
Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains

USDA Forest Service, International Institute of Tropical Forestry, Jardín Botánico Sur, 1201 Calle Ceiba, Río Piedras, PR 00926, USA
Forests 2017, 8(6), 204; https://doi.org/10.3390/f8060204
Submission received: 3 April 2017
/
Revised: 5 June 2017
/
Accepted: 5 June 2017
/
Published: 10 June 2017
(This article belongs to the Special Issue Tropical Forest Ecology and Management for the Anthropocene)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The science of ecology fundamentally aims to understand species and their relation to the environment. At sites where hurricane disturbance is part of the environmental context, permanent forest plots are critical to understand ecological vegetation dynamics through time. An overview of forest structure and species composition from two of the longest continuously measured tropical forest plots is presented. Long-term measurements, 72 years at the leeward site, and 25 years at windward site, of stem density are similar to initial and pre-hurricane values at both sites. For 10 years post-hurricane Hugo (1989), stem density increased at both sites. Following that increase period, stem density has remained at 1400 to 1600 stems/ha in the leeward site, and at 1200 stems/ha in the windward site. The forests had similar basal area values before hurricane Hugo in 1989, but these sites are following different patterns of basal area accumulation. The leeward forest site continues to accumulate and increase basal area with each successive measurement, currently above 50 m2/ha. The windward forest site maintains its basal area values close to an asymptote of 35 m2/ha. Currently, the most abundant species at both sites is the sierra palm. Ordinations to explore variation in tree species composition through time present the leeward site with a trajectory of directional change, while at the windward site, the composition of species seems to be converging to pre-hurricane conditions. The observed differences in forest structure and composition from sites differently affected by hurricane disturbance provide insight into how particular forest characteristics respond at shorter or longer time scales in relation to previous site conditions and intensity of disturbance effects.
1. Introduction
Long-term forest monitoring provides vegetation succession measurements needed to understand resiliency and recovery of forest systems from disturbance events. Two frequently evaluated components of forest succession in response to disturbance are stand level structural properties and the dynamics of species composition through time [1]. Understanding forest structure and composition during vegetation succession in tropical forests may mean following a high number of species, each with their own particular life-history traits [2,3]. The environment of the Caribbean, with its natural disturbance regime and predicted climate change scenarios serves as a template to understand forest dynamics responses to disturbance events such as hurricanes [4,5,6,7,8,9].
Our understanding of forest dynamics has benefited immensely from the ability to observe changes through time, thus chronicling disturbance events and vegetation responses in an adequate and increasing time span [10,11,12]. This short communication aims to present an overview of succession dynamics in two continuously measured sites in the same forest type [12,13], located in the Luquillo Experimental Forest (LEF), Puerto Rico. Because of differences in location relative to prevailing winds, hurricane disturbance effects and the measured responses at these two sites were different [14], but these observations have not been analyzed together to understand overall response patterns at the level of the forest type. A comparison of responses in forest composition and structure during hurricane-induced succession in mature secondary forests is presented. This includes contrasting patterns of stem density, stem structure, and basal area responses at each site, and changes in abundance of dominant species and those with contrasting life histories through time. Brief notes on some of the species that have been lost and gained to the plots throughout the measurement period are also presented. An exploratory ordination analysis is used to highlight the differences in trajectories of species composition dynamics through time and in response to hurricane disturbance at each forest site. The role of hurricane disturbance at these sites is discussed in relation to observed and predicted forest structure.
2. Methods
2.1. Study Site
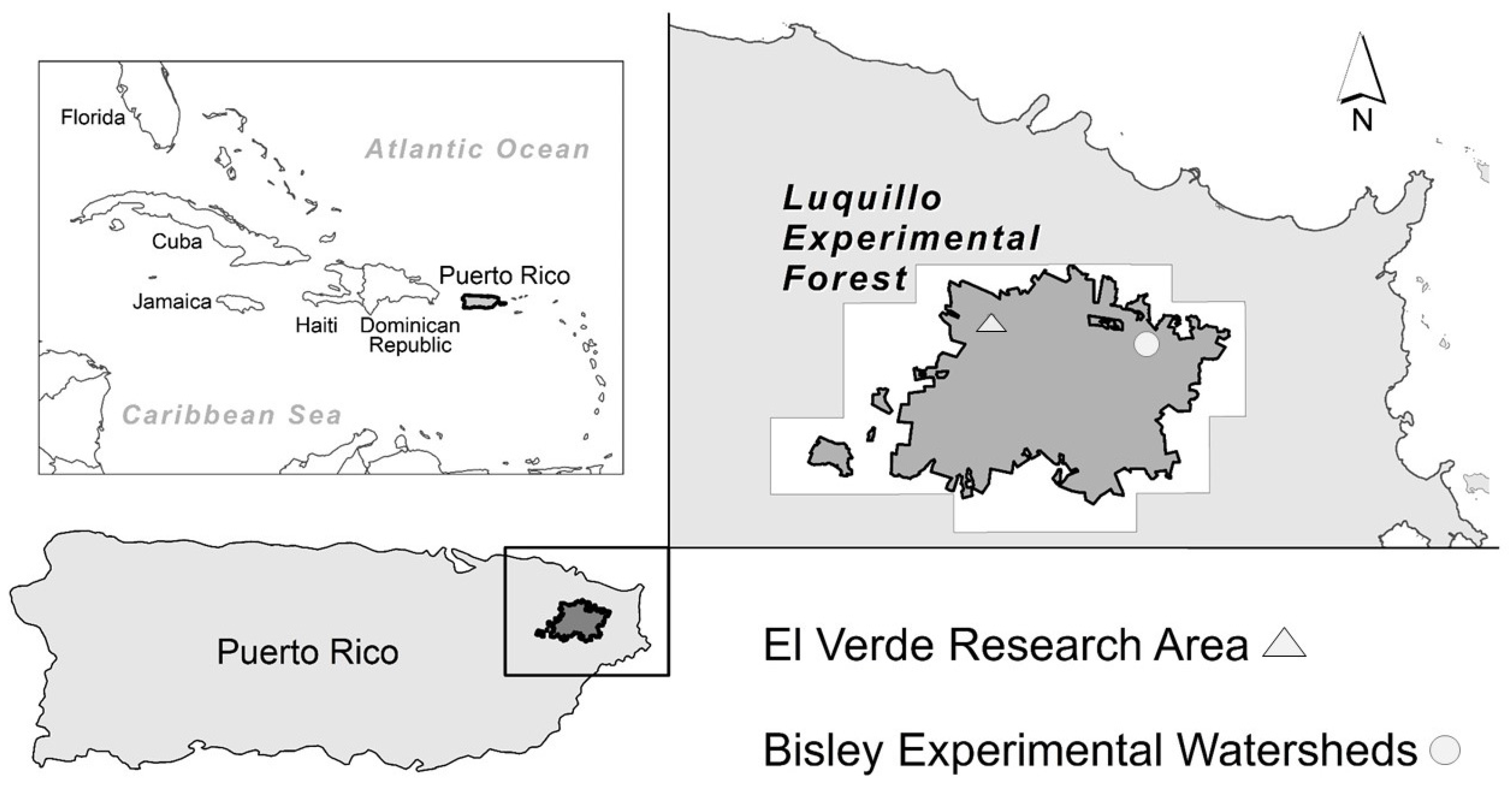
The LEF is located in northeastern Puerto Rico, with the El Verde Research Area (El Verde) on the west and the Bisley Experimental Watersheds (Bisley) on the east (Figure 1). The forests of the LEF grow in the context of environmental events such as hurricanes, volcanoes, Saharan-dust, and earthquakes [15,16,17]. The forest vegetation communities in the LEF are associated with different intensities and scales of disturbances such as treefalls, landslides, and hurricanes [18,19,20,21]. The lower montane forest contains the Dacryodes-Sloanea forest association, commonly known as tabonuco forest. This forest type occurs in the subtropical wet forest life zone, sensu Holdridge, at 200 to 600 m elevation with an average 3482 mm/yr of rainfall [17,22,23]. This forest type occupies the greatest extent of area in the LEF and is also found throughout Caribbean islands [13,14,24]. This forest type has trees with canopies that range from 25 to 30 m in height and lianas (i.e., woody vines) that form a common structural element in these forest communities (Figure 2a).
Trees in this forest type are distributed in relation to terrain geomorphology, with the dominant tree species Dacryodes excelsa Vahl (tabonuco) occupying ridges and ridge tops (Figure 2a) [25,26]. The sierra palm, Prestoea montana (synonymous with P. acuminata, var. montana) (R. Graham) Nichols, tends to occupy riparian valleys, concave areas and slopes, and can form mono-specific forest stands (Figure 2b) [27]. Other canopy level trees include Sloanea berteriana Choisy (motillo), Manilkara bidentata (A. DC.) Chev. (ausubo), Guarea guidonia L. Sleumer (guaraguao), Buchenavia capitata (Aubl.) Howard (granadillo), and Ocotea leucoxylon (Sw.) De Laness. (laurel geo) [23]. Younger tabonuco forest stands often include the tree species Tabebuia heterophylla (DC.) Britton, (roble blanco) Cecropia schreberiana Miq. (yagrumo), and Schefflera morototoni (Aubl.) Maguire, Steyerm. & Frodin (yagrumo hembra). In the past, some lower elevation tabonuco forests were selectively harvested for trees with commercially valuable wood from the species that are characteristically dominant in this forest type such as D. excelsa, M. bidentata, S. berteriana, and Magnolia splendens Urban (laurel sabino) [10,28,29,30]. The LEF has had a suite of climatic disturbance events that have been described, including hurricanes San Nicolás in 1931, San Ciprián in 1932, Santa Clara (Betsy) in 1956, Hugo in 1989, and Georges in 1998 [10,12,13]. Hurricane Hugo, the largest storm to affect the area since 1932, passed over the LEF on 18 September 1989. The hurricane defoliated entire areas, and although it reduced the aboveground biomass by 50 percent in windward Bisley [31], there was minimal biomass reduction in leeward El Verde [12]. After hurricane Hugo, eight other storms passed near the LEF [18,21]. Of those, hurricane Georges in 1998 was the largest and resulted in localized defoliation and uprooting [32]. At least six meteorological droughts of varying intensity have also been recorded during a recent 25-year study period [17,33,34].
