
Plant Debris and Its Contribution to Ecosystem Carbon Storage in Successional Larix gmelinii Forests in Northeastern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Field Investigation
2.3. Statistical Analyses
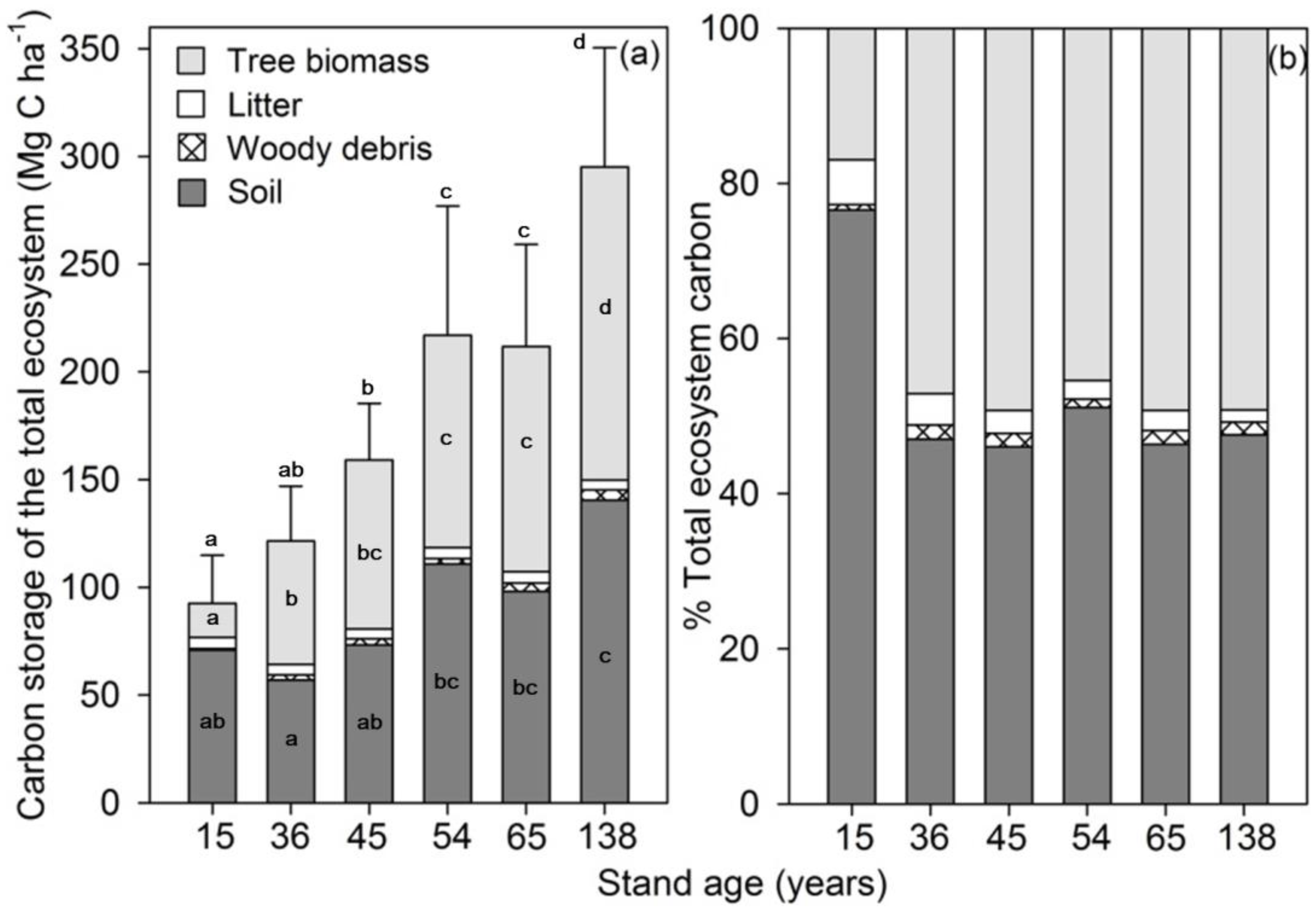
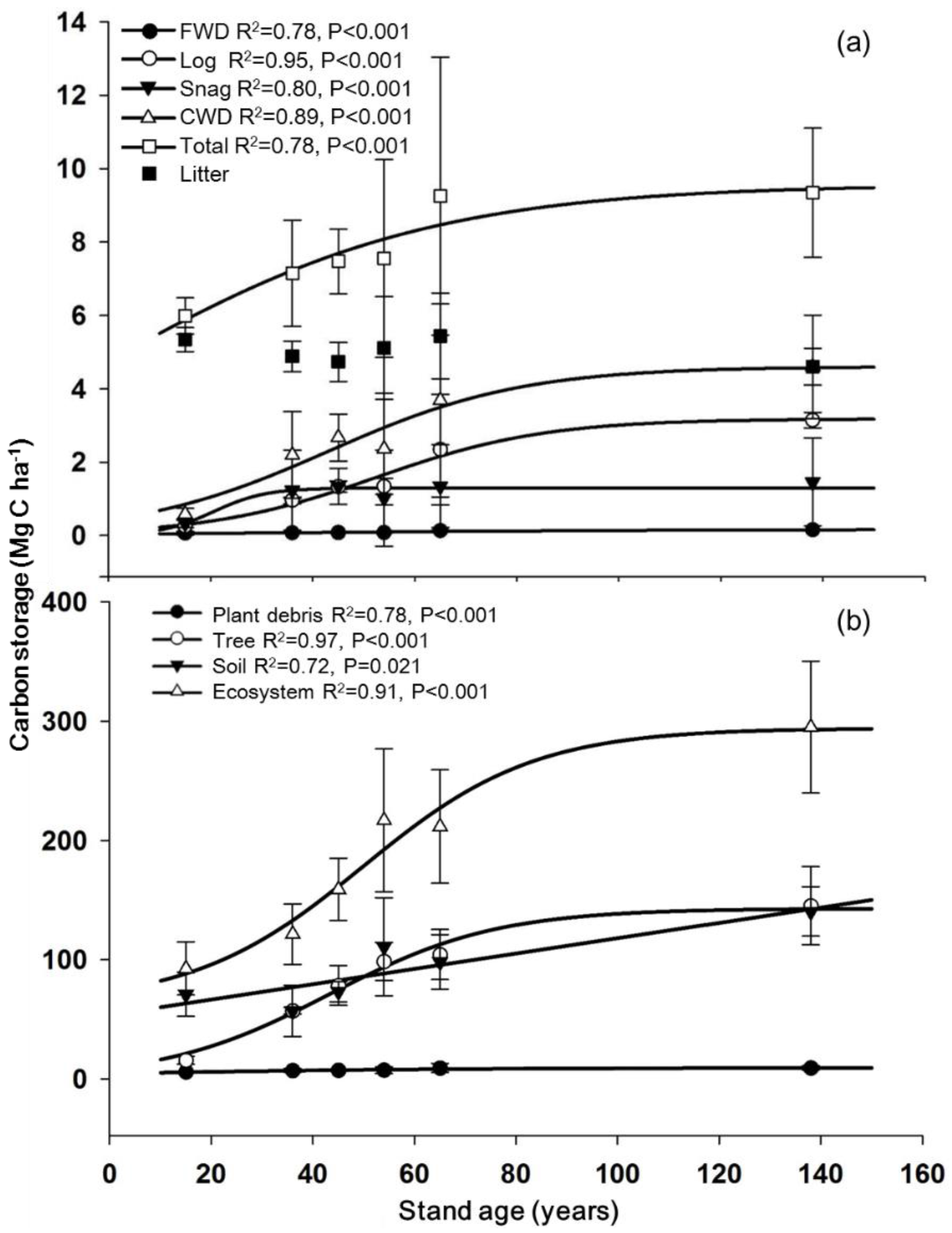
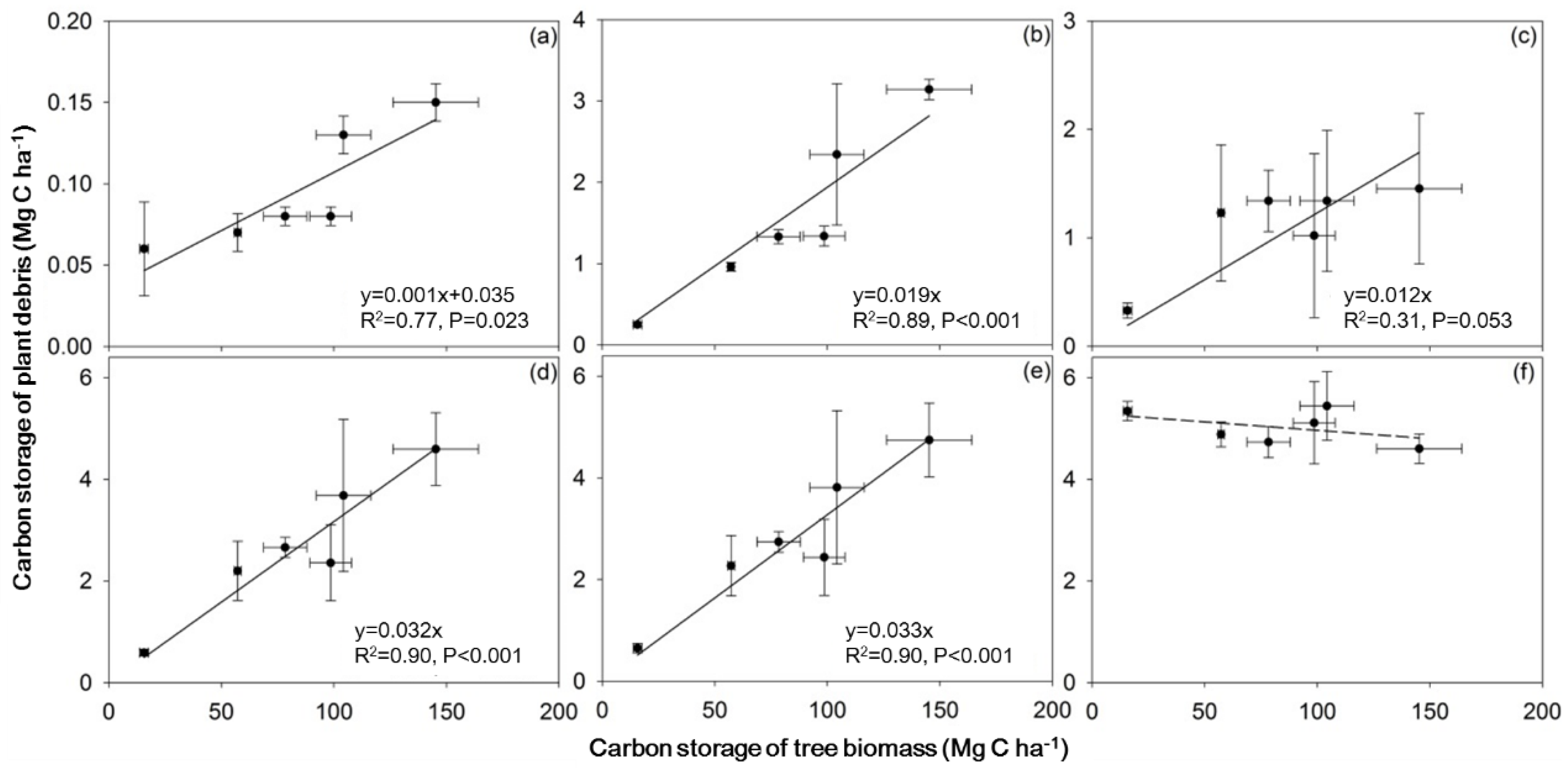
3. Results
3.1. Carbon Storage in Plant Debris
3.2. Carbon Storage in Ecosystem
3.3. Carbon Storage as a Function of Stand Age
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Berg, B.; McClaugherty, C. Decomposition as a process: Some main features. In Plant Litter; Springer: Berlin/Heidelberg, Germany, 2014; pp. 11–34. [Google Scholar]
- Hall, S.; Burke, I.; Hobbs, N. Litter and dead wood dynamics in ponderosa pine forests along a 160-year chronosequence. Ecol. Appl. 2006, 16, 2344–2355. [Google Scholar] [CrossRef]
- Triska, F.-J.; Cromack, K., Jr. The role of wood debris in forests and streams. In Forests: Fresh Perspectives from Ecosystem Analysis; Oregon State University Press: Corvallis, OR, USA, 1980; pp. 171–190. [Google Scholar]
- Brassard, B.-W.; Chen, H.-Y. Effects of forest type and disturbance on diversity of coarse woody debris in boreal forest. Ecosystems 2008, 11, 1078–1090. [Google Scholar] [CrossRef]
- Harmon, M.-E.; Franklin, J.-F.; Swanson, F.-J.; Sollins, P.; Gregory, S.-V.; Lattin, J.-D.; Anderson, N.-H.; Cline, S.-P.; Aumen, N.-G.; Sedell, J.-R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Swift, M.-J.; Heal, O.-W.; Anderson, J.-M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Tallmon, D.; Mills, L.-S. Use of logs within home ranges of California red-backed voles on a remnant of forest. J. Mammal. 1994, 75, 97–101. [Google Scholar] [CrossRef]
- Harmon, M.-E.; Hua, C. Coarse woody debris dynamics in two old-growth ecosystems. Bioscience 1991, 41, 604–610. [Google Scholar] [CrossRef]
