Clearcut | Scarify | Plant |
| NC-740 | 1.9 | 8 | 1991 | Clearcut | None | Plant |
| NC-709 | 17.2 | 35 | 1991 | Clearcut | Scarify | Plant |
| NC-842 | 2.1 | 7 | 1992 | Partial cut | None | Natural |
| NC-733 | 30.3 | 44 | 1992 | Clearcut | Scarify | Plant |
| NC-702 | 2.0 | 9 | 1993 | Clearcut | None | Plant |
| NC-747 | 8.0 | 31 | 1994 | Clearcut | None | Plant |
| NC-750 | 9.8 | 41 | 1995 | Clearcut | Scarify | Plant |
| NC-1085 | 22.6 | 47 | 1996 | Partial cut | Scarify | Plant |
| NC-1137 | 13.5 | 29 | 1997 | Clearcut | None | Plant |
| NC-927 | 22.5 | 43 | 1998 | Partial cut | None | Plant |
| NC-760 | 3.4 | 13 | 1998 | Partial cut | None | Natural |
| NC-1129 | 6.0 | 22 | 1999 | Partial cut | None | Plant |
| NC-1090 | 1.4 | 7 | 1999 | Partial cut | None | Natural |
| NC-1135 | 11.7 | 49 | 2002 | Partial cut | None | Plant |
| NC-1116 | 2.4 | 9 | 2003 | Partial cut | Scarify | Natural |
| NC-1143 | 6.7 | 28 | 2004 | Partial cut | None | Natural |
| Variable | Description | Unit | Data Source |
|---|---|---|---|
| Response variables | |||
| Presence/absence | Presence/absence of white spruce, birch, and aspen | category | Field sampling |
| Species dominance | Species dominance of white spruce, birch, and aspen | category | Field sampling |
| Basal area | Basal area of white spruce (high ≥ 0.5 m3, low < 0.5 m3), birch (high ≥ 1 m3, low < 1 m3), and aspen (high ≥ 0 m3, low < 0 m3) | category | Field sampling |
| Biomass | Biomass accumulation (high ≥ 5 t, low < 5 t) | category | Field sampling |
| Predictor variables | |||
| Harvest type | Harvest type: clearcut/partial cut | category | AKDOF FMD |
| Site preparation | Ground treatment type: none/mechanical site preparation | category | AKDOF FMD |
| Reforestation | Reforestation type: natural/planting white spruce seedlings | category | AKDOF FMD |
| Year | Year since harvest: 10–39 | continuous | AKDOF FMD |
| Size | Size of harvest unit | hectare | AKDOF FMD |
| Edge | Distance to edge of harvest unit | km | AKDOF FMD |
| White spruce | Distance to white spruce forest | km | AKDOF vegetation map |
| Birch | Distance to birch forest | km | AKDOF vegetation map |
| Aspen | Distance to aspen forest | km | AKDOF vegetation map |
| Water | Distance to water | km | AKDOF vegetation map |
| Highway | Distance to highway | km | AKDOF vegetation map |
| Forest road | Distance to forest road | km | AKDOF vegetation map |
| Urban | Distance to urban area | km | AKDOF vegetation map |
| Development | Distance to development (power line, mine etc.) | km | AKDOF vegetation map |
| Elevation | Elevation | m | GINA DEM |
| Slope | Slope | degree | GINA DEM |
| Aspect | Aspect | category | GINA DEM |
| TPI | Topographic Position Index | continuous | GINA DEM |
| Soils | Soil subgroup | category | NRCS |
| May temp | Average temperature of May | °C | SNAP |
| June temp | Average temperature of June | °C | SNAP |
| July temp | Average temperature of July | °C | SNAP |
| Aug temp | Average temperature of August | °C | SNAP |
| May precip | Precipitation sum of May | mm | SNAP |
| June precip | Precipitation sum of June | mm | SNAP |
| July precip | Precipitation sum of July | mm | SNAP |
| Aug precip | Precipitation sum of August | mm | SNAP |
| Predictions | Specificity Sensitivity | Mean Accuracy | AUC | |||
|---|---|---|---|---|---|---|
| “All Size” Group Presence/Absence | Absent | Present | ||||
| Aspen | Absent | 491 | 92 | 84.22% | 0.84 | 0.92 |
| Present | 22 | 121 | 84.62% | |||
| Birch | Absent | 176 | 91 | 65.92% | 0.68 | 0.74 |
| Present | 138 | 321 | 69.93% | |||
| White spruce | Absent | 196 | 74 | 72.59% | 0.73 | 0.78 |
| Present | 123 | 333 | 73.03% | |||
| Saplings presence/absence | Absent | Present | ||||
| Aspen | Absent | 653 | 43 | 93.82% | 0.94 | 0.98 |
| Present | 2 | 28 | 93.33% | |||
| Birch | Absent | 394 | 84 | 82.43% | 0.82 | 0.90 |
| Present | 47 | 201 | 81.05% | |||
| White spruce | Absent | 422 | 108 | 79.62% | 0.79 | 0.88 |
| Present | 44 | 152 | 77.55% | |||
| “All size” group dominance | Low | High | ||||
| Aspen | Low | 580 | 79 | 88.01% | 0.88 | 0.95 |
| High | 9 | 58 | 86.57% | |||
| Birch | Low | 296 | 84 | 77.89% | 0.78 | 0.85 |
| High | 78 | 268 | 77.46% | |||
| White spruce | Low | 377 | 115 | 76.63% | 0.76 | 0.83 |
| High | 58 | 176 | 75.21% | |||
| Sapling dominance | Low | High | ||||
| Aspen | Low | 656 | 44 | 93.71% | 0.94 | 0.97 |
| High | 2 | 24 | 92.31% | |||
| Birch | Low | 423 | 83 | 83.60% | 0.84 | 0.90 |
| High | 36 | 184 | 83.64% | |||
| White spruce | Low | 472 | 111 | 80.96% | 0.81 | 0.88 |
| High | 27 | 116 | 81.12% | |||
| Basal area | Low | High | ||||
| Aspen | Low | 103 | 6 | 94.50% | 0.94 | 0.98 |
| High | 2 | 32 | 94.12% | |||
| Birch | Low | 194 | 30 | 86.61% | 0.86 | 0.93 |
| High | 33 | 201 | 85.90% | |||
| White spruce | Low | 190 | 43 | 81.55% | 0.81 | 0.88 |
| High | 45 | 177 | 79.73% | |||
| Biomass | Low | High | 0.78 | 0.85 | ||
| Low | 319 | 87 | 78.57% | |||
| High | 72 | 261 | 78.38% | |||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morimoto, M.; Juday, G.; Young, B. Clearcutting and Site Preparation, but Not Planting, Promoted Early Tree Regeneration in Boreal Alaska. Forests 2017, 8, 12. https://doi.org/10.3390/f8010012
Morimoto M, Juday G, Young B. Clearcutting and Site Preparation, but Not Planting, Promoted Early Tree Regeneration in Boreal Alaska. Forests. 2017; 8(1):12. https://doi.org/10.3390/f8010012
Chicago/Turabian StyleMorimoto, Miho, Glenn Juday, and Brian Young. 2017. "Clearcutting and Site Preparation, but Not Planting, Promoted Early Tree Regeneration in Boreal Alaska" Forests 8, no. 1: 12. https://doi.org/10.3390/f8010012