2.1.1. Permanent Plots
The El Verde 3 plot (El Verde, 18°19’ N, 65°49’ W) is a 0.72 hectare plot that was established in 1943 by Frank Wadsworth to assess stem growth of trees of commercially valuable species [10,12]. In 1937 and in 1958, a timber-stand treatment was conducted, cutting and removing stems. The plot site has a leeward, northwest aspect and is situated on a ridge on the south side of the Quebrada Sonadora. The El Verde plot is ~0.5 km from the 16 ha Center for Tropical Forest Science (CTFS) Luquillo Forest Dynamics Plot, in what is now the El Verde Research Management Area. The El Verde Plot was measured upon establishment and then at intervals between 3 to 12 years, with recent measurement intervals set at 5 years (measurements in 1943, 1946, 1951, 1976, 1988, 1993, 1998, 2005, 2010, and 2015). In censuses of El Verde, all stems of all tree species ≥4.0 cm diameter at 1.3 m from the ground (dbh) were measured. Palms were measured for dbh and were included in the census when their internode reached 1.37 m. As new trees and palms reached the minimum diameter class, they were numbered and marked with aluminum tags. A detailed narrative of each El Verde plot assessment can be found in [12]. The Bisley Experimental Watersheds (Bisley; 18°20’ N, 65°50’ W) study area spans 13 ha of 3 monitored watersheds, tributaries to the Río Mameyes, established in 1987 as a research area with gaged streams, canopy towers for meteorological data, and sampling of forest vegetation. At this site, selective logging is presumed to have minimally occurred [29,30]. In 1989, a series of 78.54 m2 permanent forest plots, with a total cumulative sampled area of 0.71 ha, were established in Bisley [13,30,31]. These plots are measured at 5 year intervals (measurements in pre-hurricane 1989, post-hurricane 1989, 1994, 1999, 2004, 2009, and 2014). In censuses of Bisley, all stems ≥2.5 cm diameter at 1.3 m from the ground (dbh) were measured, but for comparative purposes in this communication only stems ≥4 cm dbh are included. Palms were measured for dbh and were included in the census when their internode reached 1.5 m. As new trees and palms reached the minimum diameter class, they were numbered and marked with aluminum tags.
2.1.2. Data Analyses
Data spanning 1943 to 2015 (10 censuses) for El Verde and 1989 to 2014 (7 censuses) for Bisley were used for analyses. Previous studies with data from these sites were limited to 5 censuses from Bisley and 8 from El Verde [12,13]. Analyses include data from both permanent plot sites with all stems ≥4 cm in diameter. Data on stem density and basal area are presented per hectare for all identified and living stems per census. To illustrate individual species stem density through time, six species that were dominant in terms of basal area and or stem density in the plots were selected. These species represent primary forest species (late successional) and secondary forest species (early successional), as classified by studies that combine the species-specific characteristics of seedling regeneration under different light conditions, and relative densities of various life history stages in this forest type [34,35]. The early successional species are C. schreberiana, and Psychotria berteriana DC. (cachimbo). The late successional species are D. excelsa, M. bidentata, and S. berteriana. The sierra palm, P. montana, is classified as a mature forest species although it has previously also been associated with early succession due to its ability to form slope and floodplain stands [10,14]. These six species have also been used in other studies of this forest type to explore responses to experimental hurricane disturbance [36] or to model simulations [37]. Examples of species that have been lost and gained to the plot sites throughout the measurement period are presented. To assess tree community species composition dynamics through time, two non-metric multidimensional scaling (NMS) ordination analyses were conducted with number of stems per species per census year. An NMS ordination was conducted because of its efficiency when reducing high dimensional multivariate species space (high number of species) to two dimensions, which provides ease when plotting simple 2 axis graphs. Another benefit is that NMS has minimal assumptions about relationships among variables [38,39]. The multivariate analyses contained only species that occurred in at least two censuses per study site. The NMS ordination was made using species abundance, with which a matrix was generated for each site using Bray–Curtis distance in PC Ord-6 (PC-ORD, Gleneden Beach, OR, USA; [40]). For El Verde, ordination data contained 75 species from 10 census years, and for Bisley, 62 species from 7 census years. Differences in community composition among census year (i.e., census year as the grouping variable) were also compared via simple standardized chi-square distances for species associations per census year on a matrix of species presence and absence. A value of −1 is a perfect negative association, or no similarity in composition. A value of 1 is a perfect positive association, or great similarity in composition. This is a complementary procedure to explore further comparisons of community species composition (presence and absence) among census year per site.
3. Results
3.1. Observations on Structural Characteristics
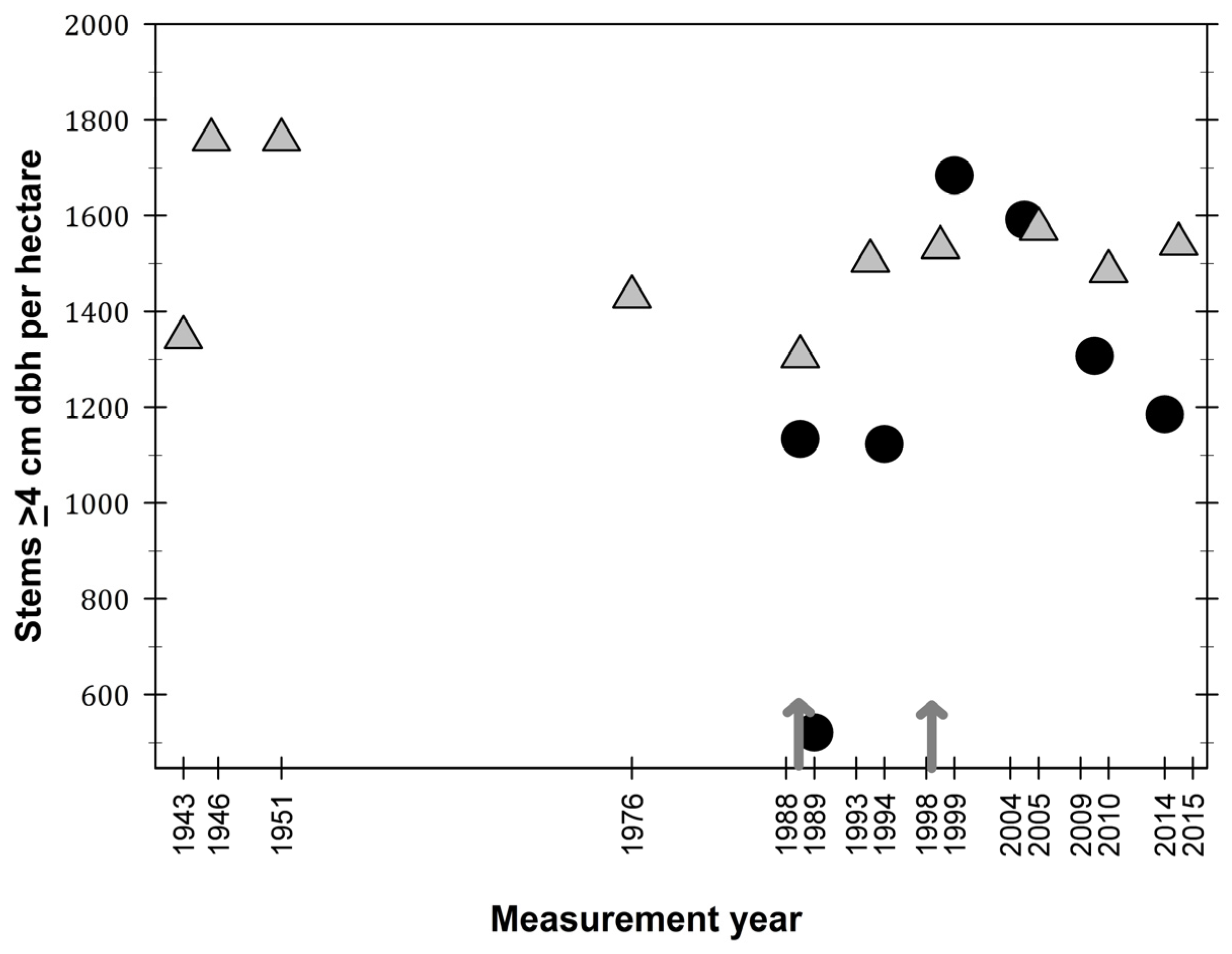
Stem density values in El Verde were between 1400 and 1800 per hectare during the first eight years of plot censuses (Figure 3). From 1976 to 2015, stem values have been constrained from 1400 to 1600 stems per hectare. Although stem density seems to have fluctuated during the 72-year study period at El Verde, these fluctuations have been minimal compared to dynamics observed at Bisley. Stem density values at Bisley were 1200 stems per hectare before hurricane Hugo, and these stem density values returned to that level five years after the hurricane. Maximum stem density values at Bisley were observed ten years after the hurricane, in 1999. The current trend in Bisley stem density is for a steady decrease in values, becoming closer to 1200 stems per hectare as observed in the last three censuses.
