Fungal Community and Ligninolytic Enzyme Activities in Quercus deserticola Trel. Litter from Forest Fragments with Increasing Levels of Disturbance

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Litter Sampling
2.2. Litter and Soil Nutrients Analyses
2.3. Enzyme Activity Assays
2.3.1. Laccase (Lac; EC 1.10.3.2)
2.3.2. Lignin Peroxidase (LiP; EC 1.11.1.14)
2.3.3. Manganese Peroxidase (MnP; EC 1.11.1.13)
2.4. Statistical Analyses of Enzyme Activity
2.5. DNA Extraction, PCR Assays, Cloning, and Sequencing
2.6. Bioinformatics and Fungal Community Analyses
3. Results
3.1. Nutrients and Vegetation in Studied Plots
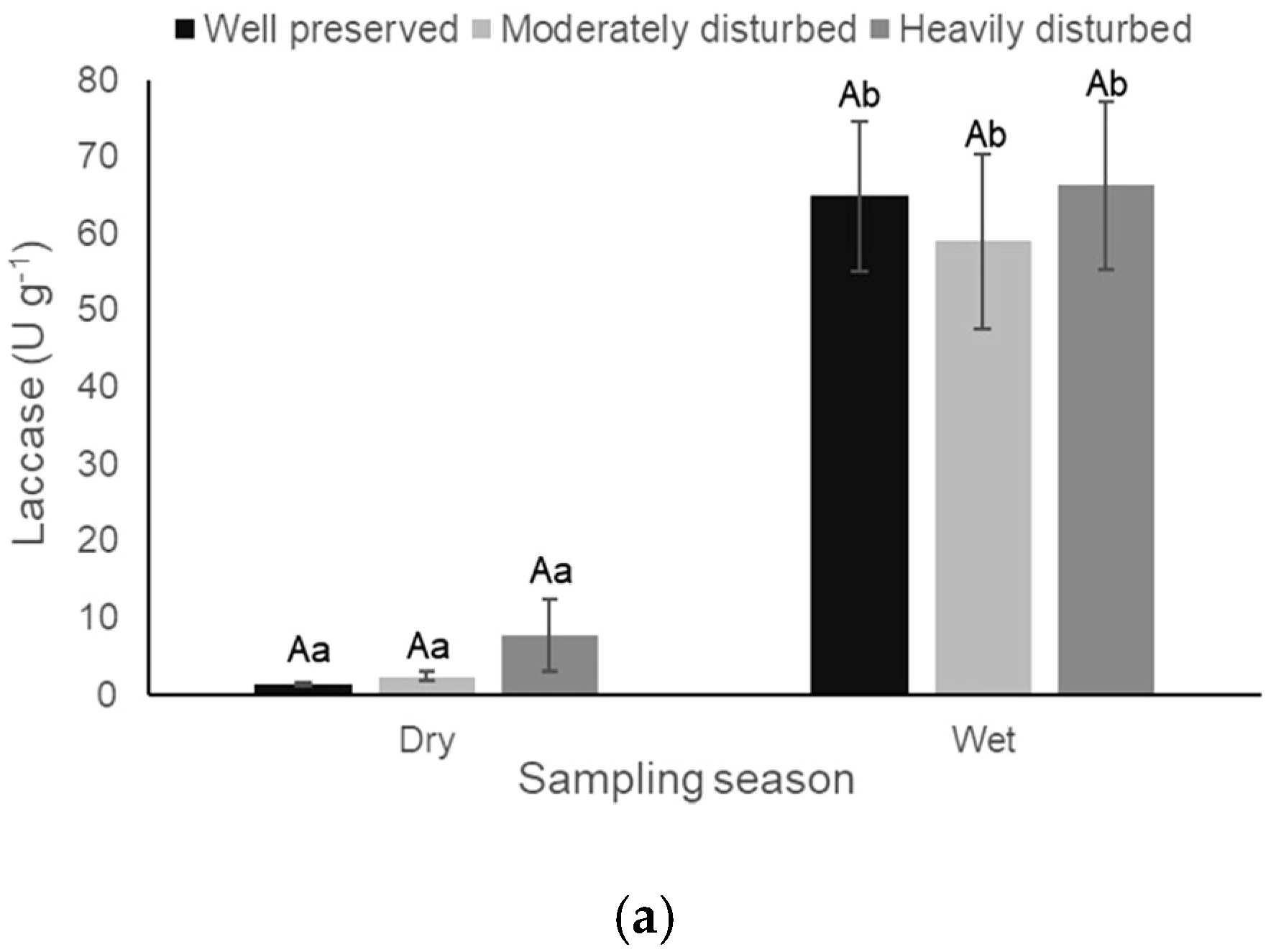
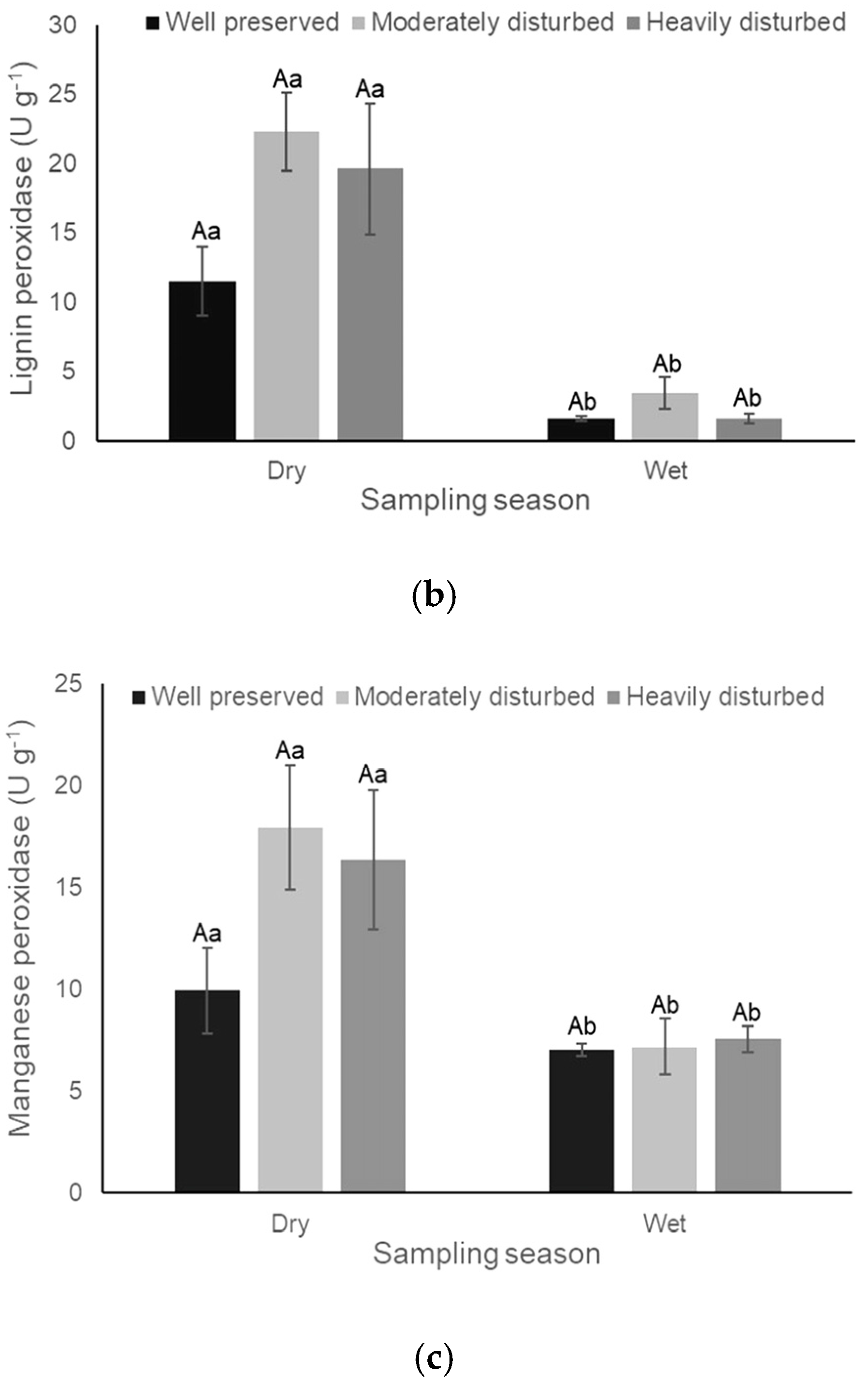
3.2. Enzyme Activity
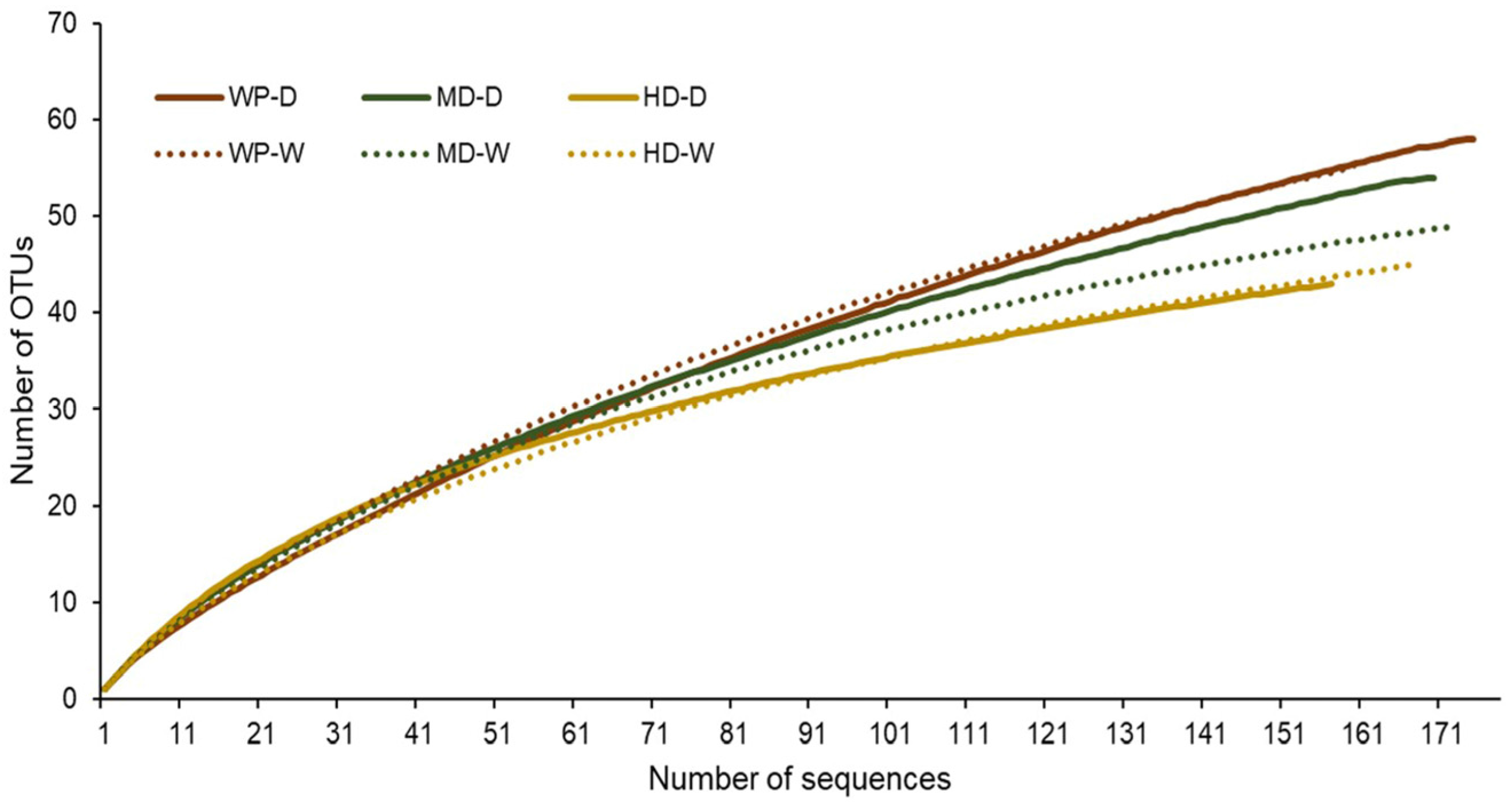
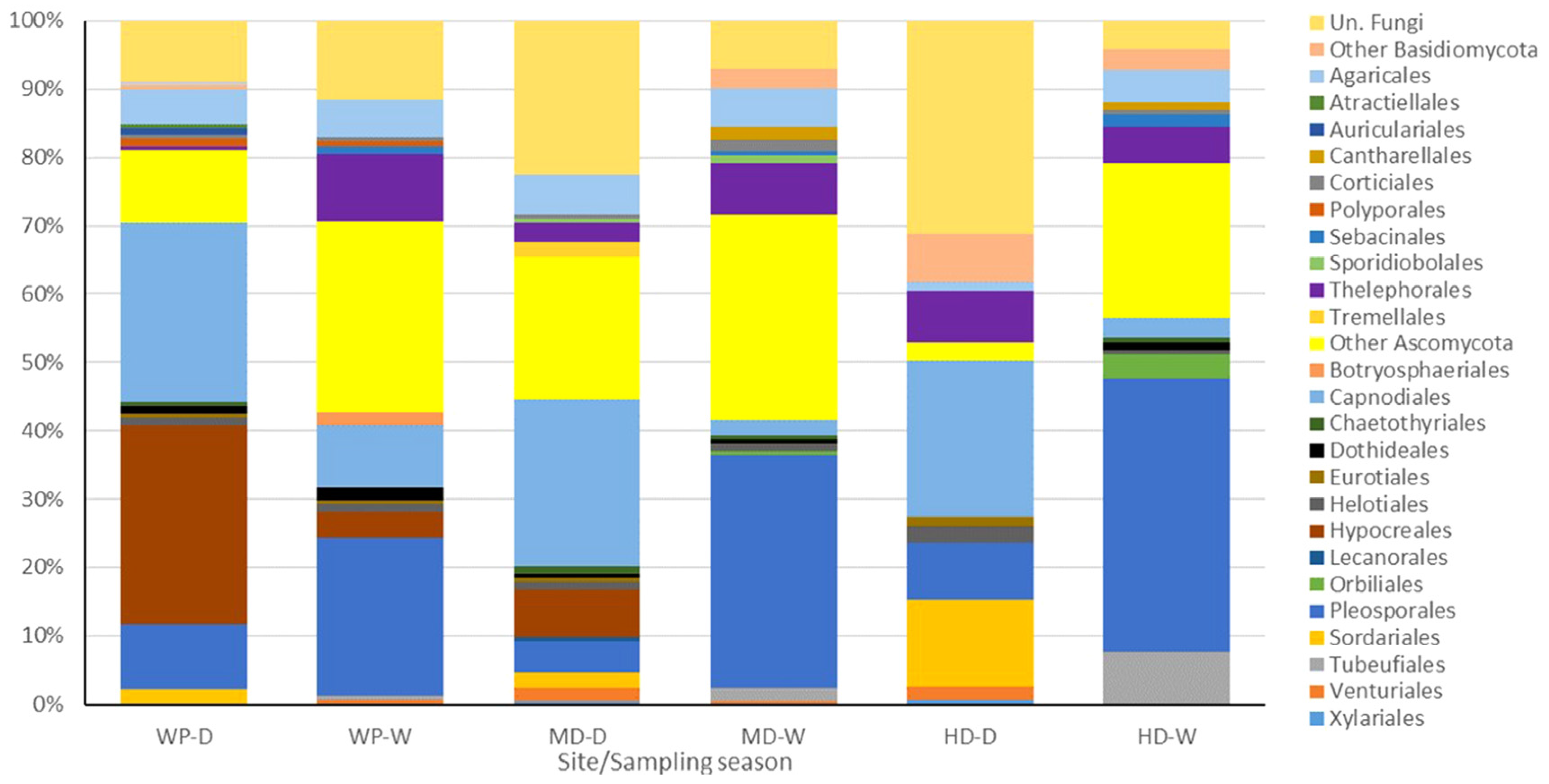
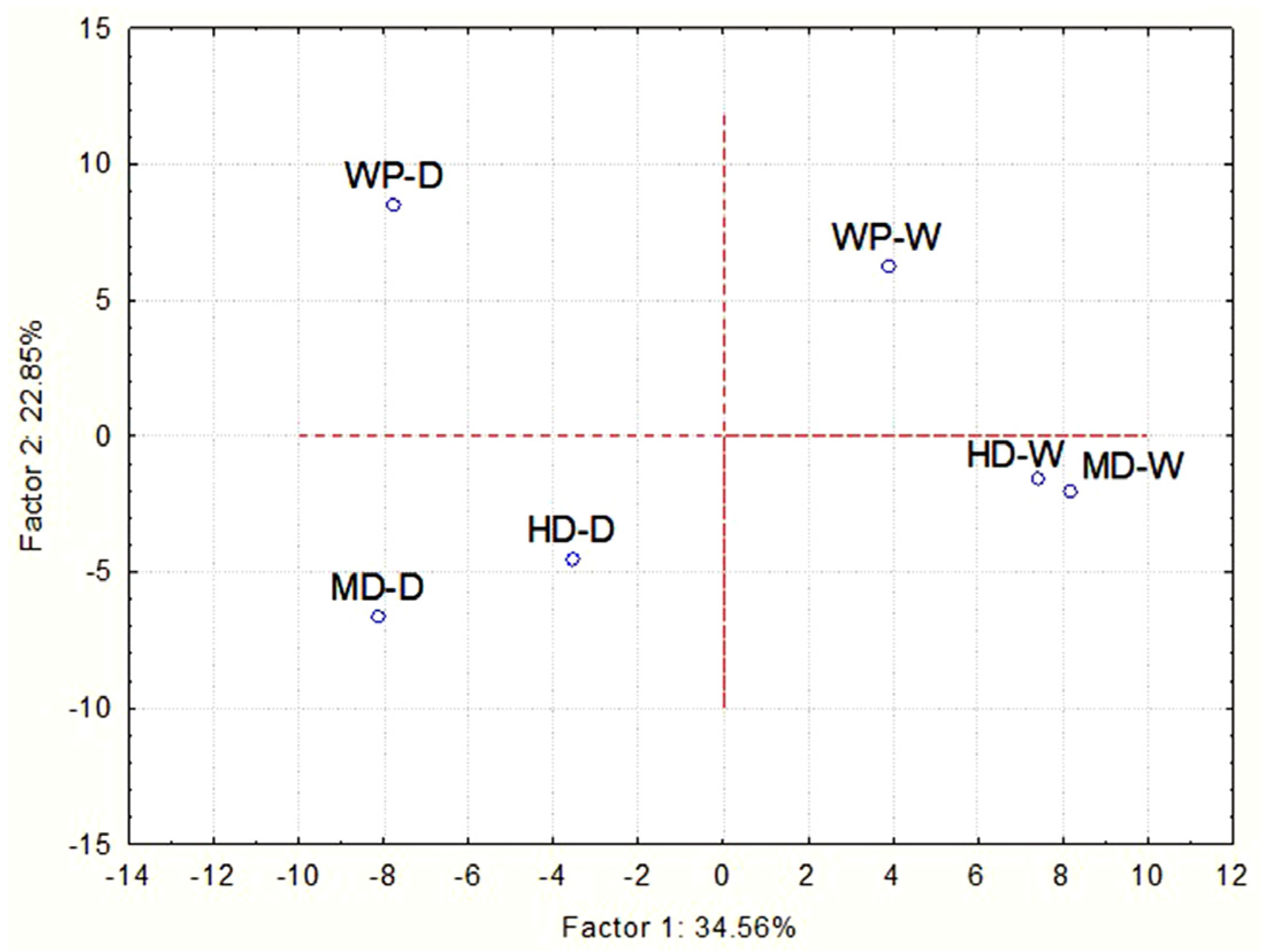
3.3. Analysis of Fungal Communities
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; Blackwell: London, UK, 1979; p. 372. [Google Scholar]
- Hättenschwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition in terrestrial ecosystems. An. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Peay, K.G.; Kennedy, P.G.; Talbot, J.M. Dimensions of biodiversity in the Earth mycobiome. Nat. Rev. Microbiol. 2016, 14, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Zak, D.R.; Pregitzer, K.S.; Burton, A.J.; Edwards, I.P.; Kellner, H. Microbial responses to a changing environment: Implications for the future functioning of terrestrial ecosystems. Fungal Ecol. 2011, 4, 386–395. [Google Scholar] [CrossRef]
- Osono, T. Diversity and functioning of fungi associated with leaf litter decomposition in Asian forests of different climatic regions. Fungal Ecol. 2011, 4, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Peršoh, D. Plant-associated fungal communities in the light of meta’omics. Fungal Divers. 2015, 75, 1–25. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Talbot, J.M.; Bruns, T.D.; Taylor, J.W.; Smith, D.P.; Branco, S.; Glassman, S.I.; Erlandson, S.; Vilgalys, R.; Liao, H.-L.; Smith, M.E.; et al. Endemism and functional convergence across the North American soil mycobiome. Proc. Natl. Acad. Sci. USA 2014, 111, 6341–6346. [Google Scholar] [CrossRef] [PubMed]
- Nixon, K.C. Infrageneric classification of Quercus (Fagaceae) and typification of sectional names. Ann. Sci. For. 1993, 50, 25–34. [Google Scholar] [CrossRef]
- Kremer, A.; Casasoli, M.; Barreneche, T.; Bodénès, C.; Sisco, P.; Kubisiak, T.; Scalfi, M.; Leonardi, S.; Bakker, S.; Buiteveld, J.; et al. Fagaceae Trees. In Forest Trees, 1st ed.; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 7, pp. 161–187. [Google Scholar]
- Nixon, K.C. Global and Neotropical distribution and diversity of Oak (genus Quercus) and Oak forests. In Ecology and Conservation of Neotropical Montane Oak Forests; Kappelle, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 3–13. [Google Scholar]
