Ecophysiological Responses of Calcicole Cyclobalanopsis glauca (Thunb.) Oerst. to Drought Stress and Calcium Supply
by
Liang Xue
1,2, 
Huadong Ren
1,2,
Wei Long
1,
Xiuhui Leng
1,
Jia Wang
1,2,
Xiaohua Yao
1,2 and
Sheng Li
1,2,* 1
Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Fuyang 311400, China
2
Observation and Research Station for Rock Desert Ecosystem, Anshun 562100, China
*
Author to whom correspondence should be addressed.
Forests 2018, 9(11), 667; https://doi.org/10.3390/f9110667
Submission received: 17 September 2018
/
Revised: 21 October 2018
/
Accepted: 23 October 2018
/
Published: 25 October 2018
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:Water deficit and high calcium (Ca2+) content and are two typical soil characteristics in the Karst region. However, the problem of whether high Ca2+ in Karst calcareous soil could increase drought tolerance in calcicole plants has not been solved. We investigated the ecophysiological responses of Cyclobalanopsis glauca (Thunb.) Oerst. cuttings to short-term drought stress and Ca2+ application. Drought stress (10% PEG-6000) markedly reduced relative water content (RWC) and water potential (WP), and enhanced the levels of reactive oxygen species (ROS) production (H2O2 and O2•−) and malondialdehyde (MDA) content in C. glauca leaves. Under drought treatment, exogenous Ca2+ application (20 mM CaCl2) markedly increased the RWC and WP, and reduced the H2O2, O2•−, and MDA content. Furthermore, water deficit induced a significant increase in the activities of antioxidant enzymes such as peroxidase (POD), superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR), and glutathione peroxidase (GPX), and increased the accumulation of osmoregulation substances. External Ca2+ alleviated drought-induced oxidative stress and osmotic stress with further increased activities of antioxidant enzymes, and enhanced the accumulation of osmoregulation substances. In addition, exogenous Ca2+ treatment alleviated the reduction of the photosynthesis rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), and chlorophyll content (SPAD), and further increased water use efficiency (WUE) under drought stress. This study confirms that exogenouos Ca2+ application induces improvements in the water status, osmotic adjustment, antioxidant defense, and photosynthesis efficiency of C. glauca under drought stress.
1. Introduction
The Karst region in southwest China is one of three distribution centers around the world [1]. Karst habitats are characterized by a double-layer hydrogeological structure and high porosity of the underlying limestone rock with serious surface water leakage, and very shallow and patchy soil with a low water retention capacity [2,3]. Rainfall rapidly disappears into subterranean streams through shafts, sinkholes, and dolines [4]. Previous studies showed that available soil water is only sufficient for plant transpiration needs for one or two weeks following saturated field capacity [5,6]. Drought stress has become one of the most constraining factors for plants in Karst habitats [4].
Plant ecophysiological characteristics might be changed by soil water deficit. Drought could induce a decrease in the relative water content (RWC) and water potential (WP) of plants [7]. Additionally, it can trigger the overproduction of reactive oxygen species (ROS), resulting in oxidative stress [8,9]. Water deficit can also reduce chlorophyll content, gas exchange parameters, stomatal conductance, and photosystem reaction center activity, resulting in the suppression of photosynthesis [10]. To cope with oxidative stress, plants have established antioxidant defense systems to eliminate ROS, including numerous antioxidant enzymes and non-enzymatic antioxidants [11]. Moreover, plants accumulate certain compatible solutes such as proline, soluble sugar, and glycine betaine to deal with drought stress [12]. Under moderate drought conditions, four Karst adapted plants with different growth forms enhanced water use efficiency (WUE), antioxidant enzymes activities, and osmoregulation substances, indicating the essential roles of these defense systems involved in drought resistance [13].
Calcium has been approved to ameliorate the adverse effects of water deficit on plants, which might be related to increased antioxidant enzyme activities and reduced membrane lipid peroxidation [14,15,16,17,18,19]. Moreover, Ca2+ application also improves plant water status by inducing more osmotic adjustment substance synthesis [15,18,20]. Additionally, it can improve stomatal movement and increase photosynthetic electron transport, thereby increasing WUE [21]. As a vital second messenger, Ca2+ also plays key roles in plant responses to drought stress [22,23,24].
Karst habitats are also characterized by the high soil calcium content of their limestone background. The calcium-rich environment reserves a lot of calcicole plants, which can live on limestone soil. However, whether the high exogenous calcium in the Karst environment could enhance the drought tolerance of calcicole plants remains to be proved. Furthermore, the possible ecophysiological mechanisms of Ca2+ involved in the drought tolerance of calcicole plants has not been illuminated. Cyclobalanopsis glauca (Thunb.) Oerst. (C. glauca) is an endemic calcicole tree species in the Karst regions of southwestern China. Because of its good ecological function and economic value, C. glauca has been used as a pioneer tree species for vegetation restoration in the Karst region. It has been reported that drought stress would damage C. glauca by aggravating oxidative stress, deteriorating water status, and inhibiting photosynthesis [25,26]. The aim of this study is to investigate the effects of exogenous Ca2+ on the tolerance of C. glauca to water deficit. We hypothesized that exogenous Ca2+ will improve water status, antioxidant enzyme activities, osmotic adjustment substances, and photosynthetic efficiency in C. glauca, enhancing its tolerance under drought stress.
2. Materials and Methods
2.1. Plant Materials and Stress Treatments
One-year-old cuttings of C. glauca were sampled from a twenty-eight-year-old seed bearer in a Karst habitat (24°47′26.6″ N, 106°33′7.81″ E) in Leye county, Guangxi Province, China. The long-term annual mean temperature of the sampling site was 16.5 °C (10 °C in January and 34 °C in July). The long-term annual mean precipitation was 1200 mm, 78.3% of which occurred from May to September. The soil type was black calcareous soil with a high calcium concentration. Cuttings were grown in a greenhouse where the temperature was 28/23 °C (day/night), the daily photoperiod was 16/8 h, and the relative humidity was about 65%. Six-month-old cuttings were subjected to different treatments in solutions containing 1/2 strength Hoagland solution at pH 6.0 as follows: drought (10% PEG-6000), drought with exogenous Ca2+ (10% PEG-6000 + 20 mM CaCl2), and the control. Each treatment and control consisted of three replicates and lasted for 0, 12, 24, 48, and 72 h, respectively. One fully expanded leaf per individual with a similar size was used for photosynthesis and water status determination per replicate. Moreover, two leaves per individual were mixed for other physiological analyses.
2.2. Leaf Relative Water Content and Water Potential
Leaf samples were collected immediately to measure their fresh weight (FW) and then submerged in distilled water for 24 h to acquire the turgid weight (TW). Subsequently, the samples were placed at a temperature of 70 °C for 72 h at constant weight to obtain the dry weight (DW). All the leaf samples were weighted using an electronic scale (0.001 g precision). The leaf RWC was calculated using the following formula: RWC (%) = ((FW − DW/TW − DW) × 100). An instantaneous leaf WP was measured at 10:00 a.m. using a WP4C Dewpoint Potential Meter (Decagon, Pullman, WA, USA).
2.3. Reactive Oxygen Species (ROS) and Malondialdehyde (MDA) Content Determination
Hydrogen peroxide (H2O2) content was determined according to Sergiev et al. [27]. 0.3 g of the fresh leaf samples were homogenized in an ice bath with 4 mL of 0.1% (w/v) trichloroacetic acid (TCA). The homogenate was centrifuged at 12,000 rpm for 15 min at 4 °C and 0.5 mL supernatant was added to 0.5 mL 50 mM potassium phosphate buffer (pH 6.8) and 1 mL 1 M potassium iodide. The absorbance of the supernatant was read at 560 nm. The H2O2 content was obtained using a standard curve.
