Temperate and Tropical Forest Canopies are Already Functioning beyond Their Thermal Thresholds for Photosynthesis

1
School of Forest Resources & Environmental Science, Michigan Technological University, 1400 Townsend Dr., Houghton, MI 49931, USA
2
U.S. Geological Survey, Southwest Biological Science Center, 2290 S. West Resource Blvd, Moab, UT 84532, USA
3
USDA Forest Service, International Institute of Tropical Forestry, Jardín Botánico Sur, 1201 Calle Ceiba, Río Piedras, PR 00926-1115, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(1), 47; https://doi.org/10.3390/f9010047
Submission received: 21 December 2017
/
Revised: 15 January 2018
/
Accepted: 15 January 2018
/
Published: 22 January 2018
(This article belongs to the Special Issue Tree Ecophysiology: Responses and Adaptation to a Changing Environment)
Abstract
:Tropical tree species have evolved under very narrow temperature ranges compared to temperate forest species. Studies suggest that tropical trees may be more vulnerable to continued warming compared to temperate species, as tropical trees have shown declines in growth and photosynthesis at elevated temperatures. However, regional and global vegetation models lack the data needed to accurately represent such physiological responses to increased temperatures, especially for tropical forests. To address this need, we compared instantaneous photosynthetic temperature responses of mature canopy foliage, leaf temperatures, and air temperatures across vertical canopy gradients in three forest types: tropical wet, tropical moist, and temperate deciduous. Temperatures at which maximum photosynthesis occurred were greater in the tropical forests canopies than the temperate canopy (30 ± 0.3 °C vs. 27 ± 0.4 °C). However, contrary to expectations that tropical species would be functioning closer to threshold temperatures, photosynthetic temperature optima was exceeded by maximum daily leaf temperatures, resulting in sub-optimal rates of carbon assimilation for much of the day, especially in upper canopy foliage (>10 m). If trees are unable to thermally acclimate to projected elevated temperatures, these forests may shift from net carbon sinks to sources, with potentially dire implications to climate feedbacks and forest community composition.
1. Introduction
Temperate and tropical forests make up much of the world’s biomass, with tropical forests alone accounting for over 60% of terrestrial global carbon [1]. Forests can mitigate the effects of climate change, such as elevated temperature, through carbon dioxide (CO2) uptake during photosynthesis. However, global surface-air temperature is expected to increase by 1 to 5 °C by 2100, with an even greater increase expected in northern latitudes, along with an increase in the frequency and intensity of short-term heat waves [2,3]. As temperatures increase, there is a theoretical thermal tipping point after which photosynthesis begins to decline while plant respiration may still be increasing. If declines in CO2 uptake are severe enough that forest-wide respiration exceeds photosynthesis, forests could become net sources of carbon to the atmosphere [4,5]. Determining where this thermal threshold exists and whether different forest types are close to shifting from carbon sources to sinks is crucial to understanding and modeling global climate feedbacks [6].
Many global vegetation models are based on Farquhar’s model of photosynthesis [7], but our ability to accurately parameterize these models is hampered by a lack of mechanistic data on physiological responses to warming and thermal acclimation potential of canopy photosynthesis and respiration, especially for tropical forests [8,9,10,11,12,13,14,15,16]. Models analyzing the responses of tropical carbon storage to elevated temperature have produced inconsistent predictions on whether these forests will remain sinks or become sources [14,17]. In part, this is because there is great variability among tropical forests in annual temperature, precipitation, soil types, geographic range, and species assemblage; likely showing a variety of responses to changing climate across different systems [18,19,20]. Additionally, the greatest uncertainties in model projections of global carbon balance over the next century are for tropical latitudes, due to a combination of large carbon fluxes and a severe lack of relevant tropical-specific data [14]. Since tropical forests in particular account for such a large proportion of the global carbon cycle, any shifts in tropical photosynthesis could alter global carbon balances and feedbacks to climate change.
Though canopy warming studies will be crucial in determining whether different forest types can physiologically acclimate [9,11,14,21,22,23], instantaneous photosynthesis measurements in response to a range of temperatures are necessary to improve models and set a baseline to determine how close these forests are to the temperature optima for photosynthesis. Even in temperate forests, there is a scarcity of data of the short-term effects of elevated temperatures on photosynthesis in mature forest canopies and for long-term temperature acclimation [24].
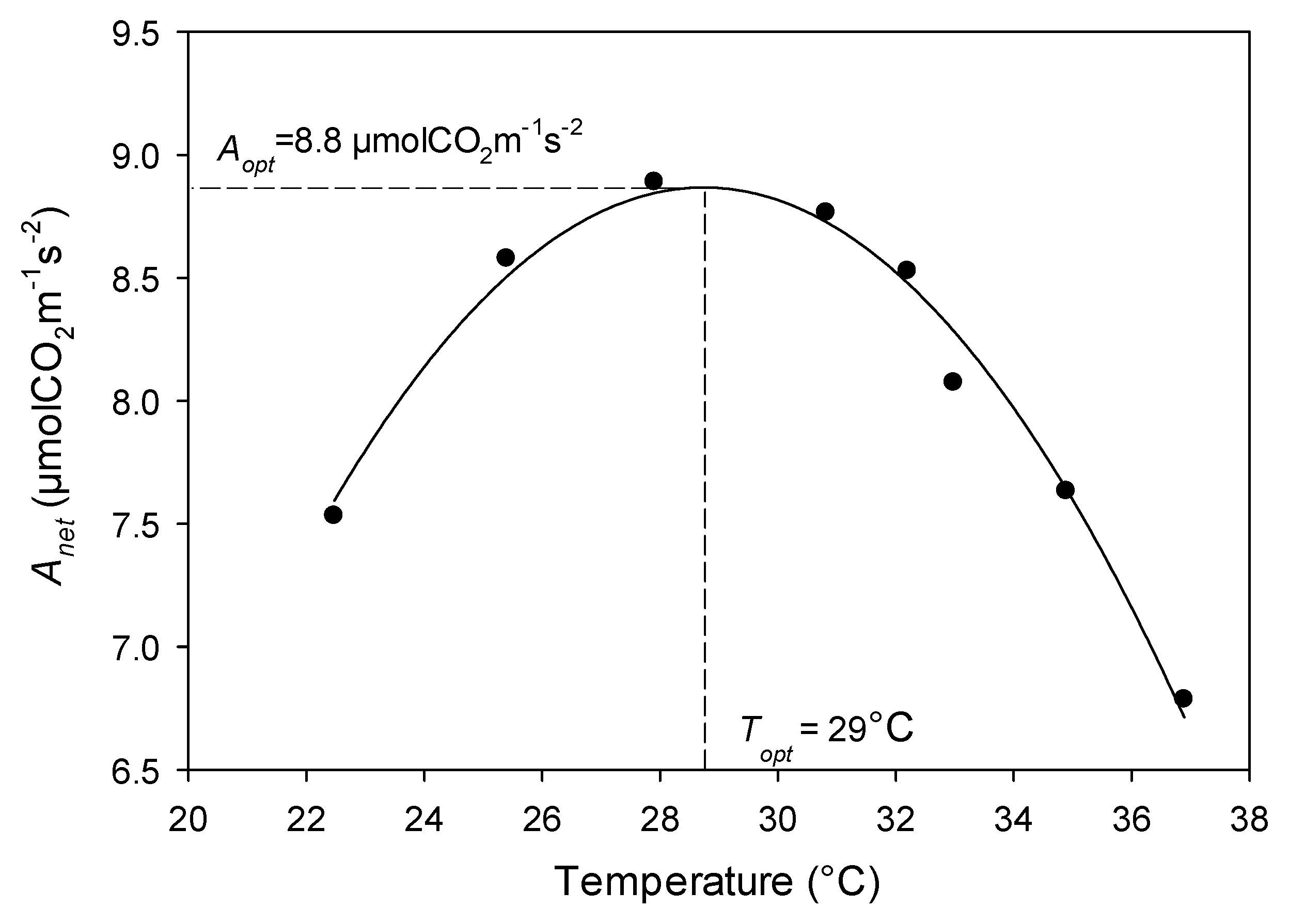
Light saturated photosynthetic response to temperature increases to a maximum (Aopt) at an optimum leaf temperature (Topt), then decreases again, following a parabolic curve (Figure 1) [7,25,26].
Below Topt, photosynthesis typically increases as the rates of biochemical reactions increase with temperature due to low-temperature limited enzymes [26,27]. In most temperate C3 species, photosynthetic decline occurs at leaf temperatures between 26 and 32 °C, due to direct biochemical limitations and/or to indirect stomatal responses to increases in vapor pressure deficit [7,26]. The two components of net photosynthesis commonly used for model parameterization are the rate of ribulose-1,5-bisphosphate (RuBP) carboxylation (Vcmax) and the rate of RuBP regeneration from the electron transport chain (Jmax) [7,28]. At elevated leaf temperatures, primary limitations to net photosynthesis include: temperature sensitivity to Ribulose-1,5-bisphosphate carboxylase oxygenase activase (Rubisco activase) and thylakoid membrane dysfunction, which affects efficiency of light reactions and electron transport [29,30], and increased rates of photorespiration due to the decrease of both specificity of Rubisco for CO2 and the relative solubility of CO2 versus O2 [31]. Irreversible leaf damage from high temperatures typically begins above 45 °C as plasma membranes in the chloroplast become permeable and proteins begin to denature [26].
Perhaps due to relatively low diurnal and inter-annual variability, tropical trees have been shown to be more susceptible to warming in comparison to temperate and boreal species, and have shown photosynthetic declines at temperatures of only 3 °C above mean ambient air temperatures [32,33]. Recent evidence suggests that lowland tropical forests, which are already consistently warm throughout the year, may already be operating close to their photosynthetic thermal optima, beyond which carbon uptake declines [5,22,34,35,36,37,38]. Due to greater diurnal, seasonal, and inter-annual variability in temperature in temperate ecosystems, the range of temperature from ambient to Topt will likely be greater in temperate species and therefore these high latitude species may not be operating as close to their thermal thresholds as tropical species [25,32,39,40,41].
For accurate model representation, it is important to measure photosynthesis across the entire vertical transect of the canopy, as upper and lower branches have different traits, are exposed to different environmental gradients, and can respond differently to temperature [42]. For example, light conditions are highly variable across a vertical gradient within canopies due to density of foliage limiting light access in the lower canopy [40,43], and light can be especially limiting in ecosystems with high annual cloud cover and mean annual precipitation (MAP), such as tropical rainforests [44]. As light availability typically increases with canopy height, leaf temperature does as well. The direct radiation hitting upper canopy leaves warms sun leaves more than diffuse scattered light in lower shaded leaves [45]. In many forests, leaf temperatures can be 1–7 °C above ambient air temperature, especially in the upper canopy [46]. Therefore, upper canopy leaves of both temperate and tropical species have been shown to have greater photosynthetic capacity, but they also may be more susceptible to heat stress due to elevated temperatures and vapor pressure deficit [47].
Various leaf functional traits can also be strong predictors of photosynthetic capacity and, in turn, photosynthetic response to elevated temperature. Leaf nitrogen (N) content in canopies has been found to correlate with greater photosynthetic capacity in various forest types [48,49,50], and leaf-area based phosphorus (P) has also shown similar patterns across upper canopy gradients as light-saturated rates of photosynthesis [51,52,53]. Additionally, leaf mass per area (LMA) is an extremely useful leaf trait that correlates with numerous within-canopy traits that are more challenging to measure (e.g., gas exchange, leaf nutrients) [54]. LMA typically increases with height in temperate and tropical forest canopies, either due to light or hydrostatic gradients [55,56,57] and since LMA has been found to have significant relationships with light saturated rates of photosynthesis (Amax), it may also be a strong predictor of maximum rates of photosynthesis under varying temperatures [12,56,58]. Thus, leaf P, N, and/or LMA may be able to predict photosynthetic temperature response parameters (Aopt and Topt). The highly weathered clay soils of tropical forests are often considered to be more limited by P while the younger temperate forests are generally considered to be more limited by N [59,60,61]. We may therefore expect P to be strongly correlated to photosynthetic parameters in the tropical forest with N more important in the temperate forest canopies [62,63,64].
Overall, this study sought to investigate how ambient leaf temperatures and instantaneous photosynthetic responses to temperature varied within mature tree canopies and across three forest types: temperate deciduous, tropical moist, and tropical wet, in order to see how close to their physiological temperature optima the dominant tree species of these forests may be operating. Our specific hypotheses were:
- (A)
- Compared to lower leaves, upper canopy leaves at all sites will have higher temperature optima for photosynthesis (Topt), higher maximum rates of photosynthesis (Aopt), and both parameters will increase with increasing light availability.
