Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau

1
State Key Laboratory of Surface Processes and Resource Ecology, College of Resources Science and Technology, Faculty of Geographical Science, Beijing Normal University, Beijing 100875, China
2
Department of Veterinary and Biomedical Sciences, University of Minnesota, 1971 Commonwealth Avenue, 205 Veterinary Science, Saint Paul, MN 55108, USA
*
Authors to whom correspondence should be addressed.
Forests 2018, 9(2), 48; https://doi.org/10.3390/f9020048
Submission received: 17 November 2017
/
Revised: 14 January 2018
/
Accepted: 15 January 2018
/
Published: 23 January 2018
(This article belongs to the Special Issue Tree Ecophysiology: Responses and Adaptation to a Changing Environment)
Abstract
:This paper measured the ecophysiological responses of Populus cathayana Rehd., Salix longistamina C. Wang et P. Y. Fu., and Ulmus pumila L. to high altitude in the Tibetan Plateau based on changes in water relations, gas exchange, and chlorophyll fluorescence. P. cathayana and U. pumila have higher survival rates than S. longistamina, but the latter has highest biomass. S. longistamina has higher water-use efficiency (WUE), lower transpiration rates (E), higher water potential (Ψ), highest light saturation point (LSP) and higher photosystem II (PSII) photochemistry efficiency (Fv’/Fm’) and non-photochemistry quenching (NPQ) than the other species, and is thus adapted to its habitat for afforestation. U. pumila has lower E, light compensation point (LCP), dark respiration (Rd), Fv’/Fm’ and electron transport rate (ETR), with higher Ψ, apparent quantum yield (AQY), net photosynthetic rate (Pn) and non-photochemical quenching (NPQ), which helps it maintain water balance and utilize weak light to survive at high altitude. Relative low WUE, Ψ, Rd, NPQ, with high E, Pn, Fv’/Fm’ and biomass, imply that P. cathayana is more suitable for shelterbelt forests than for a semi-arid habitat. These three species can adapt to high-altitude conditions by different physiological mechanisms and morphological characteristics, which can provide a theoretical basis for afforestation and forest management in the Qinghai Tibetan Plateau.
1. Introduction
Climate change affects the distribution, population structure, and growth dynamics of plants [1,2]. Survival of a species in a particular environment requires the ability to adapt to the habitat’s characteristics, and, plants have developed a wide range of physiological responses to cope with environmental stresses [3]. Plants either adjust their photosynthetic performance to their environment or adjust their morphological and physiological traits to maintain a homeostatic photosynthetic performance [4,5]. The Tibetan Plateau of western China (also known as the Qinghai-Tibetan Plateau) is the world’s largest and highest plateau. Its harsh and complex climatic conditions limit the distribution and diversities of plants due to the effects of high altitude (above 3000 m asl), which include high solar irradiance, reduced CO2 and O2, exposure to strong winds, shallow soils, low temperatures, and low availability of water and nutrients [6,7]. Although these characteristics are not favorable for plant growth, many species survive and grow under these conditions by developing special adaptation mechanisms [6]. Understanding how these adaptations affect tree growth is important for researchers interested in predicting the response of forests of the Tibetan Plateau to future climate change.
Three tree species are important parts of the plateau’s vegetation. Populus cathayana Rehd. is a native Chinese species that is distributed mainly in northern, southwestern, and central China, covering a large geographic range [8,9]. The species has strong resistance to stressful environments and is an important genetic resource, particularly for forest shelterbelts and planting to restore degraded land [10]. Ulmus pumila L. is also a native Chinese species, with a long history of cultivation. It develops a deep and widespread root system, and is highly resistant to drought and cold, especially in harsh environments such as that of the Tibetan Plateau [11]. It provides important ecosystem services such as soil and water conservation and the prevention of wind erosion and blowing sand, and also provides high-quality timber; in many of the regions where it grows, it cannot be replaced by other species [12]. Salix longistamina C. Wang et P. Y. Fu is mainly distributed in the Tibetan Plateau and the surrounding region, where it mainly grows in valleys and river plains [13]. The species can be used for protection of river banks against erosion, stabilization of sandy soils, and as a windbreak [14]. The species has strong resistance against pests and diseases, and high tolerance of drought and poor soils [15]. Like U. pumila, it has a deep and widespread root system [16].
Despite the ecological importance and many favorable characteristics of these species, their ecophysiology and adaptive mechanisms have been poorly studied, particularly at the high altitudes of the Tibetan Plateau. Plant species adapt to adverse environments by different plastic responses, such as physiological or morphological adjustments [17].
Under high illumination levels such as those on the Tibetan Plateau, damage to a plant’s photosynthetic apparatus can be severe; this damage can be revealed by a reduction in the variable fluorescence parameter [18]. The photosynthetic efficiency of many plants decreases under stress, which is correlated with a decrease in photochemical quenching of chlorophyll fluorescence and an increase in non-photochemical quenching (NPQ) [19,20]. Thus, studies of chlorophyll fluorescence provide insights into a plant’s responses to stress. These measurements have the advantage of being non-destructive and rapid, and they also provide information on the utilization and dissipation of light energy [21].
In the present study, our goal was to assess the physiological adaptations of the three local species to the stresses they encounter at high elevation in the southeastern Tibetan Plateau by examining the responses of their morphological characteristics (i.e., survival rate, biomass, height), water relations, gas exchange, and photosynthetic apparatus (i.e., fluorescence properties). Thus, we have proposed two main hypotheses: (1) these three species can adapt to high-altitude conditions using different physiological mechanisms; and (2) Salix longistamina is more adapted to harsh conditions with higher WUE and photosynthetic activity. The results of this study will contribute to our understanding of the adaptation mechanisms of three species in high altitude and light intensity, low temperature and soil moisture environment. Moreover, the present study could provide some guidance for forest management, cultivation and afforestation in the Tibetan Plateau.
2. Materials and Methods
2.1. Study Site
Our field measurements were carried out in 2010 near Sangye town in the middle reaches of Tibet’s Yarlung Zangbo River (29°19′ N, 91°30′ E), at an elevation of approximately 3500 m asl. The area has a cool-temperate semi-arid continental monsoon climate, but with considerable variation within the plateau. The total annual precipitation ranges between 300 and 450 mm. The mean annual temperature ranges between 5 and 9 °C, with maximum temperatures in June and July of 18 to 32 °C and minimum temperatures in February and January of −16 to −37 °C. The frost-free period averages 140 days, and the average total number of sunlight hours reaches 3092 annually.
Seedlings of the three species were from local seed nursery and planted in the spring of 2007 to stabilize an area of sandy soils and improve the ecological environment of a pediment on the northern slopes of the Nyainqentanglha Mountains that are covered with sand sheets, on the northern bank of the Yarlung Zangbo River. The planting density was about 625 trees per hectare. The species were planted in separate stands with one species of each stand at the same site, separated by at least 10 m to avoid competition between the mature trees (Figure 1). U. pumila and P. cathayana seedlings were planted with 1m height. The shoots of S. longistamina were cut into 15-cm long pieces, each containing three to four buds and soaked in water for 12 h. Then, the cuttings of S. longistamina were planted at the site. The study site was located about 20 m above the river, and the local water table was about 8 m below the surface. At the time of our measurements, sunrise occurred at around 08:00 and sunset occurred at around 20:00.
Soil water content was determined from soil samples taken from the center of the root zone (a depth of 60 cm) on the dates of measurement. The samples were dried to constant mass at 105 °C and the mass was expressed as a percentage of the water content. The water content was very low (Table 1). The water content is highest at 0–10 cm depth in all three stands of these tree species.
2.2. Leaf Gas Exchange
Leaf gas exchange of mature, fully expanded leaves of three individuals of each species was measured on clear, cloudless days (14 and 15 July) in 2010 between 08:00 and 20:00 using an LI-6400 portable photosynthesis system (LI-COR, Lincoln, NE, USA). Each set of three plants came from one of the three stands. Five replications were measured for the current mature foliage on each individual. For these individuals, we measured net photosynthetic rate (Pn), transpiration (E), the intercellular CO2 concentration (Ci), stomatal conductance (gs), the relative humidity (RH), air temperature (Ta), and leaf temperature (Tl). Water use efficiency (WUE) of each species was then obtained by the following equation.
WUE = Pn/E
2.3. Leaf Water Potential
After the gas-exchange measurements, we measured leaf water potential (Ψ) with a WP4 Dew-point Potential Meter (Decagon Devices, Pullman, WA, USA). We used six fully expanded leaves from the tops of each individual, with three individuals per species. Calibration of the device was checked hourly.