Superoxide anion (O2•−) was measured through a method described by Meng et al. [28]. Frozen leaf samples of 0.5 g were homogenized in an ice bath with 5 mL 50 mM phosphate buffer (pH 7.8). The homogenate was centrifuged at 12,000 rpm for 15 min at 4 °C. The mixture made up of 0.5 mL supernatant and 0.5 mL phosphate buffer (pH 7.8), and 1 mL 1 mM hydroxyl ammonium chloride was incubated at 25 °C for 1 h. Then, 1 mL 17 mM p-amino benzene sulphonic acid and 1 mL 7 mM α-naphthylamine were added to the mixture and incubation was carried out at 25 °C for 20 min. Finally, the equivalent volume of ethyl ether was added to the mixture and then it was centrifuged at 1500 rpm for 5 min. The absorbance of the water phase was measured at 530 nm.
The MDA content was determined using the method of Cao et al. [29]. The extract was dissolved in 5 mL 10% TCA and centrifuged at 12,000 rpm for 10 min, and the supernatant was then transferred to a 5 mL centrifuge tube and diluted to 4 mL with 10% TCA. The 1 mL supernatant was mixed with 1 mL 20% TCA containing 0.5% (w/v) thiobarbituric acid (TBA). The mixture was heated in boiling water for 15 min and cooled immediately; the mixture was then centrifuged at 12,000 rpm for 10 min. The absorbance of the final supernatant was measured at 532 nm, 600 nm, and 450 nm. The MDA value was estimated by means of an extinction coefficent (155 mM−1 cm−1).
2.4. Measurement Osmoregulation Substances
The total soluble sugar was measured following the anthrone sulphuric acid method [30] using sucroseas standard. Briefly, 0.5 g smashed samples were placed in 15 mL conical tubes containing 10 mL distilled water. The anthrone reagent was prepared using 0.1 g anthrone dissolved in 50 mL 95% sulphuric acid. The absorbance of the reaction solution was measured at 620 nm using a spectrophotometer (UV-1800, Shimadzu, Kyoto, Japan) after a water bath set at 100 °C for 10 min. The free proline content was measured following the ninhydrin method [30]. Briefly, 0.2 g samples were smashed in 5 mL 3% (w/v) sulfosalicylic acid. The reaction mixture contained equal volumes of proline, ninhydrin reagent, and glacial acetic acid. The absorbance of the reaction solution was measured at 520 nm using a spectrophotometer (UV-1800, Shimadzu, Kyoto, Japan) after a water bath set at 100 °C for 1 h. Glycine betaine was determined using the periodide colorimetric method according to Grieve and Grattan [31].
2.5. Measurement of Antioxidant Enzyme Activities
For enzyme extracts and assays, 0.5 g frozen leaf samples were homogenized in 8 mL 50 mM cold potassium phosphate buffer (KPB) (pH 7.8) containing 0.1 mM EDTA and 1% (w/v) polyvinypolypyrrolidone phosphate buffer (pH 6) in a chilled mortar. The homogenate was then centrifuged at 12,000 prm for 20 min at 4 °C, and the resulting supernatant was used to assay the following enzyme activities and measured using a spectrophotometer (UV-1800, Shimadzu, Kyoto, Japan).
Superoxide dismutase (SOD) activity was determined using the nitroblue tetrazolium (NBT) method as described by Beauchamp and Fridovich [32]. The assay medium contained 50 mM potassium phosphate buffer (pH 7.8), 0.1 mM EDTA, 13 mM methionine, 75 µM NBT, and 2 µM riboflavin. The reaction mixture was placed under a 15 W fluorescent lamp with 120 µmol m−2 s−1 for 15 min. The absorbance was read at 560 nm.
Catalase (CAT) activity was assayed following the method described by Chance and Maehly [33], with some modification. The reaction mixture contained 50 mM KPB (pH 7.0), 200 mM H2O2, and enzyme extract. The decomposition of H2O2 was measured by the decline in absorbance at 240 nm for 1 min.
Peroxidase (POD) activity was determined using the method described by Upadhyaya et al. [34], with some modification. The reaction mixture contained 100 mM KPB (pH 7.0), 10 mM H2O2, 20 mM guaiacol, and 1 mL of the enzyme extract. Changes in the absorbance of the reaction solution at 470 nm were measured.
The activity of ascorbate peroxidase (APX) was determined as a decrease in absorbance at 290 nm for 1 min, with minor modification [35]. The reaction was started by adding 50 µL extraction enzyme, 1.25 mL 50 mM KPB (pH 7.8), 500 µL 2mM H2O2, and 200 µL ascorbic acid (ASA).
The activity of the glutathione reductase (GR) was determined using the method of Cakmak et al. [36]. The absorbance of the reaction solution was measured at 340 nm. The reaction mixture contained 1 mM ethylene diamine tetraacetic acid (EDTA), 0.5 mM glutathione disulfide (GSSG), 0.15 mM nicotinamide adenine dinucleotide phosphate (NADPH), 100 mM sodium phosphate buffer (pH 7.8), and 0.15 mL enzyme extract. The protein content was determined using Bradford’s [37] method.
Glutathione peroxidase (GPX) activity was assayed according to the method of Lawrence and Burk [38]. The reaction mixture contained 0.2 mM NADPH, 1 mM sodium azide (pH 7.0), 1 mM glutathione (GSH), 1 unit glutathione reductase and 2 mM H2O2 and enzyme extract. The reaction was started by adding H2O2 and the absorbance was measured at 340 nm.
2.6. Photosynthetic Parameters
Chlorophyll content was determined using a SPAD-502 chlorophyll meter (Minolta Co., Tokyo, Japan). The photosynthesis rate (Pn), stomatal conductance (Gs), and transpiration rate (Tr) were measured at 10:00 a.m. using an infrared gas analyzer LI-COR 6400 (LI-COR Inc., Lincoln, NE, USA). The measurements were implemented under certain conditions of CO2 concentration (380 ppm), photosynthetic photon flux density (PPFD) of 800 µmol photons m−2 s−1 at 25 °C. WUE was estimated using the ratio between net photosynthesis and the amount of water transpiration (Pn/Tr).
2.7. Statistical Analyses
Statistical analyses were performed with SPSS version19.0 (SPSS Inc., Chicago, IL, USA). Statistical differences between measurements for different treatments were tested using an analysis of one-way variance (ANOVA) followed by Duncan’s multiple range test. Values are presented as the means ± standard deviation (SD) of three replicate samples. Significant differences were determined at p < 0.05 levels.
3. Results
Plant leaves with PEG-6000 and Ca2+ treatment after 72 h are shown in Figure 1. Compared with the control, the drought-treated leaf margin appeared to be yellowing and showed milder symptoms when supplemented with Ca2+.
3.1. Relative Water Content (RWC) and Leaf Water Potential (WP)
Compared with the control, the leaf RWC was significantly decreased at 24 h for drought stress and 48 h for Ca2+ addition (p < 0.05), respectively (Figure 2a). Meanwhile, significant decreases in the leaf instantaneous WP were also observed for drought stress and Ca2+ addition, with the lowest value of −3.4 MPa and −2.67 MPa at 72 h, respectively (Figure 2b). Under drought condition, the Ca2+-treated plants had a higher RWC than the untreated one, with up to a 4.67% increase at 72 h (Figure 2a). Compared with the drought, significant higher WPs were also observed in the Ca2+-treated plants at all treatment times.