- Aldrich, P.R.; Cavender-Bares, J. Quercus. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 89–129. [Google Scholar]
- Rzedowski, J. Vegetación de México, 1st ed.; Limusa: Distrito Federal, México, 1978; 504p. [Google Scholar]
- Gómez-Luna, B.E.; Rivera-Mosqueda, M.C.; Dendooven, L.; Vázquez-Marrufo, G.; Olalde-Portugal, V. Charcoal production at kiln sites affects C and N dynamics and associated soil microorganisms in Quercus spp temperate forests of central Mexico. Appl. Soil Ecol. 2009, 41, 50–58. [Google Scholar] [CrossRef]
- Aguilar, R.; Guilardi, A.; Vega, E.; Skutsch, M.; Oyama, K. Sprouting productivity and allometric relationships of two oak species managed for traditional charcoal making in central Mexico. Biomass Bioenergy 2012, 36, 192–207. [Google Scholar] [CrossRef]
- Chapa-Vargas, L.; Monzalvo-Santos, K. Natural protected areas of San Luis Potosí, Mexico: Ecological representativeness, risks, and conservation implications across scales. Int. J. Geogr. Inf. Sci. 2012, 26, 1625–1641. [Google Scholar] [CrossRef]
- Asbjornsen, H.; Ashton, M.S.; Vogt, D.J.; Palacios, S. Effects of habitat fragmentation on the buffering capacity of edge environments in a seasonally dry tropical oak forest ecosystem in Oaxaca, Mexico. Agric. Ecosyst. Environ. 2004, 103, 481–495. [Google Scholar] [CrossRef]
- Galicia, L.; Zarco-Arista, A.E.; Mendoza-Robles, K.I.; Palacio-Prieto, J.L.; García-Romero, A. Land use/cover, landforms and fragmentation patterns in a tropical dry forest in the southern Pacific region of Mexico. Singapore J. Trop. Geogr. 2008, 29, 137–154. [Google Scholar] [CrossRef]
- García-Oliva, F.; Covaleda, S.; Gallardo, J.F.; Prat, C.; Velázquez-Durán, R.; Etchevers, J.D. Firewood extraction affects carbon pools and nutrients in remnant fragments of temperate forest at the Mexican Transvolcanic Belt. Bosque 2014, 35, 311–324. [Google Scholar] [CrossRef]
- Rodríguez-Trejo, D.A.; Myers, R.L. Using oak characteristics to guide fire regime restoration in Mexican pine-oak and oak forests. Ecol. Restor. 2010, 28, 304–323. [Google Scholar] [CrossRef]
- Bugalho, M.N.; Caldeira, M.C.; Pereira, J.S.; Aronson, J.; Pausas, J.G. Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front. Ecol. Environ. 2011, 9, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Oliver, T.H.; Morecroft, M.D. Interactions between climate change and land use change on biodiversity: Attribution problems, risks, and opportunities. WIREs Clim. Chang. 2014, 5, 317–335. [Google Scholar] [CrossRef] [Green Version]
- Morris, S.J.; Friese, C.F.; Allen, M.F. Disturbance in natural ecosystems: Scaling from fungal diversity to ecosystem functioning. In Environmental and Microbial Relationships; Mycota, I.V., Druzhinina, I.S., Kubicek, C.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 79–98. [Google Scholar]