- Hudiburg, T.; Law, B.; Turner, D.-P.; Campbell, J.; Donato, D.; Duane, M. Carbon dynamics of Oregon and Northern California forests and potential land-based carbon storage. Ecol. Appl. 2009, 19, 163–180. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Zhang, D.; Hui, D.; Luo, Y.; Zhou, G. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef]
- Lang, G.-E. Forest turnover and the dynamics of bole wood litter in subalpine balsam fir forest. Can. J. For. Res. 1985, 15, 262–268. [Google Scholar] [CrossRef]
- Tritton, L.-M. Dead Wood in the Northern Hardwood Forest Ecosystem; Yale University: New Haven, CT, USA, 1980. [Google Scholar]
- Spies, T.-A.; Franklin, J.-F.; Thomas, T.-B. Coarse woody debris in Douglas-fir forests of western Oregon and Washington. Ecology 1988, 69, 1689–1702. [Google Scholar] [CrossRef]
- Sturtevant, B.-R.; Bissonette, J.-A.; Long, J.-N.; Roberts, D.-W. Coarse woody debris as a function of age, stand structure, and disturbance in boreal Newfoundland. Ecol. Appl. 1997, 7, 702–712. [Google Scholar] [CrossRef]
- Carmona, M.-R.; Armesto, J.-J.; Aravena, J.-C.; Pérez, C.-A. Coarse woody debris biomass in successional and primary temperate forests in Chiloé Island, Chile. For. Ecol. Manag. 2002, 164, 265–275. [Google Scholar] [CrossRef]
- Green, P.; Peterken, G.-F. Variation in the amount of dead wood in the woodlands of the Lower Wye Valley, UK in relation to the intensity of management. For. Ecol. Manag. 1997, 98, 229–238. [Google Scholar] [CrossRef]
- Noormets, A.; McNulty, S.-G.; Domec, J.-C.; Gavazzi, M.; Sun, G.; King, J.-S. The role of harvest residue in rotation cycle carbon balance in loblolly pine plantations. Respiration partitioning approach. Glob. Chang. Biol. 2012, 18, 3186–3201. [Google Scholar] [CrossRef]