3.2. O2•− and H2O2 Generation and Lipid Peroxidation
H2O2 and O2•− are the two main forms of ROS produced under adverse conditions. Compared with the control, O2•− and H2O2 contents were significantly increased (p < 0.05) at 12 h in drought stress and 24 h in Ca2+-treated plants, respectively (Figure 3a,b). A sharp accumulation of O2•− (141.56%) and H2O2 (96.39%) over the control plants was found in drought conditions at 72 h (Figure 3a,b). Drought-treated plants supplemented with Ca2+ significantly (p < 0.05) reduced the O2•− accumulation starting at 12 h (Figure 3b), and significantly (p < 0.05) reduced the H2O2 accumulation starting at 12 h with an exception for 24 h (Figure 3a). The maximum reduction of O2•− and H2O2 contents with Ca2+ treatment were 31.82% and 17.25% at 12 h, respectively (Figure 3a,b).
The MDA content in drought-stressed plants was significantly higher (p < 0.05) than the control plants starting at 24 h, with the sharpest accumulation of 38.37% over the control plants at 72 h (Figure 3c). Compared to the drought stress, the Ca2+-treated plants showed a significantly lower MDA content starting at 24 h, which reduced the MDA accumulation by 13.94% at 72 h (Figure 3c).
3.3. Osmoregulation Substances
Compared to the control, the total proline and soluble sugar contents were significantly higher (p < 0.05) under drought stress starting at 12 h, and gradually increased as drought stress intensified, with the maximum increases of 64.21% and 59.09% at 72 h, respectively (Figure 4a,b). Drought-treated plants supplemented with Ca2+ significantly (p < 0.05) further increased the accumulation of proline and soluble sugar contents (Figure 3a,b). Glycine betaine only increased significantly (p < 0.05) under moderate drought stress (12, 24 h) (Figure 4c). Compared with the drought treatment, Ca2+-treated plants significantly increased the glycine betaine contents starting at 24 h, with the maximum increase up to 26.87% at 48 h (Figure 4c).
3.4. Antioxidant Enzymes
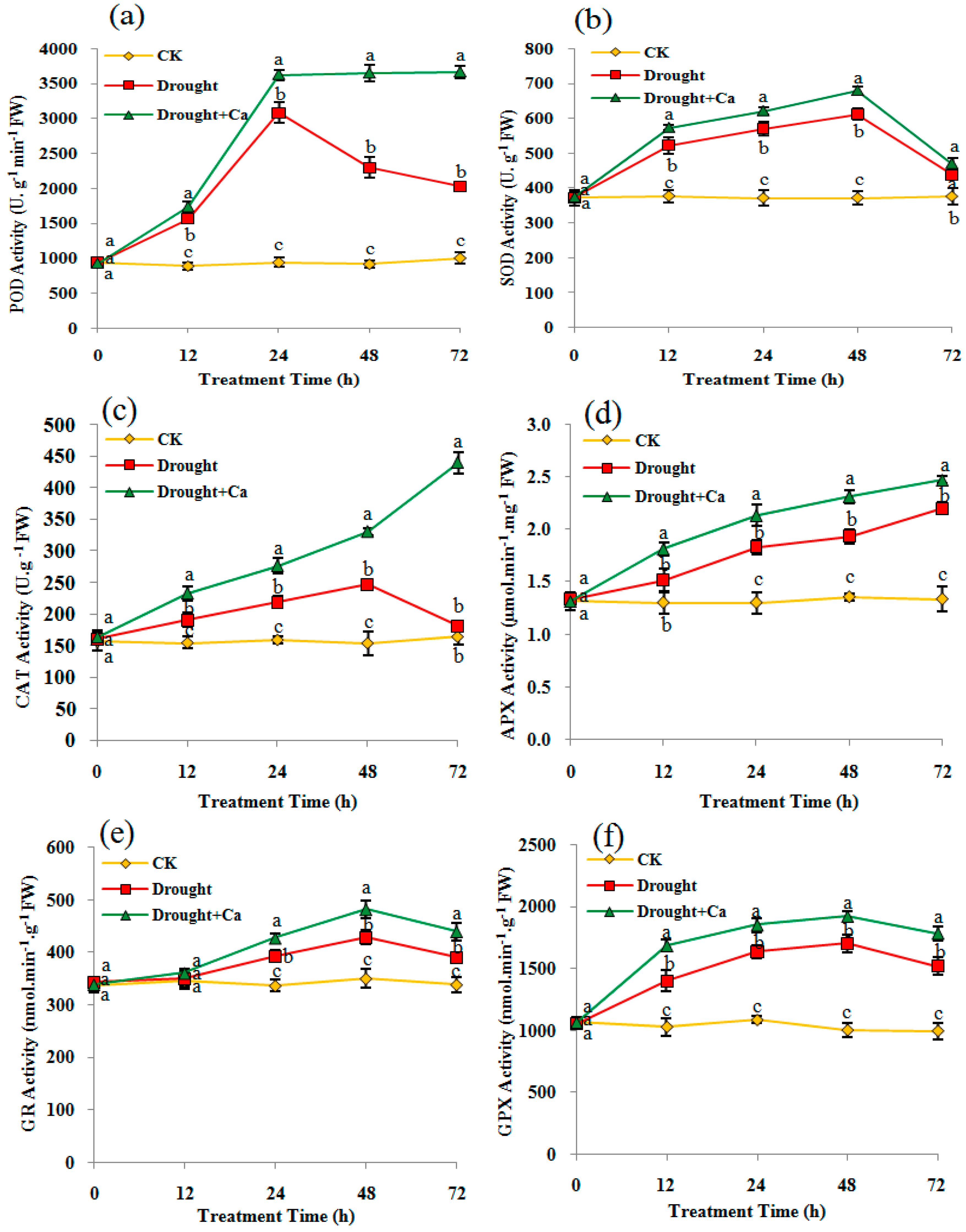
Drought stress and exogenous Ca2+ positively regulated the antioxidant enzyme activities in the C. glauca cuttings. Under drought stress, the activities of POD, SOD and GPX were significantly induced (p < 0.05) starting at 12 h, while the activities of APX and GR significantly increased at 24 h (Figure 5a,b,d–f). The activities of CAT increased from 12 h to 48 h, but showed no differences compared to the control at 72 h (Figure 5c). Compared to the drought stress, Ca2+ treatment significantly increased (p < 0.05) POD, CAT, APX, and GPX activities starting at 12 h (except for POD at 72 h) and POD and GR activities starting at 24 h (Figure 5a–f). For instance, Ca2+-treated plants remarkably increased POD and CAT activities as the drought condition intensified, with the maximum increases of 80.76% and 143.17% at 72 h, respectively (Figure 5a,c). The data showed that exogenous Ca2+ mainly increased the activities of the major antioxidant enzymes activities in C. glauca cuttings under water deficit.
3.5. Photosynthetic Parameters
The SPAD value decreased as the drought stress persisted. However, a significant increase (p < 0.05) in SPAD was observed in Ca2+-treated plants compared with the drought treatment across all stress time points (Table 1). Moreover, PEG-6000 induced drought stress significantly (p < 0.05) inhibited the Pn, Tr, and Gs of C. glauca compared to the control treatment, with the exception of a significant (p < 0.05) increase in WUE (Table 1). The largest decreases in Pn, Tr, and Gs under water deficit were 55.07%, 63.92%, and 55.08% at 72 h, respectively (Table 1). Ca2+ application significantly (p < 0.05) improved the above photosynthetic parameters at all treatment times (Table 1). Compared with the drought stress, the Pn and WUE in Ca2+-treated plants increased by 57.55% and 13.55% at 72 h, respectively (Table 1).
