Physical Properties of Seeds of Eleven Spruce Species


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Physical Properties
2.3. Statistical Analysis
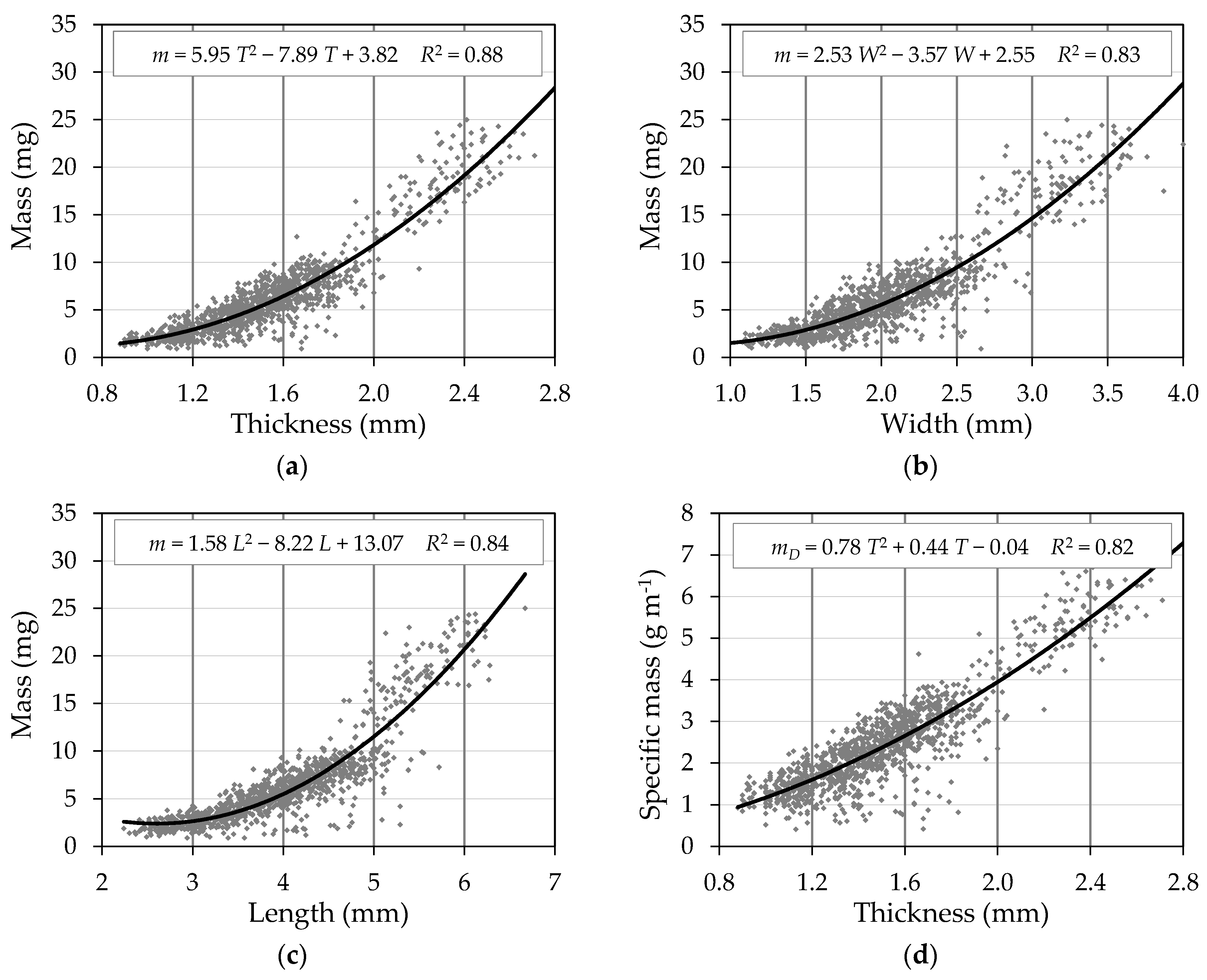
3. Results
3.1. Experimental Material
- for terminal velocity—0.25 m s−1,
- for basic dimensions (thickness, width and length)—0.1 mm,
- for the angle of external friction—1.2°,
- for seed mass—0.7 mg.
3.2. Correlations between Seed Properties
4. Discussion
- effective seedling production through the rational use of space allocated to different seeds, which is a very important consideration in container nurseries;
- rational use of seeding material with minimal loss of potentially germinable seeds;
- environmental benefits—the genetic potential of seeding material is not depleted (some trees produce seeds whose size differs from the reference range for the species).