- Chávez-Vergara, B.; Rosales-Castillo, A.; Merino, A.; Vázquez-Marrufo, G.; Oyama, K.; García-Oliva, F. Quercus species control nutrients dynamics by determining the composition and activity of the forest floor fungal community. Soil Biol. Biochem. 2016, 98, 186–195. [Google Scholar] [CrossRef]
- Morris, M.H.; Pérez-Pérez, M.A.; Smith, M.E.; Bledsoe, C.S. Multiple species of ectomycorrhizal fungi are frequently detected on individual oak root tips in a tropical cloud forest. Mycorrhiza 2008, 18, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Van der Wal, A.; Geydan, T.D.; Kuyper, T.W.; de Boer, W. A thready affair: Linking fungal diversity and community dynamics to terrestrial decomposition processes. FEMS Microbiol. Rev. 2013, 37, 477–494. [Google Scholar] [CrossRef] [PubMed]
- Azul, A.M.; Sousa, J.P.; Agerer, R.; Martín, M.P.; Freitas, H. Land use practices and ectomycorrhizal fungal communities from oak woodlands dominated by Quercus suber L. considering drought scenarios. Mycorrhiza 2010, 20, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Hallgren, S.W.; Wilson, G.W.T. Frequency of prescribed burning in an upland oak forest determines soil and litter properties and alters the soil microbial community. For. Ecol. Manag. 2012, 265, 241–247. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Trivedi, C.; Hu, H.; Anderson, I.C.; Jeffries, T.C.; Zhou, J.; Singh, B.K. Microbial regulation of the soil carbon cycle: Evidence from gene–enzyme relationships. ISME J. 2016, 10, 2593–2604. [Google Scholar] [CrossRef] [PubMed]
- Avis, P.G.; Gaswick, W.C.; Tonkovich, G.S.; Leacock, P.R. Monitoring fungi in ecological restorations of coastal Indiana, USA. Restor. Ecol. 2017, 25, 92–100. [Google Scholar] [CrossRef]
- Purahong, W.; Kapturska, D.; Pecyna, M.J.; Schloter, M.; Buscot, F.; Hofrichter, M.; Krüger, D. Influence of different forest system management practices on leaf litter decomposition rates, nutrient dynamics and the activity of ligninolytic enzymes: a case study from Central European forests. PLoS ONE 2014, 9, e93700. [Google Scholar] [CrossRef] [PubMed]
- Flores-Rentería, D.; Rincón, A.; Valladares, F.; Yuste, J.C. Agricultural matrix affects differently the alpha and beta structural and functional diversity of soil microbial communities in a fragmented Mediterranean holm oak forest. Soil Biol. Biochem. 2016, 92, 79–90. [Google Scholar] [CrossRef]
- Cornejo-Tenorio, G.; Sánchez-García, E.; Flores-Tolentino, M.; Santana-Michel, F.; Ibarra-Manríquez, G. Flora y vegetación del cerro El Águila, Michoacán, México. Bot. Sci. 2013, 91, 155–180. [Google Scholar]
- Klooster, D. Forest transitions in Mexico: Institutions and forests in a globalized countryside. Prof. Geogr. 2003, 55, 227–237. [Google Scholar] [CrossRef]