4. Discussion
The present study effectively confirmed that exogenous Ca2+ could increase the drought tolerance of Cyclobalanopsis glauca (Thunb.) Oerst. (C. glauca). Compared to plants with simple drought treatment, leaves with Ca2+ application showed less yellowing, indicating a positive role of Ca2+ in increasing comprehensive drought resistance in plants. Relative water content (RWC) and water potential (WP) are two appropriate indicators in estimating the water status of plants. Drought induced significant declines in RWC and WP in C. glauca. External Ca2+ application alleviated drought damage, as manifested by the increases in these two factors compared to simply drought-treated plants. These findings partially support the results reported in Zoysia japonica Steud. [15], Nitraria tangutorum Bobr. [39], Zea mays L. [20,40], Triticum aestivum L. [40] and Trifolium repens L. [16], indicating a crucial role of Ca2+ in improving water conditions under drought stress. External Ca2+ application can mitigate the inhibition of aquaporin under osmotic stress in Zea mays [41], which plays an essential role in facilitating the transport of water [42]. Thus, whether the relative high water status maintained by Ca2+ in this study partly contributed to the increased aquaporin expression should be further studied.
Osmotic adjustment is an important mechanism for plants to maintain water status under drought conditions [43]. Accumulations of proline, soluble sugars, and glycine betaine are known to be associated with drought tolerance in plants [15,18,44]. The calcium-induced accumulation of proline, soluble sugar, and glycine betaine indicates the important role of calcium in alleviating the impact of drought [15,18]. For instance, Ca2+ application induced proline accumulation in Zoysia japonica under drought conditions [15]. Foliar calcium spray accelerated the accumulation of proline content in maize, which enhanced its tolerance to drought stress [45]. Indeed, Knight et al. [46] reported that a gene involved in proline biosynthesis could be upregulated by exogenous Ca2+ application under drought stress, implying a molecular function of Ca2+ involved in regulating the drought-resistant gene. Among the compatible solutes, glycine betaine is recognized as a particularly effective protectant against abiotic stress [44]. Calcium chloride application further increased the glycine betaine content in Catharanthus roseus L., thereby conferring greater osmoprotection under drought stress [14]. Chen et al. [47] found that Populus euphratica Oliv. accumulated an amount of soluble sugar for osmoregulation to counter drought stress in a desert riparian forest. Xi et al. [48] reported that exogenous Ca2+ addition could increase soluble sugar content of Vitis vinifera L. during water stress. In the present study, we found that Ca2+ application resulted in a further increase in the concentrations of all three osmoregulation substances. This result indicated that calcium plays a vital role in improving water status by accelerating the accumulation of numerous osmoregulation substances during drought stress. However, massive accumulation of proline has been found to have toxic effects on plants [49]. Thus, research on accumulation and the effect of osmoregulation regulated by Ca2+ under prolonged drought stress is still needed.
The production and scavenging of ROS are unbalanced under water deficit conditions, causing oxidative damage to plants [8,9]. Therefore, drought stress induced peroxidation of membrane lipids as the excessive accumulation of ROS. MDA is considered as an important criterion of the peroxidation of membrane lipids [50]. Calcium is known to maintain membrane integrity, consequently enhancing drought tolerance in plants [51]. The absence of Ca2+ significantly increased the H2O2 and O2•− across all treatment times, accompanied by a significant enhancement of MDA in C. glauca. Plants synthesize numerous antioxidant enzymes to maintain the balance of the formation and elimination of ROS [52]. SOD is the first line of ROS defense that catalyzes O2•− dismutation into H2O2 and O2 [53]. CAT and POD are the primary antioxidant enzymes for H2O2 scavenging in plant cells [54]. APX is involved in the ascorbate-glutathione (ASA-GSH) cycle, which is an efficient antioxidant system for the detoxification of H2O2. GR also plays a major role in maintaining the pool of reduced glutathione through H2O2 removal and by activating the ascorbate-glutathione cycle. GPX catalyze the reduction of H2O2 and lipid peroxides using GSH as an electron donor [55]. In the present study, drought stress induced enhancements of the antioxidant enzyme activities analyzed, and Ca2+ treatments further improved their activities. These results indicate that calcium plays a crucial role in detoxifying drought-induced oxidative stress by improving numerous antioxidant enzyme activities in C. glauca. It is note worthy that some antioxidant enzymes (such as SOD, GR, and GPX) showed decreased activities with the extension of the time of stress even in Ca2+ application treatments. The results may imply that the effects of Ca2+ in upregulating these antioxidant enzyme activities would be limited under prolonged drought stress.
Photosynthesis is the basis of plant growth and development, and water is recognized as one of the most important factors affecting photosynthesis [56]. Drought stress depressed stamota aperture, Rubisco activity, and chlorophyll biosynthesis, resulting in a reduction of the photosynthesis rate [10]. Ca2+ plays a vital role in the regulation of plant photosynthesis, including photosynthetic electron transport, the Calvin cycle, and chlorophyll stabilization [57,58]. In the present study, the application of Ca2+ significantly improved photosynthesis-related parameters such as Pn, Tr, Gs, WUE, and SPAD in C. glauca. These findings are consistent with previous studies on Sophora viciifolia Hance. [59], Arachis hypogaea L. [60], and Cucumis sativus L. [61]. Ramalho et al. [62] investigated the effects of Ca deficiency on Coffea arabica L. photosynthesis, indicating a pivotal role of Ca2+ in maintaining photochemical efficiency and chlorophyll content. Hu et al. [58,63] studied the proteomic responses of Pinus massoniana L. and Liquidambar formosana Hance. under acid rain stress with different calcium treatments, which confirmed that Ca2+ plays a vital role in the upregulation of genes involved in photosynthetic electron transport and carbon fixation. Under atrazine stress, Ca2+ application upregulated the expression of D1 protein in Pennisetum giganteum A. Rich., which has been recognized as one of the core subunits in photosystem II (PSII) [64]. Thus, external Ca2+ treatment might upregulate genes involved in electron transport, carbon assimilation, and PSII photochemical efficiency, which jointly improve the photosynthetic capacity of C. glauca under drought stress.
Supporting our hypothesis, high exogenous calcium could enhance the short-term drought tolerance of calcicole plant C. glauca. By improving the water status and capacities of ROS scavenging and photosynthesis, the Karst calcium-rich habitat might increase the survival chances of native plants under intermittent drought conditions. Even so, water status, ROS contents, and Pn in drought-treated plants with Ca2+ application hardly reached the control level, indicating a limited role of Ca2+ in drought resistance. Moreover, as plant species, and the duration and intensity of drought stress could make such defense systems variable [65], far more studies still need to be undertaken under multiple drought conditions across various species.