2.4. Photosynthetic Light Response Curves
We obtained photosynthetic light-response curves at 2-h intervals from 9:00 to 11:00 at a range of light intensities (from 0 to 2600 µmol m−2 s−1) using a LI-6400-02 Portable Photosynthetic System (LI-COR, Lincoln, NE, USA). The irradiance response was measured at 25 °C and an RH of 50%. CO2 concentration (400 μmol mol−1) and photosynthetic parameters of each species were obtained after fitting the light response data [22]:
where A is net photosynthesis rate, Amax the maximal net photosynthesis rate, Ic the light compensation point, I the light intensity, and αc the quantum efficiency. Dark respiration rate Rd is calculated from the relationship describing the light-limited part of the photosynthesis light response curves:
A = [Amax (I − Ic) αc] / [Amax + (I − Ic) αc]
A = αcI − Rd
For I = Ic, A = 0 and Rd = αcI.
2.5. Diurnal Course of Chlorophyll Fluorescence
Variable chlorophyll a fluorescence was measured with a LI-COR portable LI-6400-40 pulse-amplitude-modulation fluorometer (Li-Cor 6400; Li-Cor, Lincoln, NE, USA). For these measurements, we maintained mature, fully expanded leaves, still attached to the branch, in darkness overnight. Before each measurement, samples were held in darkness for more than 30 min at ambient temperature. We measured maximum fluorescence (Fm) and minimum fluorescence (Fo) simultaneously in the dark. We then exposed the leaves to a photosynthetic photon flux density (PPFD) of 1000 µmol photons m−2 s−1 for more than 3 min to measure the maximum fluorescence (Fm’) and steady-state fluorescence (Fs) in the light-adapted state after Pn stabilized. We performed these measurements every 2 h from 08:00 to 20:00 on sunny days.
We calculated the maximum photochemical efficiency of PSII using the formula (Fm − Fo)/Fm = Fv/Fm, where Fv represents variable fluorescence. We estimated the effective quantum yield of PSII (ΦPSII) using the equation ΔF/F’m = (F’m − Fs)/F’m [23]. We calculated photochemical quenching (qP) as (F’m − Fs)/(F’m − Fo) [24], and non-photochemical quenching (NPQ) as Fm/Fm’ − 1 [25]. We calculated the electron transport rate (ETR) as ΦPSII × PAR × 0.5 × 0.84 [26].
2.6. Survival Rate, Height, Biomass
We examined survival rate, crown and height of three species in the middle of growing season in 2010. At the end of growing season, biomass production and allocation of the three species were measured by means of destructive sampling with three replicates. Then, biomass samples were dried at 75 °C to constant weight.
2.7. Data Analysis
Measurements for each species were compared using analysis of variance (ANOVA). Differences among the species were evaluated using the least significance difference (LSD) method and Tukey test with a significance level of p < 0.05, including morphological characteristics (height, biomass, survival rate and crown), water relations (water potential, WUE, E and VPD), gas exchange characteristics (Pn, gs, Ci), chlorophyll fluorescence parameters (i.e., Fv/Fm, NPQ). Data were standardized before the analysis. All statistical analyses were performed using SPSS 18.0 software (SPSS Inc., IBM, Chicago, IL, USA).
3. Results
3.1. Morphological Characteristics
The survival rates (over 3 years), crown, height and biomass among the three species in the study are significantly different (p < 0.05). U. pumila has the highest survival rate (Table 2, p < 0.05). The survival rates of P. cathayana, U. pumila and S. longistamina are 93.80%, 83.83% and 71.70% respectively. The crown diameter of the three species ranges from 45.00 cm × 45.00 cm to 78.00 cm × 63.00 cm (p < 0.05). S. longistamina has the largest crown and biomass, and P. cathayana has the highest height among the three species (p < 0.05).
3.2. Water Relations
The water potential, transpiration and water use efficiency are significantly different in the species (p < 0.05). The diurnal course of water potential shows similar circadian rhythms in all three species with clear diurnal changes (Figure 2A). The Ψ of P. cathayana is lower than those of the other species throughout the day (Figure 2A, p < 0.05). The VPD of the species is not significant. The difference in vapor pressure (VPD) follows a similar diurnal pattern in all stands, with the lowest value at sunrise and VPD increasing to a maximum at around 14:00 to 16:00 (Figure 2B).
The diurnal changes in E of S. longistamina and U. pumila show two peaks, the first of which appears at about 12:00 and the second appears at about 18:00, whereas P. cathayana shows a single peak at about 12:00; P. cathayana has the highest E and U. pumila the lowest (Figure 2C, p < 0.05).
The water use efficiency of S. longistamina is generally higher than that of other two species, but the diurnal course of WUE differs among the three species (p < 0.05). In the morning, the WUE of P. cathayana is lower than that of the other two species, and increases slowly to a maximum at about 14:00 and18:00. WUE in the morning follows similar patterns for S. longistamina and U. pumila, with two peaks at about 10:00 and 14:00 (Figure 2D).
3.3. Gas Exchange
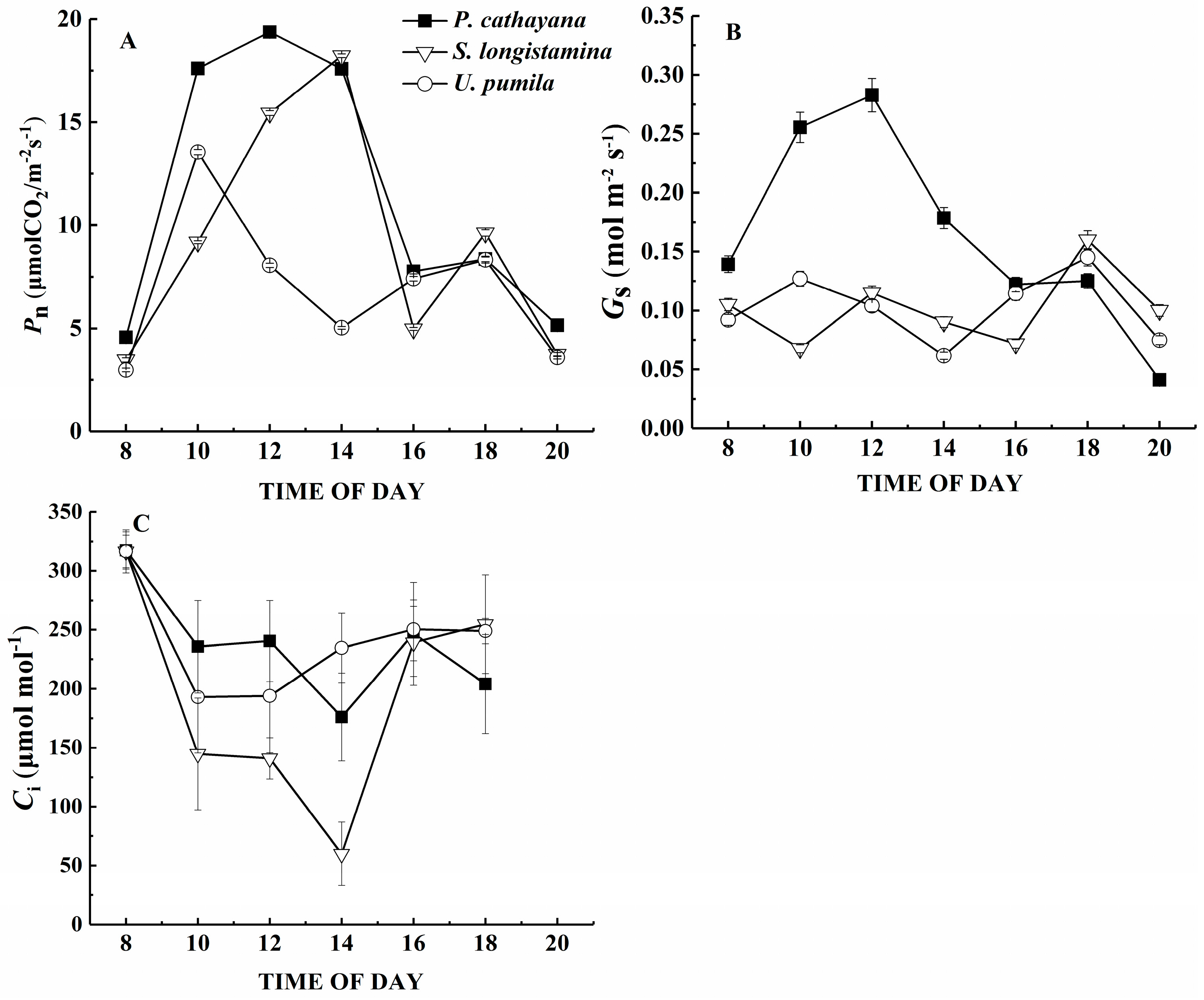
The net photosynthetic rate, stomatal conductance and intercellular carbon dioxide concentration reach a significant level among the species (p < 0.05). Pn is highest for P. cathayana and lowest for U. pumila (Figure 3A, p < 0.05). However, the diurnal changes of Pn show a similar pattern with two peaks in all three species. The diurnal trends are similar for gs, but U. pumila and S. longistamina differ little in their patterns, with the first peak appearing at 12:00 for S. longistamina, which is 2 h earlier than peak Pn for this species. Populus cathayana has a higher gs than the other two species until about 16:00 (Figure 3B,C, p < 0.05). The diurnal changes of Ci are decreased first and increased later. The Ci of S. longistamina is lowest during the day (p < 0.05).