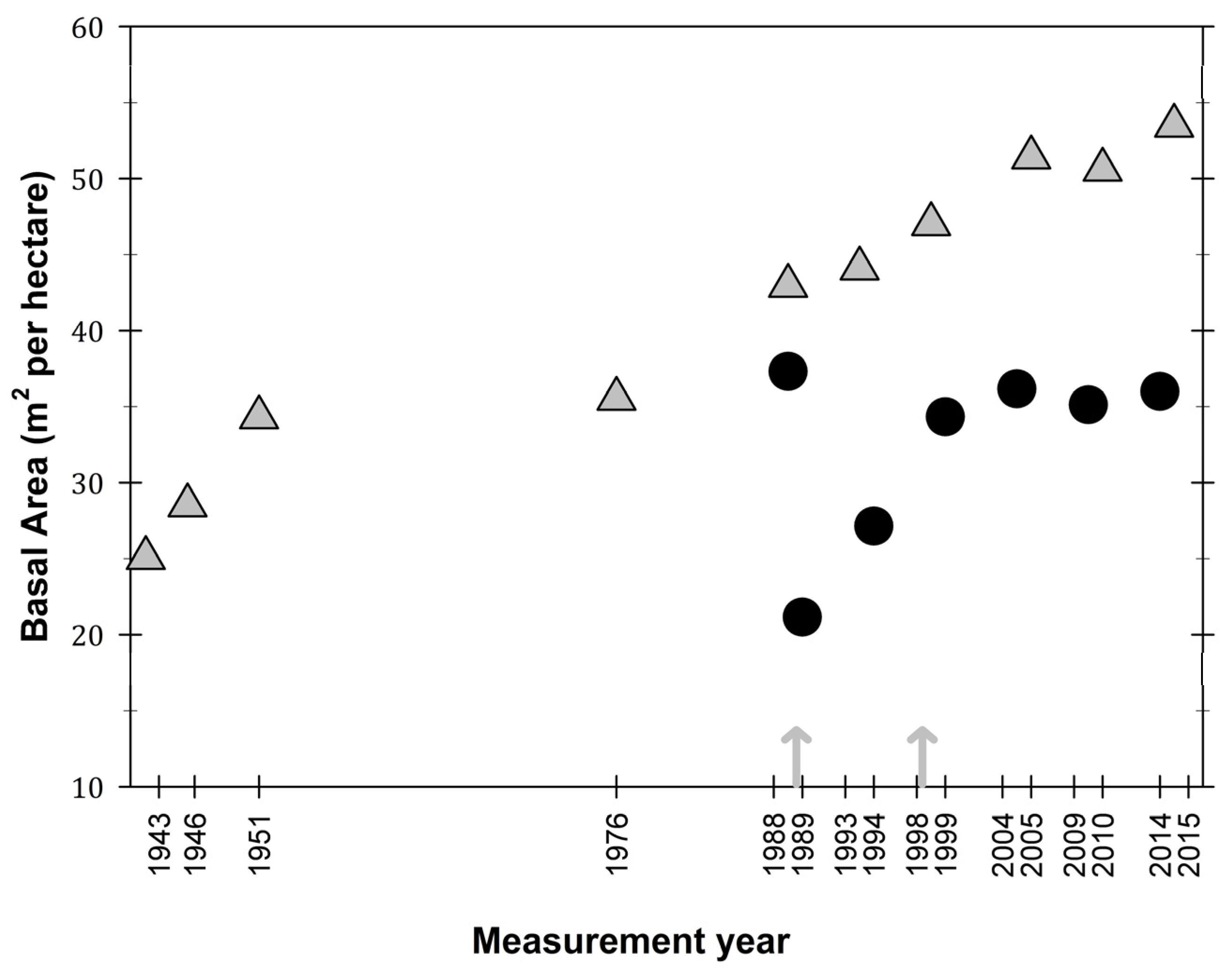
In terms of basal area, El Verde has an increasing trend with values from the last three plot censuses 2005, 2010, and 2015 almost doubled from initial plot measurements in 1943 and 1951 (Figure 4). Basal area at Bisley has maintained values between 35 and 40 m2 per hectare, which is similar to the basal area recorded at the site before hurricane Hugo (Figure 4). Although basal area decreased almost by half after hurricane Hugo, ten years later, basal area had consistent values at Bisley. Both El Verde and Bisley were at similar values of basal area (35 m2/ha) before hurricane Hugo in 1989.
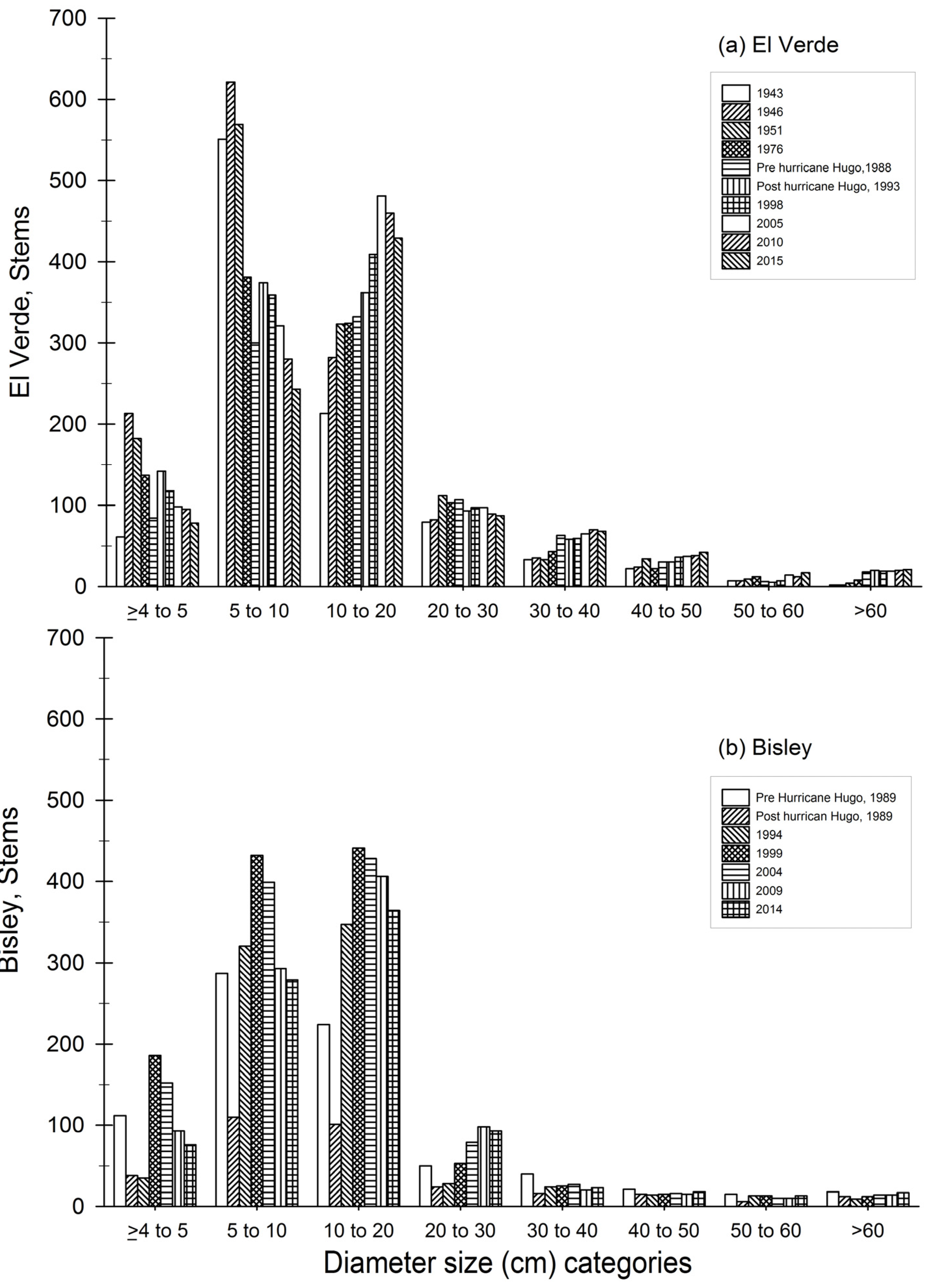
Distribution of stem diameter size classes through time reveal that at El Verde, the stem category of 10 to 20 cm consistently increased in plot censuses from 1943 to 2005 and has contained the greatest number of stems since 1998 (Figure 5). Stems in the smaller size category of 5 to 10 cm accounted for the greatest proportion of the plot in the early 1943, 1946 and 1951 censuses, but this size class decreased consistently since 1993. The categories of 30 to 40 cm and of 40 to 50 cm have both maintained an increasing trend at El Verde. Since plot establishment, El Verde stems in the >60 cm category have gradually increased. In Bisley, however, trees >60 cm reached pre-hurricane Hugo values in 2015. The greatest proportion of stems in Bisley since 1994 originate from the 10 to 20 cm size category. Similarly, in El Verde and Bisley, the greatest number of stems is in the category of 10 to 20 cm; however, Bisley currently has a greater quantity of stems in that size category than El Verde. At the Bisley study site, the 30 to 40 cm and the 40 to 50 cm categories account for a smaller component of the stems in contrast to El Verde.
3.2. Observations on Species Abundances and Plot Species Composition
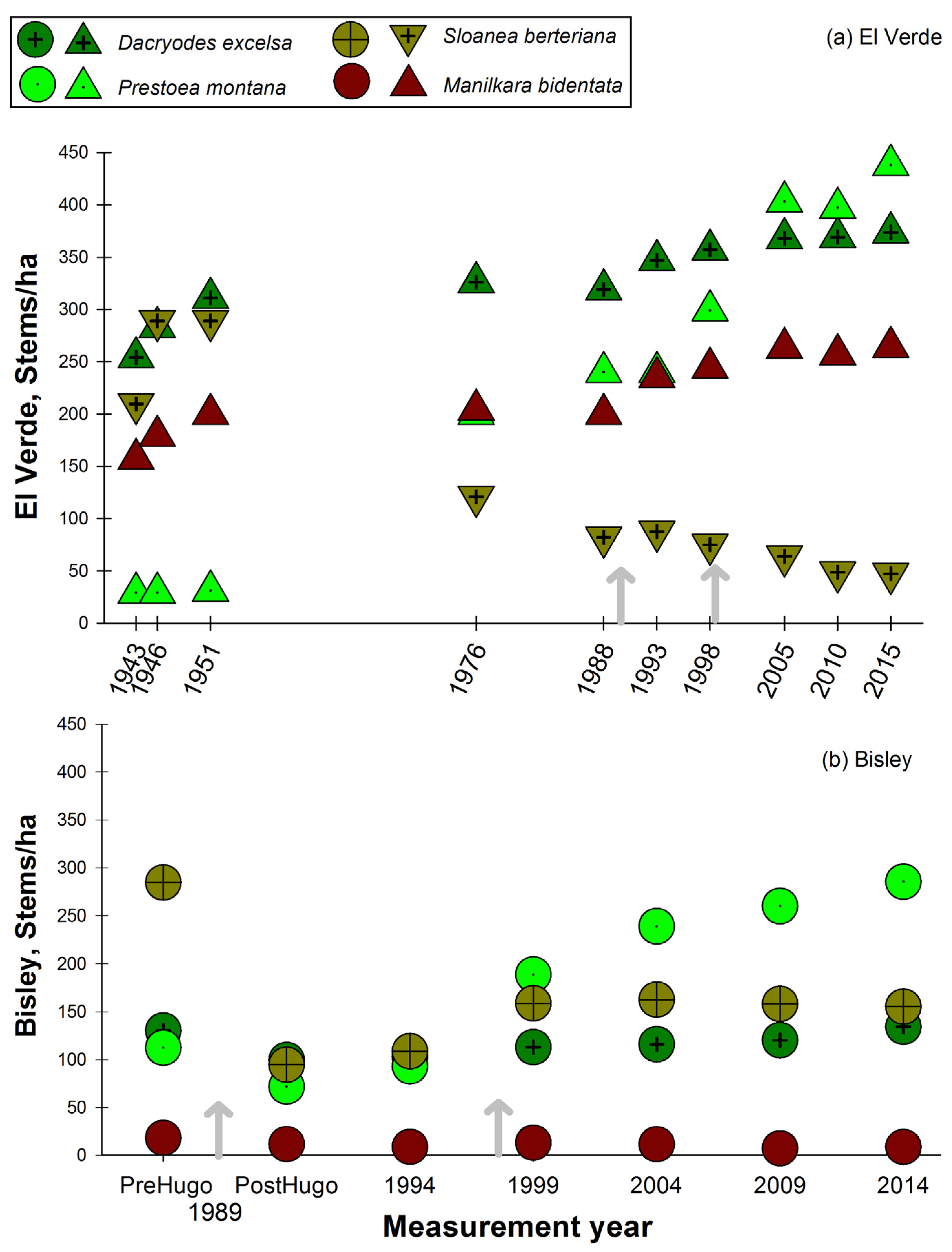
The long-lived and shade tolerant primary forest species, D. excelsa, had slightly increasing stem densities at both El Verde and Bisley (Figure 6a,b). This trend was observed even with the effect of hurricane Hugo decreasing basal area at Bisley. However, this was not the case with S. berteriana, which presented a slightly decreasing, perhaps stabilizing, trend in stem density at Bisley ten years after hurricane Hugo. At the El Verde site, S. berteriana presented a decrease in stem density after the 1951 census (Figure 6a). This decreasing trend started to stabilize in the 2005, 2010 censuses, but at much lower stem density values than in the initial 1943 census. The slow growing, long lived and dense hardwood M. bidentata in El Verde follows a very similar stem density pattern with a consistent and slight increase during the past 72 years as observed with D. excelsa. At Bisley, M. bidentata has remained at low stem density and has not changed during the study period (Figure 6b). In the case of M. bidentata, although the stem density remained constant at Bisley, it was one order of magnitude lower than at El Verde.