- (B)
- Topt and Aopt will increase with increasing mean annual temperatures across forest types, but tropical tree species will be functioning closer to photosynthetic thresholds than temperate species.
- (C)
- Topt and Aopt will increase with increasing LMA at all sites but will be better predicted by leaf N in the temperate site, and leaf P at the tropical sites.
2. Materials and Methods
Sampling took place from four canopy access towers at three sites: a temperate deciduous forest in Upper Peninsula Michigan and two sites in Puerto Rico. The Puerto Rico sites were defined as subtropical moist and subtropical wet forests, respectively by the Holdridge life zone system and both tropical forests were dominated by broadleaf evergreen tree species [19].
2.1. Temperate Deciduous Forest
The temperate deciduous study site was located at the Michigan Tech Ford Center and Forest near L’Anse, MI (46°64′ N, 88°48′ W). Mean annual temperature (MAT) was 5 °C, though mean annual growing temperature (MAGT) was 17.4 °C. Mean annual precipitation (MAP) was 879 mm from 2009 to 2011, with 401 mm occurring during the growing season, typically May through September [65]. The soil at the site was an Allouez gravelly coarse sandy loam, extending to a depth of 40 cm (http://websoilsurvey.sc.egov.usda.gov/). Acer saccharum was the dominant species on the site, making up 97% of total tree density, with only five species per hectare at the site. Other non-dominant species included Ulmus americana, Tilia americana, Betula alleghaniensis, and Ostrya virginiana. The average height of the canopy was 23 m, while the average height to live crown was 14 m [56]. For additional information about forest composition, structure, and site history, see Campione et al. [66].
Gas exchange and leaf trait data were acquired at the temperate site with a 19-m tall mobile aluminum walk-up tower (Upright, Inc., Selma, CA, USA), constructed in the summer of 2012.
2.2. Tropical Moist Forest
The tropical moist forest study site was located in El Tallonal Forest Reserve, a 114 ha privately owned natural area in the municipality of Arecibo, Puerto Rico (18°40′ N, 66°73′ W). The MAT was 25.5 °C and the MAP was 1295 mm (Table 1). The wet season at this site lasts from July to September, while the dry season is generally from January to March. This north central region of Puerto Rico is dominated by karst topography and has soils derived from limestone parent material with an overlay of Inceptisols in the low lying valleys [67]. There are an estimated 24 tree species per hectare at the site, and the dominant tree species is Castilla elastica [68]. The forest was estimated to be over 70 years old at the time of the study [68,69]. Historically, the area was used for grazing cattle and agriculture until 1950, when these lands were abandoned and naturally regenerated to forest [69].
Two identical 25 m steel towers, constructed 12 m apart in 2008, were used at the tropical moist forest site (BilJax, Archbold, OH, USA). C. elastica, Guarea guidonia, and Ocotea leucoxylon were measured from these towers. C. elastica is a fast growing, early successional broadleaved evergreen tree with relatively high LMA and light saturated photosynthetic rates (Amax) [70]. G. guidonia is a mid-successional deciduous tree native to Central and South America [71,72]. O. leucoxylon, which is also a Caribbean native, is an early successional broad-leaved evergreen tree [73].
Leaves of these three species were sampled from seven heights (7, 12, 14, 16, 20, 22, and 25 m) across both towers. Other species were accessible, however, they were not sampled if they were not present at a minimum of two heights.
2.3. Tropical Wet Forest
The tropical wet forest was located in the Luquillo Experimental Forest in northeastern Puerto Rico (18°31′ N, 65°74′ W) [19,74,75,76]. MAP of the site was 3936 mm and MAT is 25 °C, at an elevation of 361 m above sea level [74]. Temperature varies just 4 °C from month to month throughout the year with no days with temperatures below freezing. A drier season typically occurs from January through April, while the wettest period ranges from May to November [75]. The soils at the tropical wet forest are Ultisols, clayey poorly drained soil with red mottles [75]. This forest as a whole averages 10 tree species per hectare, with an average of 516 stems per ha [77,78]. The average height of the canopy was 20 m [79]. Dacryodes excelsa (common name: tabonuco), was the dominant tree species of the forest [80]. Dacryodes excelsa is one of the dominant native trees in Puerto Rico, growing at elevations between 200 to 800 m [81]. Other species located near the tower include: Prestoea montana, Casearia arborea, Inga laurina, Manilkara bidentata, and Sloanea berteriana [82]. For additional site history, see [75,83].
The tropical wet forest tower was a 24.7-m tall mobile aluminum walk-up built in 1991, with a footprint of 2.5 m2 (Upright, Inc., Selma, CA, USA) (Table 1). Dacryodes excelsa was the only species accessible across the entire vertical canopy gradient. Dacryodes excelsa is a long-lived broad-leaved evergreen species, and it is estimated that mature trees live up to 400 years with average heights of 30 to 35 m [81]. Dacryodes excelsa has historically shown relatively low light saturated photosynthesis, about 2.7 µmol CO2 m−2 s−1 [84]. Leaves were sampled from seven heights (8, 11, 15, 18, 20, 23, and 25 m).
2.4. Environmental Measurements
Diffuse non-interceptance (DIFN%), defined as diffuse light transmitted through the canopy as a proportion of incident (above canopy) radiation, was measured at each study site with an LAI-2200 (LI-COR Biosciences, Lincoln, NE, USA). DIFN% in forests is closely related to seasonally integrated photosynthetic photon flux density [85] and we use it here as a measurement of canopy openness and light availability integrated over time. ‘Above canopy’ measurements were taken at the top of the towers, and measurements were taken at every sampling height for each tower at each forest site. DIFN% was only sampled under uniformly cloudy conditions whenever possible at mid-day for all three sites, and a white diffuser cap was used to correct for direct sunlight when uniformly cloudy conditions were not present. DIFN% was measured on 1 June 2014 in Puerto Rico, and on 20 June 2014 in the temperate forest.
Leaf temperature was measured using a Fluke 572 infrared thermometer (Fluke Corporation, Everett, WA, USA). Five leaves of all accessible species were measured at every sampling height once an hour, for three days at all sites. Leaf temperature was measured from the 1–5 June and the 4–22 August at the tropical moist and tropical wet sites, and 8–16 August at the temperate deciduous site.
Air temperature was measured in thirty-minute increments and automatically logged from 26 May–27 August 2014 at the tropical moist and wet forests (HOBO Pro V2 temp/RH, Onset Computer Corporation, Cape Cod, MA, USA). Air temperature sensors in solar shields were tethered to the towers with zip ties at 5, 15, and 25 m at the tropical moist forest, and at 7, 13, 18, and 23 m at the tropical wet forest. Air temperature data were collected at the temperate deciduous forest from May to August of 2013 at heights 3, 8, and 11 m.
2.5. Sampling Design for Photosynthetic Measurements
In the temperate forest, photosynthetic temperature response curves were conducted on two leaves of Acer saccharum at every accessible height of the tower. Leaves were collected from four individual mature trees between 7 to 20 m, and from five individual understory saplings at less than 1 m in height. Photosynthetic light response curves were conducted on two healthy mature leaves at the top (20 m), middle (11 m), and understory (<1 m) to extract the light saturation point for use in temperature response curves. All physiological measurements were performed from the towers on intact leaves. All sampling from the temperate forest site took place during the growing season, from 7–21 July 2014 between 8 am and 4 pm.
In the tropical wet forest site, two mature leaves of Dacryodes excelsa were measured at every height for the temperature response curves from three accessible trees. Light response curves were taken on two healthy mature leaves at 25, 20, and 11 m to extract the light saturation point (the light concentration at which photosynthesis levels off). Sampling took place in two field campaigns: one from 6–9 June 2014 and another from 6–28 August 2014 between 8 am and 4 pm.
In the tropical moist site, two leaves of each species (C. elastica from four trees, G. guidonia two trees, and O. leucoxylon two trees) were sampled at each accessible height for the photosynthetic temperature response curves. C. elastica was sampled at 12, 14, 16, 20 and 22 m, G. guidonia at 8 and 12 m, and O. leucoxylon at 20, 22, and 25 m. The light saturation point used for these species was from a previous study [86]. Sampling took place between 9 am and 5 pm in two campaigns: one from 11–13 June 2014 and another from 13–22 August 2014.
2.6. Photosynthesis Measurements and Parameter Extractions
Photosynthesis measurements were taken with a LI-6400XT with a 6400-02B Red/Blue light source (LI-COR Biosciences, Lincoln, NE, USA). Light response curves were conducted to obtain the light saturation point (Isat) for each species, since it is a necessary parameter for the temperature response curves. Temperature response curves were conducted using a water jacket (6400-88 expanded temperature kit, Licor Biosciences, Lincoln, NE, USA) to increase and decrease leaf temperature. Leaf temperatures were achieved by introducing heated or cooled water through a Bev-A-Line tube through the water jacket and awaiting equilibrium at target temperatures. Water was continuously introduced through the system using gravity, with receptacles both above and below the measurement chamber. Using this method, we measured photosynthetic rates at 22, 24, 27, 29, 30, 31, 32, 33, and 35 °C. Relative humidity was kept between 50% and 60%, and was regulated by flow (200 umols−1 for temperate samples and 300 umols−1 for tropical samples) and desiccant. CO2 was kept at ambient concentrations of 400 ppm, and Photosynthetically active radiation (PAR)was kept at the average Isat for each species (800, 700, 1200, 1300, and 1200 µmol m−2 s−1 respectively for Dac, Cas, Acer, Oco, and Gua) as extracted from light response curves. Topt was calculated by taking the second order polynomial function on the temperature response curves and solving for the highest point on the x-axis, and Aopt was extracted by solving for the photosynthetic value from the regression equation at Topt (Figure 1) [87].
2.7. Leaf Traits
Leaf area for all leaves measured for gas exchange at the temperate site was measured using a leaf area meter (LI-3100 LI-COR Biosciences, Lincoln, NE, USA). The leaf area of leaves measured for gas exchanged at the tropical sites was measured by scanning pictures of leaves alongside a ruler (HP deskjet), then tracing the outline of the scanned image in ImageJ (Rasband, W.J 1997–2014, http://imagej.nih.gov/ij). Leaves were then dried for 24 h at 65 °C and weighed. Leaf mass per area (LMA; g m−2) was determined by taking the ratio of dry weight to leaf area. Samples were ground with a ball bearing grinder (8000 M Mixer/Mill, Spex Sample Prep, Metuchen, NJ, USA) for three minutes. Prior to elemental analysis, ground samples were dried for an additional 24 h at 65 °C, weighed with a Sartorius Cubis microbalance (Data Weighing Systems, Elk Grove, IL, USA) for 4–6 mg of leaf material, and folded in to 5 by 9 mm tin capsules. Leaf nitrogen (N) analysis was performed using an Elementar vario Microcube elemental analyzer (Elementar Inc., Hanau, Germany). Leaf phosphorus (P) analysis was analyzed with inductively coupled plasma optical emission spectrometry using a Thermo Jarrell Ash IRIS Advantage Inductively Coupled Plasma Optimal Emission Spectrometer (Precision Dynamics Corporation, San Fernando, CA, USA).
2.8. Data Analysis
An F-protected Least Significant Difference Test was performed to determine if means were significantly different among species and forest types. For all subsequent regression analyses, species data within tropical moist forest were pooled. Maximum daily leaf temperature (TLEAFmax), maximum daily air temperature (TAIRmax), and photosynthetic temperature optima (Topt) of leaves were each analyzed with height using simple linear regression. The points where TLEAFmax and Topt intersected were found by algebraically comparing where the coordinates of Topt = a1(height) + b1 and TLEAFmax = a2(height) + b2 intersected. Simple linear regression was also used to compare Topt and Aopt with foliar nutrients (Nmass, Narea, Pmass, Parea), and LMA. DIFN% was expressed as a percent of light reaching each canopy level graphically and was transformed with a natural log function for statistical analyses. All statistical analyses were performed in R (R Core Team, Vienna, Austria, 2013).
3. Results
3.1. Within-Canopy Variability of Photosynthetic Response to Temperature
Overall mean Topt values were 2.5 °C greater for both tropical forests than for the temperate forest (30 °C and 27.5 °C, respectively; Table 1). Aopt, however, was highest in the tropical moist forest (9.8 µmol CO2 m−1 s−2), followed by the temperate forest (7.3 µmol CO2 m−1 s−2), and lastly, the tropical wet forest (4.9 µmol CO2 m−1 s−2).