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jonson, O.; More, D. Przewodnik Collinsa Drzewa (Collins Tree Guide); Multico Oficyna Wydawnicza: Warszawa, Poland, 2009; pp. 100–115. ISBN 9788370736439. (In Polish) [Google Scholar]
- Houston Durrant, T.; Mauri, A.; de Rigo, D.; Caudullo, G. Picea sitchensis in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Off. EU: Luxembourg, 2016. [Google Scholar]
- Uniyal, S.K.; Uniyal, A. Climate change and large-scale degradation of spruce: common pattern across the globe. Clim. Res. 2009, 38, 261–263. [Google Scholar] [CrossRef]
- Jia, Z.R.; Wang, J.H.; Zhang, S.G. Pollen morphology and its phylogenetic implications in the genus Picea. Plant Syst. Evol. 2014, 300, 461–473. [Google Scholar] [CrossRef]
- Ghimire, B.; Lee, C.; Yang, J.; Heo, K. Comparative leaf anatomy of some species of Abies and Picea (Pinaceae). Acta Bot. Bras. 2015, 29, 346–353. [Google Scholar] [CrossRef]
- Li, L.; Sun, Y.; Zou, J.; Yue, W.; Wang, X.; Liu, J. Origin and speciation of Picea schrenkiana and Picea smithiana in the Center Asian Highlands and Himalayas. Plant Mol. Biol. Rep. 2015, 33, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Savill, P.; Wilson, S.; Mason, B.; Jinks, R.; Stokes, V.; Christian, T. Alternative spruces to Sitka and Norway. Part 2—Oriental or Caucasian spruce (Picea orientalis), and the american and asian spruces. Q. J. For. 2017, 111, 88–97. [Google Scholar]
- Croser, C.; Renault, S.; Franklin, J.; Zwiazek, J. The effect of salinity on the emergence and seedling growth of Picea mariana, Picea glauca and Pinus banksiana. Environ. Pollut. 2001, 115, 9–16. [Google Scholar] [CrossRef]
- Li, Y.; Stocks, M.; Hemmilä, S.; Källman, T.; Zhu, H.; Zhou, Y.; Chen, J.; Liu, J.; Lascoux, M. Demographic histories of four spruce (Picea) species of the Qinghai-Tibetan Plateau and neighboring areas inferred from multiple nuclear loci. Mol. Biol. Evol. 2010, 27, 1001–1014. [Google Scholar] [CrossRef] [PubMed]
- Brus, D.J.; Hengeveld, G.M.; Walvoort, D.J.J.; Goedhart, P.W.; Heidema, A.H.; Nabuurs, G.J.; Gunia, K. Statistical mapping of tree species over Europe. Eur. J. For. Res. 2012, 131, 145–157. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea abies in Europe: distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Off. EU: Luxembourg, 2016. [Google Scholar]
- Drössler, L.; Fahlvik, N.; Wysocka, N.K.; Hjelm, K.; Kuehne, C. Natural regeneration in a multi-layered Pinus sylvestris-Picea abies forest after target diameter harvest and soil scarification. Forests 2017, 8, 35. [Google Scholar] [CrossRef]
- Nieuwenhuis, M.A.; Wills, J.M. The effect of cultivation technique on root architecture of young Sitka spruce (Picea sitchensis (Bong.) Carr.) trees on surface water gleys. New For. 2002, 24, 195–213. [Google Scholar] [CrossRef]
- Li, C.; Liu, S.; Berninger, F. Picea seedlings show apparent acclimation to drought with increasing altitude in the eastern Himalaya. Trees 2004, 18, 277–283. [Google Scholar] [CrossRef]
- Dumais, D.; Prévost, M. Management for red spruce conservation in Québec: The importance of some physiological and ecological characteristics—A review. For. Chron. 2007, 83, 378–392. [Google Scholar] [CrossRef]