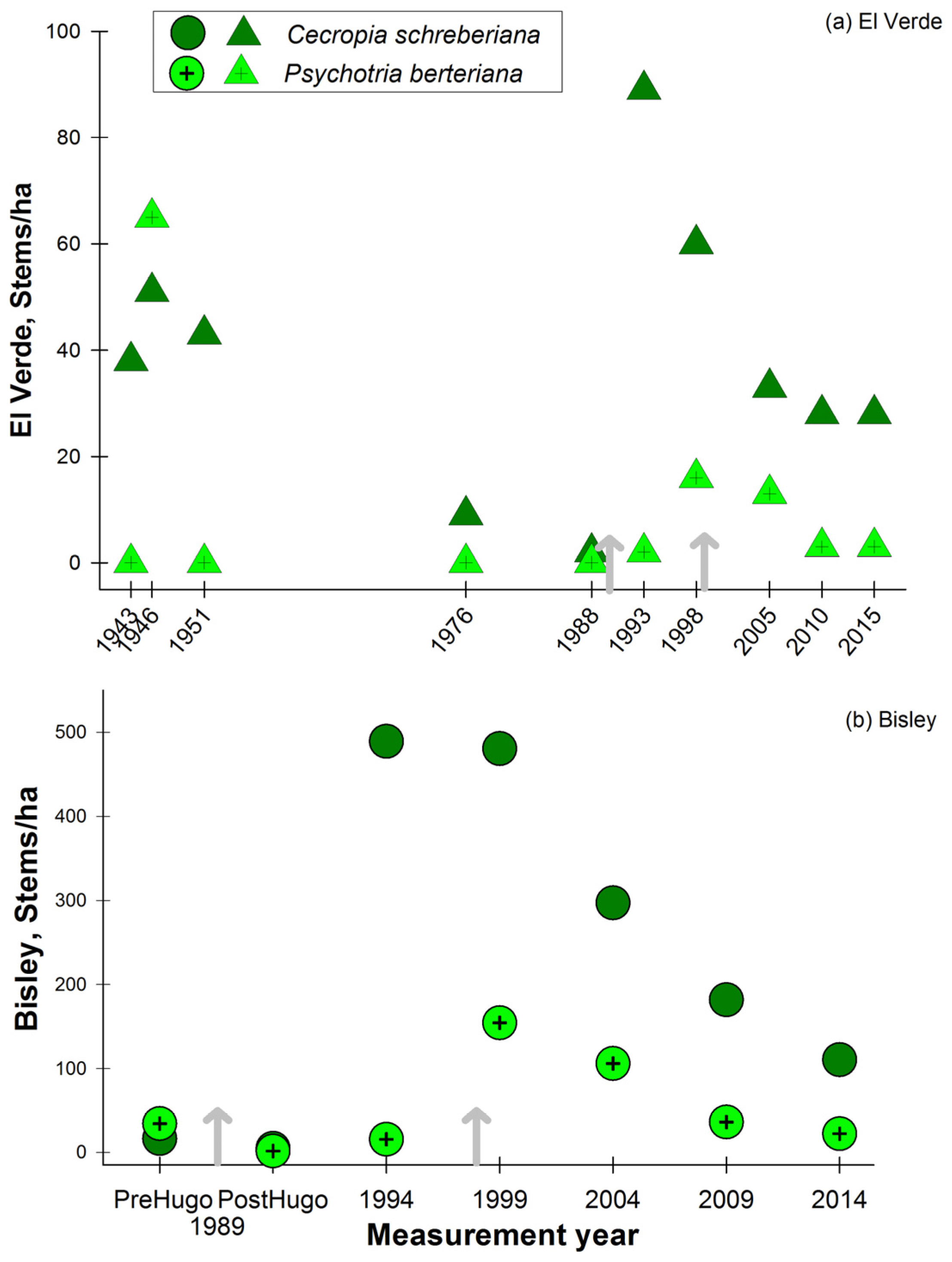
The sierra palm, P. montana, pattern of stem density has consistently increased through the study period at both Bisley and El Verde (Figure 6a,b). At El Verde, the lowest stem densities were observed in the initial censuses 1943, 1946 and 1951, and values have continued to increase since 1976 in all following censuses. At Bisley, P. montana decreased in the census after hurricane Hugo, but ten years later, values were higher than previous to hurricane Hugo (Figure 6b). The patterns that contrast the most are the stem density fluctuations observed in two of the most abundant species, P. montana and S. berteriana. The palm P. montana has increased steadily at both sites while S. berteriana has presented a consistently decreasing trend at El Verde and a relatively constant stem density 10 years post-hurricane Hugo at Bisley. At Bisley, stem densities of the shade intolerant C. schreberiana have not returned to the lower pre-hurricane Hugo densities, while at El Verde, the values are lower than the initial 1943 census (Figure 7a). In the case of P. berteriana, both Bisley and El Verde stem densities have returned to values similar to initial census conditions at each site (Figure 7a,b). Both of these species were in low abundance initially and had a dramatic increase after hurricane Hugo. Although stem densities for C. schreberiana are still higher than pre-hurricane Hugo values, these are much lower at El Verde (35–40 stem/ha) compared to Bisley (100–200 stems/ha). In the last 3 censuses at both study sites, there has been a consistently decreasing trend of early successional C. schreberiana and P. berteriana stem density (Figure 7a,b). Other changes in species observed include the tree fern Cyathea arborea, documented for the first time inside the El Verde plot in 2005, after 62 years. This is in contrast to Bisley plots where this species was present since before hurricane Hugo and has remained in constant stem abundance values. The opposite case is observed with the wind dispersed and high-light associated T. heterophylla, which has remained present in El Verde plots during the 72 years of censuses, while it disappeared from Bisley plots 5 years after hurricane Hugo. The species Magnolia splendens was rare in the initial plot measurements at Bisley and El Verde and is now absent from the plots at both sites.
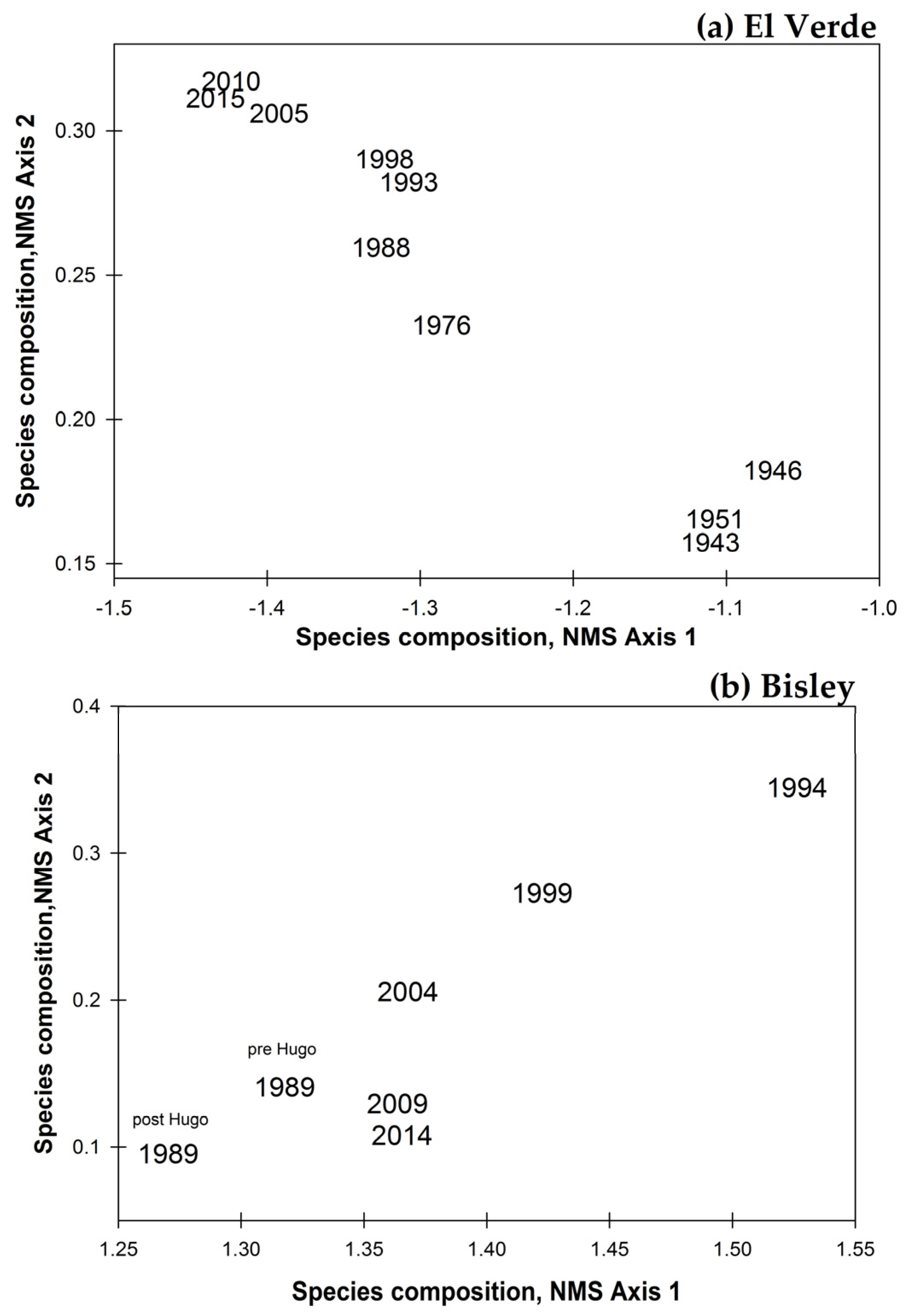
In the El Verde plot, species composition of each census through time occupies a distinct part of ordination space that continues to change through time (Figure 8a). The last three censuses at El Verde, 2005, 2010 and 2015, have high association in their species composition as evidenced by their very close scatter in the multivariate species space. This is also represented with standardized chi-square distance pairwise comparisons between the 2005 and 2010 census (0.60), and the 2010 and 2015 census (0.91). In contrast, the initial census dates, 1943, 1946, and 1951, are very distantly scattered in El Verde species ordination space in relation to the most recent census dates 2005, 2010, and 2015. This is also evidenced by the pairwise distance comparisons between 1943 and 2005 (0.11), 1946 and 2005 (0.01), 1951 and 2005 (−0.07), 1951 and 2010 (0.05), 1951 and 2015 (0.07), i.e., all very low values for similarity in species composition. The three initial censuses, 1943, 1946 and 1951, share great similarity in species composition, evidenced by the pairwise comparison distance values between 1943 and 1951 (0.74), 1946 and 1951 (0.77). These initial census dates occupy a distinctly separate area of ordination space to that of the most recent El Verde censuses. In contrast, community species composition seems to be converging in Bisley, with the most recent censuses, 2009 and 2014, occupying a closely scattered area of species ordination space as the pre-hurricane 1989 census (Figure 8b). The 2009 and 2014 census were the closest in the Bisley multivariate species space (pairwise distance 0.84), and the 2014 census was marginally similar to the species composition of the 1989 census after hurricane Hugo (0.43). The 1994 census species composition, five years after Hugo, had the greatest difference in species composition with respect to the pre-hurricane census (0.26). The species C. schreberiana and P. berteriana were at their highest stem densities in Bisley during the 1994 census and in El Verde during the 1993 census.
