Forest Type and Tree Characteristics Determine the Vertical Distribution of Epiphytic Lichen Biomass in Subtropical Forests


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
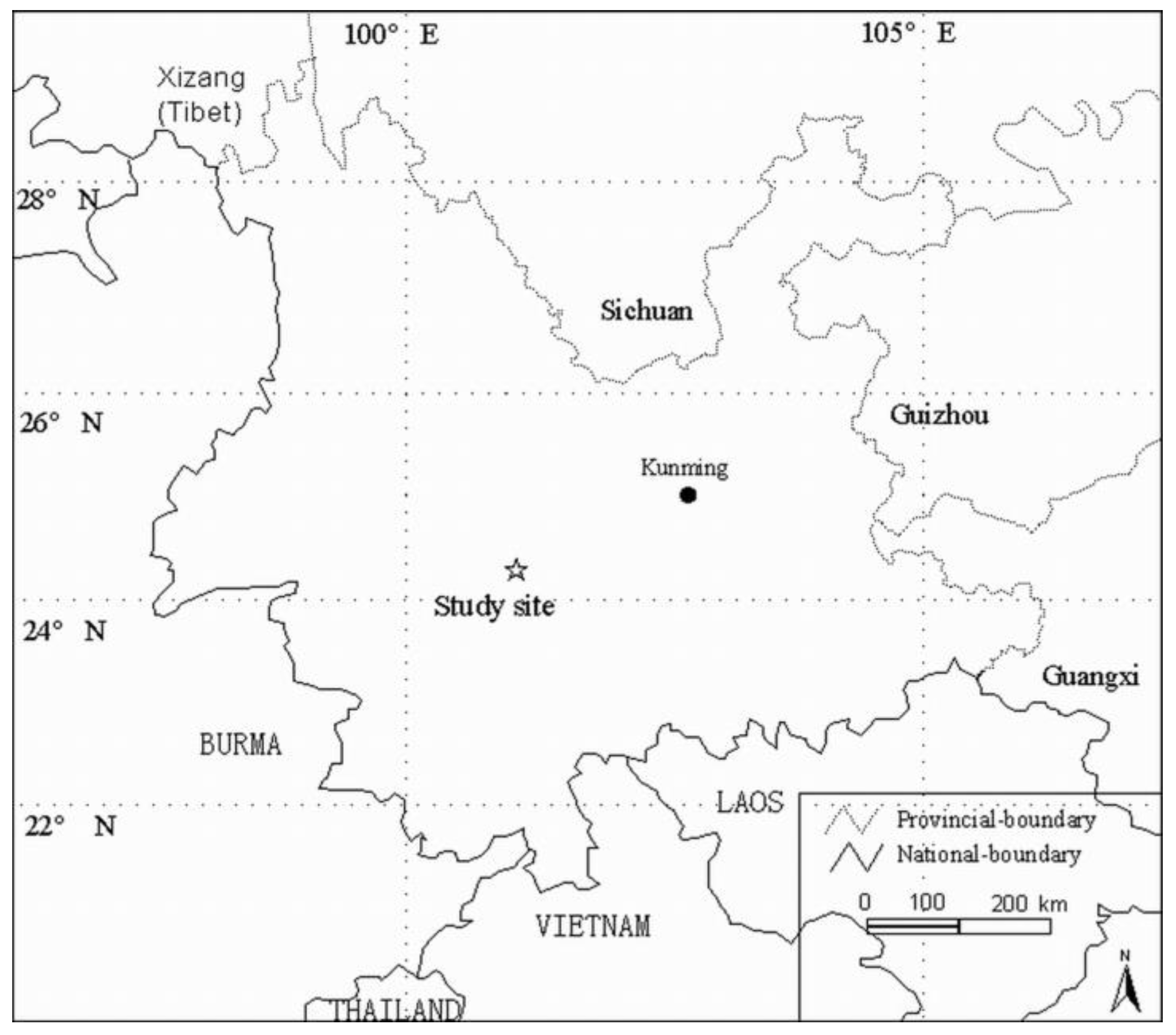
2.1. Study Area
2.2. Sampling Method
2.3. Data Analysis
3. Results
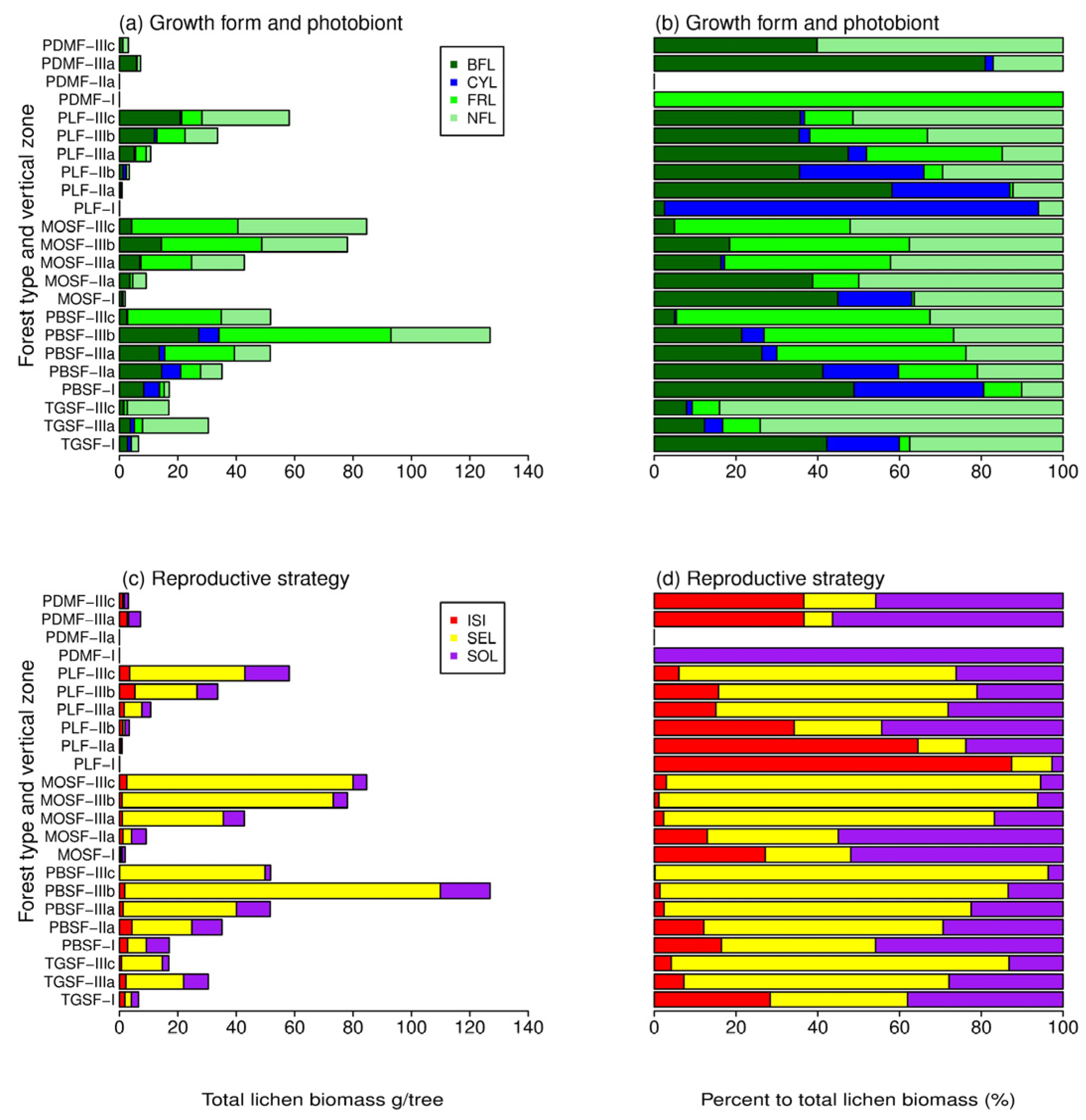
3.1. Biomass among Forest Types
3.2. Biomass among Host Species
3.3. The Effect of Tree Attributes
3.4. Vertical Distribution among Height Segments
3.5. Vertical Distribution among Vertical Zones
4. Discussion
4.1. Influences of Forest Type and Host Attribute
4.2. Vertical Distribution
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sillett, S.C.; Antoine, M.E. Lichens and Bryophytes in Forest Canopies. In Forest Canopies, 2nd ed.; Lowman, M.D., Rinker, H.B., Eds.; Academic Press: Cambridge, CA, USA, 2004; pp. 151–174. [Google Scholar]
- Ellis, C.J. Lichen epiphyte diversity: A species, community and trait-based review. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 131–152. [Google Scholar] [CrossRef]
- Rhoades, F.M. Nonvascular Epiphytes in Forest Canopies: Worldwide Distribution, Abundance, and Ecological Roles. In Forest Canopies; Lowman, M.D., Nadkarni, N.M., Eds.; Academic Press: Cambridge, CA, USA, 1995; pp. 353–408. [Google Scholar]
- Kirkpatrick, R.C.; Zou, R.J.; Dierenfeld, E.S.; Zhou, H.W. Digestion of selected foods by Yunnan snub-nosed monkey Rhinopithecus bieti (Colobinae). Am. J. Phys. Anthropol. 2001, 114, 156–162. [Google Scholar] [CrossRef]
- Van Stan, J.T.; Pypker, T.G. A review and evaluation of forest canopy epiphyte roles in the partitioning and chemical alteration of precipitation. Sci. Total Environ. 2015, 536, 813–824. [Google Scholar] [CrossRef] [PubMed]
- McCune, B. Using epiphyte litter to estimate epiphyte biomass. Bryologist 1994, 97, 396–401. [Google Scholar] [CrossRef]
- Peck, J.L.E.; McCune, B. Remnant trees and canopy lichen communities in western Oregon: A retrospective approach. Ecol. Appl. 1997, 7, 1181–1187. [Google Scholar] [CrossRef]
- Li, S.; Liu, W.Y.; Wang, L.S.; Ma, W.Z.; Song, L. Biomass, diversity and composition of epiphytic macrolichens in primary and secondary forests in the subtropical Ailao Mountains, SW China. For. Ecol. Manag. 2011, 261, 1760–1770. [Google Scholar] [CrossRef]