High photosynthetic rates in the pooled mean of the tropical moist forest were mainly driven by high rates of O. leucoxylon, 11.5 µmol CO2 m−1 s−2, compared to the next highest values by species: Castilla elastica at 9.9 µmol CO2 m−1 s−2 and Acer saccharum at 7.3 µmol CO2 m−1 s−2 (Table 2).
3.2. Relationships between Photosynthetic Temperature Optima and Maximum Leaf and Air Temperatures
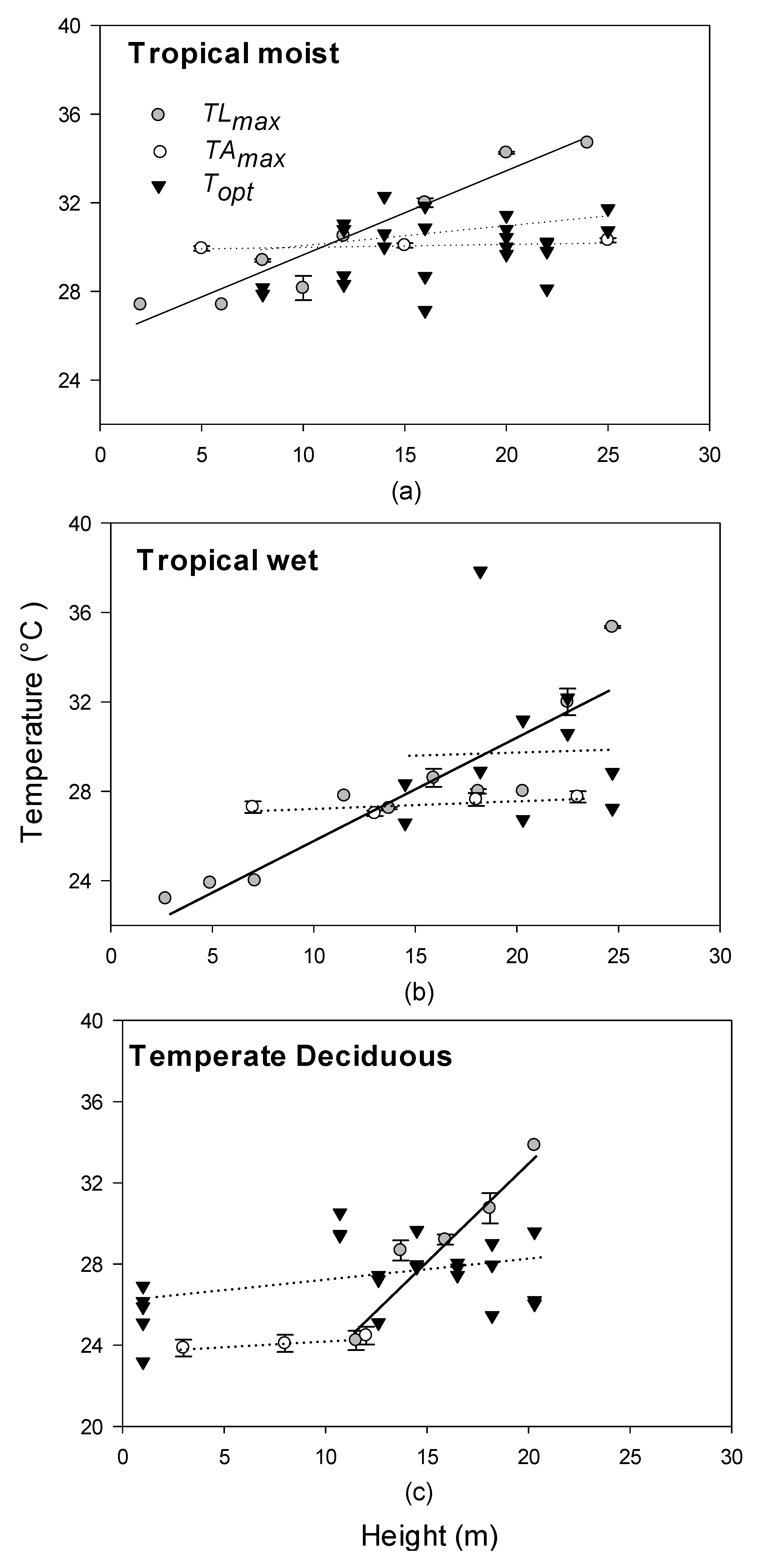
In all forest types, the relationship between TLEAFmax and height was significant and showed a steep positive increase. TLEAFmax spanned 8–10 °C in all forests from understory to canopy top (Figure 4).
TAIRmax, however, only increased significantly with height in the tropical wet forest, and slopes were relatively shallow for all forest types. We only had upper canopy air temperature data in the tropical moist forest, where TLEAFmax exceeded maximum air temperatures by 4 °C at heights of 25 m (Figure 4a).
TLEAFmax exceeded temperatures Topt in the upper canopy in all forest types, but the heights and temperatures where this occurred differed for each forest canopy type (Figure 4). In the tropical moist site, Topt exceeded TLEAFmax at 29 °C, above 11 m height (Figure 4a), whereas at the tropical wet site, Topt intersected at 22 m at a higher temperature of 31 °C (Figure 4b). At the temperate site, Topt and TLEAFmax intersected at a lower height of 11 m, and at a temperature of 28 °C (Figure 4c), indicating that mid-canopy leaf temperature is already exceeding Topt.
3.3. Variation in Leaf Traits with Forest Type and across Vertical Canopy Gradients
Mean LMA was different across each forest type, where the temperate forest mean LMA was lowest (44 g m−2), and the tropical wet forest mean LMA was the highest (122 g m−2) (Table 3).
Compared to the temperate deciduous forest, the tropical wet forest showed steeper increases of LMA in response to light. The tropical moist forest had the highest mean Narea (2.4 g m−2), followed by the tropical wet forest (1.7 g m−2), then the temperate deciduous forest (0.9 g m−2), while at the species level O. leucoxylon had the greatest mean Narea (2.9 g m−2). Nmass differed significantly among each forest type as well, with the moist forest also showing the highest mean Nmass (25 mg g−1), but with the lowest mean Nmass in the tropical wet forest (14 mg g−1, Table 2). At the species level, C. elastica and G. guidonia had the highest mean values of Nmass (25 mg g−1 and 29 mg g−1, respectively, Table 2). Narea increased linearly with height in all forest types. The relationship between Nmass and height was not significant at any site (Table 4).
Narea showed a strong correlation with light only in the temperate forest (p ≤ 0.001, Table 5), increasing log linearly, and was not significant in any other forest type. In the tropical wet forest, Nmass decreased with light and was the only forest type to show any significance between these variables (p = 0.05, Table 5).
The tropical moist forest had the highest mean Parea (0.11 g m−2), differing significantly from the two other forest types (0.09 and 0.08 g m−2 in the wet and temperate forests, respectively, Table 2). The temperate deciduous forest had the greatest mean Pmass, followed by the tropical moist and wet forests (1.7, 1.4, and 0.8 mg g−1, respectively, Table 2). A. saccharum showed the lowest value of Parea (0.08 g m−2), but had one of the highest values of Pmass (1.7 mg g−1) along with G. guidonia (1.9 mg g−1; Table 2). Parea increased linearly with height in the temperate deciduous forest only, while Pmass decreased with height in the tropical moist forest only (Table 4). Neither Parea nor Pmass responded to light environment in any forest type (Table 5).
3.4. Relationships between Photosynthetic Temperature Response Parameters and Leaf Functional Traits
4. Discussion
4.1. Effects of Height and Light on Photosynthetic-Temperature Response Curves
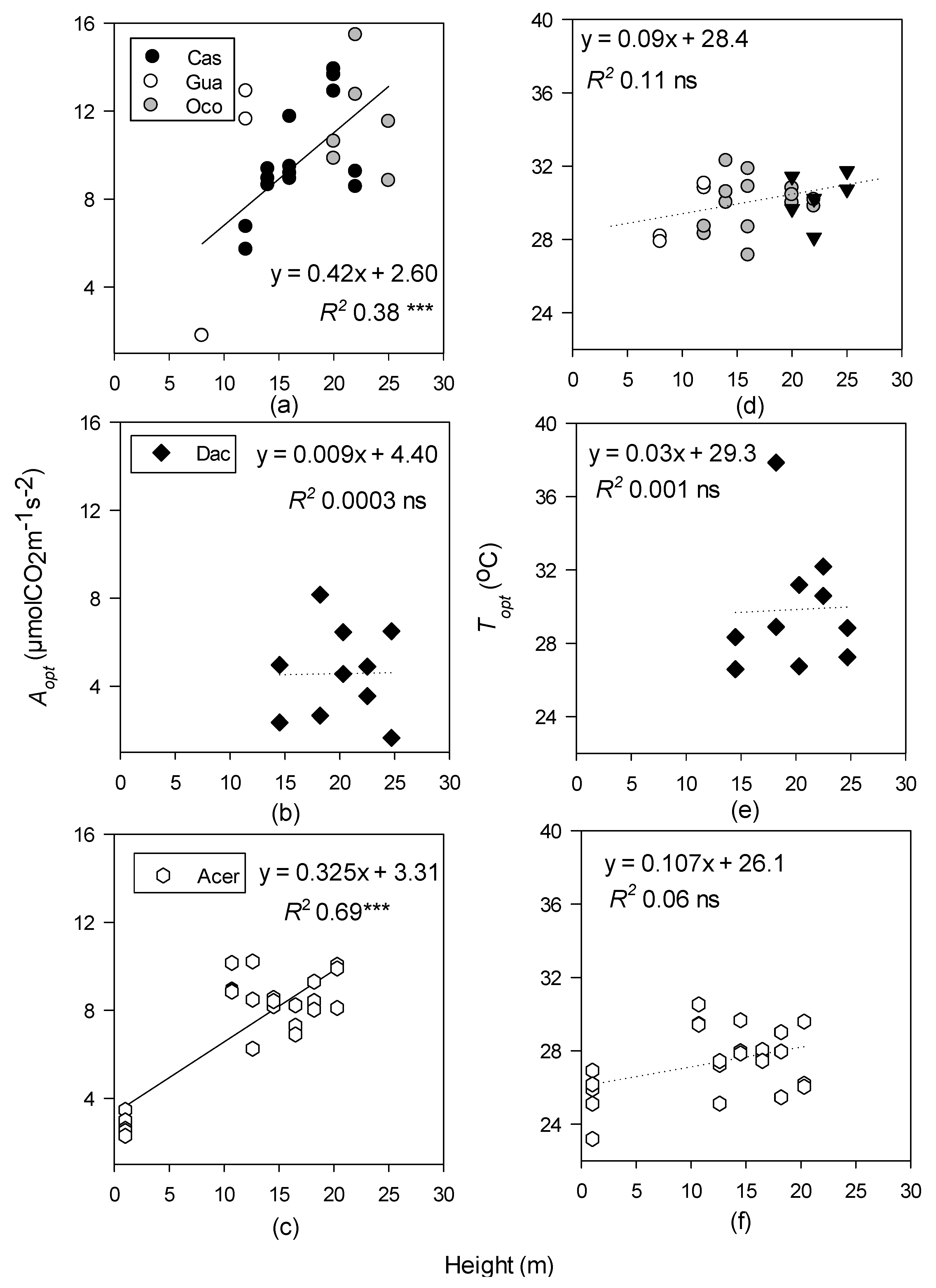
As expected, optimum photosynthetic rates (Aopt) increased with canopy height in the temperate deciduous and tropical moist forests, but not the wet forest (Figure 2a–c). This pattern could be related to the fact that the temperate and tropical moist forests had the greatest sampling range of heights, though it is also possible that height is not as significant for photosynthetic rates in the wet forest, potentially since D. excelsa is an evergreen species. Aopt did show a positive response to increasing light environment in all three forest types (Figure 3). Aopt has been correlated with light saturated rates of photosynthesis (Amax), and similar patterns have been observed in other studies [12,88]. Light-saturated upper canopy leaves typically correspond with greater photosynthetic optima in many tropical and temperate species, though some studies have also found that photosynthesis may peak near 10 m in the canopy and then decline higher in the canopy [58,89,90].
Topt was much more conservative across height and light gradients than Aopt for all forest types. Although there was a slight trend toward higher Topt in the upper canopy (Figure 4), the Topt did not show a significant increase with height in any forest type, contrary to expectations (Figure 2d–f). This could be a result of the sampling methods and the lack of accessible leaves below 10 m, especially in the tropical forests.