5. Conclusions
In this study, drought stress induced a significant decrease in RWC, WP, Pn, Gs, Tr, and SPAD, and enhanced the levels of ROS production (H2O2 and O2•−) and MDA content in calcicole Cyclobalanopsis glauca (Thunb.) Oerst. (C. glauca) leaves, implying that a reduction in water status and photosynthesis capacity combined with an increased oxidative stress were induced after exposure to water deficit conditions. Enhanced activities of antioxidant enzymes (POD, CAT, SOD, APX, GR, and GPX) and increased accumulation of osmoregulation substances (proline, soluble sugar, and glycine betaine) were also induced under drought treatment, suggesting that an antioxidant defense with osmotic adjustment was induced to enhance drought tolerance in C. glauca. Furthermore, short-term exogenous Ca2+ application alleviated the harmful ecophysiological effects produced by drought. Ca2+ amendment lowered the drought-induced oxidative stress with an increased activity of all the antioxidant enzymes mentioned. External Ca2+ further stimulated the accumulation of osmoregulation substances, which improved water status under short-term drought conditions. The Pn, Gs, Tr, SPAD, and WUE levels were also increased with external Ca2+ treatment, indicating that an improvement in photosynthesis capacity could be induced through Ca2+ application. In conclusion, high soil calcium content might enhance the tolerance of the calcicole C. glauca in a drought environment by maintaining water status, mitigating oxidative stress, and improving photosynthesis capacity.
Author Contributions
Conceptualization, L.X. and S.L.; Investigation, X.L. and W.L.; Formal Analysis, H.R. and X.Y.; Resources, J.W.; Writing, L.X.
Funding
This research was funded by Fundamental Research Funds for the Central Non-profit Research Institution of CAF (CAFYBB2016QA002), the Zhejiang Province Public Welfare Technology Application Research Project (2016c32028), the National Natural Science Foundation of China (31600586), the National Key R&D Program of China (2016YFC0502605-3), and Fundamental Research Funds of CAF (CAFYBB2017MA001).
Acknowledgments
We would like to thank Bin Liang and Bin Zhu for their assistance in collecting the plant materials used for the experiments. We are sincerely grateful to the anonymous reviewers for their valuable comments to improve the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, S.; Liu, Q.; Zhang, D. Karst Rocky Desertification in southwestern China: Geomorphology, landuse, impact and rehibilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Li, S.; Birk, S.; Xue, L.; Ren, H.; Chang, J.; Yao, X. Seasonal changes in the soil moisture distribution around bare rock outcrops within a karst rocky desertification area (Fuyuan County, Yunnan Province, China). Environ. Earth Sci. 2016, 75, 1482. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Ren, H.D.; Xue, L.; Chang, J.; Yao, X.H. Influence of bare rocks on surrounding soil moisture in the karst rocky desertification regions under drought conditions. Catena 2014, 116, 157–162. [Google Scholar] [CrossRef]
- Cao, J.; Yuan, D.; Tong, L.; Azim, M.; Yang, H.; Huang, F. An overview of Karst ecosystem in Southwest China: Current state and future management. J. Resour. Ecol. 2015, 6, 247–256. [Google Scholar]
- Zhou, Y.C.; Pan, G.X. Adaptation and adjustment of Maolan forest ecosystem to karst environment. Carsol. Sin. 2001, 20, 47–52. [Google Scholar]
- Li, A.D.; Lu, Y.F.; Wei, X.L.; Yu, L.F. Studies on the regime of soil moisture under different microhabitats in Huajiang karst valley. Carsol. Sin. 2008, 27, 56–62. [Google Scholar]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in Maize seedlings. Front. Plant Sci. 2015, 6, 1241. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef]
- Cruz, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Parry, M.A.; Andralojc, P.J.; Khan, S.; Lea, P.J.; Keys, A.J. Rubisco activity: Effects of drought stress. Ann. Bot. 2002, 89, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Gopi, R.; Lakshmanan, G.A.; Panneerselvam, R. Triadimefon induced changes in the antioxidant metabolism and ajmalicine production in Catharanthus roseus (L.) G. Don. Plant Sci. 2011, 171, 271–276. [Google Scholar]
- Deng, X.; Liu, Y.; Xu, X.; Liu, D.; Zhu, G.; Yan, X.; Wang, Z.; Yan, Y. Comparative proteome analysis of wheat flag leaves and developing grains under water deficit. Front. Plant Sci. 2018, 9, 425. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, Y.; Guo, K. Ecophysiological adaptations to drought stress of seedlings of four plant species with different growth forms in karst habitats. Chin. J. Plant Ecol. 2011, 35, 1070–1082. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Sankar, B.; Kishorekumar, A.; Gopi, R.; Somasundaram, R.; Panneerselvam, R. Water deficit stress mitigation by calcium chloride in Catharanthus roseus: Effects on oxidative stress, proline metabolism and indole alkaloid accumulation. Colloids Surf. B Biointerfaces 2007, 60, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Li, X.; Zhang, L. The Effect of Calcium Chloride on Growth, Photosynthesis, and Antioxidant Responses of Zoysia japonica under Drought Conditions. PLoS ONE 2013, 8, e68214. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Q. Exogenous calcium alters activities of antioxidant enzymes in Trifolium repens L. leaves under peg-induced water deficit. J. Plant Nutr. 2010, 33, 1874–1885. [Google Scholar] [CrossRef]
- Moustakas, M.; Sperdouli, I.; Kouna, T.; Antonopoulou, C.I.; Therios, I. Exogenous proline induces soluble sugar accumulation and alleviates drought stress effects on photosystem II functioning of Arabidopsis thaliana leaves. Plant Growth Regul. 2011, 65, 315. [Google Scholar] [CrossRef]
- Kang, J.; Zhao, W.; Zheng, Y.; Zhang, D.M.; Zhou, H.; Sun, P. Calcium chloride improves photosynthesis and water status in the C4 succulent xerophyte Haloxylon ammodendron under water deficit. Plant Growth Regul. 2017, 82, 1–12. [Google Scholar] [CrossRef]
- Li, Z.; Tan, X.F.; Lu, K.; Liu, Z.M.; Wu, L.L. The effect of CaCl2 on calcium content, photosynthesis, and chlorophyll fluorescence of tung tree seedlings under drought conditions. Photosynthetica 2017, 55, 553–560. [Google Scholar] [CrossRef]
- Marques, D.J.; Ferreira, M.M.; da Silva Lobato, A.K.; Guedes de Carvalho, J.; de Assuncao Carvalho, J.; Alves de Freitas, W.; Ribeiro Bastos, A.R.; Pereira, F.J.; Mauro de Castro, E. CaSiO3 improves water potential and gas exchange but not contribute to the production parameters of maize plants exposed to different irrigation depths. Aust. J. Crop Sci. 2015, 8, 1257–1265. [Google Scholar]
- Wang, W.H.; Chen, J.; Liu, T.W.; Chen, J.; Han, A.D.; Simon, M.; Dong, X.J.; He, J.X.; Zheng, H.L. Regulation of the calcium-sensing receptor in both stomatal movement and photosynthetic electron transport is crucial for water use efficiency and drought tolerance in Arabidopsis. J. Exp. Bot. 2014, 65, 223–234. [Google Scholar] [CrossRef] [PubMed]
- David, L.; Raoul, R.; Alain, P. Calcium in plant defence–signalling pathways. New Phytol. 2006, 171, 249–269. [Google Scholar]