- Zhang, C.; Ju, W.; Chen, J.-M.; Wang, X.; Yang, L.; Zheng, G. Disturbance-induced reduction of biomass carbon sinks of China’s forests in recent years. Environ. Res. Lett. 2015, 10, 114021. [Google Scholar] [CrossRef]
- Guo, Z.-D.; Hu, H.-F.; Li, P.; Li, N.-Y.; Fang, J.-Y. Spatio-temporal changes in biomass carbon sinks in China’s forests from 1977 to 2008. Sci. China Life Sci. 2013, 56, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, S.; Guo, Z.; Xu, B.; Fang, J. The stage-classified matrix models project a significant increase in biomass carbon stocks in China’s forests between 2005 and 2050. Sci. Rep. 2015, 5, 11203. [Google Scholar] [CrossRef] [PubMed]
- Binkley, D. A hypothesis about the interaction of tree dominance and stand production through stand development. For. Ecol. Manag. 2004, 190, 265–271. [Google Scholar] [CrossRef]
- Taylor, A.-R.; Wang, J.-R.; Chen, H.-Y.-H. Carbon storage in a chronosequence of red spruce (Picea rubens) forests in central Nova Scotia, Canada. Can. J. For. Res. 2007, 37, 2260–2269. [Google Scholar] [CrossRef]
- Noh, N.-J.; Son, Y.; Lee, S.-K.; Seo, K.-W.; Heo, S.-J.; Yi, M.-J.; Park, P.-S.; Kim, R.-H.; Son, Y.-M.; Lee, K.-H. Carbon and nitrogen storage in an age-sequence of Pinus densiflora stands in Korea. Sci. China Life Sci. 2010, 53, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Woodall, C.; Walters, B.; Oswalt, S.; Domke, G.; Toney, C.; Gray, A. Biomass and carbon attributes of downed woody materials in forests of the United States. For. Ecol. Manag. 2013, 305, 48–59. [Google Scholar] [CrossRef]
- Fang, J.-Y.; Guo, Z.-D.; Hu, H.-F.; Kato, T.; Muraoka, H.; Son, Y. Forest biomass carbon sinks in East Asia, with special reference to the relative contributions of forest expansion and forest growth. Glob. Chang. Biol. 2014, 20, 2019–2030. [Google Scholar] [CrossRef] [PubMed]
