Effects of Douglas Fir Stand Age on Soil Chemical Properties, Nutrient Dynamics, and Enzyme Activity: A Case Study in Northern Apennines, Italy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Soil Bulk Density and Physicochemical Analyses
2.3. Soil Organic Matter Properties
2.4. Soil Enzyme Activities and Functional Diversity
- β-glucosidase to chitinase + leucine aminopeptidase ratio (BG/(NAG + LAP)), as an indicator of the nitrogen limitation with respect to carbon [12];
- Chitinase + leucine aminopeptidase to acid phosphatase ratio (NAG + LAP)/PHOS), as an indicator of the phosphorus limitation with respect to nitrogen [41];
2.5. Foliar Nutrients
2.6. Statistical Analysis
3. Results
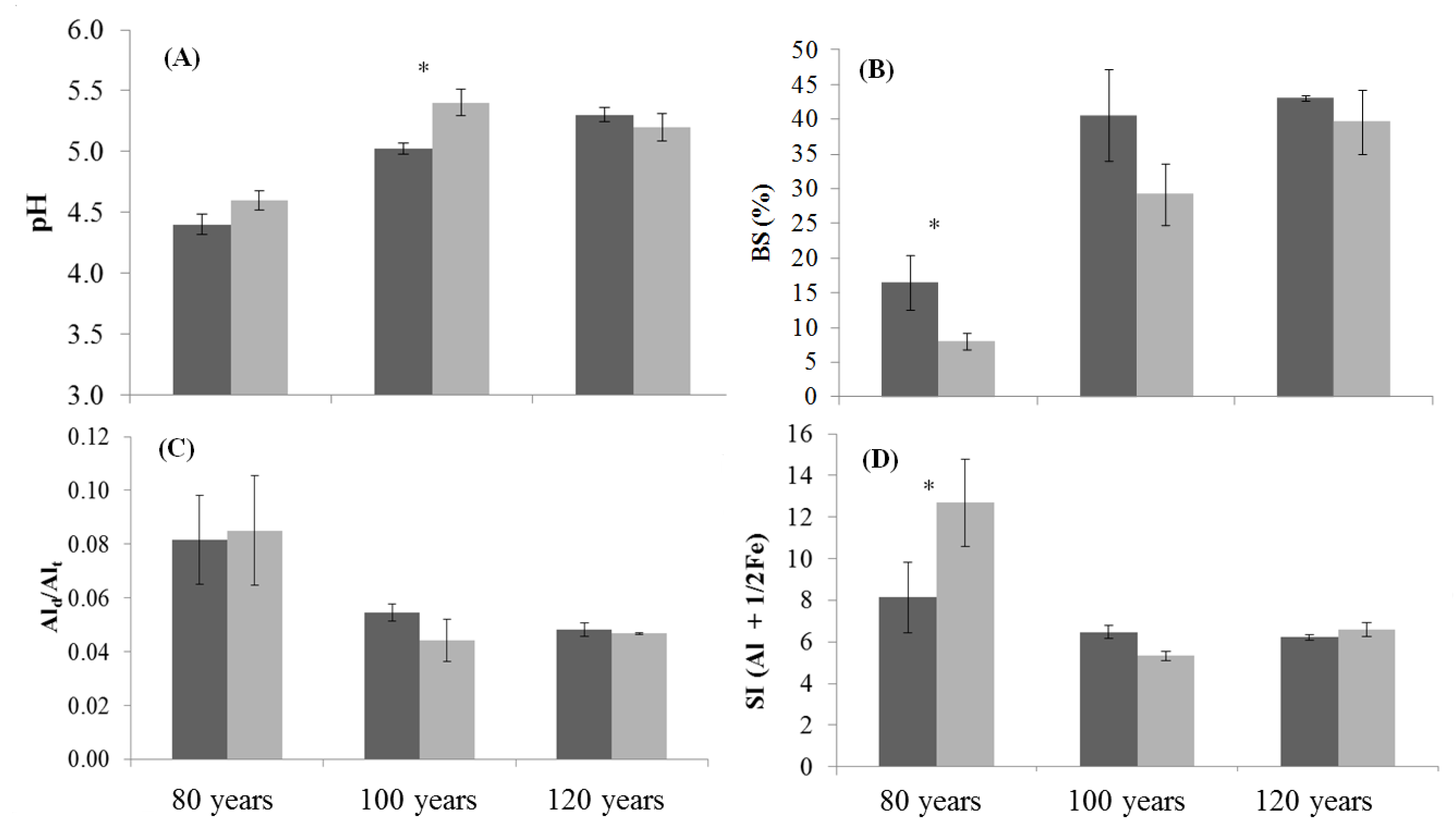
3.1. Soil Properties of Stand Age Classes
3.2. Soil Carbon and Nitrogen Stocks
3.3. Foliar Nutrients
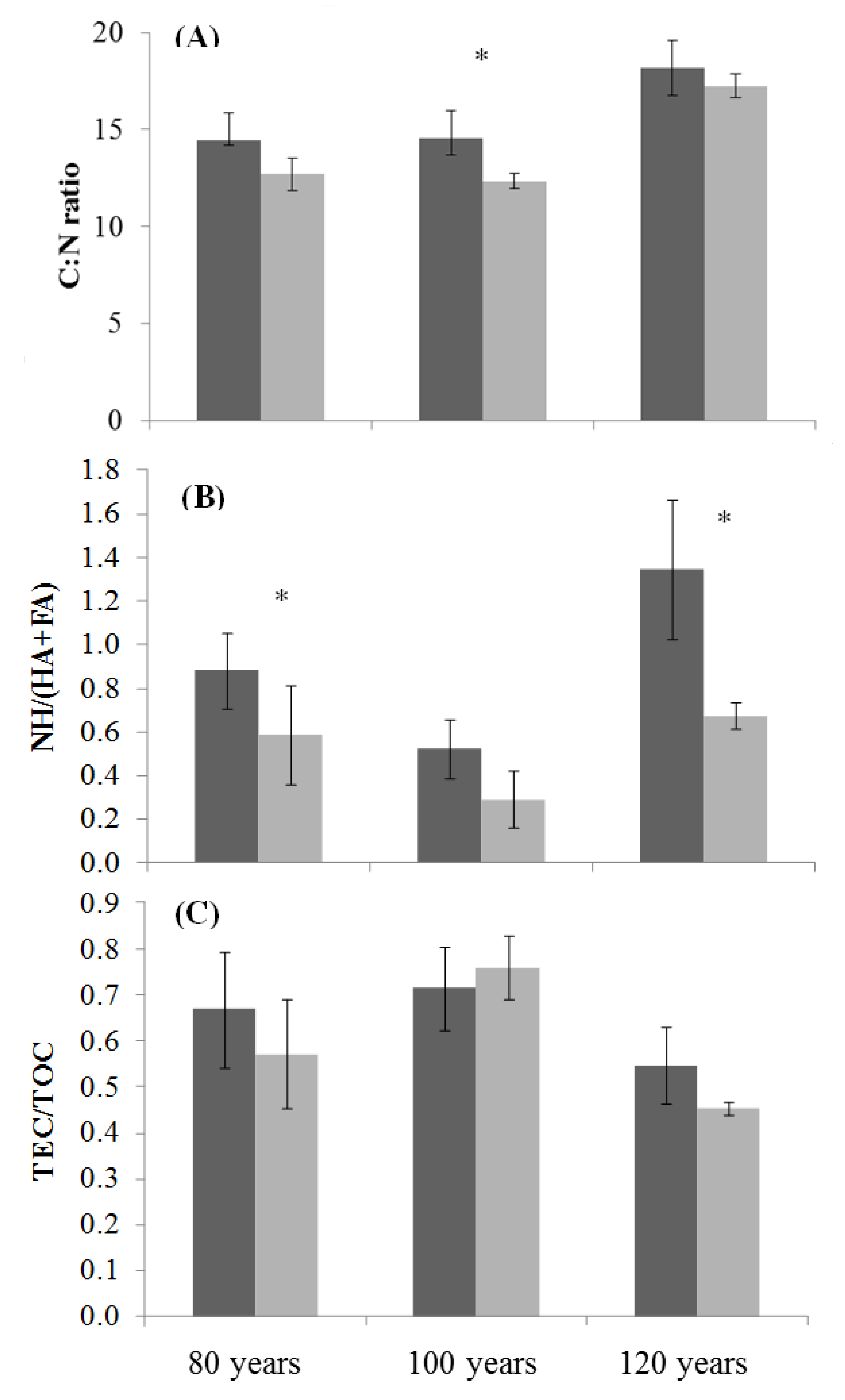
3.4. Soil Organic Matter Properties
3.5. Soil Enzyme Activity and Functional Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Watson, R.T.; Noble, I.R.; Bolin, B.; Ravin-Dranath, N.H.; Verando, D.J.; Dokken, D.J. Land Use, Land Use Change and Forestry; Special Report of the Intergovernmental Panel on Climate Change: Cambridge, UK, 2000. [Google Scholar]
- Lal, R. Soil carbon sequestration impacts on global climate change and food security. Science 2004, 304, 1623–1626. [Google Scholar] [CrossRef] [PubMed]
- Aubert, M.; Bureau, F.; Alard, D.; Bardat, J. Effect of tree mixture on the humic epipedon and vegetation diversity in managed beech forests (Normandy, France). Can. J. For. Res. 2004, 34, 233–248. [Google Scholar] [CrossRef]
- Hedde, M.; Aubert, M.; Decaëns, T.; Bureau, F. Dynamics of soil carbon in a beechwood chronosequence forest. For. Ecol. Manag. 2008, 255, 193–202. [Google Scholar] [CrossRef]
- Pickett, S.T.A. Space-for-time substitutions as an alternative to long-term studies. In Long-Term Studies in Ecology; Likens, G.E., Ed.; Springer: New York, NY, USA, 1989; pp. 110–135. [Google Scholar]