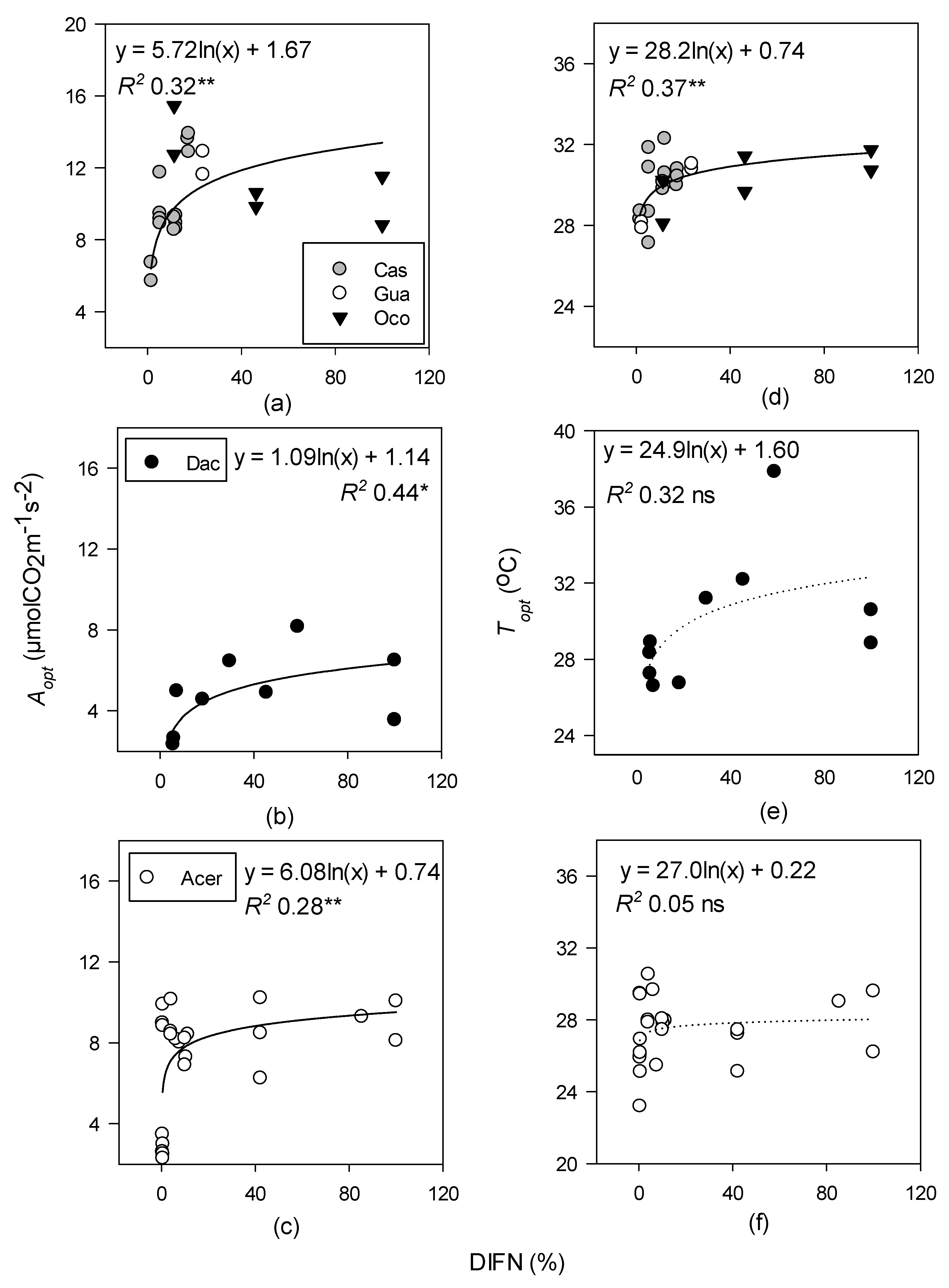
There is a scarcity of temperature response curve data that span vertical canopy gradients in all forested ecosystems [35,91,92]. Overall, we have found that neither height nor light were reliable predictors of Topt, except in the tropical moist forest, which showed a log-linear relationship with light (DIFN%) (Figure 3d). Light availability is considered one of the limiting factors for photosynthesis of tropical forest species, therefore it was surprising to find that there was a significant relationship with Aopt and DIFN% in the tropical moist and temperate deciduous forests, but not the tropical wet forest, though this could be due to the lack of measurable leaves in the sub-canopy (Figure 3a–c) [28,93]. Leaves in the upper canopy have been found to be near their photosynthetic optima, and any increases in temperature can induce stomatal closure, leading to a decline in photosynthesis (Figure 8a,b) [5,94].
4.2. Relationships between Optimal Temperatures for Photosynthesis, Leaf Temperature, and Air Temperature across Different Forest Types
As predicted, both tropical forests had higher Topt (30 °C) than the temperate forest (27 °C; Table 2). This was expected, since Topt typically corresponds with growth temperature, and mean growing season temperature in the wet tropical and moist tropical forests is more than 7 °C higher than the temperate deciduous forest [26,39]. This is consistent with data showing that temperate C3 species have Topt ranges between 20 and 35 °C, peaking in the 30–35 °C range, while species adapted to warmer growing conditions, such as tropical trees, can exhibit Topt values well above 35 °C [29]. The Topt range of our study for tropical species remained near 30 °C (Table 2).
Unlike Topt, tropical forests do not necessarily always have greater Aopt compared to temperate forests [58,95,96], as our study displayed. In the case of Dacryodes excelsa at the wet tropical field site, Aopt was significantly lower (4.9 µmol CO2 m−1 s−2) than that of the tropical moist species (average 9.8 µmol CO2 m−1 s−2) and Acer saccharum (7.3 µmol CO2 m−1 s−2) in the temperate forest (Table 2). Even though Aopt was much lower for our study’s tropical wet species than the temperate forest, due to the ability to photosynthesize year-round, net canopy photosynthesis may actually be higher in tropical forests [38,97,98].
When maximum daily leaf temperatures (TLEAFmax) exceed Topt, this indicates that photosynthesis is suppressed for at least a portion of the day due to elevated temperatures. It was hypothesized that the tropical sites would be operating closer to thermal thresholds [5], and thus show leaf temperatures closer to Topt than the temperate forest. What we found, however, is that Topt exceeded TLEAFmax in the upper canopies of all three sites. The intersection between maximum daily leaf temperatures in upper canopies and Topt was 10 m lower in the tropical moist forest than the tropical wet forest (Table 1, Figure 4b,c). The tropical moist forest canopy was operating above Topt at a lower canopy position than the wet tropical forest, which indicates that carbon uptake in the hotter, drier forest may be more susceptible to further climate-change related warming.
Leaf temperatures showed steeper increases with height than air temperatures, which did not change significantly with height in any forest (Figure 4). The differences between leaf and air temperature increased with height, with TAIRmax exceeding TLEAFmax in the lower canopy and TLEAFmax exceeding TAIRmax in the mid and upper canopy. In other studies, leaf temperature has been found to be 1–7 °C greater than corresponding air temperature, and this difference typically increases with height [46]. The greatest disparity in leaf and air temperature was in the temperate forest (about 12 °C, Figure 4). Both tropical forests had higher average leaf and air temperatures compared to the temperate forest (Figure 4). There was an average of 2 °C difference between leaf and air temperature in the tropical wet forest, a 1 °C difference in the moist forest, and a 4 °C difference in the temperate forest (Figure 4).
Temperate and tropical forests already currently experience mid-day declines in photosynthesis due to elevated temperature [46,99]. Upper canopies, which already have higher leaf and air temperature, are also much more likely to exhibit negative reactions to increases in temperature, since they are already operating near maximum rates of photosynthesis in ambient conditions [94]. The leaf temperature in upper canopies can also be up to 10 °C greater than air temperature, and leaf temperature has greater association with stomatal closure, one of the short-term causes for declines in photosynthesis [46,100].
Our study found that the temperate site showed similar positive relationships between Topt and TLEAFmax along a vertical gradient as the tropical moist site, which indicates that some temperate forests may be just as susceptible to declines in photosynthesis with rising air temperature as tropical forests, contrary to our overarching hypotheses. According to the United States Department of Agriculture Forest Service Climate Change Atlas (https://www.fs.fed.us/nrs/atlas/) and other sources, the abundance of species such as A. saccharum is expected to decline in Northern Michigan as the climate changes [99,101]. This, combined with our findings, could indicate not only a decline of A. saccharum abundance, but also a potential decline in carbon assimilation of northern hardwood forests.
Tropical trees have been shown to be more susceptible to warming in comparison to temperate and boreal species, with photosynthetic declines at temperatures just 3 °C above mean ambient air temperatures [32,33]. Due to the relative consistency of tropical lowland temperature over short and long timescales, tropical species are perhaps more likely to show strong species shifts in response to climate change, instead of an ability for individual trees to acclimate [14,17,21,102]. In particular, tropical tree species may not be able to photosynthetically acclimate to elevated leaf temperature due to the limited range of diurnal, seasonal, and inter-annual temperatures under which they have evolved [14,41]. However, there is a dearth of ecophysiological data in tropical ecosystems from warming experiments [14], and mature tree canopy warming experiments are rare in any latitude [21,22]. Accordingly, our understanding of tropical responses to warming remains limited. We acknowledge the limitations of extrapolating leaf-level results from single species to the forest level, however, the species sampled here were dominant in each forest type (e.g., Acer saccharum represented 90% of tree density), and these secondary tropical forests maintain relatively low levels of tree species diversity in comparison to other lowland tropical forests. Additionally, our data are critical to help fill data gaps in the physiological processes of tropical forest canopies [6].
4.3. Leaf Functional Traits
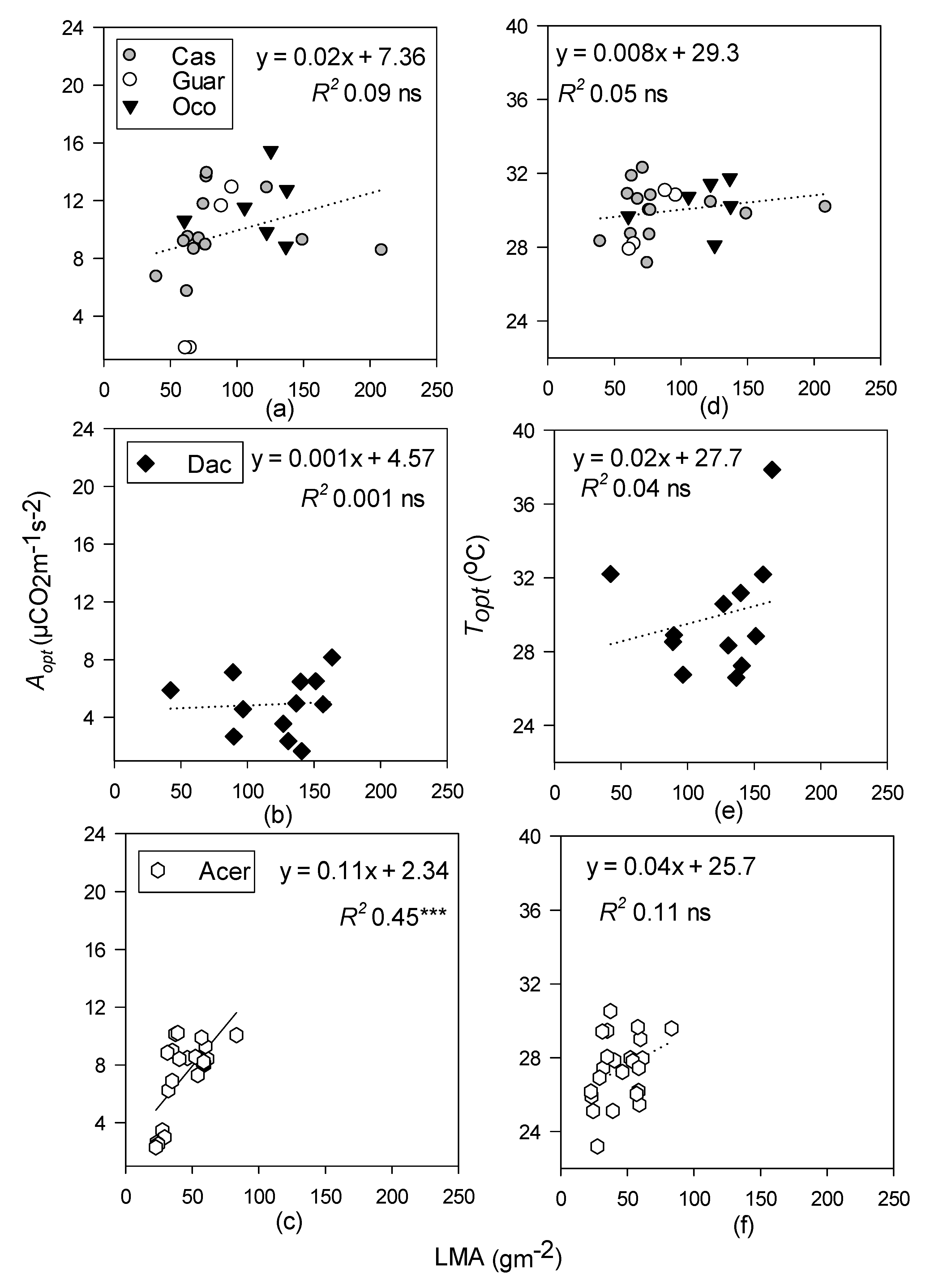
LMA was only a significant predictor of Topt and Aopt in the temperate forest (Figure 5). LMA increases with canopy height, regardless of the forest type [55], and strong correlations between LMA, height, and photosynthetic capacity have been found within A. saccharum canopies specifically [55,56,103,104]. Tropical forest LMA did not increase with Aopt, possibly because there was insufficient sub-canopy data in the tropical sites to get a broad gradient for LMA and photosynthetic capacity.