- Shao, H.B.; Song, W.Y.; Chu, L.Y. Advances of calcium signals involved in plant anti-drought. C. R. Biol. 2008, 331, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.J.; Wei, F.J.; Wang, C.; Wu, J.J.; Ratnasekera, D.; Liu, W.X.; Wu, W.H. Arabidopsis calcium-dependent protein kinase CPK10 functions in ABA and Ca2+-mediated stomatal regulation in response to drought stress. Plant Physiol. 2010, 154, 1232–1243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; You, Y.M.; Huang, Y.Q.; Li, X.K.; Zhang, J.C.; Zhang, D.N.; He, C.X. Effects of drought stress on Cyclobalanopsis glauca seedlings under simulating karst environment condition. Acta Ecol. Sin. 2012, 32, 6318–6325. [Google Scholar] [CrossRef]
- Zhang, Z.F.; You, Y.M.; Huang, Y.Q.; Li, X.K.; Zhang, J.C.; Zhang, D.N.; He, C.X. Changes of Cyclobalanopsis glauca water parameters under drought stress in karst environment. Chin. J. Ecol. 2011, 30, 664–669. [Google Scholar]
- Sergiev, I.; Alexieva, V.; Karanov, E. Effect of spermine, atrazine and combination between them on some endogenous protective systems and stress markers in plants. C. R. Acad. Bulg. Sci. 1997, 51, 121–124. [Google Scholar]
- Meng, X.; Wang, J.R.; Wang, G.D.; Liang, X.Q.; Li, X.D.; Meng, Q.W. An R2R3-MYB gene, LeAN2, positively regulated the thermo-tolerance in transgenic tomato. J. Plant Physiol. 2015, 175, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Luo, Q.; Tian, Y.; Meng, F. Physiological and proteomic analyses of the drought stress response in Amygdalus mira (Koehne) Yü et Lu roots. BMC Plant Biol. 2017, 17, 53. [Google Scholar] [CrossRef] [PubMed]
- Gao, J. Experimental Techniques of Plant Physiology; World Publishing Corporation: Xi’an, China, 2000. [Google Scholar]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 1, 764–775. [Google Scholar]
- Upadhyaya, A.; Sankhla, D.; Davis, T.D.; Sankhla, N.; Smith, B.N. Effect of paclobutrazol on the activities of some enzymes of activated oxygen metabolism and lipid peroxidation in senescing soybean leaves. J. Plant Physiol. 1985, 121, 453–461. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Cakmak, I.; Strbac, D.; Marschner, H. Activities of hydrogen peroxide-scavenging enzymes in germinating wheat seeds. J. Exp. Bot. 1993, 44, 127–132. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Yuan, X.T.; Liu, W.; Xuan, Y.N.; Zhang, Y.Y.; Yan, Y.Q. Physiological effects of exogenous Ca2+ on Nitraria tangutorum under salt stress. Plant Physiol. J. 2014, 50, 88–94. [Google Scholar]
- Harsh, N. Accumulation of osmolytes and osmotic adjustment in waterstressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonists. Environ. Exp. Bot. 2003, 50, 253–264. [Google Scholar]
- Wu, Y.; Liu, X.; Wang, W.; Zhang, S.; Xu, B. Calcium regulates the cell-to-cell water flow pathway in maize roots during variable water conditions. Plant Physiol. Biochem. 2012, 58, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef] [PubMed]
- Serraj, R.; Sinclair, T.R. Osmolyte accumulation: Can it really help increase crop yield under drought conditions? Plant Cell Environ. 2002, 25, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Murata, N. Glycinebetaine: An effective protectant against abiotic stress in plants. Trends Plant Sci. 2008, 13, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, N.; Muhammad, S.N.; Rashid, A.; Muhammad, Z.I.; Muhammad, Y.A.; Yasir, H.; Shah, F. Foliar calcium spray confers drought stress tolerance in maize via modulation of plant growth, water relations, proline content and hydrogen peroxide activity. Arch. Agron. Soil Sci. 2018, 64, 116–131. [Google Scholar]
- Knight, H.; Trewavas, A.J.; Knight, M.R. Calcium signaling in Arabidopsis thaliana responding to drought and salinity. Plant J. 1997, 12, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, H.; Chen, Y. Adaptation strategies of desert riparian forest vegetation in response to drought stress. Ecohydrology 2013, 6, 956–973. [Google Scholar] [CrossRef]
- Xi, Z.M.; Sun, W.J.; Zhang, Z.W. Effect of exogenous Ca2+ on drought resistance physiological indexes of wine grape cultivar Pinot Noir under water stress. J. Northwest A&F Univ. 2007, 35, 137–140. [Google Scholar]
- Shamsul, H.; Qaiser, H.; Mohammed, N.A.; Arif, S.W.; John, P.; Aqil, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar]
- Jiang, Y.; Huang, B. Effect of calcium on antioxidant activities and water relations associated with heat tolerance in two cool-season grasses. J. Exp. Bot. 2001, 52, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Sairam, R.K.; Srivastava, G.C.; Tyagi, A.; Meena, R.C. Role of ABA, salicylic acid, calcium and hydrogen peroxide on antioxidant enzymes induction in wheat seedlings. Plant Sci. 2005, 169, 559–570. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yi, H. Effect of sulfur dioxide on ROS production, gene expression and antioxidant enzyme activity in Arabidopsis plants. Plant Physiol. Biochem. 2012, 58, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, K.; Miyao, K.; Gaber, A.; Takeda, T.; Kanaboshi, H.; Miyasaka, H.; Shigeoka, S. Enhancement of stress tolerance in transgenic tobacco plants overexpressing Chlamydomonas glutathione peroxidase in chloroplasts or cytosol. Plant J. 2004, 37, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexa, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Hochmal, A.K.; Schulze, S.; Trompelt, K.; Hippler, M. Calcium-dependent regulation of photosynthesis. Biochim Biophys Acta 2015, 1847, 933–1003. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.J.; Wu, Q.; Liu, X.; Shen, Z.J.; Chen, J.; Liu, T.W.; Chen, J.; Zhu, C.Q.; Wu, F.H.; Chen, L.; et al. Comparative proteomic analysis reveals the effects of exogenous calcium against acid rain stress in Liquidambar formosana Hance leaves. J. Proteome Res. 2015, 15, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, P.; Chen, C.; Song, G.; Tang, H. Effect of Exogenous Ca2+ on the growth, physiological characters of Sophora viciifolia seedlings in Karst mountain area under the drought stress. J. Nucl. Agric. Sci. 2017, 31, 2039–2046. [Google Scholar]
- Gu, X.; Sun, L.; Gao, B.; Sun, Q.; Liu, C.; Zhang, J.; Li, X. Effects of calcium fertilizer application on peanut growth, physiological characteristics, yield and quality under drought stress. Chin. J. Appl. Ecol. 2015, 26, 1433–1439. [Google Scholar]
- Chen, L.L.; Wang, X.F.; Liu, M.; Yang, F.J.; Shi, Q.H.; Wei, M.; Li, Q.M. Effect of calcium and ABA on photosynthesis and related enzymes activities in cucumber seedlings under drought stress. Chin. J. Appl. Ecol. 2016, 27, 3996–4002. [Google Scholar]
- Ramalho, J.C.; Rebelo, M.C.; Santos, M.E.; Antunes, M.L.; Nunes, M. Effects of calcium deficiency on Coffea arabica. Nutrient changes and correlation of calcium levels with some photosynthetic parameters. Plant Soil 1994, 172, 87–96. [Google Scholar] [CrossRef]
- Hu, W.J.; Chen, J.; Liu, T.W.; Wu, Q.; Wang, W.H.; Liu, X.; Shen, Z.J.; Simon, M.; Chen, J.; Wu, F.H.; et al. Proteome and calcium-related gene expression in Pinus massoniana needles in response to acid rain under different calcium levels. Plant Soil 2014, 2380, 285–303. [Google Scholar] [CrossRef]
- Erinle, K.O.; Jiang, Z.; Ma, B.; Li, J.; Chen, Y.; Ur-Rehman, K.; Shahla, A.; Zhang, Y. Exogenous calcium induces tolerance to atrazine stress in Pennisetum seedlings and promotes photosynthetic activity, antioxidant enzymes and psbA gene transcripts. Ecotoxicol. Environ. Saf. 2016, 132, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Yang, Y.; Wang, G.; Yang, L.; Sun, X. Ecophysiological responses of Abies fabri seedlings to drought stress and nitrogen supply. Physiol. Plantarum 2010, 139, 335–347. [Google Scholar]
Figure 1.