- Lamontagne, J.M.; Boutin, S. Local-scale synchrony and variability in mast seed production patterns of Picea glauca. J. Ecol. 2007, 95, 991–1000. [Google Scholar] [CrossRef]
- Shibata, E.; Saito, M.; Tanaka, M. Deer-proof fence prevents regeneration of Picea jezoensis var. hondoensis through seed predation by increased woodmouse populations. J. For. Res. 2008, 13, 89–95. [Google Scholar] [CrossRef]
- Mckenzie, A.J.; Petty, S.J.; Toms, M.P.; Furness, R.W. Importance of Sitka spruce Picea sitchensis seed and garden bird-feeders for Siskins Carduelis spinus and Coal Tits Periparus ater. Bird Study 2010, 54, 236–247. [Google Scholar] [CrossRef]
- Dumais, D.; Prévost, M. Germination and establishment of natural red spruce (Picea rubens) seedlings in silvicultural gaps of different sizes. For. Chron. 2016, 92, 90–100. [Google Scholar] [CrossRef]
- Dylewski, L. Ecology and evolution of interaction between seed predators and conifers. Edukacja Ekologiczna 2016, 4, 3–8. (In Polish) [Google Scholar]
- Oleksyn, J.; Modrzyński, J.; Tjoelker, M.G.; Żytkowiak, R.; Reich, P.B.; Karolewski, P. Growth and physiology of Picea abies populations from elevational transects: Common garden evidence for altitudinal ecotypes and cold adaptation. Funct. Ecol. 1998, 12, 573–590. [Google Scholar] [CrossRef]
- Mosseler, A.; Major, J.E.; Simpson, J.D.; Daigle, B.; Lange, K.; Park, Y.-S.; Johnsen, K.H.; Rajora, O.P. Indicators of population viability in red spruce, Picea rubens. I. Reproductive traits and fecundity. Can. J. Bot. 2000, 78, 928–940. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, X.; Gu, W. Biogeographic differences in cone, needle and seed morphology among natural Picea asperata populations in Western China. For. Stud. China 2005, 7, 1–6. [Google Scholar] [CrossRef]
- Liu, G.F.; Zang, R.G.; Liu, H.; Bai, Z.Q.; Guo, Z.J.; Ding, Y. Geographic variation of seed morphological traits of Picea schrenkiana var. tianschanica in Tianshan Mountains, Xinjiang of Northwest China. Chin. J. Appl. Ecol. 2012, 23, 1455–1461. [Google Scholar]
- Grochowicz, J. Maszyny do Czyszczenia i Sortowania Nasion (Seed Cleaning and Sorting Machines); Akademia Rolnicza: Lublin, Poland, 1994; pp. 25–28. ISBN 839016129X. (In Polish) [Google Scholar]
- Rawat, B.S.; Uniyal, A.K. Variability in cone and seed characteristics and seed testing in various provenances of Himalayan spruce (Picea smithiana). J. For. Res. 2011, 22, 603–610. [Google Scholar] [CrossRef]
- Załęski, A. Nasiennictwo Leśnych Drzew I Krzewów Iglastych (Management of Coniferous Forest Trees and Shrubs for Seed Production); Oficyna Edytorska Wydawnictwo Świat: Warszawa, Poland, 1995; pp. 117–121. ISBN 8385597271. (In Polish) [Google Scholar]
- Khan, M.L. Effects of seed mass on seedling success in Artocarpus heterophyllus L., a tropical tree species of north-east India. Acta Oecol. 2004, 25, 103–110. [Google Scholar] [CrossRef]
- Parker, W.C.; Noland, T.L.; Morneault, A.E. The effects of seed mass on germination, seedling emergence, and early seedling growth of eastern white pine (Pinus strobus L.). New For. 2006, 32, 33–49. [Google Scholar] [CrossRef]