3.4. Pn-Light Response Curves
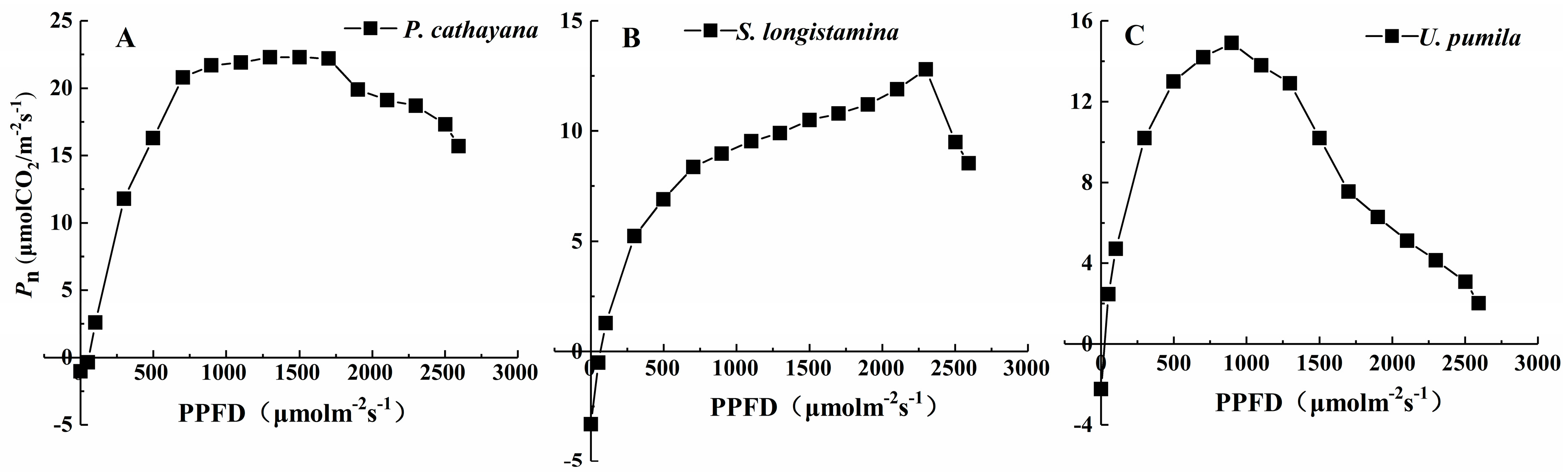
The Pn of the three species initially increases with increasing irradiance, and eventually reaches the LSP (Figure 4). The curve and Pmax for P. cathayana is obviously higher than those for the other species (p < 0.05). The differences of LSP, LCP and Rd of three species are significant (p < 0.05). S. longistamina and P. cathayana have higher LSP and LCP than U. pumila (Table 3, p < 0.05). S. longistamina has the higher Rd (3.31 µmol m−2 s−1) than other species (p < 0.05). P. cathayana and S. longistamina have the lower AQY than U. pumila (p < 0.05).
3.5. Chlorophyll Fluorescence Parameters
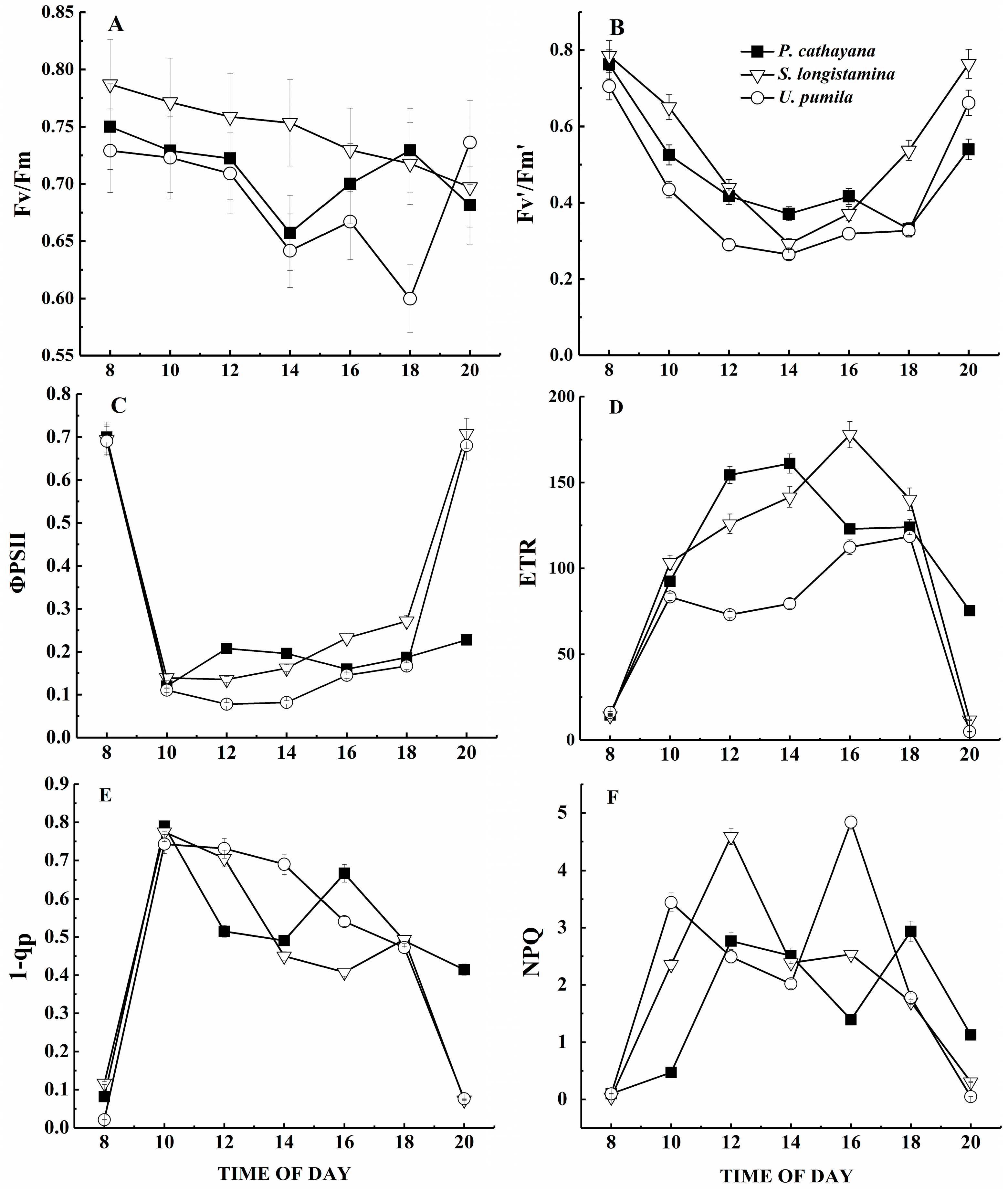
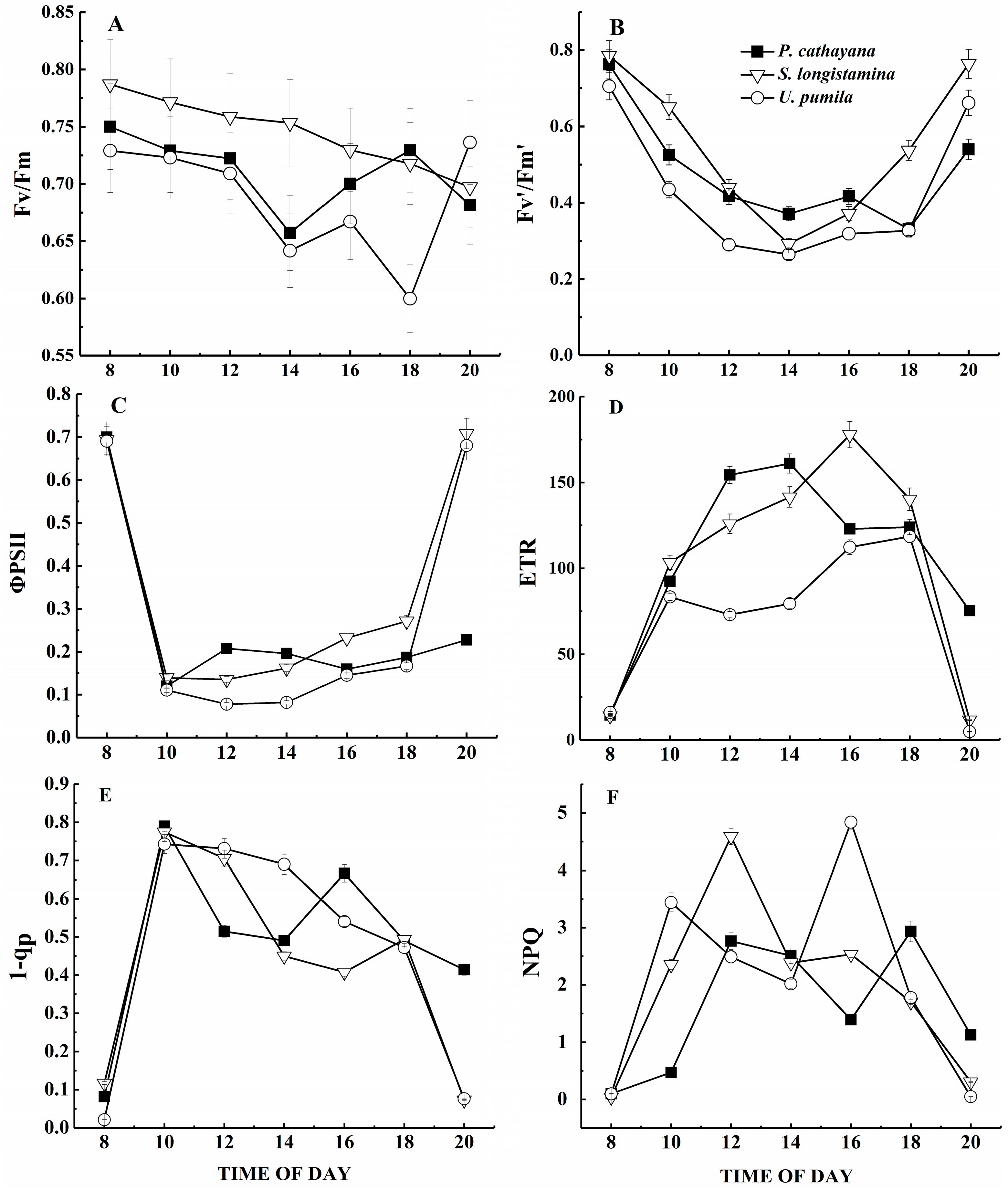
We observed distinct patterns in the diurnal course of the potential quantum yield of PSII (i.e., Fv/Fm) (Figure 5A). Fv/Fm in P. cathayana and U. pumila isn’t significantly different, but both show a mid-day depression. U. pumila shows an especially strong reduction in Fv/Fm during the day, reaching a minimum of 0.59 at around 18:00; thereafter, it increases slowly and recovers to near its value at the start of the day. However, Fv/Fm in S. longistamina decreases throughout the day, with neither a mid-day depression nor full recovery at sunset. Of the three species, S. longistamina has the highest Fv/Fm during most of the day (p < 0.05).