- Corona, P.; Scotti, R.; Tarchiani, N. Relationship between environmental factors and site index in Douglas –fir plantations in central Italy. For. Ecol. Manag. 1998, 110, 195–207. [Google Scholar] [CrossRef]
- Pavari, A.; de Philippis, A. La sperimentazione di specie forestali esotiche in Italia: La sperimentazione del primo ventennio. In Annali della Sperimentazione Agraria; Tip. Failli: Roma, Italy, 1941; Volume 38, p. 648. [Google Scholar]
- Gabbrielli, A. Aldo Pavari sperimentatore. (L’Italia Forestale e Montana) Ital. J. For. Mount. Environ. 2010, 66, 411–420. [Google Scholar] [CrossRef]
- Antisari, L.V.; Falsone, G.; Carbone, S.; Marinari, S.; Vianello, G. Douglas-fir reforestation in North Apennine (Italy): Performance on soil carbon sequestration, nutrients stock and microbial activity. Appl. Soil Ecol. 2015, 86, 82–90. [Google Scholar] [CrossRef]
- Jaffrain, J.; Gérard, F.; Meyer, M.; Ranger, J. Assessing the quality of dissolved organic matter in forest soils using ultraviolet absorption spectrophotometry. Soil Sci. Soc. Am. J. 2007, 71, 1851–1858. [Google Scholar] [CrossRef]
- Allison, S.D.; Gartner, T.; Holland, K.; Weintraub, M.; Sinsabaugh, R.L. Soil Enzymes: Linking Proteomics and Ecological Processes. In Manual of Environmental Microbiology, 3rd ed.; Hurst, C.J., Knudsen, G.R., McInerney, M.J., Stetzenbach, L.D., Walter, M.V., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 704–711. [Google Scholar]
- Sinsabaugh, R.L.; Hill, B.; Shah, J.J.F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–799. [Google Scholar] [CrossRef] [PubMed]
- IUSS. Working Group World Reference Base for Soil Resources 2006; first update 2007; World Soil Resources Reports: Rome, Italy, 2007. [Google Scholar]
- Antisari, L.V.; Marinari, S.; Dell’Abate, M.T.; Baffi, C.; Vianello, G. Plant cover and epipedon SOM stability as factors affecting brown soil profile development and microbial activity. Geoderma 2011, 161, 212–224. [Google Scholar] [CrossRef]
- Welke, S.E.; Hope, G.D. Influences of stand composition and age on forest floor processes and chemistry in pure and mixed stands of Douglas-fir and paper birch in interior British Columbia. For. Ecol. Manag. 2005, 219, 29–42. [Google Scholar] [CrossRef]