- Du, E.; Zhou, Z.; Li, P.; Hu, X.; Ma, Y.; Wang, W.; Zheng, C.; Zhu, J.; He, J.-S.; Fang, J. NEECF: A project of nutrient enrichment experiments in China’s forests. J. Plant Ecol. 2013, 6, 428–435. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.; Zhang, M.; Yan, Q.; Sun, O.-J. Soil microbial biomass carbon and nitrogen in forest ecosystems of Northeast China: A comparison between natural secondary forest and larch plantation. J. Plant Ecol. 2010, 3, 175–182. [Google Scholar] [CrossRef]
- Li, H.-K.; Lei, L.-C. Estimation and Evaluation of Forest Biomass Carbon Storage in China; China Forestry Publishing Press: Beijing, China, 2010. (In Chinese) [Google Scholar]
- Worbes, M.; Staschel, R.; Roloff, A.; Junk, W.-J. Tree ring analysis reveals age structure, dynamics and wood production of a natural forest stand in Cameroon. For. Ecol. Manag. 2003, 173, 105–123. [Google Scholar] [CrossRef]
- Bruelheide, H.; Schmid, B. Community assembly during secondary forest succession in a Chinese subtropical forest. Ecol. Monogr. 2011, 81, 25–41. [Google Scholar] [CrossRef]
- Hunt, R. Plant growth curves. In The Functional Approach to Plant Growth Analysis; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- Wang, C.; Bond-Lamberty, B.; Gower, S.-T. Carbon distribution of a well-and poorly-drained black spruce fire chronosequence. Glob. Chang. Biol. 2003, 9, 1066–1079. [Google Scholar] [CrossRef]
- Ryan, M.-G.; Binkley, D.; Fownes, J.-H. Age-related decline in forest productivity: pattern and process. Adv. Ecol. Res. 1997, 27, 213–262. [Google Scholar]
- Bond-Lamberty, B.; Wang, C.; Gower, S.-T. Net primary production and net ecosystem production of a boreal black spruce wildfire chronosequence. Glob. Chang. Biol. 2004, 10, 473–487. [Google Scholar] [CrossRef]