Fv’/Fm’ in S. longistamina and U. pumila shows similar diurnal changes (Figure 5B). Among the three species, Fv’/Fm’ is highest in S. longistamina and lowest in U. pumila (p < 0.05).
In response to the rapid increase in PPFD, ΦPSII decreases sharply after sunrise (Figure 5C). ΦPSII in S. longistamina and U. pumila increases rapidly later in the day, reaching values similar to the pre-dawn values by sunset. ΦPSII of P. cathayana does not show full recovery by sunset. ΦPSII is generally higher in S. longistamina and P. cathayana than in U. pumila (p < 0.05).
The diurnal course of ETR shows different patterns for the three species (Figure 5D). ETR of U. pumila initially increases to a peak at 10:00, then decreases until about 12:00 and subsequently recovers. ETR of P. cathayana and S. longistamina has similar patterns, but the maximum value reaches at 14:00 and 16:00, respectively. ETR is significantly higher in S. longistamina and P. cathayana than in U. pumila (p < 0.05).
In all three species, 1 − qp is significantly different, and increases sharply after sunrise, reaching a maximum value by around 10:00, then decreases slowly (Figure 5E, p < 0.05). At 16:00 and 18:00, respectively, 1 − qp reaches the second, lower peak in P. cathayana and S. longistamina; thereafter, it decreases again. S. longistamina has lowest 1 − qp during a day (p < 0.05).
After sunrise, the NPQ of all three species increases (Figure 5F), and the diurnal changes show two peaks and a mid-day depression. NPQ of S. longistamina is significantly higher than other species at 12:00 (p < 0.05).
4. Discussion
4.1. Morphological Characteristics
The survival rate of plants is influenced by many biotic and abiotic factors such as pests, water shortage, and high irradiance [27]. A high survival rate of plants suggests high adaptation ability [28]. The survival rates of P. cathayana, U. pumila and S. longistamina in the study are 93.80%, 83.83% and 71.70%, indicating these species have strong adaption ability in the Qinghai-Tibet Plateau [29]. S. longistamina has highest biomass and crown among the species, suggesting a strong tolerance to resource-limited conditions [30]. The reason probably is the strong root system and high Pn and WUE of S. longistamina. The root system of Salix is sensitive to external stress such as salinity [31]. S. longistamina may increase its root-to-shoot ratio in high irradiance conditions, which helps it absorb water and nutrients for accumulating larger biomass [32].
4.2. Water Relations
Water is one of the fundamental factors for plants influencing their growing process. However, in many regions like the Qinghai-Tibetan Plateau, water resources are becoming seriously limited. Thus, plants will utilize water sources more efficiently by changing their water regimes [33]. Ψ reflects a plant’s water status, and determines the plant’s ability to maintain its water balance by absorbing water from the soil and from neighboring cells. The higher Ψ of S. longistamina and U. pumila suggests that they sustain better water relations under water-limited conditions than P. cathayana, probably as a result of their massive root system, which can exploit deep soil moisture [34,35]. The root system of Salix has strong root anchorage, which helps roots embed in soils, and Ulmus can generate new roots rapidly and thus increase root surface area [36]. These characteristics indicate the strong water absorption capacity of these three species.
WUE is the ratio of Pn and E, which is an important physiological parameter that can help to explain a plant’s ability to maintain water equilibrium [37]. Increased WUE has been proved an adaptation to semi-arid and arid environment [38]. The higher WUE in S. longistamina indicates that it conserves water better than the other species. Its low E and high Pn support this conclusion [39]. S. longistamina and U. pumila have higher Ψ and WUE with lower E, which suggests their better water relations under water-limited conditions, and their suitability for afforestation in the Qinghai Tibetan Plateau [40]. Adversely, WUE of P. cathayana is lower because of its higher Pn and E. The higher gs of P. cathayana may result in higher E. It opens stomata to assimilate carbohydrates rapidly at first then closes to avoid water loss. This diurnal trend indicates that P. cathayana is sensitive to changes in the external environment and can utilize resources optimally [41]. The higher Pn of P. cathayana, combined with its low WUE and high −E and a large leaf area, make this species more suitable for use in a forest shelterbelt where water is abundant for the purpose of preventing erosion and decreasing wind speed to protect crops and soils [42]. These physiological adaptions help the species to survive in the semi-arid, high-altitude area.
4.3. Gas Exchange
Plant survival in a given environment depends on the plant’s ability to adapt to the habitat, moreover, plants can develop different ecophysiological characteristics even in similar habitats [43]. In the present study, three species show different gas-exchange characteristics. The higher Pn of P. cathayana and S. longistamina indicates that they have higher photosynthetic capacity than U. pumila under the conditions [44]. However, Pn of all three species remains greater than 0 μmol CO2 m−2 s−1 throughout the day during this part of the growing season, suggesting that they are capable of adapting to a high-irradiance environment. The diurnal changes in Pn show a similar pattern with two peaks in all three species. A similar phenomenon was reported for other high-altitude plants [44]. Farquhar and Sharkey [45] suggested that a reduction in photosynthetic capacity is caused by stomatal closure, as well as by non-stomatal factors. When gs and Ci decrease simultaneously, the decline in Pn is caused mainly by stomatal closure. During the mid-day depression in Pn for P. cathayana and S. longistamina, both gs and Ci decrease, so stomatal limitation of photosynthesis may explain the mid-day depression in these species. The temperature at noon reaches its peak so that these two species close stomata to avoid water loss and reduce CO2 absorption [46]. In contrast, the mid-day depression of Pn for U. pumila occurs when Ci changes in the opposite direction to gs, suggesting that the mid-day depression is primarily attributable to non-stomatal factors. High light intensity resulted in non-stomatal limitation, as well as lower Rubisco activity, electron transport rates, carboxylation efficiency and damage to PSII reaction center, which is also influenced by some biochemical indexes (e.g., plant hormone) [47]. Plants growing in the field are susceptible to photoinhibition under environmental stress [48]. Photoinhibiton decreases the photosynthetic efficiency under stress, in which input of photons exceeds demand [49]. In the present study, Fv/Fm, Fv’/Fm’, and ΦPSII of all three species decrease with increasing PPFD and then recovered in the afternoon as PPFD decreases, suggesting that photoinhibition occurs in all three species and may be another cause for the depression in Pn [50]. We will discuss this in more detail in the section Photosynthetic efficiency.