- Blanco, J.A. Forests may need centuries to recover their original productivity after continuous intensive management: An example from Douglas-fir stands. Sci. Total Environ. 2012, 437, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Perry, D.A.; Molina, R.; Amaranthus, M.P. Mycorrhizae, mycorrhizospheres, and reforestation: Current knowledge and research needs. Can. J. For. Res. 1987, 17, 929–940. [Google Scholar] [CrossRef]
- Griffiths, R.P.; Baham, J.E.; Caldwell, B.A. Soil solution chemistry of ectomycorrhizal mats in forest soil. Soil Biol. Biochem. 1994, 26, 331–337. [Google Scholar] [CrossRef]
- Ranger, J.; Allie, S.; Gelhaye, D.; Pollier, B.; Turpault, M.P.; Granier, A. Nutrient budgets for a rotation of a Douglas-fir plantation in the Beaujolais (France) based on a chronosequence study. For. Ecol. Manag. 2002, 171, 3–16. [Google Scholar] [CrossRef]
- Marques, R.; Ranger, J.; Villette, S.; Granier, A. Nutrient dynamics in a chronosequence of Douglas-fir (Pseudotsuga menziesii (Mirb). Franco) stands on the Beaujolais Mounts (France). 2. Quantitative approach. For. Ecol. Manag. 1997, 92, 167–197. [Google Scholar] [CrossRef]
- Jussy, J.H.; Colin-Belgrand, M.; Ranger, J. Production and root uptake of mineral nitrogen in a chronosequence of Douglas-fir (Pseudotsuga menziesii) in the Beaujolais Mounts. For. Ecol. Manag. 2000, 128, 197–209. [Google Scholar] [CrossRef]
- Patrone, G. Piano di Assestamento delle Foreste di Vallombrosa e di S. Antonio per il Quindicennio 1970–1984; Ministero dell’Agricoltura e delle Foreste, Azienda di Stato per le Foreste Demaniali: Rome, Italy, 1970. [Google Scholar]
- Travaglini, D. Piano di Gestione e Silvomuseo 2006–2025 della Foresta di Vallombrosa: Compartimentazione e cartografia. Ann. dell’Accad. Italiana Sci. For. 2009, 58, 139–150. [Google Scholar]
- Pavari, A. Studio Preliminare sulle Colture di Specie Forestali Esotiche in Italia. In Annali del Regio Istituto Superiore Forestale Nazionale; Ricci: Firenze, Italy, 1916; Volume 1, pp. 159–379. [Google Scholar]