- Works, M.A.; Hadley, K.S. The cultural context of forest degradation in adjacent Purépechan communities, Michoacán, Mexico. Geogr. J. 2004, 170, 22–38. [Google Scholar] [CrossRef]
- Castillo-Santiago, M.Á.; Ghilardi, A.; Oyama, K.; Hernández-Stefanoni, J.L.; Torres, I.; Flamenco-Sandoval, A.; Fernández, A.; Mas, J.F. Estimating the spatial distribution of woody biomass suitable for charcoal making from remote sensing and geostatistics in central Mexico. Energy Sust. Dev. 2013, 17, 177–188. [Google Scholar] [CrossRef]
- López, E.; Bocco, G.; Mendoza, M.; Velázquez, A.; Aguirre-Rivera, J.R. Peasant emigration and land-use change at the watershed level: A GIS-based approach in central Mexico. Agric. Syst. 2006, 90, 62–78. [Google Scholar] [CrossRef]
- Gentry, A.H. Diversity and floristic composition of Neotropical dry forests. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: Cambridge, CA, USA, 1995; pp. 146–194. [Google Scholar]
- Chávez-Vergara, B.; Merino, A.; Vázquez-Marrufo, G.; García-Oliva, F. Organic matter dynamics and microbial activity during decomposition of forest floor under two native Neotropical oak species in a temperate deciduous forest in Mexico. Geoderma 2014, 235, 133–145. [Google Scholar] [CrossRef]
- Bremmer, J.M. Nitrogen-Total. In Methods of Soil Analyses Part 3: Chemical Analyses; Spark, D.L., Page, A.L., Summer, M.E., Tabatabai, M.A., Helmke, P.A., Eds.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Huffman, E.W.D. Performance of a new automatic carbon dioxide coulometer. Microchem. J. 1977, 22, 567–573. [Google Scholar] [CrossRef]
- Parham, J.A.; Deng, S.P. Detection, quantification and characterization of β-glucosaminidase activity in soil. Soil Biol. Biochem. 2000, 32, 1183–1190. [Google Scholar] [CrossRef]
- Nagai, M.; Sato, T.; Watanabe, H.; Saito, K.; Kawata, M.; Enei, H. Purification and characterization of an extracellular laccase from the edible mushroom Lentinula edodes and decolorization of chemically different dyes. Appl. Microbiol. Biotechnol. 2002, 60, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Tien, M.; Kirk, T.K. Lignin peroxidase of Phanerochaete chrysosporium. Methods Enzymol. 1988, 161, 238–249. [Google Scholar] [CrossRef]
- Martinez, M.J.; Ruiz-Duenas, F.J.; Cuillen, F.; Martinez, A.T. Purification and catalytic properties of two manganese peroxidase isoenzymes from Pleurotus eryngii. Eur. J. Biochem. 1996, 237, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Von Ende, C.N. Repeated measures analysis: growth and other time-dependent measures. In Design and Analysis of Ecological Experiments; Scheiner, S.M., Gurevitch, J., Eds.; Chapman & Hall: New York, NY, USA, 1993; pp. 113–137. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innins, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Oxford, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular cloning. In A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; p. 2100. [Google Scholar]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Handelsman, J. Introducing DOTUR, a computer program for defining operational taxonomic units and estimating species richness. Appl. Environ. Microbiol. 2005, 71, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Magurran, A.E. Ecological Diversity and Its Measurement; Chapman and Hall: London, UK, 1996; p. 192. [Google Scholar]