4.4. Photosynthesis Curve
Pmax has been widely used to compare the ecophysiological characteristics of plants [51]. Pn of the three species increases with increasing irradiance, and reaches the LSP in all three species. Consequently, they are likely to be able to protect their photosynthetic apparatus against photo-degradation [52]. P. cathayana has higher Pmax and LSP and lower LCP and Rd, suggesting its better light use ability. P. cathayana can grow fast in high altitude conditions with higher Pn and biomass. The largest LSP of S. longistamina indicates that it has higher light-use efficiency in the study area, and largest biomass and WUE of S. longistamina suggests a better adaptation to the high irradiance [53]. LCP reflects the ability of plants to make use of low irradiance [28]. The higher the value of AQY, the more efficiently photosynthesis converts light energy and the higher the photosynthetic capacity [54]. U. pumila cannot make full use of the available PPFD for much of the day. However, the lower LCP and Rd of U. pumila with highest AQY suggests that the species can tolerate shade and utilize weak light better than the other species, thereby increasing the accumulation of photosynthate [55]. These are the main reasons why U. pumila can survive on the Tibetan Plateau despite obvious photoinhibition and low biomass compared with the other species. Therefore, the three species have different photosynthesis mechanisms to utilize light to form more carbon assimilate to survive under high-altitude conditions.
4.5. Photochemistry Efficiency
Photoinhibition has been observed in many plants growing naturally in the field, and is revealed by a mid-day decrease in Fv/Fm [56]. In our study, there are clear differences in Fv/Fm among the three species. Fv/Fm of P. cathayana and U. pumila decreases to a lower level than that of S. longistamina at mid-day, but recovers almost completely at sunset, suggesting that photoinhibition in these species is a photoprotective mechanism accomplished through thermal dissipation of excess energy, as suggested by the mid-day increase of NPQ in both species [57]. Photoinhibition can be caused by photodamage not only to the photosynthetic apparatus but also to photoprotective mechanisms through thermal dissipation of excess energy [58]. In our study, Fv/Fm remains lower than 0.80 throughout the day, indicating that all three species are stressed by the high irradiance. This suggests the presence of chronic photoinhibition and damage to PSII’s reaction centers, resulting in photodamage during the summer at the high altitude of the study area [59]. Thus, we hypothesize that photoinhibition in all three species resulted both from a photoprotective process and from photodamage.
The maximum PSII efficiency (Fv’/Fm’) can be thought of as the efficiency with which PSII’s antennae absorb photons and deliver them to open PSII reaction centers [60]. Fv’/Fm’ is higher for S. longistamina and P. cathayana than for U. pumila, and shows a proportionally smaller decrease during the day than occurs for U. pumila. This suggests that U. pumila has less capacity to utilize the high excitation energy that is available through photosynthetic electron transport. This is supported by the lower ETR for U. pumila [61].
The actual PSII efficiency can be used to estimate the rate of photochemical reactions in terms of the absorbed light that is utilized in photochemistry [23]. ΦPSII and ETR are both high in S. longistamina and P. cathayana and low in U. pumila, indicating a high degree of phenotypic plasticity of the photosynthetic apparatus in these species [62]. Our study supports previous results, in which the operating efficiency is often linearly related to Pn [23]. The decrease in actual PSII efficiency under excess light can be caused by two processes: an increase in closure of PSII reaction centers and a decrease in the efficiency of excitation capture in PSII through photoprotective thermal dissipation of excess excitation energy before it reaches the PSII reaction centers [63]. With higher photosynthetic efficiency and with lower NPQ and a lower degree of closure of the PSII reaction centers, P. cathayana exhibits less thermal dissipation and closure of the PSII reaction centers than the other species, combined with high LSP and LCP and lowest Rd, which means that it captures more excitation energy to drive photosynthetic electron transport and larger biomass [64]. In contrast, S. longistamina has higher NPQ and a lower degree of closure of the PSII reaction centers than in P. cathayana. This suggests that the higher biomass in P. cathayana and S. longistamina results, at least in part, from higher photochemical efficiency [65]. In contrast, U. pumila has higher NPQ and a higher degree of closure of the PSII reaction centers, in addition to lower ETR and LCP, implying a higher degree of thermal dissipation [66]. These results suggest that all three species can compensate for high irradiance by changing their utilization of absorbed light through changes in electron transport, but to different extents [67]. The decrease of PSII efficiency in the three species under high irradiance results from a decrease in the efficiency of excitation capture in PSII and by an increase in the degree of closure of the PSII reaction centers [68].
5. Conclusions
Our study suggests that P. cathayana, S. longistamina and U. pumila acclimate efficiently to field conditions in the semi-arid, high-altitude study area. Of the three, S. longistamina has the highest WUE, LSP, biomass, photochemistry efficiency and lowest water loss; therefore, this species is drought-tolerant and capable of surviving well in semi-arid habitats. Despite its lower WUE, U. pumila has lower E and comparable higher water potential, AQY and lowest LCP, which lets it maintain its water balance and utilize weak light to survive at high altitude. These two species can be used in afforestation for a series of ecosystem services such as soil and water conservation. In contrast, because of low WUE, LCP and Rd and highest Pn and E, P. cathayana is more suitable for sites with abundant water like forest shelterbelts than for a desert habitat. These three species all can tolerant high-altitude conditions via different physiological mechanisms and morphological characteristics. Above all, S. longistamina can utilize resources optimally in the Tibetan Plateau. Our results can provide understanding of physiological mechanisms about these species’ survival in this area. In future forest management in the region, S. longistamina can be considered as an economically important species for cultivation to prevent soil erosion and improve the harsh environment.
Acknowledgments
We thank Y.G. Liu, Beijing Normal University, for his field assistance. This study was supported by the National Basic Research Program of China (Grant No. 2013CB956001) and by the National Natural Science Foundation of China (Grant No. 40771069).
Author Contributions
Jirui Gong and Chunlai Zhang conceived and designed the experiments; An Ran and Jiaqiong Zhang performed the field and laboratory experiments; Jirui Gong analyzed the data; Jirui Gong and Zihe Zhang wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Devi, N.; Agedorn, F.; Moiseev, P.; Bugmann, H.; Shiyatov, S.; Mazepa, V.; Rigling, A. Expanding forests and changing growth forms of Siberian larch at the Polar Urals tree line during the 20th century. Glob. Chang. Biol. 2008, 14, 1581–1591. [Google Scholar] [CrossRef]
- Qian, C.; Yin, H.; Shi, Y.; Zhao, J.; Yin, C.; Luo, W. Population dynamics of Agriophyllum squarrosum, a pioneer annual plant endemic to mobile sand dunes, in response to global climate change. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Okunlola, G.O.; Olusanya, A.O. Physiological response of the three most cultivated pepper species (capsicum spp.) in Africa to drought stress imposed at three stages of growth and development. Sci. Hortic. 2017, 224, 198–205. [Google Scholar] [CrossRef]