- Nocentini, S. Le specie forestali esotiche: La sperimenazione di Aldo Pavari e le prospettive attuali. L’Italia For. Mont. 2010, 65, 449–457. [Google Scholar] [CrossRef]
- Sanesi, G.; Certini, G. The umbric epipedon in the N Apennines, Italy—An example from the Vallombrosa Forest. J. Plant Nutr. Soil Sci. 2005, 168, 392–398. [Google Scholar] [CrossRef]
- Ferretti, F. Le prove di introduzione delle specie forestali esotiche: L’esperienza condotta a Vallombrosa. Primo Contrib. Ann. ISSA 1998, 29, 93–114. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle Size Analysis. In Methods of Soil Analysis; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; Volume 9, pp. 383–411. [Google Scholar]
- Orsini, L.; Rémy, J.C. Utilisaion du chlorure de cobaltihexamine pour la détermination simultanée de la capacité d’échange et des bases échangeables des sols. Sci. Sol. 1976, 4, 269–279. [Google Scholar]
- Ciesielski, H.; Sterckeman, T. Determination of CEC and exchangeable cations in soils by means of cobalt hexamine trichloride. Effects of experimental conditions. Agronomie 1997, 17, 1–8. [Google Scholar] [CrossRef]
- Schwertmann, U.; Fitzpatrick, R.W.; Taylor, R.M.; Lewis, D.G. The influence of aluminum on iron oxides. Part ii. Preparation and properties of A1-substituted hematites. Clays Clay Miner. 1979, 27, 105–112. [Google Scholar] [CrossRef]
- Mehra, O.P.; Jackson, M.L. Iron oxide removal from soils and clays by a dithionite-citrate system buffered with sodium bicarbonate. Clays Clay Miner. 1960, 7, 317–327. [Google Scholar] [CrossRef]
- Antisari, L.V.; Dell’Abate, M.T.; Buscaroli, A.; Gherardi, M.; Nisini, L.; Vianello, G. Role of soil organic matter characteristics in a pedological survey: “Bosco Frattona” natural reserve (Site of Community Importance, Italy) case study. Geoderma 2010, 156, 302–315. [Google Scholar] [CrossRef]