- Chao, A. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Cao, L.; Zhang, R. Bacterial and fungal taxon changes in soil microbial community composition induced by short-term biochar amendment in red oxidized loam soil. World J. Microbiol. Biotechol. 2014, 30, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Handelsman, J. Introducing SONS, a tool for OTU-based comparisons of membership and structure between microbial communities. Appl. Environ. Microbiol. 2006, 72, 6773–6779. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Hamady, M.; Knight, R. UniFrac an online tool for comparing microbial community diversity in a phylogenetic context. BMC Bioinformatics 2006, 7, 371. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Coq, S.; Barantal, S.; Handa, I.T. Leaf traits and decomposition in tropical rainforests: revisiting some commonly held views and towards a new hypothesis. New Phytol. 2011, 189, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Bonanomi, G.; Cesarano, G.; Gaglione, S.A.; Ippolito, F.; Sarker, T.; Rao, M.A. Soil fertility promotes decomposition rate of nutrient poor, but not nutrient rich litter through nitrogen transfer. Plant Soil 2017, 412, 397–411. [Google Scholar] [CrossRef]
- Kubartová, A.; Ranger, J.; Berthelin, J.; Beguiristain, T. Diversity and decomposing ability of saprophytic fungi from temperate forest litter. Microb. Ecol. 2009, 58, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Salamanca, E.F.; Kaneko, N.; Katagiri, S.; Nagayama, Y. Nutrient dynamics and lignocellulose degradation in decomposing Quercus serrata leaf litter. Ecol. Res. 1998, 13, 199–210. [Google Scholar] [CrossRef]
- Gessner, M.O.; Swan, C.M.; Dang, C.K.; McKie, B.G.; Bardgett, R.D.; Wall, D.H.; Hättenschwiler, S. Diversity meets decomposition. Trends Ecol. Evol. 2010, 25, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Sheffer, E.; Canham, C.D.; Kigel, J.; Perevolotsky, A. Countervailing effects on pine and oak leaf litter decomposition in human-altered Mediterranean ecosystems. Oecologia 2015, 177, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Arslan, H.; Guleryuz, G.; Kırmızı, S. Nitrogen mineralisation in the soil of indigenous oak and pine plantation forests in a Mediterranean environment. Eur. J. Soil Biol. 2010, 46, 11–17. [Google Scholar] [CrossRef]
- Barba, J.; Lloret, F.; Yuste, J.C. Effects of drought-induced forest die-off on litter decomposition. Plant Soil 2016, 402, 91–101. [Google Scholar] [CrossRef]