4. Discussion
The patterns of vegetation dynamics observed have components that are broadly representative of tabonuco forests and also display site-specific responses associated with location in relation to hurricane disturbance events. Site-specific responses such as accumulated basal area depend on the magnitude of effects and frequency of disturbance events [7,14,24,41,42]. Tabonuco forest’s structural attributes such as basal area are able to recover from disturbances that remove as much as half of the above-ground biomass [13] and also have the capacity to continue accumulating basal area through time when cumulative disturbance effects are minimal [12].
The factors that contribute to these responses are various and include the life-history patterns of dominant species in this forest type [9,37,43]. The patterns presented in common at both El Verde and Bisley sites were those of the dominant species in these long-term plots, which had stem densities with stable and mostly increasing trends in all censuses after hurricane Hugo in 1989. In this forest type, the attributes of the dominant species, D. excelsa, M. bidentata and P. montana, drive the combination of responses of stem density and basal area at these long-term plot sites [14,24,35]. The resistance and increases observed from dominant species drive forest structure responses. This was also found in modeled simulations of tabonuco forest with individual-based species responses to disturbance, where low temporal variation was observed as the response of dominant species [37]. At both long-term study sites, a rapid response of increased stem density from the early successional species C. schreberiana and P. berteriana is a proxy for canopy formation and light availability, which starts the process of facilitation and changing conditions in the understory while increasing basal area [2,20,36,37]. Once the canopy closes and light availability changes, these early successional species not only respond negatively to canopy closure, but they are also susceptible to pathogens due to their high intra-species post-hurricane densities [44,45]. However, the patterns that contrast the most are those between stem density fluctuations for two of the most abundant species, P. montana and S. berteriana. The palm P. montana has increased steadily at both sites while S. berteriana has presented a trend towards decrease. An increase in palm density in tabonuco forest stands had been noted at El Verde since the 1976 measurements [10] and discussed in a long-term analysis of palm forests compared with tabonuco forests stands [14]. The decreasing trend observed in the El Verde plot with S. berteriana could be due to initial stem density values being relatively high, as these could have been still responding to the effects of the previous San Ciprián hurricane [10]. Because decreases in stem density of S. berteriana at El Verde, and to a lesser degree at Bisley, occur simultaneously with increases in P. montana, another possibility could be that perhaps P. montana could be occupying sites for S. berteriana recruitment as both of these species are found in wet and concave locations [13,26,27,30,34].
These two tabonuco forest sites experienced different disturbance conditions and are at different points in their response to disturbance or successional trajectories. Due to a difference in their location and geographical aspect, the plots seem to be responding to effects from different disturbance events. Analyses of the effect of the passage of hurricanes over Caribbean, Philippines and Australian forests highlighted the importance of site location and aspect relative to wind direction in the interpretation of hurricane effects [9,14,36,42,43,46,47,48,49]. These are fundamental observations when assessing forest responses to these events. The increases in basal area at El Verde seem primarily due to increases in already established stems of the 30 to 40 cm and 40 to 50 cm diameter categories, which are trees that have been long-time residents of the plot and are resisting and growing through time. This is not the case in the Bisley plot, where trees of the 30 to 40 cm and 40 to 50 cm dbh categories are less abundant, contribute proportionally less to basal area, and have not had a net increase in stem density during the study period. Similar windward and leeward site responses were observed in palm forest stands in the LEF. Greater hurricane Hugo effects were measured on the windward slopes where a long-term palm forest study plot is located, while much fewer effects were observed in a palm forest study plot on a leeward slope [14].
The effects from hurricane disturbances maintain these forests in a constant state of structural and compositional change in response to intensity of the cumulative effects. At Bisley, there was a greater successional trajectory dynamic due to the stronger hurricane effect on stem density and basal area. At El Verde, the hurricane defoliation had minimal immediate effects on stem density and basal area, while increasing canopy opening and light to the forest floor [2,20,36].
Thus, El Verde has been maintained in a directional successional transition, as effects from hurricane Hugo were minimal, and it is still in a response trajectory from the effects of San Ciprián in 1932 [10,12]. When hurricane Georges reached the Bisley site, it was within the first ten years of recovery from the changes in structure that resulted following hurricane Hugo. Although no significant effects were observed in terms of stem density and basal area at Bisley following hurricane George in 1998 [13], the forest site benefited from the canopy defoliation and associated increases of light to the forest floor by continuing or extending on the particular succession trajectory set by hurricane Hugo. At the El Verde site, it seems that due to the time since hurricane San Ciprián and the minimal effects from Hugo, the forest has continued on a succession path defined by increased growth, accrued structure and basal area that continues surpassing initial measurement conditions [10,12]. The apparent asymptote at Bisley for basal area values is likely due to the steady loss of the accumulated large stems of early successional species being replaced by smaller shade tolerant stems of the dominant species. If the next hurricane at Bisley has moderate or minimal effects on forest structure, limited to defoliation type effects, then basal area should be expected to increase, as has been observed at El Verde.
In response to disturbance events such as hurricanes, forest community dynamics depend on the interaction of processes that occur at different time scales. Tree structure can recover to pre-disturbance values within a decade in the tabonuco forest of the LEF. This is due to the tree species community being able to shift composition in such a way that only the dominant species are maintained in association with a flux of changes from other species. During the 72 years of vegetation censuses at El Verde and the 25 years at Bisley, concomitant changes in species composition have allowed for forest structure components such as basal area to either recover and maintain pre-disturbance values (such as observed in Bisley), or to progressively increase (as observed in El Verde). If there is now a period of hurricane-free conditions for another 10–15 years, perhaps the tabonuco forests at the LEF will slow down their pattern of accretion of basal area under undisturbed and closed canopy conditions. Before hurricane Hugo in 1989, both El Verde and Bisley were at similar values of basal area, but because of the different disturbance effects, their responses diverged. The defoliation and trimming effect seems to be what has allowed El Verde to increase in basal area, due to a combination of the temporary increase in C. schreberiana, and the increases in basal area contributed by the stems of already established and resistant dominant species that are now at the canopy level. It is proposed that another storm or hurricane disturbance in Bisley, similar to hurricane Georges, will produce the canopy defoliation needed to accumulate greater basal area at this forest site.
Following hurricanes, recovery and resilience of different components of the forest ecosystem have their own response intervals [20]. Tree structural characteristics and processes such as litterfall rates are the fastest to recover [31,34]. On the other hand, tree and understory species composition shift in more dramatic ways and are not able to completely return to pre-disturbance conditions at the same time scales [13,50]. Species composition occurs in the context of both changes in environmental conditions set after the particular disturbance event, plus the pool of species present at the site. The range of species at a site is limited by the constraints of their individual life history dynamics and responses to disturbance [37,41,51]. The insights into forest structure and composition point to the nuances of changes in species dynamics during succession, with individual species population dynamics and interactions among species still in need of further exploration.
Acknowledgments
This USDA Forest Service (USFS) International Institute of Tropical Forestry (IITF) research was conducted in collaboration with the University of Puerto Rico (UPR) as part of IITF contributions to the Luquillo Long-Term Ecological Research Program (LTER) in the LEF. Assistance with measurements of the El Verde plot was provided by USFS SRS-FIA and SUNY-ESF collaborators via Cooperative Grants and Agreements (N IITF-98-CA-008 and 05-DG-11120101-016) from the USFS IITF and Edna Bailey Sussman and Farnsworth Funds- SUNY-ESF. This is a contribution part of IITF’s 75th Anniversary Research Symposium. The revision and suggestions from F.H. Wadsworth, D.J. Lodge, A.E. Lugo, S.A. Sloan and three anonymous reviewers improved this manuscript. The findings, conclusions, and views expressed in this manuscript are those of the author and do not necessarily represent the views of the USDA Forest Service.
Conflicts of Interest
The author declares no conflict of interest.