- Springer, U.; Klee, J. Prüfung der Leistungsfähigkeit von einigen wichtigeren Verfahren zur Bestimmung des Kohlenstoffs mittels Chromschwefelsäure sowie Vorschlag einer neuen Schnellmethode. Z. Pflanzenernähr. Düng. Bodenkd. 1954, 64, 1–26. [Google Scholar] [CrossRef]
- Sequi, P.; de Nobili, M.; Leita, L.; Cercignani, G. A new index of humification. Agrochimica 1986, 30, 175–179. [Google Scholar]
- McGinnis, M.L.; Holub, S.M.; Myrold, D.D. Regional assessment of soil microbial functional diversity of Douglas-fir forests. Soil Sci. Soc. Am. J. 2014, 78, 125–135. [Google Scholar] [CrossRef]
- Marx, M.C.; Wood, M.; Jarvis, S.C. A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol. Biochem. 2001, 33, 1633–1640. [Google Scholar] [CrossRef]
- Vepsalainen, M.; Kukkonen, S.; Vestberg, M.; Sirvio, H.; Niemi, R.M. Application of soil enzyme activity test kit in a field experiment. Soil Biol. Biochem. 2001, 33, 1665–1672. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef] [Green Version]
- Moscatelli, M.C.; Secondi, L.; Marabottini, R.; Papp, R.; Stazi, S.R.; Mania, E.; Marinari, S. Assessment of soil microbial functional diversity: Land use and soil properties affect CLPP-MicroResp and enzymes responses. Pedobiologia 2018, 66, 36–42. [Google Scholar] [CrossRef]
- Zeglin, L.H.; Bottomley, P.J.; Jumpponen, A.; Rice, C.W.; Arango, M.; Lindsley, A.; McGowan, A.; Mfombep, P.; MYrold, D.D. Altered precipitation regime affects the function and composition of soil microbial communities on multiple time scales. Ecology 2013, 94, 2334–2345. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.C.; Smith, K.L. Soil microbial diversity and the sustainability of agricultural soils. Plant Soil 1995, 170, 75–86. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef]
- Linder, S. Foliar analysis for detecting and correcting nutrient imbalances in Norway spruce. Ecol. Bull. 1995, 44, 178–190. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Schoeneberger, P.J.; Wysocki, D.A.; Benham, E.C. Field Book for Describing and Sampling Soils; Version 3.0; Natural Resources Conservation Service, National Soil Survey Center: Lincoln, NE, USA, 2012.