- Becker, V.; Reeder, J.; Stetler, R. Biomass and habitat of nitrogen fixing lichens in an oak forest in the North Carolina Piedmont. Bryologist 1977, 80, 93–99. [Google Scholar] [CrossRef]
- Pike, L.H. The importance of epiphytic lichens in mineral cycling. Bryologist 1978, 81, 247–257. [Google Scholar] [CrossRef]
- Becker, V.E. Nitrogen fixing lichens in forests of the Southern Appalachian Mountains of North Carolina. Bryologist 1980, 83, 29–39. [Google Scholar] [CrossRef]
- Boucher, V.; Nash, T., III. The role of the fruticose lichen Ramalina menziesii in the annual turnover of biomass and macronutrients in a blue oak woodland. Bot. Gaz. 1990, 151, 114–118. [Google Scholar] [CrossRef]
- Clement, J.P.; Shaw, D.C. Crown structure and the distribution of epiphyte functional group biomass in old-growth Pseudotsuga menziesii trees. Ecoscience 1999, 6, 243–254. [Google Scholar] [CrossRef]
- Campbell, J.; Coxson, D.S. Canopy microclimate and arboreal lichen loading in subalpine spruce-fir forest. Can. J. Bot. 2001, 79, 537–555. [Google Scholar]
- Benson, S.; Coxson, D.S. Lichen colonization and gap structure in wet-temperate rainforests of northern interior British Columbia. Bryologist 2002, 105, 673–692. [Google Scholar] [CrossRef]
- Ellyson, W.J.T.; Sillett, S.C. Epiphyte communities on Sitka spruce in an old-growth redwood forest. Bryologist 2003, 106, 197–211. [Google Scholar] [CrossRef]
- Boudreault, C.; Bergeron, Y.; Coxson, D.S. Factors controlling epiphytic lichen biomass during postfire succession in black spruce boreal forests. Can. J. For. Res. 2009, 39, 2168–2179. [Google Scholar] [CrossRef]
- Liu, C.; Ilvesniemi, H.; Westman, C.J. Biomass of arboreal lichens and its vertical distribution in a boreal coniferous forest in central Finland. Lichenologist 2000, 32, 495–504. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Lie, M.; Ohlson, M. Epiphytic lichen biomass in a boreal Norway spruce forest. Lichenologist 2008, 40, 257–266. [Google Scholar] [CrossRef]
- Lang, G.E.; Reiners, W.A.; Pike, L.H. Structure and biomass dynamics of epiphytic lichen communities of balsam fir forests in New Hampshire. Ecology 1980, 61, 541–550. [Google Scholar] [CrossRef]