- Mukhortova, L.; Trefilova, O.; Krivobokov, L.; Klimchenko, A.; Vedrova, E. Coarse woody debris stock in forest ecosystems on latitudinal gradient of Central Siberia. In Proceedings of the 16th International Multidisciplinary Scientific GeoConferences SGEM2016, Albena Resort, Bulgaria, 28 June–6 July 2016. [Google Scholar]
- Mukhortova, L.-V. Carbon and nutrient release during decomposition of coarse woody debris in forest ecosystems of Central Siberia. Folia For. Pol. 2012, 54, 71–83. [Google Scholar]
- Pregitzer, K.-S.; Euskirchen, E.-S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Peichl, M.; Arain, M.-A. Above-and belowground ecosystem biomass and carbon pools in an age-sequence of temperate pine plantation forests. Agr. For. Meteorol. 2006, 140, 51–63. [Google Scholar] [CrossRef]
- Zhang, Q.-Z.; Wang, C.-K. Carbon density and distribution of six Chinese temperate forests. Sci. China Life Sci. 2010, 53, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-X.; Hu, X.-Y.; Yao, H.; Liu, G.-H.; Ji, C.-J.; Fang, J.-Y. A significant carbon sink in temperate forests in Beijing: Based on 20-year field measurements in three stands. Sci. China Life Sci. 2015, 58, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B; Wang, X; Fang, J; Piao, S.; Shen, H.; Zhao, S.; Peng, C. Altitudinal changes in carbon storage of temperate forests on Mt Changbai, northeast China. J. Plant Res. 2010, 123, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhu, J.; Wang, C.; Zheng, T.; Wu, Q.; Yao, H.; Fang, J. Impacts of fire severity and post-fire reforestation on carbon pools in boreal larch forests in Northeast China. J. Plant Ecol. 2016, 9, 1–9. [Google Scholar] [CrossRef]
- Harmon, M.-E.; Krankina, O.-N.; Yatskov, M.; Matthews, E. Predicting broad-scale carbon stores of woody detritus from plot-level data. In Assessment Methods for Soil Carbon; Lai, R., Kimble, J., Stewart, B.-A., Eds.; CRC Press: New York, NY, USA, 2001; pp. 533–552. [Google Scholar]