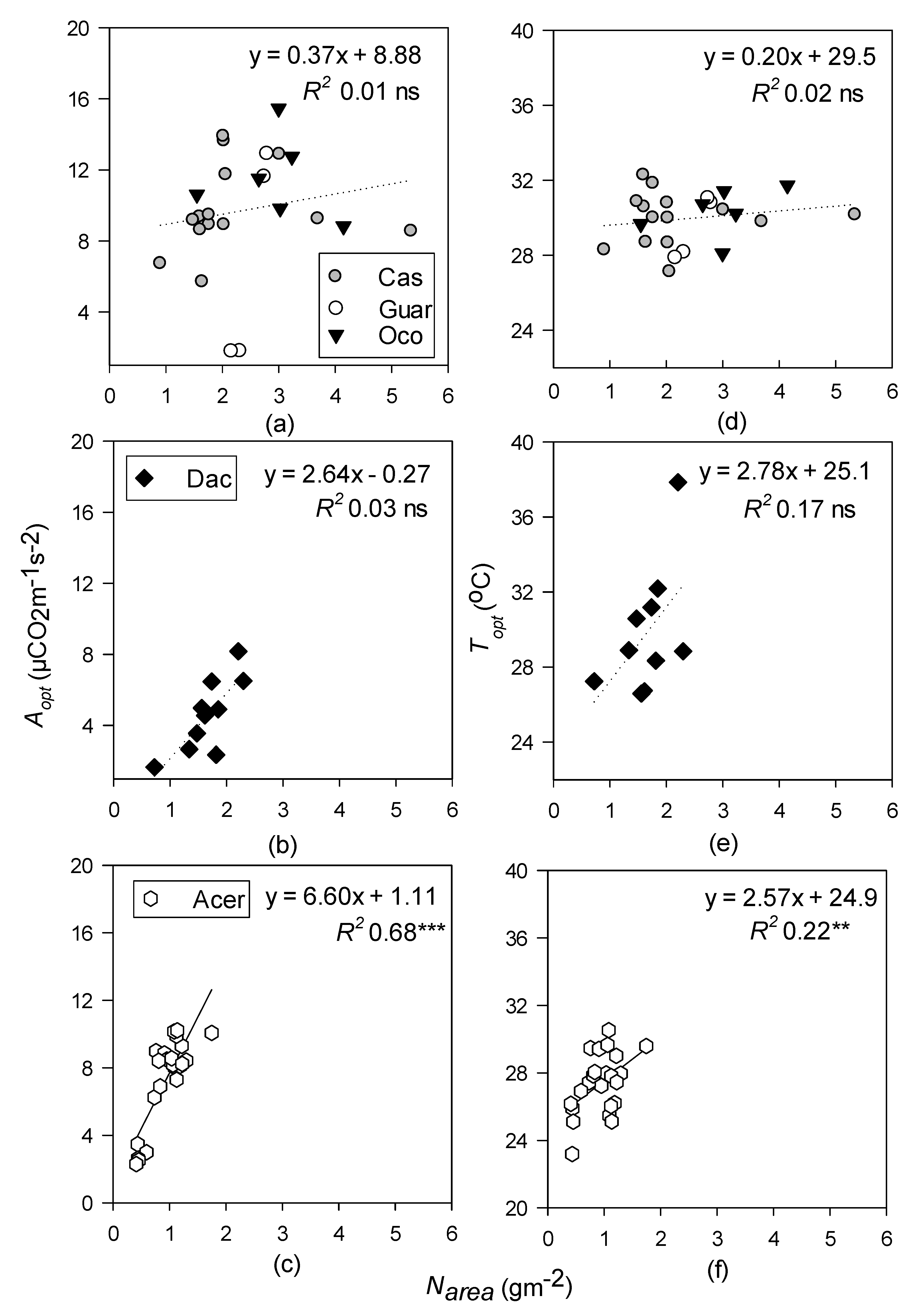
Nitrogen had stronger correlations with photosynthetic parameters on a leaf area basis than on the mass basis, which differs from other findings [50,105], and, as predicted, the relationship was strongest in the temperate forest compared to the tropical forests. Given that temperate forests are typically considered more N limited than that of tropical forests [105], the stronger relationship between photosynthetic parameters and N in this forest was in line with expectations [96]. While Narea was significantly related to the Aopt of both the tropical wet and temperate deciduous forests, the R2 was much lower (0.32 and 0.68, respectively) in the wet forest than that of the temperate forest, indicating a stronger relationship between N and photosynthesis in the temperate forest than the tropical ones (Figure 6). The tropical moist forest had a very low R2 and no significance between N and Aopt and Topt, which may also be due to variability in the relationships across the three species pooled, compared to the singular species sampled at the other sites. Very young or very old tropical forests can also have N limitations for photosynthesis, which may explain why the wet forest, which was found in the older of the two sampled tropical forests, also had a significant relationship with Aopt and Narea [106].
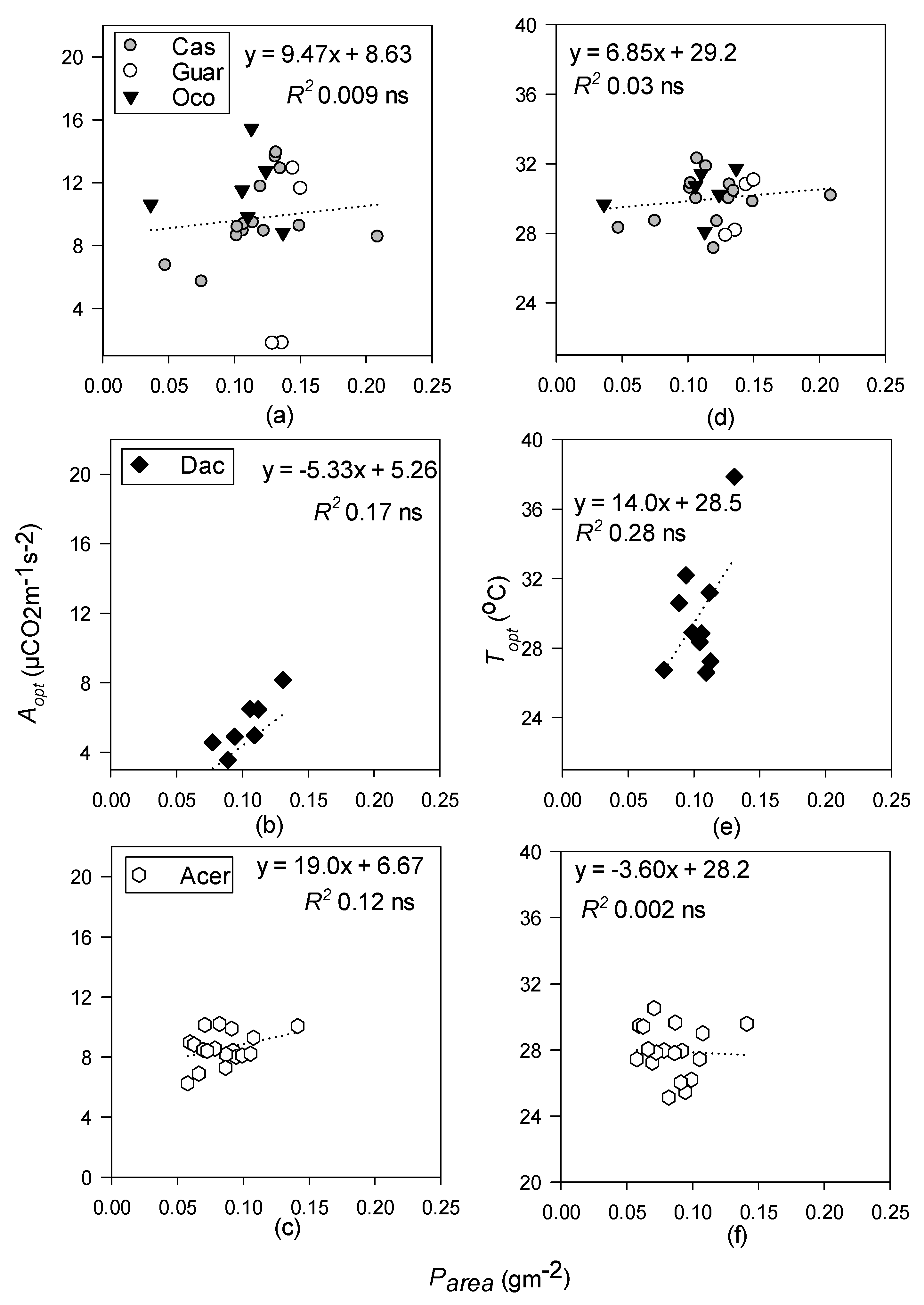
Concentrations of P per unit leaf area were not related to either Topt or Aopt for any forest type, rejecting our hypothesis that it would be significant in tropical forests. The highest Parea values were in the tropical moist forest, while Pmass was highest in the temperate forest (Table 3). The latter was expected, since P is an important substrate in photosynthetic metabolism and RuBP regeneration and is generally considered to be less limiting in temperate forests than that of tropical ecosystems [62,63,107,108].
Other studies have found significant correlations between photosynthetic capacity and leaf functional traits, yet relationships between Topt and these traits have rarely been measured [58,95,96]. Contrary to our results, studies in tropical forests have shown significant relationships between leaf P and photosynthesis, while temperate forests often show relationships between photosynthesis and leaf N [97,109,110].
5. Conclusions
In the upper canopy of all forest types, TLEAFmax was found to exceed the temperature optima for photosynthesis, indicating that leaves in the upper canopies of these forests are already showing declines as a result of high temperatures. The tropical moist forest, which had the highest mean annual temperature of all sites, had the greatest amount of canopy operating above the thermal optima. Contrary to expectations, the thermal optima for photosynthesis exceeded maximum leaf temperature in the temperate forest, more so than the tropical wet forest. Unless forest canopies can acclimate to increasing temperature and shift thermal optima for photosynthesis higher, a majority of the canopy of each site could be operating above the thermal optima by 2100, leading to potential global decreases in carbon storage.
As seen in previous studies, LMA and N (on an area and mass basis) were significant predictors of Aopt for the temperate forest and showed a strong correlation with Topt as well. The relationships in both tropical forests, however, were not as pronounced. Contrary to predictions, P was not significantly related to Aopt or Topt in either tropical forest. This could indicate that N and P were not limiting in the respective forests, though additional sampling could further elaborate on this. For future studies, more data on the mechanistic responses of photosynthesis will be needed, such as rates of electron transport and RuBP carboxylation, to determine what is limiting. In situ warming experiments could prove to be very useful in determining if acclimation to temperature does occur and to what extent. While additional data for model parameterization are needed, atmosphere-biosphere models are only as accurate as their underlying species-specific or plant functional type-specific physiological data, and different species show different thermal tolerances [8]. This could be critical for atmosphere-biosphere models and improving parameterization and for understanding changes in carbon flux due to climate change.