- Certini, G.; Agnelli, A.; Corti, G.; Capperucci, A. Composition and mean residence time of molecular weight fractions of organic matter extracted from two soils under different forest species. Biogeochemistry 2004, 71, 299–316. [Google Scholar] [CrossRef]
- Agnelli, A.; Trumbore, S.E.; Corti, G.; Ugolini, F.C. The dynamics of organic matter in soil rock fragments investigated by 14C dating and 13C measurements. Eur. J. Soil Sci. 2002, 53, 147–159. [Google Scholar] [CrossRef]
- Perry, D.A. Forest Ecology; Johns Hopkins University Press: Baltimore, MD, USA, 1994. [Google Scholar]
- Sinsabaugh, R.L.; Shah, J.J.F. Ecoenzymatic stoichiometry of recalcitrant organic matter decomposition: The growth rate hypothesis in reverse. Biogeochemistry 2011, 102, 31–43. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Shah, J.J.F. Ecoenzymatic stoichiometry and ecological theory. Ann. Rev. Ecol. Evol. Syst. 2012, 43, 313–343. [Google Scholar] [CrossRef]
- Wallenstein, M.; Allison, S.D.; Ernakovitch, J.; Steinweg, M.; Sinsabaugh, R.L. Controls on the temperature sensitivity of soil enzymes: A key driver of in situ enzyme activity rates. Soc. Biol. 2011, 22, 245–258. [Google Scholar] [CrossRef]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Marinari, S.; Bonifacio, E.; Moscatelli, M.C.; Antisari, L.V.; Vianello, G. Soil development and microbial functional diversity: Proposal for a methodological approach. Geoderma 2013, 192, 437–445. [Google Scholar] [CrossRef]
- Bonneau, M. Le diagnostic foliaire. Revue Forestičre Française 1988, 40, 19–26. [Google Scholar] [CrossRef]
- Kupka, I.; Podrázský, V.; Kubeček, J. Soil-forming effect of Douglas-fir at lower altitudes—A case study. J. For. Sci. 2013, 59, 345–351. [Google Scholar] [CrossRef]
- Egli, M.; Fitze, P.; Mirabella, A. Weathering and evolution of soils formed on granitic, glacial deposits: Results from chronosequences of Swiss alpine environments. Catena 2001, 45, 19–47. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Profile | Horizons | Boundary | Color Munsell | Structure | Texture | Consistence | Roots | Rock Fragments | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Master | Depth (cm) | D | T | dry | Moist | G | S | T | D | M | S | P | Q | S | S | V% | R | |||
| 80 | DOUG 8 | Oi | 4–2 | A | S | |||||||||||||||