- Goodale, C.-L.; Apps, M.-J.; Birdsey, R.-A.; Field, C.-B.; Heath, L.-S.; Houghton, R.-A.; Jenkins, J.-C.; Kohlmaier, G.-H.; Kurz, W.; Liu, S.; et al. Forest carbon sinks in the Northern Hemisphere. Ecol. Appl. 2002, 12, 891–899. [Google Scholar] [CrossRef]
- Martin, J.-L.; Gower, S.-T.; Plaut, J.; Holmes, B. Carbon pools in a boreal mixedwood logging chronosequence. Glob. Chang. Biol. 2005, 11, 1883–1894. [Google Scholar] [CrossRef]
- Pan, Y.-D.; Birdsey, R.-A.; Fang, J.-Y.; Houghton, R.; Kauppi, P.-E.; Kurz, W.-A.; Phillips, O.-L.; Shvidenko, A.; Lewis, S.-L.; Canadell, J.-G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Amanzadeh, B.; Sagheb-Talebi, K.; Foumani, B.-S.; Fadaie, F.; Camarero, J.-J.; Linares, J.-C. Spatial distribution and volume of dead wood in unmanaged Caspian beech (Fagus orientalis) forests from northern Iran. Forests 2013, 4, 751–765. [Google Scholar] [CrossRef]
- Nalder, I.-A.; Wein, R.-W. Long-term forest floor carbon dynamics after fire in upland boreal forests of western Canada. Glob. Biogeochem. Cycle 1999, 13, 951–968. [Google Scholar] [CrossRef]
- Domke, G.-M.; Woodall, C.-W.; Walters, B.-F.; Smith, J.-E. From models to measurements: comparing downed dead wood carbon stock estimates in the US forest inventory. PLoS ONE 2013, 8, e59949. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-H.; Sun, M.-L. Caloric values and carbon contents of dominant trees in Xiaoxing’anling forest region. Acta Ecol. Sin. 2009, 29, 953–959, (In Chinese with English abstract). [Google Scholar]
- Zhang, Q.; Wang, C.; Wang, X.; Quan, X. Carbon concentration variability of 10 Chinese temperate tree species. For. Ecol. Manag. 2009, 258, 722–727. [Google Scholar] [CrossRef]