- Jones, T.A.; Reekie, E.G. Effect of seed size on seedling growth response to elevated CO2 in Picea abies and Picea rubens. Plant Biol. 2007, 9, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Upadhaya, K.; Pandey, H.N.; Law, P.S. The effect of seed mass on germination, seedling survival and growth in Prunus jenkinsii Hook.f. & Thoms. Turk. J. Bot. 2007, 31, 31–36. [Google Scholar] [CrossRef]
- Barbour, J.R.; Carvaiho, J.P.F. Response of Rocky Mountain juniper (Juniperus scopulorum) seeds to seed conditioning and germination treatments. Seed Technol. 2009, 31, 43–54. [Google Scholar]
- Norden, N.; Daws, M.I.; Antoine, C.; Gonzalez, M.A.; Garwood, N.C.; Chave, J. The relationship between seed mass and mean time to germination for 1037 tree species across five tropical forests. Funct. Ecol. 2009, 23, 203–210. [Google Scholar] [CrossRef]
- Mtambalika, K.; Munthali, C.; Gondwe, D.; Missanjo, E. Effect of seed size of Afzelia quanzensis on germination and seedling growth. Int. J. For. Res. 2014. [Google Scholar] [CrossRef]
- Chaisurisri, K.; Edwards, D.G.W.; El-Kassaby, Y.A. Effects of seed size on seedling attributes in Sitka spruce. New For. 1994, 8, 81–87. [Google Scholar]
- Kaliniewicz, Z.; Grabowski, A.; Liszewski, A.; Fura, S. Analysis of correlations between selected physical attributes of Scots pine seeds. Tech. Sci. 2011, 14, 13–22. [Google Scholar]
- Kaliniewicz, Z.; Markowski, P.; Anders, A.; Jadwisieńczak, B.; Rawa, T.; Szczechowicz, D. Basic physical properties of Norway spruce (Picea abies (L.) Karst.) seeds. Tech. Sci. 2016, 19, 103–115. [Google Scholar]
- Kaliniewicz, Z.; Poznański, A. Variability and correlation of selected physical attributes of small-leaved lime (Tilia cordata Mill.) seeds. Sylwan 2013, 157, 39–46. (In Polish) [Google Scholar]
- Mohsenin, N.N. Physical Properties of Plant and Animal Materials; Gordon and Breach Science Public: New York, NY, USA, 1986; pp. 1–891. ISBN 9780677213705. [Google Scholar]
- Kaliniewicz, Z. Analysis of frictional properties of cereal seed. Afr. J. Agric. Res. 2013, 8, 5611–5621. [Google Scholar] [CrossRef]
- Rabiej, M. Statystyka z Programem Statistica (Statisctics in Statistica Software); Helion: Gliwice, Poland, 2012; pp. 1–344. ISBN 9788324641109. (In Polish) [Google Scholar]
- Kluczyński, B. Yielding and quality of Norway spruce (Picea abies (L.) Karst) seeds in dependence on the part of crown and chosen biological and site-related characteristics of trees. Sylwan 1992, 5, 25–35. (In Polish) [Google Scholar]
- Oleksyn, J.; Reich, P.B.; Tjoelker, M.G.; Chalupka, W. Biogeographic differences in shoot elongation pattern among European Scots pine populations. For. Ecol. Manag. 2001, 148, 207–220. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Latitude, seed predation and seed mass. J. Biogeogr. 2003, 30, 105–128. [Google Scholar] [CrossRef]
- Mezquida, E.T.; Benkman, C.W. The geographic selection mosaic for squirrels, crossbills and Aleppo pine. J. Evol. Biol. 2005, 18, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Tylek, P. Separation of fir, larch and spruce seeds based on seed mass. Prz. Tech. Rol. Leśn. 1999, 3, 17–19. (In Polish) [Google Scholar]