| Oe/Oa | 2–0 | A | W | 10YR3/3 | 7.5YR3/2 | |||||||||||||||
| A1 | 0–4 | C | W | 10YR4/3 | 7.5YR3/3 | 1 | f | GR | l/cl | S | VFR | (w)s | (w)p | 1 | f | FGR | 2 | 2 | ||
| A2 | 4–12 | C | S | 10YR4/4 | 7.5YR3/3 | 1 | f | ABK | l | S | VFR | (w)ss | (w)ps | 2 | f/m | FGR | 3 | 2 | ||
| Bw | 12–20 | C | S | 10YR5/4 | 10YR4/4 | 1 | f | ABK | sl | SH | FR | (w)so | (w)ps | 0 | f | MGR | 3 | 1 | ||
| BC | 20–24 | D | I | 10YR6/4 | 10YR4/4 | 1 | m | ABK | sl | SH | FR | (w)ss | (w)ps | 0 | f | MGR | 4 | 1 | ||
| C | 24–40+ | U | 10YR6/4 | 10YR6/4 | 0 | m | SG | sl | MH | FI | (w)ss | (w)ps | MGR | 8 | 1 | |||||
| 100 | DOUG 10 | Oi | 3–2 | A | S | |||||||||||||||
| Oe | 2–0 | A | S | |||||||||||||||||
| A1 | 0–5 | C | W | 10YR4/3 | 7.5YR2.5/3 | 1 | f/m | SBK | l | S | VFR | (w)ss | (w)p | 1 | f/m | 0 | ||||
| A2 | 5–16/17 | C | W | 10YR4/4 | 7.5YR3/3 | 1 | f | SBK | l | S | FR | (w)so | (w)ps | 1 | f/m | FGR | 2 | 2 | ||
| Bw | 16/17–31 | C | S | 10YR5/4 | 7.5YR4/6 | 1 | f | ABK | sl | SH | FR | (w)so | (w)ps | 1 | m | MGR | 10 | 1 | ||
| C | 21–40+ | U | 10YR6/4 | 10YR4/6 | 0 | m | SG | sl | MH | FR | (w)ss | (w)p | CGR | 16 | 1 | |||||
| 120 | DOUG 12 | Oi | 5–4 | A | S | |||||||||||||||
| Oe | 4–2.5 | A | S | |||||||||||||||||
| Oa | 2.5–0 | A | S | 10YR3/2 | 5YR3/1 | |||||||||||||||
| A1 | 0–8 | A | W | 10YR4/3 | 5YR3/2 | 1 | f/m | GR | sl | S | VFR | (w)so | (w)ps | 3 | f/m | CGR | 7 | 2 | ||
| A2 | 8–25 | C | W | 10YR4/4 | 5YR4/4 | 1 | f | SBK | sl | S | VFR | (w)so | (w)ps | 2 | f | MGR | 8 | 2 | ||
| Bw | 25–31 | C | S | 10YR5/4 | 5YR6/4 | 1 | f/m | ABK | sl | SH | FR | (w)so | (w)ps | 0 | f | FGR | 7 | 1 | ||
| C | 31–60+ | U | 10YR6/4 | 7.5YR5/8 | 0 | m | SG | l | SH | FR | (w)so | (w)ps | ||||||||
| Stand Age (years) | Horizons | Depth | Year | pH | Corg | Sand | Silt | Clay | CEC | BS |
|---|---|---|---|---|---|---|---|---|---|---|
| cm | g kg−1 | g kg−1 | Cmol(+) kg−1 | % | ||||||
| 80 | A1 | 0–3 | 80 | 4.4 ± 1.2 | 47.7 ± 1.6 | 364 ± 23 | 359 ± 25 | 277 ± 21 | 16.0 ± 1.2 | 22.7 ± 0.3 |
| A2 | 3–12 | 80 | 4.2 ± 1.4 | 35.0 ± 2.0 | 407 ± 56 | 336 ± 21 | 257 ± 25 | 13.9 ± 0.9 | 16.8 ± 0.4 | |
| Bw | 12–20 | 80 | 4.7 ± 1.6 | 32.1 ± 1.1 | 721 ± 32 | 239 ± 16 | 40 ± 11 | 11.9 ± 1.1 | 12.0 ± 1.0 | |