Acknowledgements
This research was supported by USDA Forest Service awards to SCR (13-IA-11120101-008), MAC (13-JV-11120101-033) and TEW (11-DG-11120101-012) and the U.S. Department of Energy Office of Science, Office of Biological and Environmental Research Terrestrial Ecosystem Sciences Program, under Award Numbers DE-SC-0012000 and DE-SC-0011806. We are grateful to Omar Gutierrez del Arroyo, Adam Coble, Mickey Jarvi, and Sarah Harttung for volunteering their time to help in the field. We are grateful to comments from Dave Hoover and two anonymous reviewers, which significantly improved the paper. This research was supported by USDA Forest Service awards to SCR (13-IA-11120101-008), MAC (13-JV-11120101-033) and TEW (11-DG-11120101-012). Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author Contributions
M.A.C., S.C.R., and T.E.W. conceived and designed the experiments; A.C.M. performed the experiments; A.C.M. and M.A.C. analyzed the data; M.A.C., T.E.W., and S.C.R. contributed reagents/materials/analysis tools; A.C.M. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pan, Y.D.; Birdsey, R.A.; Phillips, O.L.; Jackson, R.B. The structure, distribution, and biomass of the world’s forests. Ann. Rev. Ecol. Evol. Syst. 2013, 44, 593–622. [Google Scholar] [CrossRef]
- Kirtman, B.P.; Min, D.; Infanti, J.M.; Kinter, J.L., III; Paolino, D.A.; Zhang, Q.; van den Dool, H.; Saha, S.; Mendez, M.P.; Becker, E. The North American Multi-Model Ensemble (NMME): Phase-1 seasonal to interannual prediction, phase-2 toward developing intra-seasonal prediction. Bull. Am. Meteorol. Soc. 2013, 95, 595–601. [Google Scholar]
- Alexander, L.; Allen, S.; Bindoff, N.; Bréon, F.; Church, J.; Cubasch, U.; Emori, S.; Forster, P.; Friedlingstein, P.; Gillett, N. Working group I Contribution to the IPCC Fifth Assessment Report: The Physical Science Basis Summary. In Climate Change; IPCC: Bern, Switzerland, 2013. [Google Scholar]
- Wood, T.E.; Cavaleri, M.A.; Reed, S.C. Tropical forest carbon balance in a warmer world: A critical review spanning microbial-to ecosystem-scale processes. Biol. Rev. 2012, 87, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.E.; Goulden, M.L. Are tropical forests near a high temperature threshold? J. Geophys. Res. Biogeosci. 2008, 113. [Google Scholar] [CrossRef]
- Randerson, J.T.; Hoffman, F.M.; Thornton, P.E.; Mahowald, N.M.; Lindsay, K.; LEE, Y.H.; Nevison, C.D.; Doney, S.C.; Bonan, G.; Stöckli, R. Systematic assessment of terrestrial biogeochemistry in coupled climate—Carbon models. Glob. Chang. Biol. 2009, 15, 2462–2484. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Medlyn, B.; Dreyer, E.; Ellsworth, D.; Forstreuter, M.; Harley, P.; Kirschbaum, M.; Le Roux, X.; Montpied, P.; Strassemeyer, J.; Walcroft, A. Temperature response of parameters of a biochemically based model of photosynthesis. II. A review of experimental data. Plant Cell Environ. 2002, 25, 1167–1179. [Google Scholar] [CrossRef]
- Gonzalez-Meler, M.A.; Rucks, J.S.; Aubanell, G. Mechanistic insights on the responses of plant and ecosystem gas exchange to global environmental change: Lessons from Biosphere 2. Plant Sci. 2014, 226, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Atkin, O.; Atkinson, L.; Fisher, R.; Campbell, C.; Zaragoza Castells, J. Using temperature-dependent changes in leaf scaling relationships to quantitatively account for thermal acclimation of respiration in a coupled global climate-vegetation model. Glob. Chang. Biol. 2008, 14, 2709–2726. [Google Scholar] [CrossRef]
- Smith, N.; Dukes, J. Plant respiration and photosynthesis in global-scale models: Incorporating acclimation to temperature and CO2. Glob. Chang. Biol. 2013, 19, 45–63. [Google Scholar] [CrossRef] [PubMed]
- Weerasinghe, L.K.; Creek, D.; Crous, K.Y.; Xiang, S.; Liddell, M.J.; Turnbull, M.H.; Atkin, O.K. Canopy position affects the relationships between leaf respiration and associated traits in a tropical rainforest in Far North Queensland. Tree Physiol. 2014, 34, 564–584. [Google Scholar] [CrossRef] [PubMed]
- Huntingford, C.; Zelazowski, P.; Galbraith, D.; Mercado, L.M.; Sitch, S.; Fisher, R.; Lomas, M.; Walker, A.P.; Jones, C.D.; Booth, B.B. Simulated resilience of tropical rainforests to CO2-induced climate change. Nat. Geosci. 2013, 6, 268–273. [Google Scholar] [CrossRef]
- Cavaleri, M.A.; Reed, S.C.; Smith, W.K.; Wood, T.E. Urgent need for warming experiments in tropical forests. Glob. Chang. Biol. 2015, 21, 2111–2121. [Google Scholar] [CrossRef] [PubMed]
- Vårhammar, A.; Wallin, G.; McLean, C.M.; Dusenge, M.E.; Medlyn, B.E.; Hasper, T.B.; Nsabimana, D.; Uddling, J. Photosynthetic temperature responses of tree species in Rwanda: Evidence of pronounced negative effects of high temperature in montane rainforest climax species. New Phytol. 2015, 206, 1000–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.M.; Pearson, D.; Booth, B.B.; Friedlingstein, P.; Huntingford, C.; Jones, C.D.; Luke, C.M. Sensitivity of tropical carbon to climate change constrained by carbon dioxide variability. Nature 2013, 494, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.A. Sources or sinks? The responses of tropical forests to current and future climate and atmospheric composition. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Richards, P.W. The Tropical Rain Forest: An Ecological Study; The University Press: Cambridge, UK, 1952. [Google Scholar]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, CA, USA, 1967. [Google Scholar]
- Marín-Spiotta, E.; Sharma, S. Carbon storage in successional and plantation forest soils: A tropical analysis. Glob. Ecol. Biogeogr. 2013, 22, 105–117. [Google Scholar] [CrossRef]
- Slot, M.; Rey-Sanchez, C.; Gerber, S.; Lichstein, J.W.; Winter, K.; Kitajima, K. Thermal acclimation of leaf respiration of tropical trees and lianas: Response to experimental canopy warming, and consequences for tropical forest carbon balance. Glob. Chang. Biol. 2014, 20, 2915–2926. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.E. An in situ leaf and branch warming experiment in the Amazon. Biotropica 2011, 43, 658–665. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Glob. Chang. Biol. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Filewod, B.; Thomas, S.C. Impacts of a spring heat wave on canopy processes in a northern hardwood forest. Glob. Chang. Biol. 2014, 20, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Hikosaka, K.; Ishikawa, K.; Borjigidai, A.; Muller, O.; Onoda, Y. Temperature acclimation of photosynthesis: Mechanisms involved in the changes in temperature dependence of photosynthetic rate. J. Exp. Bot. 2006, 57, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.; Bjorkman, O. Photosynthetic response and adaptation to temperature in higher plants. Ann. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Graham, E.A.; Mulkey, S.S.; Kitajima, K.; Phillips, N.G.; Wright, S.J. Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons. Proc. Natl. Acad. Sci. USA 2003, 100, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F.; Way, D.A.; Kubien, D.S. Rubisco, Rubisco activase, and global climate change. J. Exp. Bot. 2008, 59, 1581–1595. [Google Scholar] [CrossRef] [PubMed]
- Salvucci, M.E.; Crafts-Brandner, S.J. Relationship between the heat tolerance of photosynthesis and the thermal stability of Rubisco activase in plants from contrasting thermal environments. Plant Physiol. 2004, 134, 1460–1470. [Google Scholar] [CrossRef] [PubMed]
- Von Caemmerer, S.; Quick, W.P. Rubisco: Physiology in vivo. In Photosynthesis; Leegood, R.C., Sharkey, T.D., von Caemmerer, S., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 85–113. [Google Scholar]
- Way, D.A.; Oren, R. Differential responses to changes in growth temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef] [PubMed]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.A.; Piper, S.; Keeling, C.; Clark, D.B. Tropical rain forest tree growth and atmospheric carbon dynamics linked to interannual temperature variation during 1984–2000. Proc. Natl. Acad. Sci. USA 2003, 100, 5852–5857. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Beadle, C.; Loughhead, S. Photosynthetic temperature responses of Eucalyptus globulus and Eucalyptus nitens. Tree Physiol. 1996, 16, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Slot, M.; Winter, K. In situ temperature response of photosynthesis of 42 tree and liana species in the canopy of two Panamanian lowland tropical forests with contrasting rainfall regimes. New Phytol. 2017, 214, 1103–1117. [Google Scholar] [CrossRef] [PubMed]
- Slot, M.; Winter, K. Photosynthetic acclimation to warming in tropical forest tree seedlings. J. Exp. Bot. 2017, 68, 2275–2284. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.-H.; Zeng, J.; Zhang, Y.-J.; Slot, M.; Gamo, M.; Hirano, T.; Kosugi, Y.; Da Rocha, H.R.; Saleska, S.R.; Goulden, M.L. Optimum air temperature for tropical forest photosynthesis: Mechanisms involved and implications for climate warming. Environ. Res. Lett. 2017. [Google Scholar] [CrossRef]
- Cunningham, S.; Read, J. Comparison of temperate and tropical rainforest tree species: Photosynthetic responses to growth temperature. Oecologia 2002, 133, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M. Direct and indirect climate change effects on photosynthesis and transpiration. Plant Biol. 2004, 6, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Rey-Sánchez, A.C.; Slot, M.; Posada, J.M.; Kitajima, K. Spatial and seasonal variation in leaf temperature within the canopy of a tropical forest. Clim. Res. 2016, 71, 75–89. [Google Scholar] [CrossRef]
- Sefcik, L.T.; Zak, D.R.; Ellsworth, D.S. Photosynthetic responses to understory shade and elevated carbon dioxide concentration in four northern hardwood tree species. Tree Physiol. 2006, 26, 1589–1599. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. Photosynthesis and resource distribution through plant canopies. Plant Cell Environ. 2007, 30, 1052–1071. [Google Scholar] [CrossRef] [PubMed]
- Van de Weg, M.J.; Meir, P.; Williams, M.; Girardin, C.; Malhi, Y.; Silva-Espejo, J.; Grace, J. Gross primary productivity of a high elevation tropical montane cloud forest. Ecosystems 2014, 17, 751–764. [Google Scholar] [CrossRef]
- Dai, Y.; Dickinson, R.E.; Wang, Y.-P. A two-big-leaf model for canopy temperature, photosynthesis, and stomatal conductance. J. Clim. 2004, 17, 2281–2299. [Google Scholar] [CrossRef]
- Koch, G.W.; Amthor, J.S.; Goulden, M.L. Diurnal patterns of leaf photosynthesis, conductance and water potential at the top of a lowland rain forest canopy in Cameroon: Measurements from the Radeau des Cimes. Tree Physiol. 1994, 14, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Catovsky, S.; Holbrook, N.; Bazzaz, F. Coupling whole-tree transpiration and canopy photosynthesis in coniferous and broad-leaved tree species. Can. J. For. Res. 2002, 32, 295–309. [Google Scholar] [CrossRef]
- Reich, P.B.; Ellsworth, D.S.; Walters, M.B. Leaf structure (specific leaf area) modulates photosynthesis–nitrogen relations: Evidence from within and across species and functional groups. Funct. Ecol. 1998, 12, 948–958. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Field, C.H.; Mooney, H.A. Photosynthesis-Nitrogen Relationship in Wild Plants. In On the Economy of Plant Form and Function, Proceedings of the Sixth Maria Moors Cabot Symposium, Evolutionary Constraints on Primary Productivity, Adaptive Patterns of Energy Capture in Plants, Harvard Forest, Petersham, MA, USA, 1 August 1983; Cambridge University Press: Cambridge, NY, USA, 1986. [Google Scholar]
- Rozendaal, D.; Hurtado, V.; Poorter, L. Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Funct. Ecol. 2006, 20, 207–216. [Google Scholar] [CrossRef]
- Crous, K.Y.; Ellsworth, D.S. Canopy position affects photosynthetic adjustments to long-term elevated CO2 concentration (FACE) in aging needles in a mature Pinus taeda forest. Tree Physiol. 2004, 24, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.P.; Beckerman, A.P.; Gu, L.; Kattge, J.; Cernusak, L.A.; Domingues, T.F.; Scales, J.C.; Wohlfahrt, G.; Wullschleger, S.D.; Woodward, F.I. The relationship of leaf photosynthetic traits—Vcmax and Jmax—To leaf nitrogen, leaf phosphorus, and specific leaf area: A meta-analysis and modeling study. Ecol. Evol. 2014, 4, 3218–3235. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Cavaleri, M.A.; Oberbauer, S.F.; Clark, D.B.; Clark, D.A.; Ryan, M.G. Height is more important than light in determining leaf morphology in a tropical forest. Ecology 2010, 91, 1730–1739. [Google Scholar] [CrossRef] [PubMed]
- Coble, A.; Cavaleri, M. Light drives vertical gradients of leaf morphology in a sugar maple (Acer saccharum) forest. Tree Physiol. 2014, 34, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Coble, A.P.; Autio, A.; Cavaleri, M.A.; Binkley, D.; Ryan, M.G. Converging patterns of vertical variability in leaf morphology and nitrogen across seven Eucalyptus plantations in Brazil and Hawaii, USA. Trees 2014, 28, 1–15. [Google Scholar] [CrossRef]
- Xiang, S.; Reich, P.B.; Sun, S.C.; Atkin, O.K. Contrasting leaf trait scaling relationships in tropical and temperate wet forest species. Funct. Ecol. 2013, 27, 522–534. [Google Scholar] [CrossRef]
- Vitousek, P.M. Beyond global warming: Ecology and global change. Ecology 1994, 75, 1861–1876. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R. Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere. Proc. Natl. Acad. Sci. USA 2006, 103, 10316–10321. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M. Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 1984, 65, 285–298. [Google Scholar] [CrossRef]
- Bloomfield, K.J.; Farquhar, G.D.; Lloyd, J. Photosynthesis-nitrogen relationships in tropical forest tree species as affected by soil phosphorus availability: A controlled environment study. Funct. Plant Biol. 2014, 41, 820–832. [Google Scholar] [CrossRef]
- Raaimakers, D.; Boot, R.; Dijkstra, P.; Pot, S. Photosynthetic rates in relation to leaf phosphorus content in pioneer versus climax tropical rainforest trees. Oecologia 1995, 102, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Gleason, S.M.; Read, J.; Ares, A.; Metcalfe, D.J. Phosphorus economics of tropical rainforest species and stands across soil contrasts in Queensland, Australia: Understanding the effects of soil specialization and trait plasticity. Funct. Ecol. 2009, 23, 1157–1166. [Google Scholar] [CrossRef]
- Jarvi, M.P.; Burton, A.J. Acclimation and soil moisture constrain sugar maple root respiration in experimentally warmed soil. Tree Physiol. 2013, 33, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Campione, M.A.; Nagel, L.M.; Webster, C.R. Herbaceous-layer community dynamics along a harvest-intensity gradient after 50 years of consistent management. Open J. For. 2012, 2, 97. [Google Scholar] [CrossRef]
- Monroe, W.H. The Karst Landforms of Puerto Rico; US Department of the Interior, Geological Survey: Washington, DC, USA, 1976.