- Yao, F.; Chen, Y.; Yan, Z.; Li, P.; Han, W.; Fang, J. Biogeographic patterns of structural traits and C:N:P stoichiometry of tree twigs in China’s forests. PLoS ONE 2015, 10, e0116391. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand (years) | 15 | 36 | 45 | 54 | 65 | 138 |
|---|---|---|---|---|---|---|
| Location | 48.03° N | 48.08° N | 48.10° N | 48.05° N | 48.06° N | 48.08° N |
| 129.26° E | 129.26° E | 129.24° E | 129.17° E | 129.09° E | 129.07° E | |
| Altitude (m) | 370 | 368 | 288 | 329 | 279 | 345 |
| Slope (°, aspect) | 9.6 N | 13.8 SW | 1.2 NW | 1.1 SE | 1.6 S | 1.9 NW |
| Mean DBH (cm) | 7.9 | 19.6 | 21.9 | 14.6 | 15.7 | 15.7 |
| DBHMax (cm) | 10.5 | 28.3 | 30.9 | 47.2 | 53.7 | 55.7 |
| Mean Height (m) | 8.0 | 12.7 | 13.4 | 10.7 | 10.3 | 12.6 |
| HeightMax (m) | 9.3 | 15.4 | 16.1 | 20.6 | 25.0 | 29.6 |
| TBA (m2/ha) | 14.6 | 31.7 | 44.5 | 48.1 | 48.6 | 63.1 |
| Stand Density (stems/ha) | 2192 | 1075 | 1058 | 900 | 758 | 900 |
| Forest Origin | Plantation | Plantation | Plantation | Secondary | Secondary | Primary |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Zhou, X.; Fang, W.; Xiong, X.; Zhu, B.; Ji, C.; Fang, J. Plant Debris and Its Contribution to Ecosystem Carbon Storage in Successional Larix gmelinii Forests in Northeastern China. Forests 2017, 8, 191. https://doi.org/10.3390/f8060191
Zhu J, Zhou X, Fang W, Xiong X, Zhu B, Ji C, Fang J. Plant Debris and Its Contribution to Ecosystem Carbon Storage in Successional Larix gmelinii Forests in Northeastern China. Forests. 2017; 8(6):191. https://doi.org/10.3390/f8060191
Chicago/Turabian StyleZhu, Jianxiao, Xuli Zhou, Wenjing Fang, Xinyu Xiong, Biao Zhu, Chengjun Ji, and Jingyun Fang. 2017. "Plant Debris and Its Contribution to Ecosystem Carbon Storage in Successional Larix gmelinii Forests in Northeastern China" Forests 8, no. 6: 191. https://doi.org/10.3390/f8060191