| Bc | 20–24 | 80 | 4.4 ± 0.8 | 28.9 ± 1.5 | 692 ± 58 | 245 ± 19 | 63 ± 8 | 10.8 ± 0.5 | 8.6 ± 0.9 | |
| C | 24–40+ | 80 | 4.8 ± 0.8 | 27.8 ± 2.3 | 615 ± 63 | 314 ± 29 | 71 ± 10 | 9.1 ± 0.4 | 7.9 ± 0.5 | |
| 100 | A1 | 0–5 | 100 | 5.0 ± 0.9 | 53.4 ± 2.8 | 369 ± 36 | 420 ± 38 | 211 ± 15 | 17.9 ± 1.3 | 47.6 ± 1.2 |
| A2 | 5–16/17 | 100 | 5.1 ± 1.3 | 33.9 ± 1.9 | 361 ± 45 | 416 ± 35 | 223 ± 16 | 18.5 ± 1.5 | 46.6 ± 0.9 | |
| Bw | 16/17–31 | 100 | 5.5 ± 1.1 | 22.4 ± 1.4 | 604 ± 25 | 323 ± 24 | 73 ± 10 | 16.4 ± 0.9 | 34.1 ± 1.2 | |
| C | 21–40 | 100 | 5.6 ± 1.2 | 12.3 ± 2.1 | 716 ± 45 | 231 ± 21 | 53 ± 5 | 11.5 ± 0.7 | 37.0 ± 1.1 | |
| 120 | A1 | 0–8 | 120 | 5.2 ± 1.4 | 91.2 ± 1.8 | 620 ± 36 | 239 ± 21 | 141 ± 19 | 38.9 ± 1.1 | 43.8 ± 1.1 |
| A2 | 8–22 | 120 | 5.4 ± 1.5 | 34.7 ± 1.3 | 639 ± 72 | 209 ± 32 | 152 ± 11 | 26.4 ± 1.3 | 42.4 ± 0.4 | |
| Bw | 22–31 | 120 | 5.4 ± 1.2 | 43.9 ± 1.9 | 662 ± 34 | 217 ± 19 | 121 ± 9 | 19.5 ± 1.1 | 45.3 ± 0.5 | |
| C | 31–40+ | 120 | 5.0 ± 1.8 | 18.3 ± 2.3 | 473 ± 38 | 383 ± 22 | 144 ± 10 | 8.0 ± 0.9 | 34.1 ± 0.3 | |
| Stand Age (years) | Al | Fe | Mn | N | Al/N | Fe/N | Mn/N |
|---|---|---|---|---|---|---|---|
| µg g−1 | % | ||||||
| 80 | 268 ± 25 | 120 ± 11 | 249 ± 31 | 1.6 ± 0.1 | 144 ± 29 | 72 ± 8 | 112 ± 27 |
| 100 | 354 ± 13 | 158 ± 12 | 440 ± 20 | 1.4 ± 0.1 | 222 ± 53 | 107 ± 28 | 287 ± 24 |
| 120 | 545 ± 32 | 364 ± 17 | 285 ± 18 | 1.3 ± 0.1 | 372 ± 38 | 256 ± 25 | 195 ± 19 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vittori Antisari, L.; Papp, R.; Vianello, G.; Marinari, S. Effects of Douglas Fir Stand Age on Soil Chemical Properties, Nutrient Dynamics, and Enzyme Activity: A Case Study in Northern Apennines, Italy. Forests 2018, 9, 641. https://doi.org/10.3390/f9100641
Vittori Antisari L, Papp R, Vianello G, Marinari S. Effects of Douglas Fir Stand Age on Soil Chemical Properties, Nutrient Dynamics, and Enzyme Activity: A Case Study in Northern Apennines, Italy. Forests. 2018; 9(10):641. https://doi.org/10.3390/f9100641
Chicago/Turabian StyleVittori Antisari, Livia, Ruxandra Papp, Gilmo Vianello, and Sara Marinari. 2018. "Effects of Douglas Fir Stand Age on Soil Chemical Properties, Nutrient Dynamics, and Enzyme Activity: A Case Study in Northern Apennines, Italy" Forests 9, no. 10: 641. https://doi.org/10.3390/f9100641