- McCune, B. Gradients in epiphyte biomass in three Pseudotsuga-Tsuga forests of different ages in western Oregon and Washington. Bryologist 1993, 96, 405–411. [Google Scholar] [CrossRef]
- Rhoades, F.M. Biomass of epiphytic lichens and bryophytes on Abies lasiocarpa on a Mt. Baker lava flow, Washington. Bryologist 1981, 81, 39–47. [Google Scholar] [CrossRef]
- Kobylinski, A.; Fredeen, A.L. Vertical distribution and nitrogen content of epiphytic macrolichen functional groups in sub-boreal forests of central British Columbia. For. Ecol. Manag. 2014, 329, 118–128. [Google Scholar] [CrossRef]
- Edwards, R.Y.; Soos, J.; Ritcey, R.W. Quantitative observations on epidendric lichens used as food by caribou. Ecology 1960, 41, 425–431. [Google Scholar] [CrossRef]
- Pike, L.H.; Rydell, R.A.; Denison, W.C. A 400-year-old Douglas fir tree and its epiphytes: Biomass, surface area, and their distributions. Can. J. For. Res. 1977, 7, 680–699. [Google Scholar] [CrossRef]
- Barkman, J.J. Phytosociology and Ecology of Cryptogamic Epiphytes; Van Gorcum: Assen, The Netherlands, 1958. [Google Scholar]
- Kermit, T.; Gauslaa, Y. The vertical gradient of bark pH of twigs and macrolichens in a Picea abies canopy not affected by acid rain. Lichenologist 2001, 33, 353–359. [Google Scholar] [CrossRef]
- Pipp, A.K.; Henderson, C.; Callaway, R.M. Effects of forest age and forest structure on epiphytic lichen biomass and diversity in a Douglas-fir forest. Northwest Sci. 2001, 75, 12–24. [Google Scholar]
- Fritz, Ö. Vertical distribution of epiphytic bryophytes and lichens emphasizes the importance of old beeches in conservation. Biodivers. Conserv. 2009, 18, 289–304. [Google Scholar] [CrossRef]
- Kovaleva, N.M.; Ivanova, G.A. Features of the biomass distribution of epiphytic lichens on Scotch pine (Lower Angara Region). Contemp. Probl. Ecol. 2012, 5, 319–322. [Google Scholar] [CrossRef]
- McCune, B.; Amsberry, K.A.; Camacho, F.J.; Clery, S.; Cole, C.; Emerson, C.; Felder, G.; French, P.; Greene, D.; Harris, R.; et al. Vertical profile of epiphytes in a Pacific Northwest old-growth forest. Northwest Sci. 1997, 71, 145–152. [Google Scholar]
- Färber, L.; Solhaug, K.A.; Esseen, P.A.; Bilger, W.; Gauslaa, Y. Sunscreening fungal pigments influence the vertical gradient of pendulous lichens in boreal forest canopies. Ecology 2014, 95, 1464–1471. [Google Scholar] [CrossRef] [PubMed]