- Rothstein, D.E.; Vitousek, P.M.; Simmons, B.L. An exotic tree alters decomposition and nutrient cycling in a Hawaiian montane forest. Ecosystems 2004, 7, 805–814. [Google Scholar] [CrossRef]
- Ceccon, E.; Sánchez, I.; Powers, J.S. Biological potential of four indigenous tree species from seasonally dry tropical forest for soil restoration. Agrofor. Syst. 2015, 9, 455–467. [Google Scholar] [CrossRef]
- Voříšková, J.; Brabcová, V.; Cajthaml, T.; Baldrian, P. Seasonal dynamics of fungal communities in a temperate oak forest soil. New Phytol. 2014, 201, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Criquet, S.; Farnet, A.M.; Tagger, S.; Le Petit, J. Annual variations of phenoloxidase activities in an evergreen oak litter: Influence of certain biotic and abiotic factors. Soil Biol. Biochem. 2000, 32, 1505–1513. [Google Scholar] [CrossRef]
- Kellner, H.; Luis, P.; Pecyna, M.J.; Barbi, F.; Kapturska, D.; Krüger, D.; Zak, D.R.; Marmeisse, R.; Vandenbol, M.; Hofrichter, M. Widespread occurrence of expressed fungal secretory peroxidases in forest soils. PLoS ONE 2014, 9, e95557. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Šnajdr, J.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Valášková, V. Responses of the extracellular enzyme activities in hardwood forest to soil temperature and seasonality and the potential effects of climate change. Soil Biol. Biochem. 2013, 56, 60–68. [Google Scholar] [CrossRef]
- Papa, S.; Pellegrino, A.; Fioretto, A. Microbial activity and quality changes during decomposition of Quercus ilex leaf litter in three Mediterranean woods. Appl. Soil Ecol. 2008, 40, 401–410. [Google Scholar] [CrossRef]
- Voříšková, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J.P.; Schaeffer, S.M. Microbial control over carbon cycling in soil. Front. Microbiol. 2012, 3, 348. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Pande, K.; Upadhyay, V.P.; Singh, J.S. Fungal communities associated with the decomposition of a common leaf litter (Quercus leucotrichophora A Camus) along an elevational transect in the Central Himalaya. Biol. Fert. Soils 1990, 9, 245–251. [Google Scholar] [CrossRef]
- Sadaka, N.; Ponge, J.F. Fungal colonization of phyllosphere and litter of Quercus rotundifolia Lam in a holm oak forest (High Atlas, Morocco). Biol. Fertil. Soils 2003, 39, 30–36. [Google Scholar] [CrossRef]
- Shirouzu, T.; Hirose, D.; Fukasawa, Y.; Tokumasu, S. Fungal succession associated with the decay of leaves of an evergreen oak, Quercus myrsinaefolia. Fungal Divers. 2009, 34, 87–109. [Google Scholar]
- Baldrian, P.; Voříšková, J.; Dobiášová, P.; Merhautová, V.; Lisá, L.; Valášková, V. Production of extracellular enzymes and degradation of biopolymers by saprotrophic microfungi from the upper layers of forest soil. Plant Soil 2011, 338, 111–125. [Google Scholar] [CrossRef]