Typical pictures of plant leaf with PEG-6000 and Ca2+ treatment after 72 h.

Figure 2.
(a) Relative water content (RWC), (b) Water potential (WP) in C. glauca leaves under drought stress and amended with Ca2+. Different small letters indicate a significant difference between treatments (p < 0.05) within each sampling time. Control group with no drought or Ca2+ treatment, CK; Drought-treated plants with no Ca2+ amendment, Drought; and Ca2+ amended drought-treated plants, Drought + Ca. All data show the means ± SD of three replicates.
Figure 2.
(a) Relative water content (RWC), (b) Water potential (WP) in C. glauca leaves under drought stress and amended with Ca2+. Different small letters indicate a significant difference between treatments (p < 0.05) within each sampling time. Control group with no drought or Ca2+ treatment, CK; Drought-treated plants with no Ca2+ amendment, Drought; and Ca2+ amended drought-treated plants, Drought + Ca. All data show the means ± SD of three replicates.
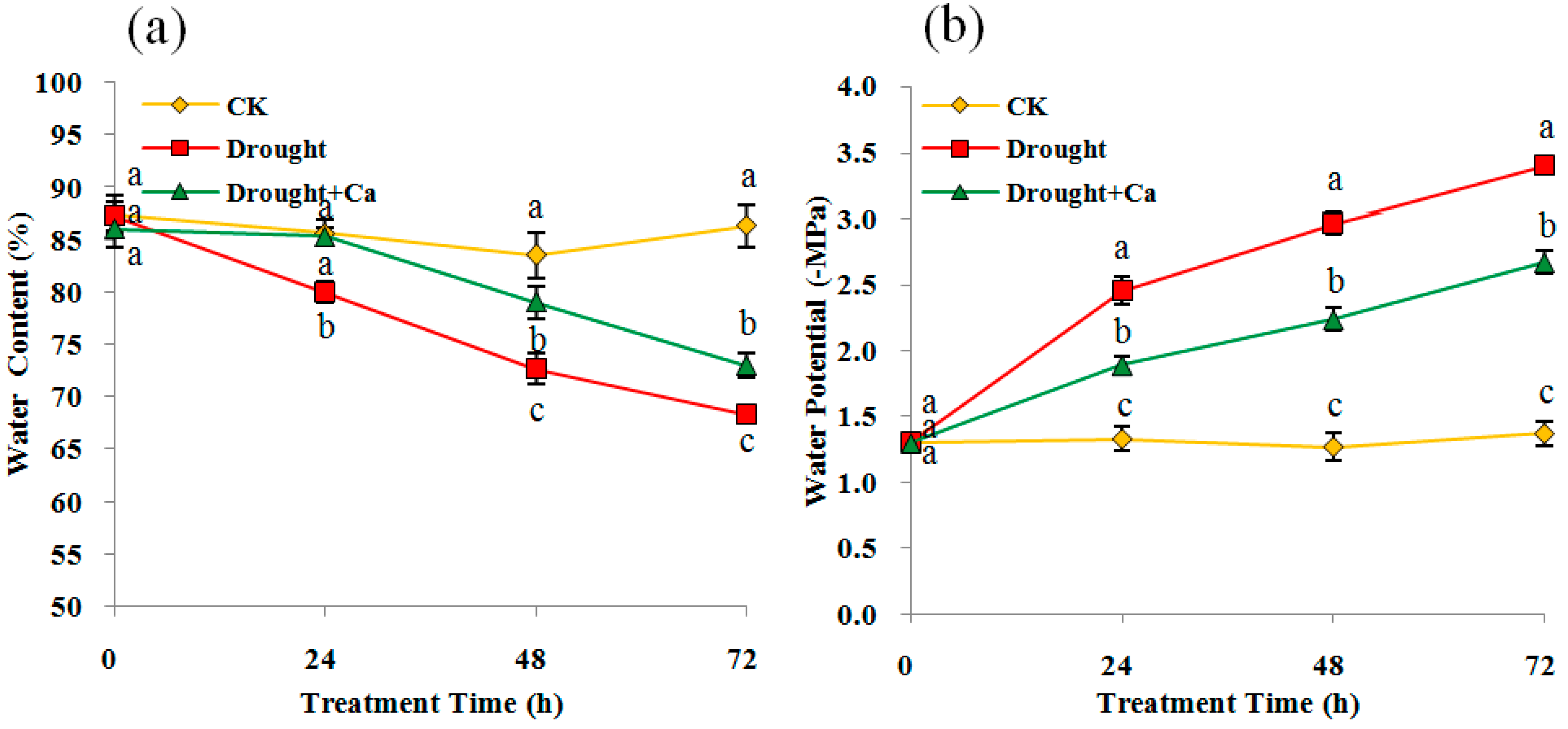
Figure 3.
(a) H2O2 content, (b) O2•− content, (c) Lipid Peroxidation (MDA content) in C. glauca leaves under drought stress and amended with Ca2+. The details of the treatments are given in the legend of Figure 2.
Figure 3.
(a) H2O2 content, (b) O2•− content, (c) Lipid Peroxidation (MDA content) in C. glauca leaves under drought stress and amended with Ca2+. The details of the treatments are given in the legend of Figure 2.