- Lugo, A.E.; da Silva, J.F.; Uribe, A.M.S. Balance de carbono en un bosque novedoso de Castilla elastica: Resultados preliminares. Asoc. Maest. Cienc. Puerto Rico 2008, 22, 13–27. [Google Scholar]
- Ewel, J.J.; Whitmore, J.L. Ecological life zones of Puerto Rico and US Virgin Islands. In Ecological Life Zones of Puerto Rico and US Virgin Islands; US Department of Agriculture: Rio Piedras, PR, USA, 1973. [Google Scholar]
- Kitajima, K.; Mulkey, S.S.; Wright, S.J. Variation in crown light utilization characteristics among tropical canopy trees. Ann. Bot. 2005, 95, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Brako, L.; Zarucchi, J.L. Catalogue of the Flowering Plants and Gymnosperms of Peru; Missouri Botanical Garden: St. Louis, MO, USA, 1993. [Google Scholar]
- Comita, L.S.; Thompson, J.; Uriarte, M.; Jonckheere, I.; Canham, C.D.; Zimmerman, J.K. Interactive effects of land use history and natural disturbance on seedling dynamics in a subtropical forest. Ecol. Appl. 2010, 20, 1270–1284. [Google Scholar] [CrossRef] [PubMed]
- Myster, R.W.; Walker, L.R. Plant successional pathways on Puerto Rican landslides. J. Trop. Ecol. 1997, 13, 165–173. [Google Scholar] [CrossRef]
- Scatena, F.; Silver, W.; Siccama, T.; Johnson, A.; Sanchez, M. Biomass and nutrient content of the Bisley experimental watersheds, Luquillo Experimental Forest, Puerto Rico, before and after Hurricane Hugo, 1989. Biotropica 1993, 25, 15–27. [Google Scholar] [CrossRef]
- Harris, N.L.; Lugo, A.E.; Brown, S.; Heartsill Scalley, T. Luquillo Experimental Forest: Research History and Opportunities; USDA Forest Service: Washington, DC, USA, 2012.
- Lugo, A.E.; Batlle, C.T.R. Leaf production, growth rate, and age of the palm Prestoea montana in the Luquillo Experimental Forest, Puerto Rico. J. Trop. Ecol. 1987, 151–161. [Google Scholar] [CrossRef]
- Weaver, P.; Medina, E.; Pool, D.; Dugger, K.; Gonzales-Liboy, J.; Cuevas, E. Ecological observations in the dwarf cloud forest of the Luquillo Mountains in Puerto Rico. Biotropica 1986, 18, 79–85. [Google Scholar] [CrossRef]
- Frangi, J.L.; Lugo, A.E. A flood plain palm forest in the Luquillo Mountains of Puerto Rico five years after Hurricane Hugo. Biotropica 1998, 30, 339–348. [Google Scholar] [CrossRef]
- Reagan, D.P.; Waide, R.B. The Food Web of a Tropical Rain Forest; University of Chicago Press: Chicago, IL, USA, 1996. [Google Scholar]
- Heartsill-Scalley, T.; Scatena, F.; Estrada, C.; McDowell, W.; Lugo, A. Disturbance and long-term patterns of rainfall and throughfall nutrient fluxes in a subtropical wet forest in Puerto Rico. J. Hydrol. 2007, 333, 472–485. [Google Scholar] [CrossRef]
- Lugo, A.E.; Wadsworth, F.H. Dacryodes excelsa vahl tabonuco. Silv. N. Am. 1990, 2, 284. [Google Scholar]
- Zimmerman, J.K.; Everham III, E.M.; Waide, R.B.; Lodge, D.J.; Taylor, C.M.; Brokaw, N.V. Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: Implications for tropical tree life histories. J. Ecol. 1994, 911–922. [Google Scholar] [CrossRef]
- Birdsey, R.A.; Weaver, P.L. Forest Area Trends in Puerto Rico; US Department of Agriculture: New Orleans, LA, USA, 1987.
- Ducrey, M. Influence of Shade on Photosynthetic Gas Exchange of Seven Tropical Rain-Forest Species from Guadeloupe (French West Indies). In Annales des Sciences Forestières; EDP Sciences: Les Ulis, France, 1994; pp. 77–94. [Google Scholar]
- Gendron, F.; Messier, C.; Comeau, P.G. Comparison of various methods for estimating the mean growing season percent photosynthetic photon flux density in forests. Agric. For. Meteorol. 1998, 92, 55–70. [Google Scholar] [CrossRef]
- Silva, L.C.R.; Anand, M. Probing for the influence of atmospheric CO2 and climate change on forest ecosystems across biomes. Glob. Ecol. Biogeogr. 2013, 22, 83–92. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Rada, F.; Azócar, A.; García-Núñez, C.; Cabrera, H.M. Gas exchange and low temperature resistance in two tropical high mountain tree species from the Venezuelan Andes. Acta Oecol. 2000, 21, 203–211. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Watanabe, Y.; Yoneda, R.; Ninomiya, I.; Koike, T. Changes in photosynthesis and leaf characteristics with tree height in five dipterocarp species in a tropical rain forest. Tree Physiol. 2006, 26, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Kenzo, T.; Yoneda, R.; Sano, M.; Araki, M.; Shimizu, A.; Tanaka-Oda, A.; Chann, S. Variations in leaf photosynthetic and morphological traits with tree height in various tree species in a Cambodian tropical dry evergreen forest. Jpn. Agric. Res. Q. 2012, 46, 167–180. [Google Scholar] [CrossRef]
- Delagrange, S.; Messier, C.; Lechowicz, M.J.; Dizengremel, P. Physiological, morphological and allocational plasticity in understory deciduous trees: Importance of plant size and light availability. Tree Physiol. 2004, 24, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Körner, C.H.; Diemer, M. In situ photosynthetic responses to light, temperature and carbon dioxide in herbaceous plants from low and high altitude. Funct. Ecol. 1987, 1, 179–194. [Google Scholar] [CrossRef]
- Ferrar, P.J.; Slatyer, R.O.; Vranjic, J.A. Photosynthetic temperature acclimation in Eucalyptus species from diverse habitats, and a comparison with Nerium oleander. Funct. Plant Biol. 1989, 16, 199–217. [Google Scholar]
- Chazdon, R.L.; Pearcy, R.W.; Lee, D.W.; Fetcher, N. Photosynthetic responses of tropical forest plants to contrasting light environments. In Tropical Forest Plant Ecophysiology; Springer: Boston, MA, USA, 1996; pp. 5–55. [Google Scholar]
- Leakey, A.; Press, M.; Scholes, J. High-temperature inhibition of photosynthesis is greater under sunflecks than uniform irradiance in a tropical rain forest tree seedling. Plant Cell Environ. 2003, 26, 1681–1690. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. From tropics to tundra: Global convergence in plant functioning. Proc. Natl. Acad. Sci. USA 1997, 94, 13730–13734. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Cernusak, L.A.; Winter, K.; Dalling, J.W.; Holtum, J.A.M.; Jaramillo, C.; Korner, C.; Leakey, A.D.B.; Norby, R.J.; Poulter, B.; Turner, B.L.; et al. Tropical forest responses to increasing atmospheric CO2: Current knowledge and opportunities for future research. Funct. Plant Biol. 2013, 40, 531–551. [Google Scholar] [CrossRef]
- Malhi, Y. The productivity, metabolism and carbon cycle of tropical forest vegetation. J. Ecol. 2012, 100, 65–75. [Google Scholar] [CrossRef]
- Peters, E.B.; Wythers, K.R.; Zhang, S.; Bradford, J.B.; Reich, P.B. Potential climate change impacts on temperate forest ecosystem processes. Can. J. For. Res. 2013, 43, 939–950. [Google Scholar] [CrossRef]
- Leakey, A.; Scholes, J.; Press, M. Physiological and ecological significance of sunflecks for dipterocarp seedlings. J. Exp. Bot. 2005, 56, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Iverson, L.R.; McKenzie, D. Tree-species range shifts in a changing climate: Detecting, modeling, assisting. Landsc. Ecol. 2013, 28, 879–889. [Google Scholar] [CrossRef]
- Janzen, D.H. Why mountain passes are higher in the tropics. Am. Nat. 1967, 233–249. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B. Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia 1993, 96, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, D.S.; Reich, P.B. Leaf mass per area, nitrogen content and photosynthetic carbon gain in Acer saccharum seedlings in contrasting forest light environments. Funct. Ecol. 1992, 423–435. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S.; Uhl, C. Photosynthesis-nitrogen relations in Amazonian tree species. Oecologia 1994, 97, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M. The flux of carbon between the chloroplast and cytoplasm. In Plant Physiology, Biochemistry and Molecular Biology; Longmans: London, UK, 1997; pp. 382–400. [Google Scholar]
- Rychter, A.M.; Rao, I. Role of phosphorus in photosynthetic carbon metabolism. Handb. Photosynth. 2005, 2, 123–148. [Google Scholar]
- Bassow, S.L.; Bazzaz, F.A. Intra-and inter-specific variation in canopy photosynthesis in a mixed deciduous forest. Oecologia 1997, 109, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, A.; Kitayama, K. Relationship between photosynthetic phosphorus-use efficiency and foliar phosphorus fractions in tropical tree species. Ecol. Evol. 2013, 3, 4872–4880. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Example photosynthetic temperature response curve from Dacryodes excelsa. Photosynthesis increases with temperature to a thermal optimum, then rapidly begins to decline. The temperature at maximum photosynthesis is Topt, and the photosynthetic rate at this temperature is Aopt.
Figure 1.
Example photosynthetic temperature response curve from Dacryodes excelsa. Photosynthesis increases with temperature to a thermal optimum, then rapidly begins to decline. The temperature at maximum photosynthesis is Topt, and the photosynthetic rate at this temperature is Aopt.