References
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef]
- Brokaw, N.V.L.; Walker, L.R. Summary of the effects of Caribbean hurricanes on vegetation. Biotropica 1991, 23, 442–447. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Willig, M.R.; Walker, L.R.; Silver, W.L. Introduction: Disturbance and Caribbean ecosystems. Biotropica 1996, 28, 414–423. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- López-Marrero, T.; Heartsill-Scalley, T. Get up, stand up: Environmental situation, threats, and opportunities in the insular Caribbean. Caribb. Stud. 2012, 40, 3–14. [Google Scholar] [CrossRef]
- Lugo, A.E.; Helmer, E.H.; Santiago Valentín, E. Caribbean landscapes and their biodiversity. Interciencia 2012, 37, 705–710. [Google Scholar]
- Lugo, A.E. Visible and invisible effects of hurricanes on forest ecosystems: An international review. Austral Ecol. 2012, 33, 368–398. [Google Scholar] [CrossRef]
- Chai, S.L.; Healey, J.R.; Tanner, E.V. Evaluation of forest recovery over time and space using permanent plots monitored over 30 years in a Jamaican montane rain forest. PLoS ONE 2012, 7, e48859. [Google Scholar] [CrossRef] [PubMed]
- Luke, D.; McLaren, K.; Wilson, B. Modeling hurricane exposure in a Caribbean lower montane tropical wet forest: The Effects of frequent, intermediate disturbances and topography on forest structural dynamics and composition. Ecosystems 2016, 19, 1178–1195. [Google Scholar] [CrossRef]
- Crow, T.R. A rainforest chronicle: A 30-year record of change in structure and composition at El Verde, Puerto Rico. Biotropica 1980, 12, 42–55. [Google Scholar] [CrossRef]
- Johnston, M.H. Successional Change and Species/Site Relationships in a Puerto Rican Tropical Forest. Ph.D. Thesis, State University of New York, New York, NY, USA, 1991; p. 253. [Google Scholar]
- Drew, A.P.; Boley, J.D.; Zhao, Y.; Johnston, M.H.; Wadsworth, F.H. Sixty-two years of change in subtropical wet forest structure and composition at El Verde, Puerto Rico. Interciencia 2009, 34, 34–40. [Google Scholar]
- Heartsill, S.; Tamara, S.; Frederick, N.; Lugo, A.E.; Moya, S.; Estrada Ruiz, C.R. Changes in structure, composition, and nutrients during 15 years of hurricane-induced succession in a subtropical wet forest in Puerto Rico. Biotropica 2010, 42, 455–463. [Google Scholar] [CrossRef]
- Lugo, A.E.; Frangi, J.L. Long-term response of Caribbean palm forests to hurricanes. Caribb. Nat. 2016, 157–175. [Google Scholar]
- Walker, L.R.; Zarin, D.J.; Fetcher, N.; Myster, R.W.; Johnson, A.H. Ecosystem development and plant succession on landslides in the Caribbean. Biotropica 1996, 28, 566–576. [Google Scholar] [CrossRef]
- Coll, M.; Fonseca, A.C.; Cortés, J. The mangrove and others vegetation associations in de Gandoca lagoon, Limón, Costa Rica. Rev. Biol. Trop. 2001, 49, 321–329. [Google Scholar] [PubMed]
- Heartsill Scalley, T.; Scatena, F.N.; Estrada, C.; McDowell, W.H.; Lugo, A.E. Disturbance and long-term patterns of rainfall and throughfall nutrient fluxes in a subtropical wet forest in Puerto Rico. J. Hydrol. 2007, 333, 472–485. [Google Scholar] [CrossRef]
- Gonzalez, G.; Waide, R.B.; Willig, M.R. Advancements in the understanding of spatiotemporal gradients in tropical landscapes: A Luquillo focus and global perspective. Ecol. Bull. 2013, 54, 245–250. [Google Scholar]
- Thomas, S.C.; Martin, A.R.; Mycroft, E.E. Tropical trees in a wind-exposed island ecosystem: Height-diameter allometry and size at onset of maturity. J. Ecol. 2015, 103, 594–605. [Google Scholar] [CrossRef]
- Brokaw, N.; Zimmerman, J.K.; Willig, M.; Camilo, G.; Covich, A.; Crowl, T.; Fetcher, N.; Haines, B.; Lodge, J.; Lugo, A.E.; et al. A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response; Brokaw, N., Crowl, T.A., Lugo, A.E., McDowell, W.H., Scatena, F.N., Waide, R.B., Willig, M.R., Eds.; Oxford University Press: New York, NY, USA, 2012; pp. 201–271. [Google Scholar]
- Taylor, M.A.; Stephenson, T.S.; Chen, A.A.; Stephenson, K.A. Climate change and the Caribbean: Review and response. Caribb. Stud. 2012, 40, 169–200. [Google Scholar] [CrossRef]
- Beard, J.S. The Natural Vegetation of the Windward and Leeward Islands; Clarendon Press: Oxford, UK, 1949; p. 192. [Google Scholar]
- Alvarez Ruiz, M. Effects of Human Activities on Stand Structure and Composition, and Genetic Diversity of Dacryodes Excelsa Vahl (Tabonuco); University of Puerto Rico: San Juan, Puerto Rico, 2002; p. 266. [Google Scholar]
- Lugo, A.E.; Scatena, F.N. Ecosystem-level properties of the Luquillo Experimental Forest with emphasis on the Tabonuco Forest. In Tropical Forests: Management and Ecology; Springer: New York, NY, USA, 1995; pp. 59–108. [Google Scholar]
- Basnet, K.; Scatena, F.N.; Likens, G.E.; Lugo, A.E. Ecological consequences of root grafting in Tabonuco (Dacryodes excelsa) trees in the Luquillo Experimental Forest, Puerto Rico. Biotropica 1993, 25, 28–35. [Google Scholar] [CrossRef]
- Weaver, P.L. Sloanea Berteriana Choisy Motillo. Elaeocarpaceae. Elaeocarpus Family; USDA Forest Service, International Institute of Tropical Forestry: Rio Piedras, Puerto Rico, 1997; p. 7.
- Lugo, A.E.; Francis, J.K.; Frangi, J.L. Prestoea Montana (R. Graham) Nichols. Sierra Palm. Palmaceae. Palm Family; US Department of Agriculture, Forest Service, International Institute of Tropical Forestry: Rio Piedras, Puerto Rico, 1998; p. 9.
- Weaver, P.L. Ecological observations on Magnolia splendens urban in the Luquillo mountains of Puerto Rico. Caribb. J. Sci. 1987, 23, 340–351. [Google Scholar]
- Garcia-Montiel, D.; Scatena, F.N. The effect of human activity on the structure and composition of a tropical forest in Puerto Rico. Forest Ecol. Manag. 1994, 63, 57–78. [Google Scholar] [CrossRef]
- Scatena, F.N. An Introduction to the Physiography and History of the Bisley Experimental Watersheds in the Luquillo Mountains of Puerto Rico; Gen. Tech. Rep. SO-72. U.S.D.A; Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1989; p. 22.
- Scatena, F.N.; Moya, S.; Estrada, C.; Chinea, J.D. The first five year in the reorganization of aboveground biomass and nutrient use following Hurricane Hugo in the Bisley Experimental Watersheds, Luquillo Experimental Forest, Puerto Rico. Biotropica 1996, 28, 424–440. [Google Scholar] [CrossRef]
- Ostertag, R.; Silver, W.L.; Lugo, A.E. Factors affecting mortality and resistance to damage following hurricanes in a rehabilitated subtropical moist forest. Biotropica 2005, 37, 16–24. [Google Scholar] [CrossRef]
- Larsen, M.C. Analysis of 20th century rainfall and streamflow to characterize drought and water resources in Puerto Rico. Phys. Geogr. 2000, 21, 494–521. [Google Scholar]
- Beard, K.H.; Vogt, K.A.; Vogt, D.J.; Scatena, F.N.; Covich, A.P.; Sigurdardottir, R.; Siccama, T.G.; Crowl, T.A. Structural and functional responses of a subtropical forest to 10 year of hurricanes and droughts. Ecol. Monogr. 2005, 75, 345–361. [Google Scholar] [CrossRef]
- Lugo, A.E.; Zimmerman, J.K. Ecological Life Histories. In Tropical Tree Seed Manual; Vozzo, J.A., Ed.; USDA Forest Service: Washington, DC, USA, 2002; pp. 191–213. [Google Scholar]
- Shiels, A.B.; Zimmerman, J.K.; García-Montiel, D.C.; Jonckheere, I.; Holm, J.; Horton, D.; Brokaw, N. Plant responses to simulated hurricane impacts in a subtropical wet forest, Puerto Rico. J. Ecol. 2010, 98, 659–673. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Murphy, L.; Sabat, A.M.; Fetcher, N.; Haines, B.L. Natural disturbance and human land use as determinants of tropical forest dynamics: Results from a forest simulator. Ecol. Monogr. 2009, 79, 423–443. [Google Scholar] [CrossRef]
- Johnson, D.E. Applied Multivariate Methods for Data Analysts; Duxbury Press: Pacific Grove, CA, USA, 1998; p. 567. [Google Scholar]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MJM Software Design: Gleneden Beach, OR, USA, 2002; p. 300. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD. In Multivariate Analysis of Ecological Data; Version 5.1; Mjm Software: Gleneden Beach, OR, USA, 2006. [Google Scholar]
- Lugo, A.E. Effects and outcomes of Caribbean hurricanes in a climate change scenario. Sci. Total Environ. 2000, 262, 243–251. [Google Scholar] [CrossRef]
- Boose, E.R.; Foster, D.R.; Fluet, M. Hurricane impacts to tropical and temperate forest landscapes. Ecol. Monogr. 1994, 64, 369–400. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Tanner, E.V.J.; Healey, J.R. Damage and responsiveness of Jamaican montane forest tree species after disturbance by a hurricane. Ecology 1995, 76, 2562–2580. [Google Scholar] [CrossRef]
- Lodge, D.J.; Cantrell, S. Fungal communities in wet tropical forests: Variation in time and space. Can. J. Bot. 1995, 73, 1391–1398. [Google Scholar] [CrossRef]
- Lodge, D.J. Microorganisms. In The Food Wed of a Tropical Forest; Reagan, D.P., Waide, R.B., Eds.; University of Chicago press: Chicago, IL, USA, 1996; pp. 53–108. [Google Scholar]
- Yap, S.L.; Davies, S.J.; Condit, R. Dynamic response of a Philippine dipterocarp forest to typhoon disturbance. J. Veg. Sci. 2016, 27, 133–143. [Google Scholar] [CrossRef]
- Murphy, H.T.; Metcalfe, D.J.; Bradford, M.G.; Ford, A.J. Community divergence in a tropical forest following a severe cyclone. Austral Ecol. 2014, 39, 696–709. [Google Scholar] [CrossRef]
- Foster, D.R.; Knight, D.H.; Franklin, J.F. Landscape patterns and legacies resulting from large, infrequent forest disturbances. Ecosystems 1988, 1, 497–510. [Google Scholar] [CrossRef]
- Willig, M.R.; Bloch, C.P.; Covich, A.P.; Hall, C.A.S.; Lodge, D.J.; Lugo, A.E.; Silver, W.L.; Waide, R.B.; Walker, L.R.; Zimmerman, J.K. Long-term research in the Luquillo Mountains, synthesis and foundations for the future. In A Caribbean Forest Tapestry: The Multidimensional Nature of Disturbance and Response; Brokaw, N., Crowl, T.A., Lugo, A.E., Mcdowell, W.H., Scatena, F.N., Waide, R.B., Willig, M.R., Eds.; Oxford University Press: New York, NY, USA, 2012; pp. 361–441. [Google Scholar]
- Imbert, D.; Labbe, P.; Rousteau, A. Hurricane damage and forest structure in Guadeloupe, French West Indies. J. Trop. Ecol. 1996, 12, 663–680. [Google Scholar] [CrossRef]
- Royo, A.A.; Scalley, T.H.; Moya, S.; Scatena, F.N. Non-arborescent vegetation trajectories following repeated hurricane disturbance: Ephemeral versus enduring responses. Ecosphere 2011, 2, 77. [Google Scholar] [CrossRef]
Figure 1.