- Schneider, T.; Keiblinger, K.M.; Schmid, E.; Sterflinger-Gleixner, K.; Ellersdorfer, G.; Roschitzki, B.; Richter, A.; Eberl, L.; Zechmeister-Boltenstern, S.; Riedel, K. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. ISME J. 2012, 6, 1749–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosca, E.; Montecchio, L.; Scattolin, L.; Garbaye, J. Enzymatic activities of three ectomycorrhizal types of Quercus robur L in relation to tree decline and thinning. Soil Biol. Biochem. 2007, 39, 2897–2904. [Google Scholar] [CrossRef]
- Floudas, D.; Binder, M.; Riley, R.; Barry, K.; Blanchette, R.A.; Henrissat, B.; Martínez, A.T.; Otillar, R.; Spatafora, J.W.; Yadav, J.S.; et al. The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bödeker, I.; Clemmensen, K.E.; Boer, W.; Martin, F.; Olson, Å.; Lindahl, B.D. Ectomycorrhizal Cortinarius species participate in enzymatic oxidation of humus in northern forest ecosystems. New Phytol. 2014, 203, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.; Nicolás, C.; Bentzer, J.; Ellström, M.; Smits, M.; Rineau, F.; Canbäck, B.; Floudas, D.; Carleer, R.; Lackner, G.; et al. Ectomycorrhizal fungi decompose soil organic matter using oxidative mechanisms adapted from saprotrophic ancestors. New Phytol. 2016, 209, 1705–1719. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi–potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Richard, F.; Moreau, P.A.; Selosse, M.A.; Gardes, M. Diversity and fruiting patterns of ectomycorrhizal and saprobic fungi in an old-growth Mediterranean forest dominated by Quercus ilex L. Can. J. Bot. 2004, 82, 1711–1729. [Google Scholar] [CrossRef]
- Azul, A.M.; Castro, P.; Sousa, J.P.; Freitas, H. Diversity and fruiting patterns of ectomycorrhizal and saprobic fungi as indicators of land-use severity in managed woodlands dominated by Quercus suber-a case study from southern Portugal. Can. J. For. Res. 2009, 39, 2404–2417. [Google Scholar] [CrossRef]
- Persha, L.; Agrawal, A.; Chhatre, A. Social and ecological synergy: local rulemaking, forest livelihoods, and biodiversity conservation. Science 2011, 331, 1606–1608. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Nemergut, D.; Knight, R.; Craine, J.M. Changes through time: Integrating microorganisms into the study of succession. Res. Microbiol. 2010, 161, 635–642. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coordinates | Well Preserved (WP) | Moderately Disturbed (MD) | Heavily Disturbed (HD) | |
|---|---|---|---|---|
| 19°32′13.20″ N, 101°17′60.00″ W | 19°32′18.24″ N, 101°17′56.40″ W | 19°32′6.00″ N, 101°18′3.60″ W | ||
| Stand characteristics | ||||
| Number of Quercus deserticola Trel. trees | 171 | 154 | 39 | |
| Mean tree DBH ± standard error (cm) | 12.1 ± 0.3 | 12.3 ± 0.25 | 15.1 ± 0.6 | |
| Aboveground biomass (Mg ha−1) | 42.7 | 46.3 | 27.4 | |
| Mean litter mass ± standard error (Mg ha−1) | 1.5 ± 0.25 | 1.0 ± 0.15 | 1.1 ± 0.1 | |