- Brodo, I.M.; Sharnoff, S.D.; Sharnoff, S. Lichens of North America; Yale University Press: New Haven, CT, USA, 2001. [Google Scholar]
- Wang, L.S. Lichens of Yunnan in China; Shanghai Scientific and Technical Publishers: Shanghai, China, 2012. [Google Scholar]
- Li, S.; Liu, W.Y.; Li, D.W. Epiphytic lichens in subtropical forest ecosystems in southwest China: Species diversity and implications for conservation. Biol. Conserv. 2013, 159, 88–95. [Google Scholar] [CrossRef]
- Aptroot, A. Lichenized and saprobic fungal biodiversity of a single Elaeocarpus tree in Papua New Guinea, with the report of 200 species of ascomycetes associated with one tree. Fungal Divers. 2001, 6, 1–11. [Google Scholar]
- Li, S.; Liu, W.Y.; Li, D.W. Bole epiphytic lichens as potential indicators of environmental change in subtropical forest ecosystems in southwest China. Ecol. Indic. 2013, 29, 93–104. [Google Scholar] [CrossRef]
- Li, S.; Liu, W.Y.; Li, D.W.; Song, L.; Shi, X.M.; Lu, H.Z. Species richness and vertical stratification of epiphytic lichens in subtropical primary and secondary forests in southwest China. Fungal Ecol. 2015, 17, 30–40. [Google Scholar] [CrossRef]
- Johansson, D. Ecology of vascular epiphytes in West African rain forest. Acta Phytogeogr. Suec. 1974, 59, 1–136. [Google Scholar]
- Cornelissen, J.H.C.; Ter Steege, H. Distribution and ecology of epiphytic bryophytes and lichens in dry evergreen forest of Guyana. J. Trop. Ecol. 1989, 5, 131–150. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 21 July 2017).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S.; Christensen, R.H.B.; Singmann, H.; Dai, B.; Grothendieck, G.; Green, P. lme4: Linear Mixed-Effects Models Using ‘Eigen’ and S4. R Package Version 1.1-13. Available online: http://cran.r-project.org/web/packages/lme4/index.html (accessed on 22 July 2017).
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Nakagawa, S.; Schielzeth, H. Coefficient of determination R2 and intra-class correlation coefficient ICC from generalized linear mixed-effects models revisited and expanded. BioRxiv 2017. [Google Scholar] [CrossRef]
- Jaeger, B. r2glmm: Computes R Squared for Mixed (Multilevel) Models. R Package Version 0.1.2. Available online: https://github.com/bcjaeger/r2glmm (accessed on 24 July 2017).