Location of Puerto Rico in the context of the Caribbean, and inset of the El Verde Research Area and Bisley Experimental Watersheds, both long-term sites in the Luquillo Experimental Forest.
Figure 1.
Location of Puerto Rico in the context of the Caribbean, and inset of the El Verde Research Area and Bisley Experimental Watersheds, both long-term sites in the Luquillo Experimental Forest.
Figure 2.
Views of vegetation from the Luquillo Experimental Forest, Puerto Rico. Top panel (a) Tabonuco, Dacryodes excelsa, the dominant tree species in forests at lower montane elevations; Bottom panel (b) Sierra palm, Prestoea montana, with stems found at all elevations and able to form mono-specific forest stands. Photos by Jerry Bauer.
Figure 2.
Views of vegetation from the Luquillo Experimental Forest, Puerto Rico. Top panel (a) Tabonuco, Dacryodes excelsa, the dominant tree species in forests at lower montane elevations; Bottom panel (b) Sierra palm, Prestoea montana, with stems found at all elevations and able to form mono-specific forest stands. Photos by Jerry Bauer.

Figure 3.
Density of stems (≥4 cm dbh) per hectare in the El Verde permanent plot (grey triangles) and in Bisley (filled circles), Luquillo Experimental Forest, Puerto Rico. Arrows represent hurricane Hugo (1989) and Georges (1998).
Figure 3.
Density of stems (≥4 cm dbh) per hectare in the El Verde permanent plot (grey triangles) and in Bisley (filled circles), Luquillo Experimental Forest, Puerto Rico. Arrows represent hurricane Hugo (1989) and Georges (1998).
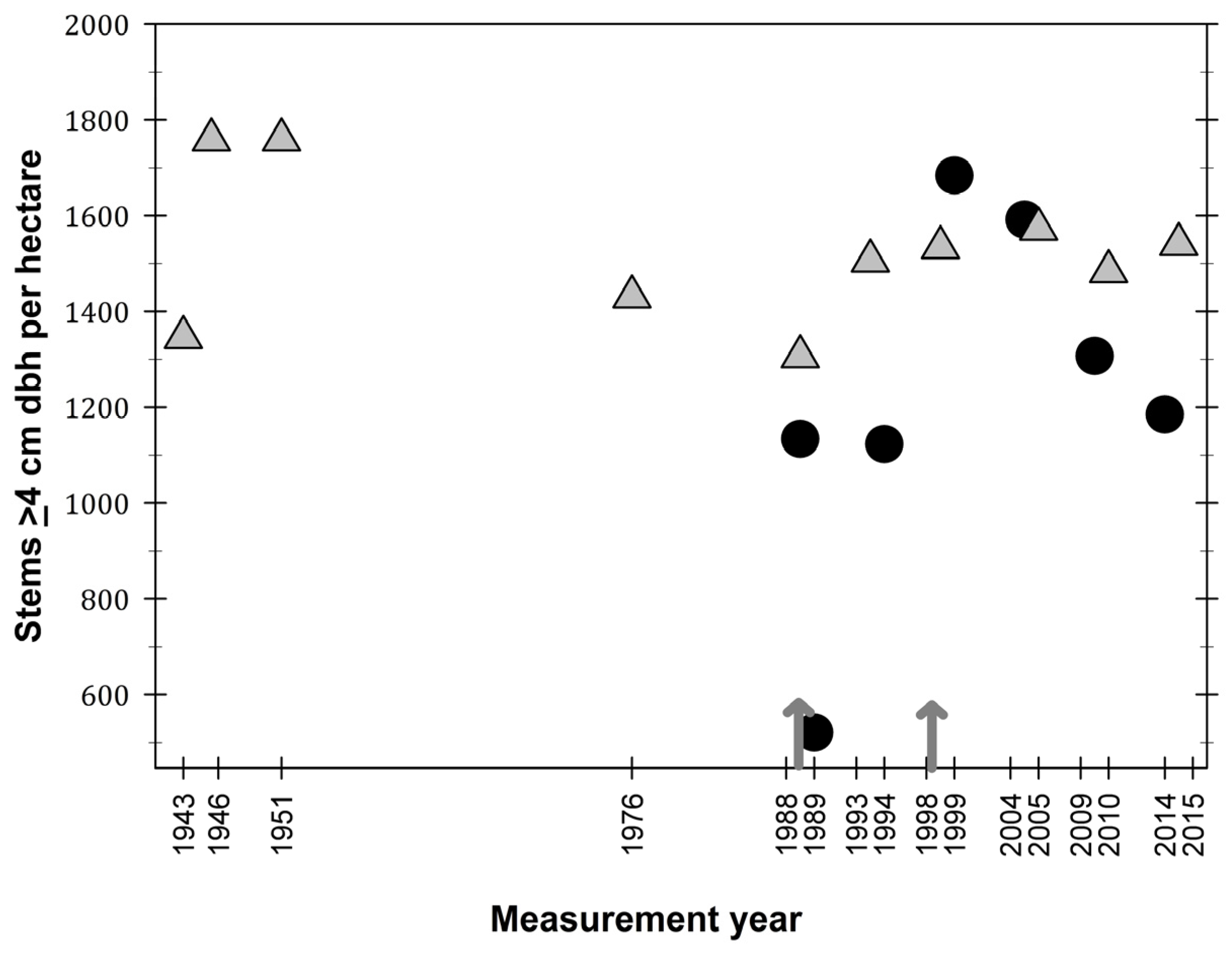
Figure 4.
Basal area (m2/ha) values in the El Verde permanent plot site (grey triangles) and the Bisley site (filled circles), Luquillo Experimental Forest, Puerto Rico. Arrows represent hurricane Hugo (1989) and Georges (1998).
Figure 4.
Basal area (m2/ha) values in the El Verde permanent plot site (grey triangles) and the Bisley site (filled circles), Luquillo Experimental Forest, Puerto Rico. Arrows represent hurricane Hugo (1989) and Georges (1998).
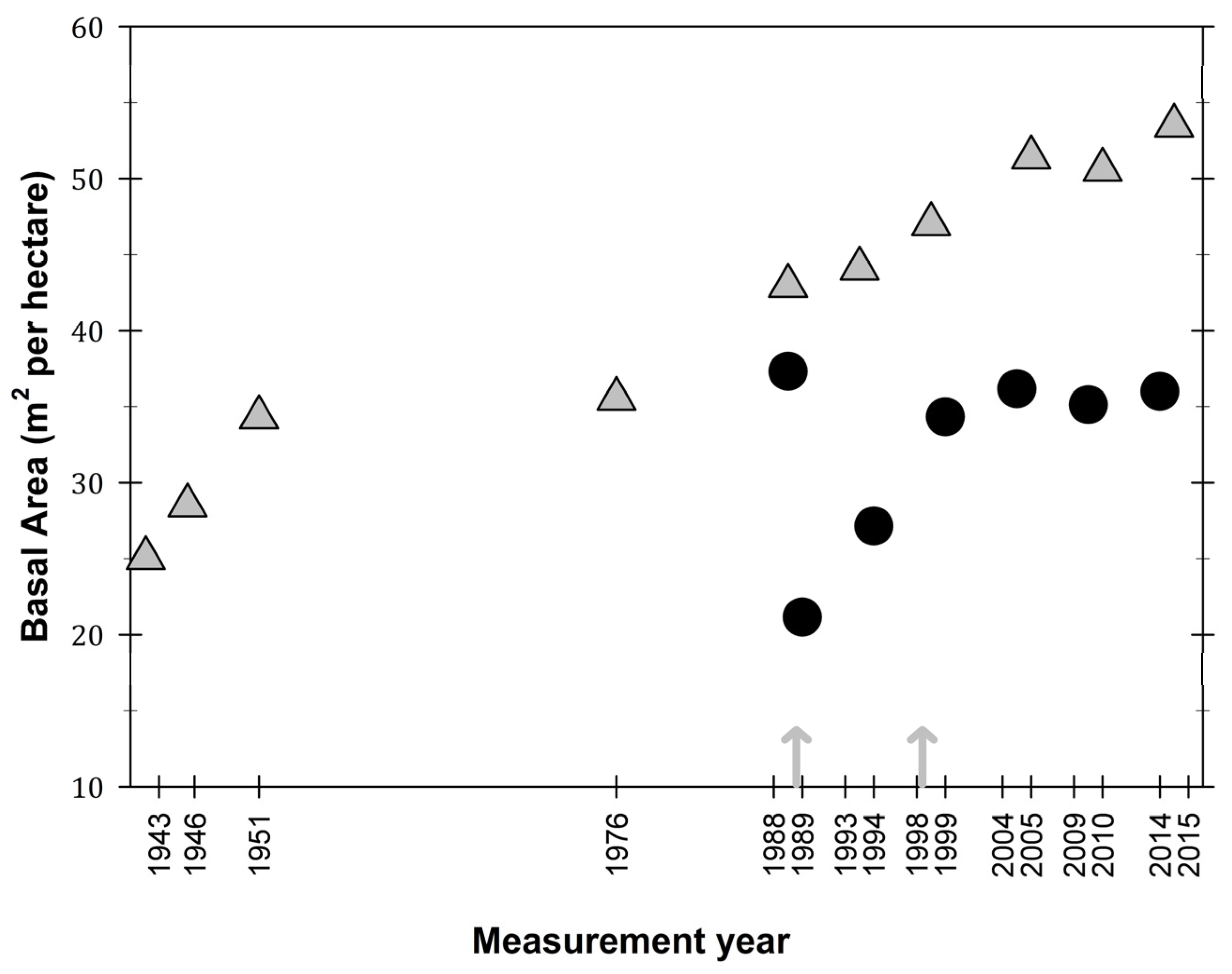
Figure 5.