| Temperature on sampling dates (°C) 1 | 34.1, 25.6 | 34.3, 26.1 | 35, 26.4 | |
| Nutrient concentrations | ||||
| Litter 2 | ||||
| pH | 5.9–6.1 | 6.0–6.3 | 6.1–6.2 | |
| Carbon (mg g−1) | 417 | 391 | 473 | |
| Nitrogen (mg g−1) | 10.3 | 8.9 | 10.4 | |
| Phosphorus (mg g−1) | 0.34 | 0.42 | 0.54 | |
| C:N | 40 | 44 | 45 | |
| C:P | 1227 | 935 | 876 | |
| N:P | 30 | 21 | 19 | |
| Soil 2 | ||||
| Carbon (mg g−1) | 42.2 | 58.5 | 51.3 | |
| Nitrogen (mg g−1) | 3.4 | 2.33 | 2.5 | |
| Phosphorus (mg g−1) | 0.36 | 0.17 | 0.55 | |
| C:N | 12 | 25 | 20 | |
| C:P | 132 | 344 | 93 | |
| N:P | 11 | 14 | 4 | |
| Enzyme | Between Subjects | Within Subjects | |
|---|---|---|---|
| Site | Sampling Season | Interaction (Site:Season) | |
| Laccase | 0.28 (0.75) | 106.42 (<0.0001) | 0.12 (0.88) |
| Manganese peroxidase | 1.65 (0.23) | 24.37 (<0.0001) | 2.42 (0.13) |
| Lignin peroxidase | 3.55 (0.061) | 52.73 (<0.0001) | 1.76 (0.21) |
| Well Preserved (WP) | Moderately Disturbed (MD) | Heavily Disturbed (HD) | ||||
|---|---|---|---|---|---|---|
| Dry | Rainy | Dry | Rainy | Dry | Rainy | |
| No. of OTUs | 58 | 56 | 54 | 49 | 43 | 45 |
| No. of singletons | 33 | 30 | 30 | 19 | 18 | 19 |
| Sequences per group | ||||||
| Ascomycota | 145 | 116 | 113 | 124 | 83 | 133 |
| Basidiomycota | 17 | 29 | 21 | 37 | 25 | 28 |
| Unidentified | 16 | 19 | 39 | 12 | 49 | 7 |
| Shannon-Weaver (H’) | 3.39 | 3.51 | 3.16 | 3.24 | 3.16 | 3.12 |
| Simpson (1-D) | 0.96 | 0.93 | 0.94 | 0.93 | 0.90 | 0.92 |
| Chao1 | 100 | 103 | 98 | 61 | 60 | 60 |
| WP-W | MD-D | MD-W | HD-D | HD-W | |
|---|---|---|---|---|---|
| WP-D | 0.253 | 0.352 | 0.099 | 0.296 | 0.104 |
| WP-W | 0.160 | 0.301 | 0.133 | 0.304 | |
| MD-D | 0.102 | 0.308 | 0.096 | ||
| MD-W | 0.106 | 0.667 | |||
| HD-D | 0.099 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosales-Castillo, J.A.; Oyama, K.; Vázquez-Garcidueñas, M.S.; Aguilar-Romero, R.; García-Oliva, F.; Vázquez-Marrufo, G. Fungal Community and Ligninolytic Enzyme Activities in Quercus deserticola Trel. Litter from Forest Fragments with Increasing Levels of Disturbance. Forests 2018, 9, 11. https://doi.org/10.3390/f9010011
Rosales-Castillo JA, Oyama K, Vázquez-Garcidueñas MS, Aguilar-Romero R, García-Oliva F, Vázquez-Marrufo G. Fungal Community and Ligninolytic Enzyme Activities in Quercus deserticola Trel. Litter from Forest Fragments with Increasing Levels of Disturbance. Forests. 2018; 9(1):11. https://doi.org/10.3390/f9010011
Chicago/Turabian StyleRosales-Castillo, Jesús A., Ken Oyama, Ma. Soledad Vázquez-Garcidueñas, Rafael Aguilar-Romero, Felipe García-Oliva, and Gerardo Vázquez-Marrufo. 2018. "Fungal Community and Ligninolytic Enzyme Activities in Quercus deserticola Trel. Litter from Forest Fragments with Increasing Levels of Disturbance" Forests 9, no. 1: 11. https://doi.org/10.3390/f9010011