- Forman, R.T.T. Canopy lichens with blue-green algae: A nitrogen source in a Colombian rain forest. Ecology 1975, 56, 1176–1184. [Google Scholar] [CrossRef]
- Neitlich, P.N. Lichen Abundance and Biodiversity Along a Chronosequence from Young Managed Stands to Ancient Forest. Master’s Thesis, University of Vermont, Burlington, VT, USA, 1993. [Google Scholar]
- Chen, L.; Liu, W.Y.; Wang, G.S. Estimation of epiphytic biomass and nutrient pools in the subtropical montane cloud forest in the Ailao Mountains, South-Western China. Ecol. Res. 2010, 25, 315–325. [Google Scholar] [CrossRef]
- Rawat, S.; Upreti, D.K.; Singh, R.P. Estimation of epiphytic lichen litter fall biomass in three temperate forests of Chamoli district, Uttarakhand, India. Trop. Ecol. 2011, 52, 193–200. [Google Scholar]
- Wolf, J.H.D. Diversity patterns and biomass of epiphytic bryophytes and lichens along an altitudinal gradient in the northern Andes. Ann. Mo. Bot. Gard. 1993, 81, 928–960. [Google Scholar] [CrossRef]
- Stone, D.F. Epiphyte succession on Quercus garryana branches in the Willamette Valley of western Oregon. Bryologist 1989, 92, 81–94. [Google Scholar] [CrossRef]
- Sillett, S.C.; Neitlich, P.N. Emerging themes in epiphyte research in westside forests with special reference to cyanolichens. Northwest Sci. 1996, 70, 54–60. [Google Scholar]
- Ellis, C.J.; Coppins, B.J. Contrasting functional traits maintain lichen epiphyte diversity in response to climate and autogenic succession. J. Biogeogr. 2006, 33, 1643–1656. [Google Scholar] [CrossRef]
- Coote, L.; Smith, G.F.; Kelly, D.L.; O’Donoghue, S.; Dowding, P.; Iremonger, S.; Mitchell, F.J.G. Epiphytes of Sitka spruce (Picea sitchensis) plantations in Ireland and the effects of open spaces. Biodivers. Conserv. 2007, 16, 4009–4024. [Google Scholar] [CrossRef]
- Hale, M.E. The Biology of Lichens; Edward Arnold: London, UK, 1967. [Google Scholar]
- Hedenås, H.; Ericson, L. Epiphytic macrolichens as conservation indicators: Successional sequence in Populus tremula stands. Biol. Conserv. 2000, 93, 43–53. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Solhaug, K.A. Differences in the susceptibility to light stress between epiphytic lichens of ancient and young boreal forest stands. Funct. Ecol. 1996, 10, 344–354. [Google Scholar] [CrossRef]
- Dettki, H.; Klintberg, P.; Esseen, P.A. Are epiphytic lichens in young forests limited by local dispersal? Ecoscience 2000, 7, 317–325. [Google Scholar] [CrossRef]
- Hilmo, O.; Såstad, S.M. Colonization of old-forest lichens in a young and an old boreal Picea abies forest: An experimental approach. Biol. Conserv. 2001, 102, 251–259. [Google Scholar] [CrossRef]
- Rogers, R.W. Ecological strategies of lichens. Lichenologist 1990, 22, 149–162. [Google Scholar] [CrossRef]
- Ellis, C.J.; Coppins, B.J. Reproductive strategy and the compositional dynamics of crustose lichen communities on aspen (Populus tremula L.) in Scotland. Lichenologist 2007, 39, 377–391. [Google Scholar] [CrossRef]
- Liu, W.Y.; Fox, J.E.D.; Xu, Z.F. Nutrient fluxes in bulk precipitation, throughfall and stemflow in montane subtropical moist forest on Ailao Mountains in Yunnan, South-West China. J. Trop. Ecol. 2002, 18, 527–548. [Google Scholar] [CrossRef]
- Xu, H.Q.; Liu, W.Y. Species diversity and distribution of epiphytes in the montane moist evergreen broad-leaved forest in Ailao Mountain, Yunnan. Biodivers. Sci. 2005, 13, 137–147. [Google Scholar] [CrossRef][Green Version]