- Załęski, A. The effect of different methods of Norway spruce seed extraction on their vitality and predisposition to long-term storage. For. Res. Pap. 2003, 1, 7–22. (In Polish) [Google Scholar]
- Yehnjong, P.S.; Zavada, M.S.; Liu, C. Characterization and ecological significance of a seed bank from the Upper Pennsylvanian Wise Formation, southwest Virginia. Acta Palaeobot. 2017, 57, 165–175. [Google Scholar] [CrossRef]
- Kaliniewicz, Z.; Markowski, P.; Anders, A.; Tylek, P.; Krzysiak, Z.; Fura, S. Influence of the age of parent stand on selected physical properties of Norway spruce seeds. Sylwan 2017, 161, 548–557. (In Polish) [Google Scholar]
- Mikola, J. The effect of seed size and duration of growth on the height of Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karst.) provenances and progenies at the nursery stage. Silva Fenn. 1980, 14, 84–94. [Google Scholar] [CrossRef]
- Kaliniewicz, Z.; Tylek, P.; Markowski, P.; Anders, A.; Rawa, T.; Zadrożny, M. Determination of shape factors and volume coefficients of seeds from selected coniferous trees. Tech. Sci. 2012, 15, 217–228. [Google Scholar]
- Pike, C.C.; Warren, J.C.; Montgomery, R.A. Allometry of early growth in selected and wild sources of white spruce, Picea glauca (Moench) Voss. New For. 2016, 47, 131–141. [Google Scholar] [CrossRef]
- Colas, F.; Lamhamedi, M.S. Production of a new generation of seeds through the use of somatic clones in controlled crosses of black spruce (Picea mariana). New For. 2014, 45, 1–20. [Google Scholar] [CrossRef]
- Mabille, F.; Abecassis, J. Parametric modelling of wheat grain morphology: a new perspective. J. Cereal Sci. 2003, 37, 43–53. [Google Scholar] [CrossRef]
- Markowski, M.; Żuk-Gołaszewska, K.; Kwiatkowski, D. Influence of variety on selected physical and mechanical properties of wheat. Ind. Crop. Prod. 2013, 47, 113–117. [Google Scholar] [CrossRef]
- Kaliniewicz, Z.; Anders, A.; Markowski, P.; Jadwisieńczak, K.; Rawa, T. Influence of cereal seed orientation on external friction coefficients. Trans. ASABE 2016, 59, 1073–1081. [Google Scholar] [CrossRef]
- Tylek, P. Friction and elasticity as separation properties of beech nuts. Sylwan 2006, 5, 51–58. [Google Scholar]
- Kaliniewicz, Z.; Mańkowski, S.; Tylek, P.; Krzysiak, Z.; Peda, W. Correlations between the physical properties of silver fir seeds. Acta Agroph. 2018, 25, 197–212. (In Polish) [Google Scholar] [CrossRef]
- Tylek, P. Problems of pneumatic selection of forest tree seeds. Sylwan 1999, 12, 65–72. (In Polish) [Google Scholar]
- Tylek, P. Analysis of aerodynamic properties of common fir and commmon beech. Inż. Rol. 2011, 6, 247–253. (In Polish) [Google Scholar]
- Kaliniewicz, Z.; Tylek, P.; Anders, A.; Markowski, P.; Rawa, T.; Ołdakowski, M.; Wąsowski, L. An analysis of the physical properties of seeds of selected deciduous tree species. Balt. For. 2016, 22, 169–174. [Google Scholar]



{kind=link}
{kind=link}
| Spruce Species | Property | |||||
|---|---|---|---|---|---|---|