- Liang, E.Y.; Wang, Y.F.; Xu, Y. Growth variation in Abies georgei var. smithii along altitudinal gradients in the Sygera Mountains, Southeastern Tibetan Plateau. Trees 2010, 24, 363–373. [Google Scholar] [CrossRef]
- Najafabadi, M.Y.; Ehsanzadeh, P. Photosynthetic and antioxidative upregulation in drought-stressed sesame (Sesamum indicum L.) subjected to foliar-applied salicylic acid. Photosynthetica 2017, 4, 611–622. [Google Scholar] [CrossRef]
- Wonsick, M.M.; Pinker, R.T. The radiative environment of the Tibetan plateau. Int. J. Clim. 2014, 34, 2153–2162. [Google Scholar] [CrossRef]
- Roupioz, L.; Jia, L.; Nerry, F.; Menenti, M. Estimation of daily solar radiation budget at kilometer resolution over the Tibetan plateau by integrating Modis data products and a DEM. Remote Sens. 2016, 8, 504. [Google Scholar] [CrossRef]
- Chen, C.; Peng, Y.H. AFLP analysis of genetic diversity in Populus cathayana Rehd. originating from Southeastern Qinghai–Tibetan Plateau of China. Pak. J. Bot. 2010, 42, 117–127. [Google Scholar]
- Song, Y.; Ma, K.; Ci, D.; Zhang, Z.; Zhang, D. Biochemical, physiological and gene expression analysis reveals sex-specific differences in Populus tomentosa floral development. Physiol. Plant. 2013, 150, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Li, Z.; Wu, F.; Tang, M. Comparative photochemistry activity and antioxidant responses in male and female Populus cathayana cuttings inoculated with arbuscular mycorrhizal fungi under salt. Sci. Rep. 2016, 6, 37663. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Li, Y.; Liu, W.; Xu, H.; Sun, O.J. Changes in water use with growth in Ulmus pumila, in semiarid sandy land of northern China. Trees 2014, 28, 41–52. [Google Scholar] [CrossRef]
- Park, G.; Lee, D.; Kim, K.; Batkhuu, N.O.; Tsogtbaatar, J.; Zhu, J.J.; Jin, Y.; Park, P.S.; Hyun, O.; Kim, H.S. Morphological characteristics and water-use efficiency of siberian elm trees (Ulmus pumila L.) within arid regions of northeast Asia. Forests 2016, 7, 280. [Google Scholar] [CrossRef]
- Guillermo, N.; Doffo, S.E.; Monteoliva, M.E.; Rodríguez, V.M.; Luquez, C. Physiological responses to alternative flooding and drought stress episodes in two willow (Salix spp.) clones. Can. J. For. Res. 2016, 47, 174–182. [Google Scholar] [CrossRef]
- Karrenberg, S.; Edwards, P.J.; Kollmann, J. The life history of Salicaceae living in the active zone of floodplains. Freshw. Biol. 2002, 47, 733–748. [Google Scholar] [CrossRef]
- Pusz, W.; Urbaniak, J. Foliar diseases of willows (Salix spp.) in selected locations of the Karkonosze mts. (the giant mts). Eur. J. Plant Pathol. 2017, 148, 45–51. [Google Scholar] [CrossRef]
- Yang, H.; Chu, J.; Lu, Q.; Gao, T. Relationships of native trees with grasses in a temperate, semi-arid sandy ecosystem of northern China. Appl. Veg. Sci. 2014, 17, 338–345. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, L.; Zhou, K.; Li, Y.; Zhao, Z. Changes in protein profile of Platycladus orientalis, (L.) roots and leaves in response to drought stress. Tree Genet. Genomes 2017, 13, 76. [Google Scholar] [CrossRef]
- Robakowski, P.; Laitat, E. Effects of an enhanced ultraviolet–B irradiation on photosynthetic apparatus of several forests coniferous tree species from different locations. Acta Physiol. Plant. 1999, 21, 283–296. [Google Scholar] [CrossRef]
- Lukaszek, M.; Poskuta, J. Chlorophyll a fluorescence kinetics and CO2 exchange rates in light and in darkness in leaves of tall fescue as affected by UV radiation. Acta Physiol. Plant. 1996, 18, 345–350. [Google Scholar]
- Liu, X.; Wang, Z.; Bao, W.; Li, X.M. Photosynthetic responses of two pleurocarpous mosses to low-level nitrogen addition: A study in an old-growth fir forest. J. Bryol. 2015, 37, 15–22. [Google Scholar] [CrossRef]
- Bilger, W.G.; Schreiber, U.; Bock, M. Determination of the quantum efficiency of photosystem II and of non–photochemical quenching of chlorophyll fluorescence in the field. Oecologia 1995, 102, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Drake, B.G.; Read, M. Carbon dioxide assimilation, photosynthetic efficiency, and respiration of a Chesapeake Bay salt marsh. J. Ecol. 1981, 69, 405–423. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F.H. The use of fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Bilger, W.; Björkman, O. Role of the xanthophylls cycle in photoprotection elucidated by measurements of light–induced absorbance changes, fluorescence and photosynthesis in Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Demmig, B.; Björkman, O. Comparison of the effects of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 1987, 171, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Ceulemans, R.; Deraedt, W. Production physiology and growth potential of poplars under short-rotation forestry culture. For. Ecol. Manag. 1999, 121, 9–23. [Google Scholar] [CrossRef]
- Gong, J.R.; Zhang, X.S.; Huang, Y.M. Comparison of the performance of several hybrid poplar clones and their potential suitability for use in northern China. Biomass Bioenergy 2011, 35, 2755–2764. [Google Scholar] [CrossRef]
- Kaczmarek, D.J.; Coyle, D.R.; Coleman, M.D. Survival and growth of a range of Populus clones in central South Carolina USA through age ten: Do early assessments reflect longer-term survival and growth trends. Biomass Bioenergy 2013, 49, 260–272. [Google Scholar] [CrossRef]
- Major, J.E.; Mosseler, A.; Malcolm, J.W.; Heartz, S. Salinity tolerance of three Salix, species: Survival, biomass yield and allocation, and biochemical efficiencies. Biomass Bioenergy 2017, 105, 10–22. [Google Scholar] [CrossRef]
- Dong, Y.; Yan, M.A.; Wang, H.; Zhang, J.; Zhang, G.; Yang, M.S. Assessment of tolerance of willows to saline soils through electrical impedance measurements. For. Ecosyst. 2013, 15, 32–40. [Google Scholar]
- Mamashita, T.; Larocque, G.R.; Desrochers, A.; Beaulieu, J.; Thomas, B.R.; Mosseler, A.; Major, J.; Sidders, D. Short-term growth and morphological responses to nitrogen availability and plant density in hybrid poplars and willows. Biomass Bioenergy 2015, 81, 88–97. [Google Scholar] [CrossRef]
- Galmés, J.; Flexas, J.; Savé, R.; Medrano, H. Water relations and stomatal characteristics of Mediterranean plants with different growth forms and leaf habits: Responses to water stress and recovery. Plant Soil 2007, 290, 139–155. [Google Scholar] [CrossRef]
- Chen, G.C.; Liu, Z.; Zhang, J.; Owens, G. Phytoaccumulation of copper in willow seedlings under different hydrological regimes. Ecol. Eng. 2012, 44, 285–289. [Google Scholar] [CrossRef]
- Cao, Y.; Ma, C.; Chen, G.; Zhang, J.; Xing, B. Physiological and biochemical responses of Salix integrathunb. under copper stress as affected by soil flooding. Environ. Pollut. 2017, 225, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Samarakoon, M.B.; Yagisawa, J. Effects of root architecture, physical tree characteristics, and soil shear strength on maximum resistive bending moment for overturning Salix babylonica, and Juglans ailanthifolia. Landsc. Ecol. Eng. 2012, 8, 69–79. [Google Scholar] [CrossRef]
- Davies, W.J.; Wilkinson, S.; Loveys, B. Stomatal control by chemical signalling and the exploitation of this mechanism to increase water use efficiency in agriculture. New Phytol. 2002, 153, 449–460. [Google Scholar] [CrossRef]
- Maud, V.; Smith, H.K.; David, C.; Jennifer, D.; Harriet, T.; Marijke, S.; Bastien, C.; Taylor, G. Adaptive mechanisms and genomic plasticity for drought tolerance identified in European black poplar (Populus nigral.). Tree Physiol. 2016, 36, 909–928. [Google Scholar]