- Wang, G.S.; Liu, W.Y.; Fu, Y.; Yang, G.P. Comparison of physical and chemical properties and microbial biomass and enzyme activities of humus from canopy and forest floor in a montane moist evergreen broad-leaved forest in Ailao Mts., Yunnan. Acta Ecol. Sin. 2008, 28, 1328–1336. [Google Scholar]
- Li, S.; Liu, W.Y.; Li, D.W.; Li, Z.X.; Song, L.; Chen, K.; Fu, Y. Slower rates of litter decomposition of dominant epiphytes in the canopy than on the forest floor in a subtropical montane forest, southwest China. Soil Biol. Biochem. 2014, 70, 211–220. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Primary Dwarf Mossy Forest (PDMF) | Primary Lithocarpus Forest (PLF) | Middle-Aged Oak Secondary Forest (MOSF) | Populus bonatii Secondary Forest (PBSF) | Ternstroemia gymnanthera Secondary Forest (TGSF) |
|---|---|---|---|---|---|
| Dominant canopy species | Lithocarpus crassifolius A. Camus; Rhododendron irroratum Franch. | Lithocarpus xylocarpus (Kurz) Markgr.; Lithocarpus hancei (Benth.) Rehder; Castanopsis wattii (King ex Hook. f.) A. Camus | Lithocarpus hancei; Vaccinium duclouxii (H. Lév.) Hand.-Mazz. | Populus bonatii Levl. | Ternstroemia gymnanthera (Wigrt et Arn.) Bedd |
| Stand age (year) | >300 | >300 | 49 | 36 | <20 |
| Basal area (m2/ha) | 48.00 | 77.31 | 55.17 | 29.88 | 42.45 |
| Canopy openness (%) | 30.00 | 5.80 | 31.00 | 51.67 | 31.67 |
| Mean diameter at breast height (dbh, cm) | 7.91 | 19.23 | 9.84 | 8.50 | 4.31 |
| Tree density (trees/ha) | 8273 | 1656 | 5903 | 4697 | 22,933 |
| Tree species richness | 15.90 | 15.72 | 15.73 | 8.87 | 5.67 |
| Species | Functional Group | Forest Type | ||||
|---|---|---|---|---|---|---|
| PDMF | PLF | MOSF | PBSF | TGSF | ||
| Anzia leucobatoides f. hypomelaena A. Zahlbruckner | NFL/SEL | 0.000 | 3.906 | 3.026 | 0.161 | 0.000 |
| Anzia physoidea A. L. Sm | NFL/SEL | 0.000 | 0.220 | 0.063 | 0.000 | 0.000 |
| Bryoria confusa (D. D. Awasthi) Brodo & D. Hawksw. | FRL/SEL | 0.000 | 0.351 | 2.280 | 0.000 | 0.000 |
| Cetrelia olivetorum (Nyl.) W. L. Culb. & C. F. Culb. | BFL/SOL | 3.393 | 7.563 | 7.172 | 16.521 | 2.713 |
| Cladonia coniocraea (Flök.) Spreng. | FRL/SOL | 0.001 | 0.221 | 0.005 | 0.017 | 0.015 |
| Coccocarpia erythroxyli (Spreng.) Swinscow & Krog | CYL/SEL | <0.001 | 0.000 | 0.002 | 0.038 | 0.008 |
| Collema fasciculare (L.) Weber ex F. H. Wigg. | CYL/SEL | 0.000 | 0.000 | 0.000 | 0.000 | 0.008 |
| Erioderma meiocarpum Nyl. | CYL/SEL | 0.000 | 0.000 | 0.000 | 0.017 | 0.000 |
| Everniastrum cirrhatum (Fr.) Hale ex Sipman | NFL/SEL | <0.001 | 1.586 | 15.308 | 20.403 | 3.601 |
| Everniastrum nepalense (Taylor) Hale ex Sipman | NFL/SEL | 0.000 | 8.265 | 22.522 | 2.871 | 14.532 |
| Everniastrum rhizodendroideum (J. C. Wei & Y. M. Jiang) Sipman | NFL/SEL | 0.000 | 0.017 | 0.753 | 0.060 | 0.641 |
| Heterodermia boryi (Fée) Hale | NFL/SEL | 0.000 | 1.660 | 0.481 | 1.409 | 0.048 |
| Heterodermia comosa (Eschw.) Follmann & Redón | NFL/SEL | 0.164 | 0.424 | 1.979 | 1.238 | 0.240 |
| Heterodermia dendritica (Pers.) Poelt | NFL/SEL | 0.163 | 3.475 | 10.195 | 2.673 | 1.787 |
| Heterodermia hypoleuca (Mühlbr.) Trevis. | NFL/SEL | 0.116 | 0.437 | 0.003 | 14.434 | 0.000 |
| Heterodermia obscurata (Nyl.) Trevis. | NFL/SOL | 0.000 | 0.000 | 0.000 | 0.000 | 0.218 |
| Hypogymnia yunnanensis Y. M. Jiang & J. C. Wei | NFL/SEL | 0.219 | 3.230 | 9.999 | <0.001 | 2.154 |