| Terminal Velocity v (m s−1) | Thickness T (mm) | Width W (mm) | Length L (mm) | Angle of External Friction α (°) | Mass m (mg) | |
| Blue spruce | 6.1 ± 0.5 c | 1.44 ± 0.13 c | 1.77 ± 0.18 c | 3.70 ± 0.34 d | 27 ± 4 c | 4.2 ± 0.9 c |
| Jezo spruce | 5.8 ± 1.1 b | 1.49 ± 0.18 d | 1.98 ± 0.27 d | 3.91 ± 0.41 e | 29 ± 6 d,e | 4.8 ± 2.0 d |
| Lijiang spruce | 6.4 ± 0.9 d | 1.63 ± 0.20 f | 2.13 ± 0.28 e | 4.46 ± 0.42 g | 29 ± 4 d,e | 7.0 ± 2.0 f,g |
| Meyer’s spruce | 6.5 ± 0.7 d | 1.53 ± 0.18 e | 2.00 ± 0.26 d | 3.92 ± 0.42 e | 29 ± 5 d,e | 6.1 ± 1.7 e |
| Morinda spruce | 8.3 ± 0.6 g | 2.32 ± 0.18 g | 3.19 ± 0.29 g | 5.52 ± 0.44 h | 23 ± 2 a | 18.6 ± 3.1 h |
| Norway spruce | 7.6 ± 0.9 f | 1.54 ± 0.20 e | 2.19 ± 0.26 f | 4.23 ± 0.48 f | 29 ± 5 e | 6.9 ± 2.3 f,g |
| Oriental spruce | 6.9 ± 0.7 e | 1.63 ± 0.16 f | 2.08 ± 0.26 e | 3.98 ± 0.38 e | 26 ± 4 b | 6.5 ± 1.8 e,f |
| Red spruce | 5.7 ± 0.7 b | 1.21 ± 0.12 b | 1.61 ± 0.20 b | 3.15 ± 0.29 c | 29 ± 5 d,e | 3.0 ± 0.8 b |
| Schrenk’s spruce | 6.6 ± 0.9 d | 1.63 ± 0.16 f | 2.24 ± 0.27 f | 4.54 ± 0.42 g | 28 ± 5 c,d | 7.4 ± 1.9 g |
| Sitka spruce | 5.3 ± 0.4 a | 1.10 ± 0.11 a | 1.47 ± 0.23 a | 2.97 ± 0.24 b | 30 ± 5 e | 2.3 ± 0.5 a |
| White spruce | 5.79 ± 0.6 b | 1.21 ±0.14 b | 1.43 ± 0.17 a | 2.76 ± 0.22 a | 29 ± 5 d,e | 2.4 ± 0.6 a |
| Spruce Species | Indicator | |||||
|---|---|---|---|---|---|---|
| Geometric Mean Diameter D (mm) | Aspect Ratio T/W (%) | Aspect Ratio T/L (%) | Aspect Ratio W/L (%) | Sphericity Index Φ (%) | Specific Mass mD (g m−1) | |
| Blue spruce | 2.11 ± 0.15 c | 81.81 ± 8.50 f | 39.17 ± 3.93 c | 48.24 ± 5.74 a,b | 57.27 ± 3.71 b,c | 1.99 ± 0.37 c |
| Jezo spruce | 2.25 ± 0.20 d | 76.16 ± 11.29 c,d | 38.19 ± 4.60 b,c | 50.84 ± 7.17 c,d | 57.72 ± 3.92 b,c,d | 2.10 ± 0.76 c |
| Lijiang spruce | 2.48 ± 0.23 f | 77.52 ± 10.98 d,e | 36.72 ± 4.71 a | 47.79 ± 5.63 a | 55.83 ± 3.76 a | 2.77 ± 0.62 d,e |
| Meyer’s spruce | 2.28 ± 0.21 d | 77.40 ± 10.59 d,e | 39.27 ± 4.34 c | 51.24 ± 5.79 c,d | 58.46 ± 3.41 d | 2.64 ± 0.53 d |
| Morinda spruce | 3.44 ± 0.21 h | 73.06 ± 7.59 a,b | 42.08 ± 3.53 d | 57.98 ± 5.65 e | 62.39 ± 3.14 g | 5.38 ± 0.64 f |
| Norway spruce | 2.42 ± 0.26 e | 70.60 ± 7.79 a | 36.44 ± 2.75 a | 51.98 ± 4.53 d | 57.35 ± 2.31 b,c | 2.79 ± 0.63 d,e |
| Oriental spruce | 2.38 ± 0.21 e | 79.07 ± 9.64 e | 41.17 ± 3.83 d | 52.58 ± 6.03 d | 59.92 ± 3.40 e | 2.70 ± 0.56 d |
| Red spruce | 1.83 ± 0.14 b | 75.75 ± 9.22b c,d | 38.66 ± 4.44 c | 51.63 ± 7.56 d | 58.29 ± 4.53 c,d | 1.61 ± 0.39 b |
| Schrenk’s spruce | 2.54 ± 0.19 g | 73.49 ± 10.74 b,c | 35.99 ± 3.89 a | 49.63 ± 6.43 b,c | 56.17 ± 3.50 a | 2.89 ± 0.64 e |
| Sitka spruce | 1.68 ± 0.12 a | 76.12 ± 11.20 c,d | 37.12 ± 4.17 a,b | 49.62 ± 8.11 b,c | 56.70 ± 4.21 a,b | 1.36 ± 0.25 a |
| White spruce | 1.68 ± 0.14 a | 85.29 ± 9.21 g | 43.95 ± 5.07 e | 51.85 ± 6.00 d | 60.95 ± 4.09 f | 1.40 ± 0.31 a |
| Spruce Species | Coefficient of Correlation between Seed Mass m and: | ||||
|---|---|---|---|---|---|
| Terminal Velocity v | Thickness T | Width W | Length L | Angle of External Friction α | |
| Blue spruce | 0.610 * | 0.538 * | 0.459 * | 0.516 * | 0.101 |