- Wikberg, J.; Ogren, E. Variation in drought resistance, drought acclimation and water conservation in four willow cultivars used for biomass production. Tree Physiol. 2007, 27, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Podlaski, S.; Pietkiewicz, S.; Chołuj, D.; Horaczek, T.; Wiśniewski, G.; Gozdowski, D.; Kalaji, H.M. The relationship between the soil water storage and water-use efficiency of seven energy crops. Photosynthetica 2017, 55, 210–218. [Google Scholar] [CrossRef]
- Landhaüsser, S.M.; Stadt, K.J.; Lieffers, V.J. Photosynthetic strategies of summer green and evergreen understory herbs of the boreal mixed wood forest. Oecologia 1997, 112, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, G.S.; Van Rees, K.C.J. Soil organic carbon sequestration by shelterbelt agroforestry systems in Saskatchewan. Can. J. Soil Sci. 2017, 97, 394–409. [Google Scholar] [CrossRef]
- Kattge, J.; Knorr, W.; Raddatz, T.; Wirth, C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Glob. Chang. Biol. 2009, 15, 976–991. [Google Scholar] [CrossRef]
- Weih, M.; Bonosi, L.; Ghelardini, L. Optimizing nitrogen economy under drought: Increased leaf nitrogen is an acclimation to water stress in willow (Salix spp.). Ann. Bot. 2011, 108, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Feng, D.; Wang, Y.; Lu, T.; Zhang, Z.; Han, X. Proteomics analysis reveals a dynamic diurnal pattern of photosynthesis-related pathways in maize leaves. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Wang, L.; Zhou, Q.; Huang, X. Stomatal and non-stomatal factors regulated the photosynthesis of soybean seedlings in the present of exogenous bisphenol A. Ecotoxicol. Environ. Saf. 2017, 145, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W., III; Diaz, M.; Winter, K. Diurnal changes in photochemical efficiency, the reduction state of Q. radiationless energy dissipation, and non-photochemical fluorescence quenching in cacti exposed to natural sunlight in northern Venezuela. Oecologia 1989, 80, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Meng, Z.; Zhang, G.; Qi, M.; Sun, Z.; Liu, Y.; Li, T. Sub-high temperature and high light intensity induced irreversible inhibition on photosynthesis system of tomato plant (Solanum lycopersicum L.). Front. Plant Sci. 2017, 8, 182. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pan, X.L.; Hong, L. Responses of photosystem II (PSII) function in leaves and samaras of Ulmus pumila to chilling and freezing temperatures and subsequent recovery. Int. J. Agric. Biol. 2012, 14, 739–744. [Google Scholar]
- Kaipiainen, E.L. Parameters of photosynthesis light curve in Salix dasyclados and their changes during the growth season. Russ. J. Plant Physiol. 2009, 56, 445–453. [Google Scholar] [CrossRef]
- Ögren, E.; Sjöström, M. Estimation of the effect of photoinhibition on the carbon gain in leaves of a willow canopy. Planta 1990, 181, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Zhao, C.Y.; Li, J.; Yan, Y.Y.; Yu, B.; Han, M. Growth and leaf gas exchange in Populus euphratica across soil water and salinity gradients. Photosynthetica 2013, 51, 321–329. [Google Scholar] [CrossRef]
- Cai, Z.Q. Shade delayed flowering and decreased photosynthesis, growth and yield of Sacha Inchi (Plukenetia volubilis) plants. Ind. Crop. Prod. 2011, 34, 1235–1237. [Google Scholar] [CrossRef]
- Ďurkovič, J.; Čaňová, I.; Priwitzer, T.; Biroščíková, M.; Kapraľ, P.; Saniga, M. Field assessment of photosynthetic characteristics in Micropropagated and grafted Wych elm (Ulmus glabra, huds.) trees. Plant Cell Tiss. Organ Cult. 2010, 101, 221–228. [Google Scholar] [CrossRef]
- Gong, J.R.; Zhao, A.F.; Huang, Y.M.; Zhang, X.S.; Zhang, C.L. Water relations, gas exchange, photochemical efficiency, and Peroxidative stress of four plant species in the Heihe drainage basin of Northern China. Photosynthetica 2006, 44, 355–364. [Google Scholar] [CrossRef]
- Xue, W.; Li, X. Moderate shade environment facilitates establishment of desert phreatophytic species Alhagi sparsifolia, seedlings by enlarge fine root biomass. Acta Physiol. Plant 2017, 39, 7. [Google Scholar] [CrossRef]
- Liu, M.Z.; Jiang, G.M.; Li, Y.G.; Niu, S.L.; Cui, H.X. Gas exchange, photochemical efficiency, and leaf water potential in three Salix species. Photosynthetica 2003, 1, 393–398. [Google Scholar] [CrossRef]
- Contin, D.R.; Soriani, H.H.; Hernández, I.; Furriel, R.P.M.; Munné-Bosch, S.; Martinez, C.A. Antioxidant and photoprotective defenses in response to gradual water stress under low and high irradiance in two Malvaceae tree species used for tropical forest restoration. Trees 2014, 28, 1705–1722. [Google Scholar] [CrossRef]
- Liu, J.; Last, R.L. A chloroplast thylakoid lumen protein is required for proper photosynthetic acclimation of plants under fluctuating light environments. Proc. Natl. Acad. Sci. USA 2017, 114, 8110–8117. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Ungerer, P.; Zhang, H.; Ruban, A.V. Direct impact of the sustained decline in the photosystem ii efficiency upon plant productivity at different developmental stages. J. Plant Physiol. 2017, 212, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, L.; Zhang, X.; Korpelainen, H.; Li, C. Sexual differences in photosynthetic activity, ultrastructure and phytoremediation potential of Populus cathayana exposed to lead and drought. Tree Physiol. 2013, 33, 1043–1060. [Google Scholar] [CrossRef] [PubMed]
- Demmig–Adams, B.; Adams, W.W., III; Logan, B.A.; Verhoeven, A.S. Xanthophyll cycle dependent energy dissipation and flexible PSII efficiency in plants acclimated to light stress. Aust. J. Plant Physiol. 1995, 22, 249–261. [Google Scholar]
- Ding, L.; Wang, K.J.; Jiang, G.M.; Li, Y.G.; Jiang, C.D.; Liu, M.Z.; Niu, S.L.; Peng, Y. Diurnal variation of gas exchange, chlorophyll fluorescence, and xanthophyll cycle components of maize hybrids released in different years. Photosynthetica 2006, 44, 26–31. [Google Scholar] [CrossRef]
- Pierangelini, M.; Ryšánek, D.; Lang, I.; Adlassnig, W.; Holzinger, A. Terrestrial adaptation of green algae Klebsormidium and Zygnema (charophyta) involves diversity in photosynthetic traits but not in CO2 acquisition. Planta 2017, 246, 971–986. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V. Quantifying the efficiency of photoprotection. Phil. Trans. R. Soc. B 2017, 372, 1730. [Google Scholar] [CrossRef] [PubMed]
- Randriamanana, T.R.; Nissinen, K.; Moilanen, J.; Nybakken, L.; Julkunen-Tiitto, R. Long-term UV-B and temperature enhancements suggest that females of Salix myrsinifolia, plants are more tolerant to UV-B than males. Environ. Exp. Bot. 2014, 109, 296–305. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, G.; Guerra, C.; Jesus, C.; Amaral, J.; Santos, C. Effect of irradiance during acclimatization on content of proline and phytohormones in micropropagated Ulmus minor. Biol. Plant. 2013, 57, 769–772. [Google Scholar] [CrossRef]
Figure 1.
Photos of three species grown in the study site, Yarlung Zangbo River area, Tibet. (A) Study site; (B) Populus cathayana; (C) Ulmus pumila; (D) Salix longistamina.
Figure 1.
Photos of three species grown in the study site, Yarlung Zangbo River area, Tibet. (A) Study site; (B) Populus cathayana; (C) Ulmus pumila; (D) Salix longistamina.

Figure 2.
Diurnal course of (A) leaf water potential (Ψ), (B) vapor-pressure deficit (VPD), (C) transpiration rate (E), and (D) water-use efficiency (WUE) of P. cathayana, S. longistamina, and U. pumila. Each point represents the mean and standard error of five replicates.
Figure 2.
Diurnal course of (A) leaf water potential (Ψ), (B) vapor-pressure deficit (VPD), (C) transpiration rate (E), and (D) water-use efficiency (WUE) of P. cathayana, S. longistamina, and U. pumila. Each point represents the mean and standard error of five replicates.
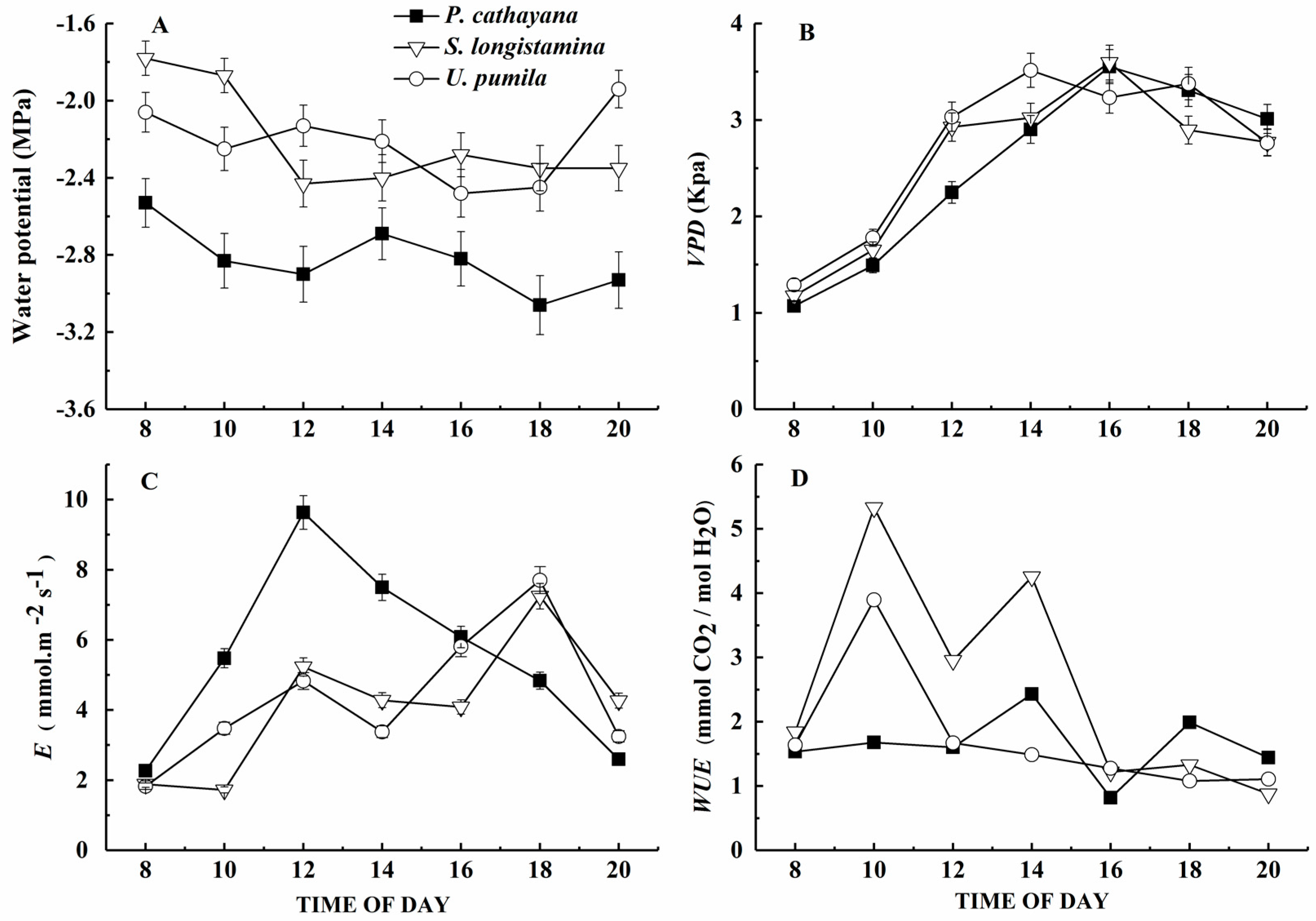
Figure 3.