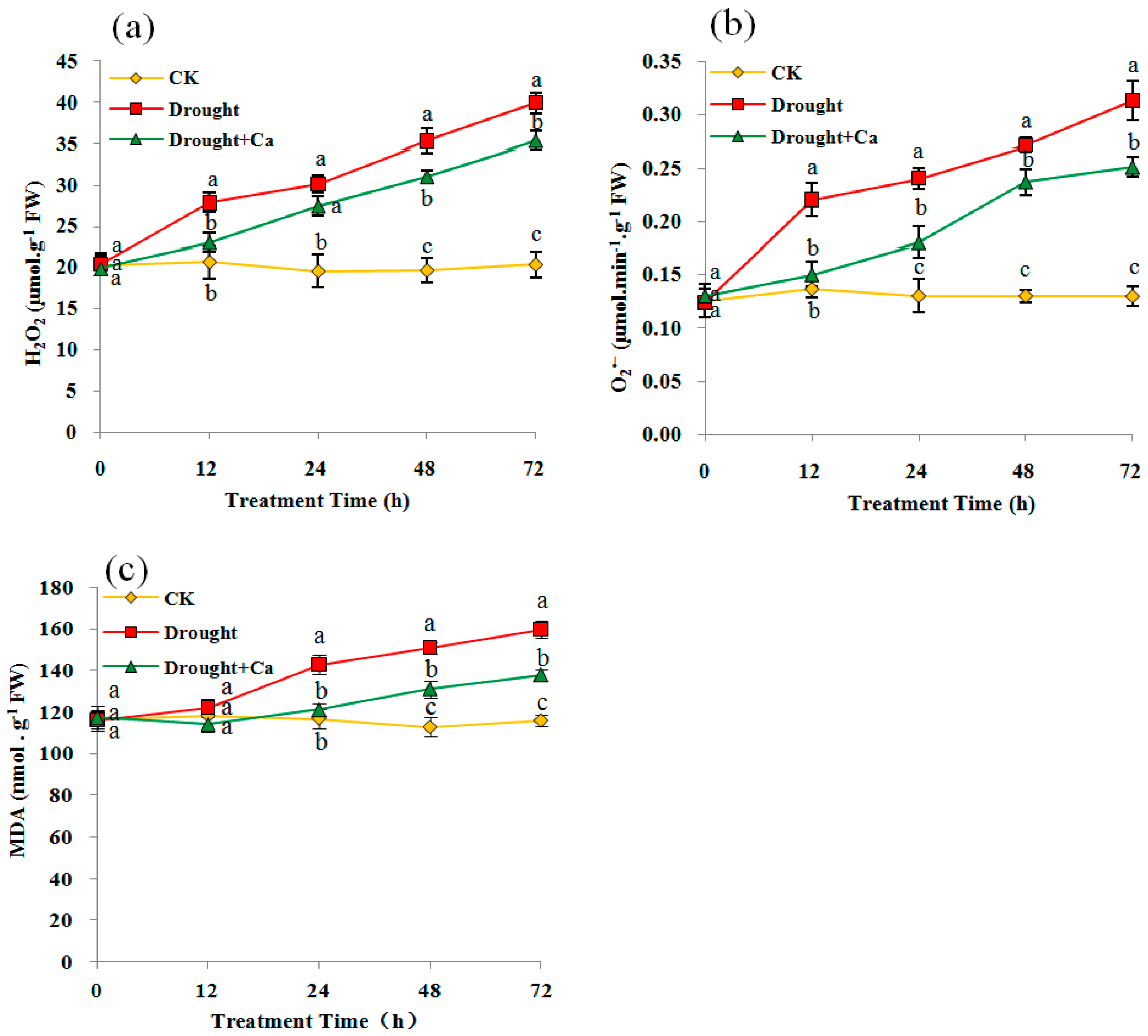
Figure 4.
(a) Proline content, (b) Soluble sugar content, (c) Glycine betaine content in C. glauca leaves under drought stress and amended with Ca2+. The details of the treatments are given in the legend of Figure 2.
Figure 4.
(a) Proline content, (b) Soluble sugar content, (c) Glycine betaine content in C. glauca leaves under drought stress and amended with Ca2+. The details of the treatments are given in the legend of Figure 2.

Figure 5.
(a) Peroxidase (POD) activity, (b) Superoxide dismutase (SOD) activity, (c) Catalase (CAT) activity, (d) Ascorbate peroxidase (APX) activity, (e) Glutathione reductase GR activity, (f) Glutathione peroxidase (GPX) activity in C. glauca leaves under drought stress and amended with Ca2+. The details of the treatments are given in the legend of Figure 2.
Figure 5.
(a) Peroxidase (POD) activity, (b) Superoxide dismutase (SOD) activity, (c) Catalase (CAT) activity, (d) Ascorbate peroxidase (APX) activity, (e) Glutathione reductase GR activity, (f) Glutathione peroxidase (GPX) activity in C. glauca leaves under drought stress and amended with Ca2+. The details of the treatments are given in the legend of Figure 2.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Photosynthetic parameters in C. glauca leaves under drought stress and amended with Ca2+.
| Treatment Time (h) | Treatment Type | SPAD SPAD Value | Pn (µmol m−2 s−1) | Tr (mmol m−2 s−1) | Gs (mmol m−2 s−1) | WUE (µmol CO2 mmol H2O−1) |
|---|---|---|---|---|---|---|
| 0 | CK | 50.50 ± 1.80 a | 5.63 ± 0.47 a | 3.57 ± 0.15 a | 0.23 ± 0.03 a | 1.58 ± 0.07 a |
| Drought | 51.00 ± 3.61 a | 5.77 ± 0.23 a | 3.67 ± 0.15 a | 0.23 ± 0.01 a | 1.57 ± 0.02 a | |
| Drought + Ca2+ | 50.33 ± 1.53 a | 5.83 ± 0.15 a | 3.70 ± 0.10 a | 0.23 ± 0.02 a | 1.58 ± 0.02 a | |
| 24 | CK | 50.40 ± 2.62 a | 6.00 ± 0.44 a | 3.50 ± 0.20 a | 0.22 ± 0.01 a | 1.71 ± 0.05 a |
| Drought | 45.00 ± 1.50 b | 4.40 ± 0.26 b | 2.42 ± 0.10 b | 0.16 ± 0.01 b | 1.82 ± 0.02 b | |
| Drought + Ca2+ | 47.63 ± 0.81 c | 5.01 ± 0.25 c | 2.77 ± 0.15 c | 0.20 ± 0.01 c | 1.81 ± 0.05 c | |
| 48 | CK | 49.01 ± 1.73 a | 5.60 ± 0.43 a | 3.53 ± 0.23 a | 0.23 ± 0.01 a | 1.58 ± 0.02 a |
| Drought | 42.33 ± 0.76 b | 3.70 ± 0.36 b | 2.04 ± 0.10 b | 0.12 ± 0.01 b | 1.81 ± 0.01 b | |
| Drought + Ca2+ | 46.13 ± 1.46 c | 4.38 ± 0.38 c | 2.21 ± 0.12 c | 0.15 ± 0.01 c | 1.98 ± 0.00 c | |
| 72 | CK | 49.03 ± 3.61 a | 6.07 ± 0.32 a | 3.51 ± 0.10 a | 0.22 ± 0.02 a | 1.73 ± 0.05 a |
| Drought | 40.57 ± 1.36 b | 3.13 ± 0.29 b | 1.75 ± 0.13 b | 0.10 ± 0.02 b | 1.85 ± 0.08 b | |
| Drought + Ca2+ | 43.00 ± 1.73 c | 3.73 ± 0.25 c | 1.99 ± 0.12 c | 0.13 ± 0.01 c | 1.95 ± 0.20 c |
Note: Control group with no drought or Ca2+ treatment, CK; Drought-treated plants with no Ca2+ amendment, Drought; and Ca2+-amended drought-treated plants, Drought + Ca2+. Photosynthesis rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), chlorophyll content (SPAD), and further increased water use efficiency (WUE). Different small letters indicate a significant difference between treatments (p < 0.05) within each sampling time.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xue, L.; Ren, H.; Long, W.; Leng, X.; Wang, J.; Yao, X.; Li, S. Ecophysiological Responses of Calcicole Cyclobalanopsis glauca (Thunb.) Oerst. to Drought Stress and Calcium Supply. Forests 2018, 9, 667. https://doi.org/10.3390/f9110667
AMA Style
Xue L, Ren H, Long W, Leng X, Wang J, Yao X, Li S. Ecophysiological Responses of Calcicole Cyclobalanopsis glauca (Thunb.) Oerst. to Drought Stress and Calcium Supply. Forests. 2018; 9(11):667. https://doi.org/10.3390/f9110667
Chicago/Turabian StyleXue, Liang, Huadong Ren, Wei Long, Xiuhui Leng, Jia Wang, Xiaohua Yao, and Sheng Li. 2018. "Ecophysiological Responses of Calcicole Cyclobalanopsis glauca (Thunb.) Oerst. to Drought Stress and Calcium Supply" Forests 9, no. 11: 667. https://doi.org/10.3390/f9110667
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.