| Jezo spruce | 0.866 * | 0.573 * | 0.264 * | 0.455 * | −0.594 * |
| Lijiang spruce | 0.765 * | 0.658 * | 0.653 * | 0.597 * | −0.320 * |
| Meyer’s spruce | 0.617 * | 0.747 * | 0.575 * | 0.798 * | −0.488 * |
| Morinda spruce | 0.418 * | 0.688 * | 0.529 * | 0.753 * | 0.045 |
| Norway spruce | 0.654 * | 0.881 * | 0.759 * | 0.920 * | −0.190 * |
| Oriental spruce | 0.782 * | 0.748 * | 0.640 * | 0.778 * | −0.302 * |
| Red spruce | 0.533 * | 0.479 * | 0.354 * | 0.453 * | −0.164 |
| Schrenk’s spruce | 0.783 * | 0.503 * | 0.387 * | 0.434 * | −0.167 |
| Sitka spruce | 0.515 * | 0.592 * | 0.349 * | 0.476 * | −0.138 |
| White spruce | 0.374 * | 0.613 * | 0.443 * | 0.599 * | −0.149 |
| Spruce Species | Seed Fraction | Percentage (%) | Coefficient of Variation (%) of Seed Mass | |
|---|---|---|---|---|
| Fraction | Total | |||
| Blue spruce | I (T ≤ 1.40 mm) | 39.1 | 21.31 | 22.08 |
| II (T = 1.41 ÷ 1.50 mm) | 33.9 | 14.43 | ||
| III (T > 1.50 mm) | 27.0 | 18.31 | ||
| Jezo spruce | I (T ≤ 1.40 mm) | 30.1 | 38.80 | 41.15 |
| II (T = 1.41 ÷ 1.55 mm | 34.4 | 30.93 | ||
| III (T > 1.55 mm) | 35.5 | 38.26 | ||
| Lijiang spruce | I (T ≤ 1.55 mm) | 30.4 | 26.53 | 28.46 |
| II (T = 1.56 ÷ 1.75 mm) | 41.9 | 23.76 | ||
| III (T > 1.75 mm) | 27.7 | 20.11 | ||
| Meyer’s spruce | I (T ≤ 1.40 mm) | 26.7 | 24.37 | 27.61 |
| II (T = 1.41 ÷ 1.60 mm) | 40.6 | 18.61 | ||
| III (T > 1.60 mm) | 32.7 | 20.73 | ||
| Morinda spruce | I (T ≤ 2.25 mm) | 35.5 | 14.34 | 16.73 |
| II (T = 2.26 ÷ 2.40 mm) | 34.6 | 13.11 | ||
| III (T > 2.40 mm) | 29.9 | 10.32 | ||
| Norway spruce | I (T ≤ 1.45 mm) | 34.5 | 19.73 | 33.61 |
| II (T = 1.46 ÷ 1.60 mm) | 35.3 | 15.26 | ||
| III (T > 1.60 mm) | 30.2 | 23.54 | ||
| Oriental spruce | I (T ≤ 1.55 mm) | 26.6 | 26.01 | 27.55 |
| II (T = 1.56 ÷ 1.70 mm) | 40.7 | 20.41 | ||
| III (T > 1.70 mm) | 32.7 | 16.02 | ||
| Red spruce | I (T ≤ 1.15 mm) | 33.6 | 26.13 | 28.00 |
| II (T = 1.16 ÷ 1.25 mm) | 30.0 | 23.12 | ||
| III (T > 1.25 mm) | 36.4 | 23.37 | ||
| Schrenk’s spruce | I (T ≤ 1.55 mm) | 29.1 | 23.12 | 25.59 |
| II (T = 1.55 ÷ 1.70 mm) | 40.9 | 18.33 | ||
| III (T > 1.70 mm) | 30.0 | 23.39 | ||
| Sitka spruce | I (T ≤ 1.05 mm) | 35.8 | 19.94 | 22.40 |
| II (T = 1.06 ÷ 1.15 mm) | 35.8 | 17.92 | ||
| III (T > 1.15 mm) | 28.4 | 16.63 | ||
| White spruce | I (T ≤ 1.15 mm) | 32.7 | 25.12 | 26.68 |
| II (T = 1.16 ÷ 1.25 mm) | 32.7 | 22.08 | ||
| III (T > 1.25 mm) | 34.6 | 18.82 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaliniewicz, Z.; Żuk, Z.; Kusińska, E. Physical Properties of Seeds of Eleven Spruce Species. Forests 2018, 9, 617. https://doi.org/10.3390/f9100617
Kaliniewicz Z, Żuk Z, Kusińska E. Physical Properties of Seeds of Eleven Spruce Species. Forests. 2018; 9(10):617. https://doi.org/10.3390/f9100617
Chicago/Turabian StyleKaliniewicz, Zdzisław, Zbigniew Żuk, and Elżbieta Kusińska. 2018. "Physical Properties of Seeds of Eleven Spruce Species" Forests 9, no. 10: 617. https://doi.org/10.3390/f9100617
APA StyleKaliniewicz, Z., Żuk, Z., & Kusińska, E. (2018). Physical Properties of Seeds of Eleven Spruce Species. Forests, 9(10), 617. https://doi.org/10.3390/f9100617