Diameter size category distribution for each census year for El Verde (a) top panel and (b) Bisley (bottom panel). Each bar represents a census year.
Figure 5.
Diameter size category distribution for each census year for El Verde (a) top panel and (b) Bisley (bottom panel). Each bar represents a census year.
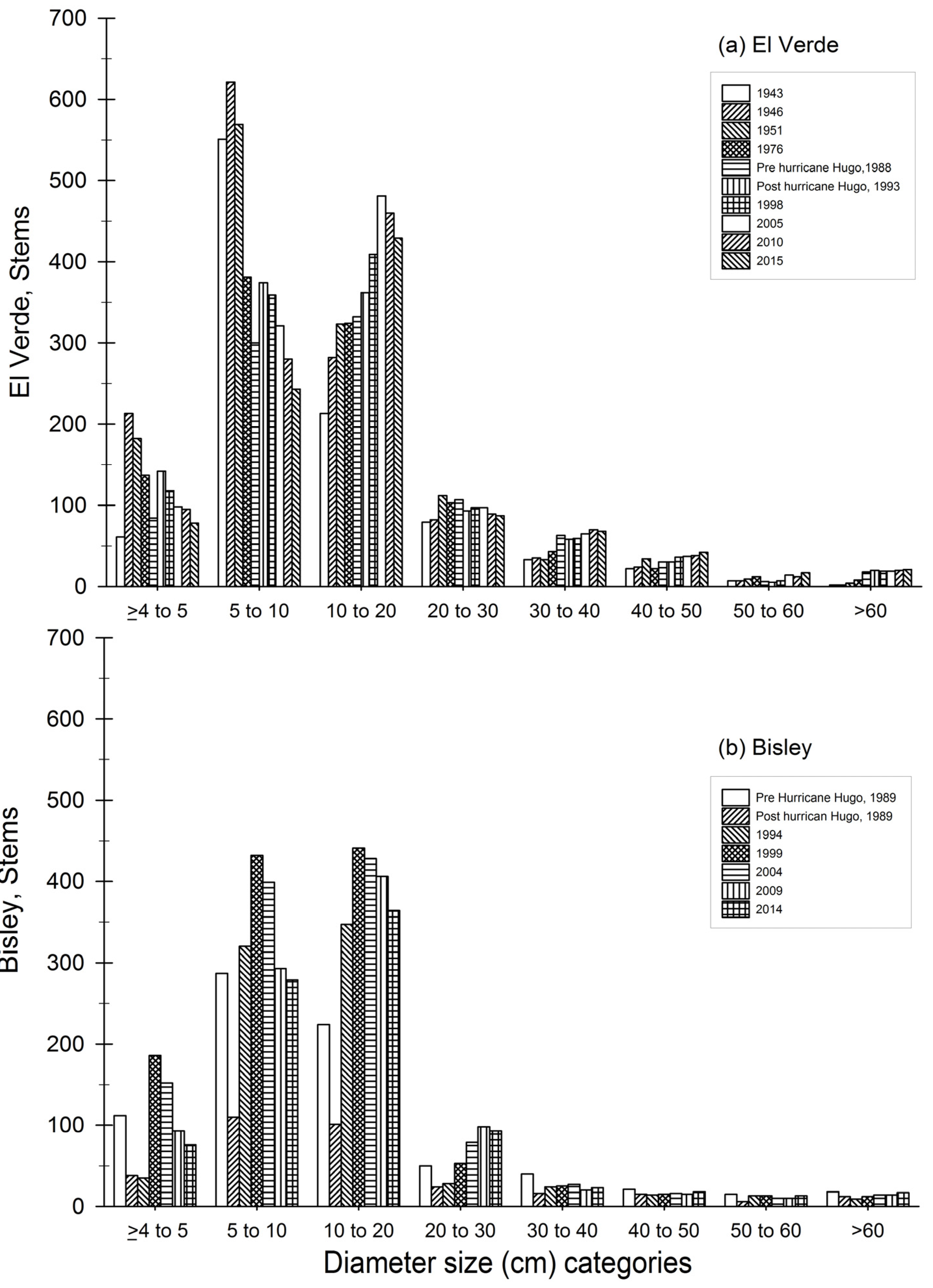
Figure 6.
Stem density per each census year for selected shade tolerant/late successional species in two long-term research plots, Luquillo Experimental Forest, Puerto Rico. Panel (a) El Verde, with triangle symbols; panel (b) Bisley with circle symbols.
Figure 6.
Stem density per each census year for selected shade tolerant/late successional species in two long-term research plots, Luquillo Experimental Forest, Puerto Rico. Panel (a) El Verde, with triangle symbols; panel (b) Bisley with circle symbols.
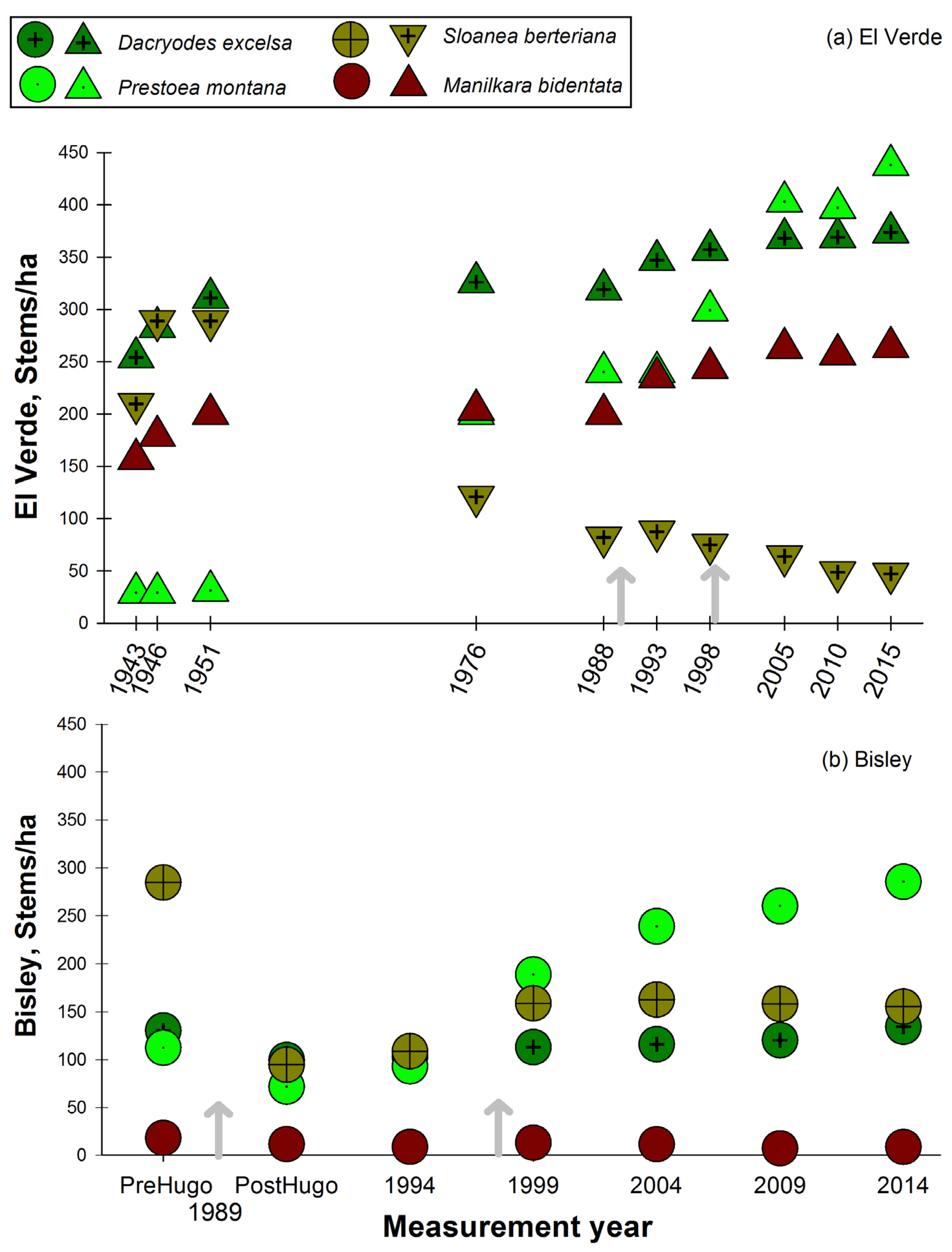
Figure 7.
Stem density per each census year for selected shade intolerant/early successional species in two long-term research plots, Luquillo Experimental Forest, Puerto Rico. Panel (a) El Verde, with triangle symbols; panel (b) Bisley with circle symbols.
Figure 7.
Stem density per each census year for selected shade intolerant/early successional species in two long-term research plots, Luquillo Experimental Forest, Puerto Rico. Panel (a) El Verde, with triangle symbols; panel (b) Bisley with circle symbols.

Figure 8.
Non-metric multidimensional scaling (NMS) ordination based on species composition and abundance per census of two tabonuco forest permanent plot sites at Luquillo Experimental Forest, Puerto Rico. Top panel (a) El Verde and bottom panel (b) Bisley.
Figure 8.
Non-metric multidimensional scaling (NMS) ordination based on species composition and abundance per census of two tabonuco forest permanent plot sites at Luquillo Experimental Forest, Puerto Rico. Top panel (a) El Verde and bottom panel (b) Bisley.

© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Heartsill Scalley, T. Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains. Forests 2017, 8, 204. https://doi.org/10.3390/f8060204
AMA Style
Heartsill Scalley T. Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains. Forests. 2017; 8(6):204. https://doi.org/10.3390/f8060204
Chicago/Turabian StyleHeartsill Scalley, Tamara. 2017. "Insights on Forest Structure and Composition from Long-Term Research in the Luquillo Mountains" Forests 8, no. 6: 204. https://doi.org/10.3390/f8060204
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.