| Hypotrachyna adducta (Nyl.) Hale | NFL/SEL | 0.074 | 1.676 | 7.476 | 0.353 | 2.135 |
| Hypotrachyna pseudosinuosa (Asahina) Hale | NFL/SOL | 1.290 | 8.685 | 7.394 | 0.437 | 4.388 |
| Hypotrachyna revoluta (Flörke) Hale | NFL/SOL | 0.000 | 0.000 | 0.000 | 0.000 | 0.015 |
| Hypotrachyna sinuosa (Sm.) Hale | NFL/SOL | <0.001 | 0.002 | 0.000 | 0.051 | 0.000 |
| Leioderma sorediatum D. J. Galloway & P. M. Jørg. | CYL/SOL | <0.001 | 0.165 | 0.099 | 0.018 | 0.237 |
| Leptogium azureum (Sw. ex Ach.) Mont. | CYL/SEL | 0.000 | 0.140 | 0.301 | 1.103 | 0.140 |
| Leptogium menziesii (Sm.) Mont. | CYL/SEL | 0.000 | 0.154 | 0.103 | 3.013 | 0.000 |
| Leptogium saturninum (Dicks.) Nyl. | CYL/ISL | 0.000 | 0.000 | 0.020 | 0.134 | 0.000 |
| Lobaria isidiophora Yoshim. | BFL/ISL | 0.000 | 2.231 | 0.000 | 1.179 | 0.000 |
| Lobaria isidiosa (Müll. Arg.) Vain. | CYL/ISL | 0.000 | 0.000 | 0.000 | 1.143 | 0.137 |
| Lobaria kurokawae Yoshim. | CYL/SEL | 0.000 | 0.010 | 0.000 | 0.000 | 0.000 |
| Lobaria retigera (Bory) Trevis. | CYL/ISL | 0.138 | 1.371 | 0.252 | 2.704 | 1.097 |
| Menegazzia terebrata (Hoffm.) A. Massal. | NFL/SOL | <0.001 | 1.371 | 1.320 | 1.072 | 0.292 |
| Myelochroa aurulenta (Tuck.) Elix & Hale | NFL/SOL | 0.000 | 0.000 | 0.000 | 0.000 | 3.650 |
| Myelochroa irrugans (Nyl.) Elix & Hale | NFL/SEL | 0.000 | 0.215 | 0.137 | 0.373 | 0.757 |
| Myelochroa subaurulenta (Ny1) Elix & Hale | NFL/SEL | 0.000 | 0.000 | 0.000 | 0.000 | 1.383 |
| Nephroma helveticum Ach. | CYL/ISL | 0.001 | 0.242 | 0.000 | 0.103 | 0.748 |
| Nephromopsis ornata (Müll. Arg.) Hue | BFL/SEL | 0.000 | 8.850 | 6.994 | 0.102 | 0.004 |
| Nephromopsis pallescens (Schaer.) Y. S. Park | BFL/SEL | 0.000 | 1.969 | 2.742 | 0.693 | 0.796 |
| Nephromopsis stracheyi (Bab.) Müll. Arg. | BFL/SEL | 0.000 | 0.535 | 0.000 | 0.000 | 0.000 |
| Oropogon asiaticus Asahina | FRL/SEL | 0.000 | 0.000 | 0.000 | 0.519 | 0.000 |
| Pannaria rubiginosa (Thunb.) Delise | CYL/SEL | 0.000 | 0.000 | 0.000 | 4.215 | 0.000 |
| Parmelia adaugescens Nyl. | NFL/SEL | 0.000 | 0.789 | 0.773 | 0.041 | 0.653 |
| Parmelina quercina (Willd.) Hale | NFL/SEL | 0.000 | 1.259 | 3.348 | 0.059 | 0.682 |
| Parmotrema eciliatum (Nyl.) Hale | BFL/SEL | 0.000 | 0.589 | 0.065 | 8.074 | 0.000 |
| Parmotrema mellissii (C. W. Dodge) Hale | BFL/ISL | 0.000 | 0.000 | 0.000 | 0.000 | 0.146 |
| Parmotrema reticulatum (Taylor) M. Choisy | BFL/SOL | <0.001 | 4.645 | 1.387 | 14.888 | 1.326 |
| Parmotrema tinctorum (Desper. ex Nyl.) Hale | BFL/ISL | 2.994 | 5.589 | 4.647 | 0.863 | 2.527 |
| Peltigera rufescens (Weiss) Humb. | CYL/SEL | 0.000 | 0.000 | 0.000 | 0.000 | 0.132 |
| Phaeophyscia ciliate (Hoffm.) Moberg | NFL/SEL | 0.000 | 0.000 | 0.000 | 0.796 | 0.000 |
| Ramalina conduplicans Vain. | FRL/SEL | 0.000 | 3.421 | 16.915 | 57.671 | 0.297 |
| Ramalina sinensis Jatta | FRL/SEL | 0.000 | 0.000 | 0.000 | 0.191 | 0.000 |
| Rimelia cetrata (Ach.) Hale & A. Fletcher. | BFL/SEL | 0.000 | 0.000 | 0.000 | 0.000 | 0.122 |
| Sticta duplolimbata (Hue) Vain. | CYL/ISL | 0.000 | 0.380 | 0.004 | 0.237 | 0.000 |
| Sticta fuliginosa (Dicks.) Ach. | CYL/ISL | 0.000 | 0.361 | 0.000 | 0.609 | 0.007 |
| Sticta gracilis (Müll. Arg.) A. Zahlbruckner | CYL/SEL | 0.000 | 0.026 | 0.009 | 0.200 | 0.205 |
| Sticta nylanderiana A. Zahlbruckner | BFL/SEL | 0.000 | 1.712 | 0.000 | 0.000 | 0.000 |
| Sulcaria sulcata (Lév.) Bystrek ex Brodo & D. Hawksw. | FRL/SEL | 0.000 | 0.027 | 0.706 | 3.920 | 0.000 |