Diurnal course of (A) net photosynthetic rate (Pn); (B) leaf stomatal conductance (gs); and (C) the internal CO2 concentration (Ci) in P. cathayana, S. longistamina, and U. pumila. Each point represents the mean and standard error of five replicates.
Figure 3.
Diurnal course of (A) net photosynthetic rate (Pn); (B) leaf stomatal conductance (gs); and (C) the internal CO2 concentration (Ci) in P. cathayana, S. longistamina, and U. pumila. Each point represents the mean and standard error of five replicates.
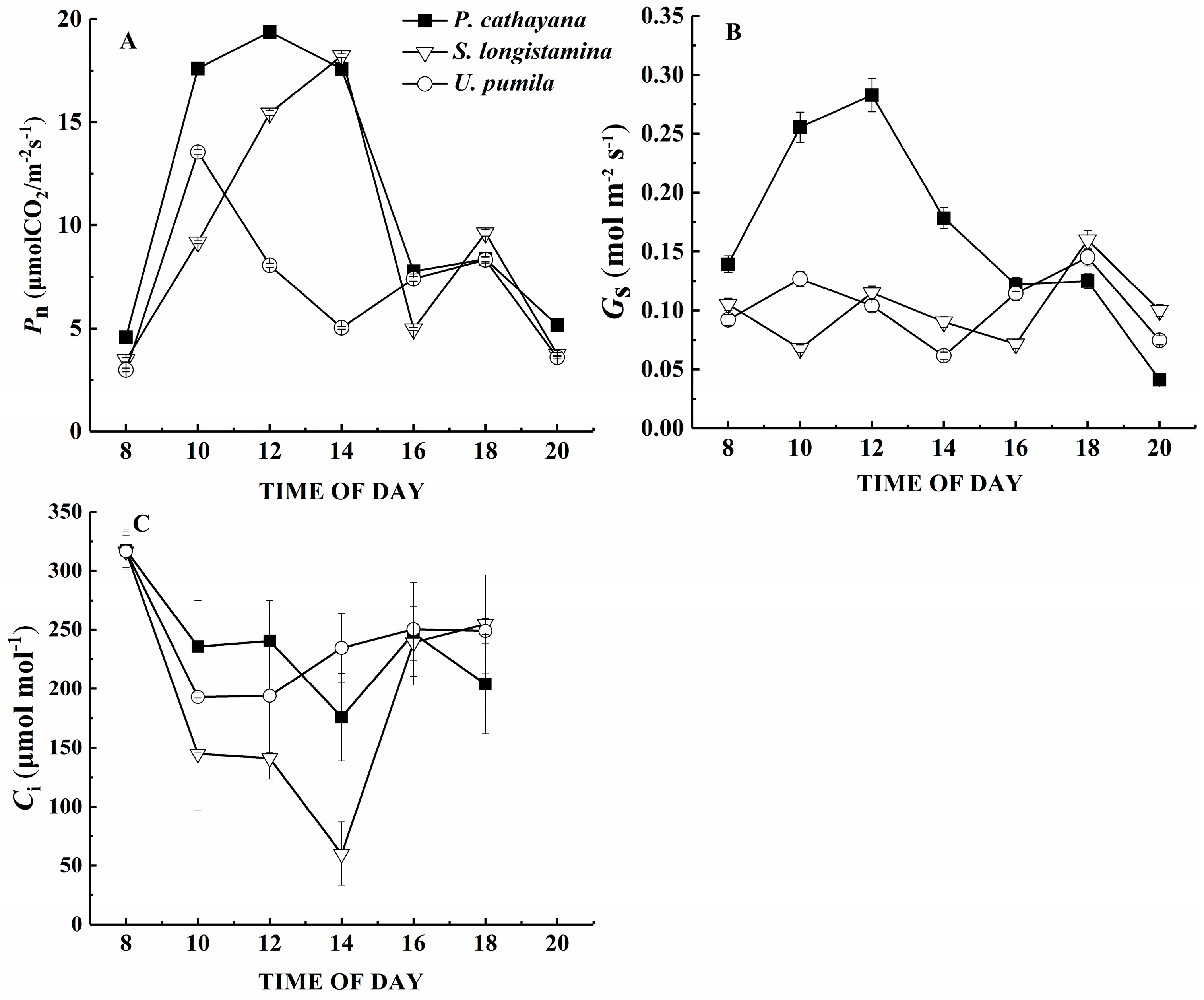
Figure 4.
The light saturation curves for net photosynthesis (Pn) as a function of irradiance (PPFD) for the leaves of P. cathayana, S. longistamina, and U. pumila. Values represent the means of five replicates.
Figure 4.
The light saturation curves for net photosynthesis (Pn) as a function of irradiance (PPFD) for the leaves of P. cathayana, S. longistamina, and U. pumila. Values represent the means of five replicates.

Figure 5.
Diurnal course of (A) potential quantum efficiency of photosystem II (Fv/Fm); (B) the maximum efficiency of photosystem II (Fv’/Fm’); (C) the quantum yield of photosystem II (ΦPSII); (D) the electron transport rate (ETR); (E) the degree of closure of the PSII reaction centers (1 − qp); and (F) non-photochemical quenching (NPQ). Values represent means and standard errors of five replicates.
Figure 5.
Diurnal course of (A) potential quantum efficiency of photosystem II (Fv/Fm); (B) the maximum efficiency of photosystem II (Fv’/Fm’); (C) the quantum yield of photosystem II (ΦPSII); (D) the electron transport rate (ETR); (E) the degree of closure of the PSII reaction centers (1 − qp); and (F) non-photochemical quenching (NPQ). Values represent means and standard errors of five replicates.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Soil water content of 4 depths in 3 stands at the study site.
| Species | Depth (cm) | |||
|---|---|---|---|---|
| 0–10 | 10–20 | 20–40 | 40–60 | |
| Populus cathayana | 2.78 | 1.94 | 2.32 | 1.38 |
| Ulmus pumila | 1.28 | 0.55 | 0.79 | 1.52 |
| Salix longistamina | 2.34 | 2.15 | 1.23 | 0.95 |
Table 2.
Morphological characteristics of the three species grown at the study site.
| Species | Survival Rates (%) | Crown Diameter (cm × cm) | Height (cm) | Total Biomass (g) |
|---|---|---|---|---|
| Populus cathayana | 83.83 ± 2.75 | 45.00 × 45.00 | 182.33 ± 3.18 | 381.41 ± 7.08 |
| Ulmus pumila | 93.80 ± 5.52 | 56.00 × 67.00 | 167.00 ± 5.77 | 209.61 ± 4.73 |
| Salix longistamina | 71.70 ± 3.06 | 78.00 × 63.00 | 122.75 ± 1.73 | 608.97 ± 29.94 |
Table 3.
Differences in photosynthetic parameters between P. cathayana, S. longistamina, and U. pumila: maximum photosynthetic rate (Pmax), light-saturation point (LSP), light-compensation point (LCP), dark respiration rate (Rd), and apparent quantum yield (AQY). Values represent means and standard errors of six replicates.
Table 3.
Differences in photosynthetic parameters between P. cathayana, S. longistamina, and U. pumila: maximum photosynthetic rate (Pmax), light-saturation point (LSP), light-compensation point (LCP), dark respiration rate (Rd), and apparent quantum yield (AQY). Values represent means and standard errors of six replicates.
| Species | Pmax (µmol CO2 m−2 s−1) | LSP (µmol m−2 s−1) | LCP (µmol m−2 s−1) | Rd (µmol m−2 s−1) | AQY (µmol CO2 µmol−1) |
|---|---|---|---|---|---|
| P. cathayana | 22.30 ± 2.14 | 1791.67 ± 33.29 | 39.89 ± 3.69 | 1.02 ± 0.09 | 0.0448 ± 0.0012 |
| S. longistamina | 12.80 ± 3.11 | 2290.00 ± 28.35 | 73.01 ± 4.54 | 3.31 ± 0.15 | 0.0454 ± 0.0014 |
| U. pumila | 14.90 ± 2.32 | 998.33 ± 13.58 | 26.35 ± 2.25 | 2.23 ± 0.12 | 0.0686 ± 0.0011 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gong, J.; Zhang, Z.; Zhang, C.; Zhang, J.; Ran, A. Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau. Forests 2018, 9, 48. https://doi.org/10.3390/f9020048
AMA Style
Gong J, Zhang Z, Zhang C, Zhang J, Ran A. Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau. Forests. 2018; 9(2):48. https://doi.org/10.3390/f9020048
Chicago/Turabian StyleGong, Jirui, Zihe Zhang, Chunlai Zhang, Jiaqiong Zhang, and An Ran. 2018. "Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau" Forests 9, no. 2: 48. https://doi.org/10.3390/f9020048
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.