| Usnea baileyi (Stirt.) A. Zahlbruckner | FRL/SOL | 0.000 | <0.001 | 0.000 | 0.000 | 0.028 |
| Usnea florida (L.) Weber ex F. H. Wigg. | FRL/SEL | 0.000 | 12.237 | 56.740 | 18.906 | 3.588 |
| Usnea nidifica Taylor | FRL/SOL | 0.000 | 0.497 | 0.000 | 0.001 | 0.000 |
| Usnea rubicunda Stirt. | FRL/SOL | 0.000 | 0.000 | 0.000 | 0.082 | 0.000 |
| Usnea sp. | FRL/SEL | 0.000 | 0.007 | 1.889 | 0.014 | 0.000 |
| Lichen Biomass | R2GLMM(m) | R2GLMM(c) | RΣ2 | Partial R2height | Partial R2dbh | Partial R2height:dbh | |
|---|---|---|---|---|---|---|---|
| Total | All | 0.195 | 0.767 | 0.418 | 0.122 * | 0.073 | 0.047 |
| Branch | 0.183 | 0.745 | 0.389 | 0.096 * | 0.076 * | 0.048 | |
| Trunk | 0.104 | 0.659 | 0.251 | 0.126 * | 0.048 | 0.049 * | |
| Broadly-lobed foliose lichens | All | 0.283 | 0.663 | 0.446 | 0.095 * | 0.121 ** | 0.077 * |
| Branch | 0.294 | 0.609 | 0.437 | 0.081 | 0.142 * | 0.093 * | |
| Trunk | 0.158 | 0.500 | 0.220 | 0.096 | 0.083 * | 0.082 ** | |
| Cyanolichens | All | 0.170 | 0.360 | 0.345 | 0.164 *** | 0.097 *** | 0.093 *** |
| Branch | 0.370 | 0.560 | 0.558 | 0.332 *** | 0.269 *** | 0.272 *** | |
| Trunk | 0.150 | 0.329 | 0.265 | 0.111 *** | 0.019 * | 0.016 * | |
| Fruticose lichens | All | 0.081 | 0.727 | 0.197 | 0.080 | 0.025 | 0.022 |
| Branch | 0.091 | 0.725 | 0.213 | 0.089 | 0.024 | 0.022 | |
| Trunk | 0.004 | 0.269 | 0.008 | 0.000 | 0.008 | 0.008 | |
| Narrowly-lobed foliose lichens | All | 0.186 | 0.734 | 0.375 | 0.088 * | 0.017 | 0.005 |
| Branch | 0.190 | 0.722 | 0.375 | 0.084 * | 0.021 | 0.007 | |
| Trunk | 0.010 | 0.639 | 0.041 | 0.019 | 0.022 | 0.025 | |
| Isidiate lichens | All | 0.018 | 0.912 | 0.013 | 0.005 *** | 0.003 *** | 0.003 *** |
| Branch | 0.231 | 0.456 | 0.301 | 0.071 * | 0.049 | 0.031 | |
| Trunk | 0.167 | 0.310 | 0.297 | 0.138 *** | 0.013 | 0.012 | |
| Sexual lichens | All | 0.200 | 0.788 | 0.437 | 0.146 * | 0.032 | 0.017 |
| Branch | 0.198 | 0.789 | 0.432 | 0.123 * | 0.026 | 0.010 | |
| Trunk | 0.017 | 0.345 | 0.048 | 0.010 | 0.018 | 0.015 | |
| Sorediate lichens | All | 0.162 | 0.579 | 0.276 | 0.020 | 0.076 * | 0.042 |
| Branch | 0.153 | 0.521 | 0.252 | 0.009 *** | 0.076 ** | 0.040 *** | |
| Trunk | 0.007 | 0.664 | 0.122 | 0.003 | 0.118 *** | 0.105 *** | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Liu, S.; Shi, X.-M.; Liu, W.-Y.; Song, L.; Lu, H.-Z.; Chen, X.; Wu, C.-S. Forest Type and Tree Characteristics Determine the Vertical Distribution of Epiphytic Lichen Biomass in Subtropical Forests. Forests 2017, 8, 436. https://doi.org/10.3390/f8110436
Li S, Liu S, Shi X-M, Liu W-Y, Song L, Lu H-Z, Chen X, Wu C-S. Forest Type and Tree Characteristics Determine the Vertical Distribution of Epiphytic Lichen Biomass in Subtropical Forests. Forests. 2017; 8(11):436. https://doi.org/10.3390/f8110436
Chicago/Turabian StyleLi, Su, Shuai Liu, Xian-Meng Shi, Wen-Yao Liu, Liang Song, Hua-Zheng Lu, Xi Chen, and Chuan-Sheng Wu. 2017. "Forest Type and Tree Characteristics Determine the Vertical Distribution of Epiphytic Lichen Biomass in Subtropical Forests" Forests 8, no. 11: 436. https://doi.org/10.3390/f8110436
APA StyleLi, S., Liu, S., Shi, X.-M., Liu, W.-Y., Song, L., Lu, H.-Z., Chen, X., & Wu, C.-S. (2017). Forest Type and Tree Characteristics Determine the Vertical Distribution of Epiphytic Lichen Biomass in Subtropical Forests. Forests, 8(11), 436. https://doi.org/10.3390/f8110436