Figure 2.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with canopy height (m), where the data were collected for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: *** p < 0.001 and ns (non-significant). Dotted lines represent non-significant regressions.
Figure 2.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with canopy height (m), where the data were collected for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: *** p < 0.001 and ns (non-significant). Dotted lines represent non-significant regressions.

Figure 3.
Log-linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with natural log of percent diffuse non-interceptance [ln(DIFN] for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: * p < 0.05, ** p < 0.01, and ns (non-significant). Dotted lines represent non-significant regressions.
Figure 3.
Log-linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with natural log of percent diffuse non-interceptance [ln(DIFN] for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: * p < 0.05, ** p < 0.01, and ns (non-significant). Dotted lines represent non-significant regressions.

Figure 4.
Comparisons with height (m) of: photosynthetic temperature optima (Topt), mean maximum daily leaf temperature (TLEAFmax), and mean maximum daily air temperature (TAIRmax) in: (a) tropical moist forest; (b) tropical wet forest; (c) temperate deciduous forest. Species-level data within the tropical moist forest were pooled. Dotted lines represent non-significant regressions.
Figure 4.
Comparisons with height (m) of: photosynthetic temperature optima (Topt), mean maximum daily leaf temperature (TLEAFmax), and mean maximum daily air temperature (TAIRmax) in: (a) tropical moist forest; (b) tropical wet forest; (c) temperate deciduous forest. Species-level data within the tropical moist forest were pooled. Dotted lines represent non-significant regressions.

Figure 5.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with leaf mass per area (LMA, g m−2) for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: *** p < 0.001 and ns (non-significant). Dotted lines represent non-significant regressions.
Figure 5.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with leaf mass per area (LMA, g m−2) for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: *** p < 0.001 and ns (non-significant). Dotted lines represent non-significant regressions.

Figure 6.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with nitrogen on a leaf area basis (Narea) for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: ** p < 0.01, *** p < 0.001, and and ns (non-significant). Dotted lines represent non-significant regressions.
Figure 6.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with nitrogen on a leaf area basis (Narea) for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Levels of significance: ** p < 0.01, *** p < 0.001, and and ns (non-significant). Dotted lines represent non-significant regressions.

Figure 7.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with phosphorus on a leaf area basis (Parea) for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Dotted lines represent non-significant regressions. Levels of significance: ns (non-significant).
Figure 7.
Linear regressions of optimum temperature (Topt; °C) and photosynthetic rates at Topt (Aopt; µ mol CO2 m−1 s−2) with phosphorus on a leaf area basis (Parea) for tropical moist (a,d); tropical wet (b,e); and temperate deciduous forests (c,f). Species-level data within the tropical moist forest were pooled. Dotted lines represent non-significant regressions. Levels of significance: ns (non-significant).

Figure 8.
(a) Leaf temperature (°C) vs. stomatal conductance (conductance, mol H2O/m2s) by species; (b) Vapor pressure deficit (VPD, kPa) by species vs. stomatal conductance (conductance, mol H2O/m2s). All forest types have been pooled together.
Figure 8.
(a) Leaf temperature (°C) vs. stomatal conductance (conductance, mol H2O/m2s) by species; (b) Vapor pressure deficit (VPD, kPa) by species vs. stomatal conductance (conductance, mol H2O/m2s). All forest types have been pooled together.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Site information for each tower. Species measured, elevation (meters above sea level), mean annual precipitation (MAP, mm), mean annual growing season temperature (MAGT, °C), tower height.
Table 1.
Site information for each tower. Species measured, elevation (meters above sea level), mean annual precipitation (MAP, mm), mean annual growing season temperature (MAGT, °C), tower height.
| Site | Species Measured | Species Code | Elevation (m) | MAP (mm) | MAGT (°C) | Tower Height (m) |
|---|---|---|---|---|---|---|
| Temperate Deciduous | Acer saccharum | Acer | 189 | 879 | 17.4 | 19 |
| Tropical Moist | Ocotea leucoxylon | Oco | 122 | 1295 | 25.5 | 25 |
| Guarea guidonia | Gua | |||||
| Castilla elastica | Cas | |||||
| Tropical Wet | Dacryodes excelsa | Dac | 361 | 3936 | 24.8 | 24.7 |
Table 2.
Mean values by species (see Table 1 for species codes) and forest type of gas exchange. Standard errors are displayed in parentheses. The number of samples (n) corresponds with the number of temperature response curves. Parameters include: temperature optimum for photosynthesis (Topt, °C), maximum rate of photosynthesis in response to temperature (Aopt, µmol CO2 m−1 s−2), leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Letter codes show significant differences among means using Tukey’s mean separation procedures (α = 0.05) (Table 2).
Table 2.
Mean values by species (see Table 1 for species codes) and forest type of gas exchange. Standard errors are displayed in parentheses. The number of samples (n) corresponds with the number of temperature response curves. Parameters include: temperature optimum for photosynthesis (Topt, °C), maximum rate of photosynthesis in response to temperature (Aopt, µmol CO2 m−1 s−2), leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Letter codes show significant differences among means using Tukey’s mean separation procedures (α = 0.05) (Table 2).
| Species/Forest | n | Topt | Aopt |
|---|---|---|---|
| Acer | 23 | 27.4(0.4)b | 7.31(0.55)b |
| Oco | 6 | 30.3(0.5)a | 11.51(0.96)a |
| Gua | 4 | 29.5(0.8)ab | 7.03(3.03)ab |
| Cas | 14 | 30.0(0.4)a | 9.88(0.65)a |
| Dac | 12 | 29.9(0.9)a | 4.90(0.59)b |
| Temperate Deciduous | 23 | 27.4(0.4)B | 7.31(0.55)A |
| Tropical Moist | 24 | 30.0(0.3)A | 9.76(0.69)B |
| Tropical Wet | 12 | 29.9(0.9)A | 4.90(0.59)A |
Table 3.
Mean values by species (see Table 1 for species codes) and forest type of leaf trait variables. Standard errors are displayed in parentheses. The number of samples (n) corresponds with the number of temperature response curves. Parameters include: temperature optimum for photosynthesis (Topt, °C), maximum rate of photosynthesis in response to temperature (Aopt, µmol CO2 m−1 s−2), leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Letter codes show significant differences among means using Tukey’s mean separation procedures (α = 0.05).
Table 3.
Mean values by species (see Table 1 for species codes) and forest type of leaf trait variables. Standard errors are displayed in parentheses. The number of samples (n) corresponds with the number of temperature response curves. Parameters include: temperature optimum for photosynthesis (Topt, °C), maximum rate of photosynthesis in response to temperature (Aopt, µmol CO2 m−1 s−2), leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Letter codes show significant differences among means using Tukey’s mean separation procedures (α = 0.05).
| Species/Forest | n | LMA | Parea | Pmass | Narea | Nmass |
|---|---|---|---|---|---|---|
| Acer | 23 | 44.5(3.3)c | 0.085(0.005)b | 1.72(0.04)a | 0.94(0.07)c | 21.2(0.7)b |
| Oco | 6 | 114.7(11.8)ab | 0.104(0.014)ab | 0.88(0.06)c | 2.93(0.34)a | 25.5(1.0)ab |
| Gua | 4 | 77.7(8.6)abc | 0.140(0.005)a | 1.85(0.15)a | 2.16(0.27)ab | 29.2(4.8)a |
| Cas | 14 | 87.7(11.7)b | 0.118(0.010)a | 1.43(0.07)b | 2.21(0.30)ab | 25.1(0.5)a |
| Dac | 12 | 122.0(10.3)a | 0.096(0.007)ab | 0.80(0.03)c | 1.73(0.13)b | 14.6(0.7)c |
| Temperate Deciduous | 23 | 44.5(3.3)C | 0.085(0.005)B | 1.72(0.04)A | 0.94(0.07)C | 21.2(0.7)B |
| Tropical Moist | 24 | 92.8(7.9)B | 0.118(0.007)A | 1.36(0.08)B | 2.38(0.21)A | 25.9(0.9)A |
| Tropical Wet | 12 | 122.0(10.3)A | 0.096(0.007)AB | 0.80(0.03)C | 1.73(0.13)B | 14.6(0.7)C |
LMA increased linearly with height in all three forest types (Table 4).
Table 4.
R2, p-values, n, slopes, and intercepts for linear regressions between canopy height and vs. leaf traits for each forest type. Species data within the tropical moist forest were pooled. Traits include: leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Levels of significance: * p < 0.05, ** p < 0.01, *** p < 0.001, and ns (non-significant).
Table 4.
R2, p-values, n, slopes, and intercepts for linear regressions between canopy height and vs. leaf traits for each forest type. Species data within the tropical moist forest were pooled. Traits include: leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Levels of significance: * p < 0.05, ** p < 0.01, *** p < 0.001, and ns (non-significant).
| Height | LMA | Parea | Pmass | Narea | Nmass | |
|---|---|---|---|---|---|---|
| Temperate Deciduous | R2 | 0.72 | 0.062 | 0.14 | 0.76 | 0.03 |
| p-value | <0.001 *** | <0.001 *** | 0.13 ns | <0.001 *** | 0.42 ns | |
| n | 23 | 17 | 17 | 23 | 23 | |
| slope | 2.00 | 0.004 | −0.02 | 0.04 | 0.09 | |
| intercept | 19.9 | 0.01 | 2.03 | 0.41 | 20.1 | |
| Tropical Moist | R2 | 0.44 | 0.03 | 0.5 | 0.36 | 0.06 |
| p-value | <0.001 ** | 0.42 ns | <0.001 *** | <0.001 *** | 0.26 ns | |
| n | 24 | 24 | 24 | 24 | 24 | |
| slope | 5.2 | 0.001 | −0.05 | 0.12 | −0.20 | |
| intercept | 4.60 | 0.10 | 2.34 | 0.29 | 29.3 | |
| Tropical Wet | R2 | 0.49 | 0.30 | 0.17 | 0.37 | 0.14 |
| p-value | 0.02 * | 0.08 ns | 0.21 ns | 0.05 * | 0.28 ns | |
| n | 11 | 11 | 11 | 11 | 11 | |
| slope | 4.10 | 0.002 | −0.01 | 0.002 | −0.23 | |
| intercept | 50.1 | 0.05 | 0.94 | 1.80 | 18.7 |
Percent DIFN and LMA showed significant log-linear relationships with height in the tropical wet and temperate deciduous forests, but not in the tropical moist forest canopy (Table 5).
Table 5.
R2, p-values, n, slopes, and intercepts for linear regressions between light (ln(DIFN)) vs. leaf traits for each forest type. Species data within the tropical moist forest were pooled. Traits include: leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Levels of significance: * p < 0.05, ** p < 0.01, *** p < 0.001, and ns (non-significant).
Table 5.
R2, p-values, n, slopes, and intercepts for linear regressions between light (ln(DIFN)) vs. leaf traits for each forest type. Species data within the tropical moist forest were pooled. Traits include: leaf mass per area (LMA, g m−2), phosphorus on a leaf area basis (Parea, g m−2), phosphorus on a leaf mass basis (Pmass, mg g−1), nitrogen on a leaf area basis (Narea, g m−2), and nitrogen on a leaf mass basis (Nmass, mg g−1). Levels of significance: * p < 0.05, ** p < 0.01, *** p < 0.001, and ns (non-significant).
| ln (DIFN) | LMA | Parea | Pmass | Narea | Nmass | |
|---|---|---|---|---|---|---|
| Temperate Deciduous | R2 | 0.41 | 0.25 | 0.00 | 0.49 | 0.04 |
| p-value | 0.001 *** | 0.039 ns | 0.94 ns | <0.001 *** | 0.37 ns | |
| n | 23 | 17 | 17 | 23 | 23 | |
| slope | 5.39 | 0.01 | 0.002 | 0.12 | 0.36 | |
| intercept | 60.5 | 0.005 | 1.72 | 1.30 | 22.3 | |
| Tropical Moist | R2 | 0.04 | 0.02 | 0.02 | 0.01 | 0.02 |
| p-value | 0.37 ns | 0.50 ns | 0.47 ns | 0.65 ns | 0.45 ns | |
| n | 24 | 24 | 24 | 24 | 24 | |
| slope | 6.39 | 0.004 | −0.05 | 0.08 | −0.58 | |
| intercept | 107 | 0.12 | 1.20 | 2.56 | 24.6 | |
| Tropical Wet | R2 | 0.56 | 0.31 | 0.27 | 0.01 | 0.37 |
| p-value | 0.01 ** | 0.07 ns | 0.10 ns | 0.81 ns | 0.05 * | |
| n | 11 | 11 | 11 | 11 | 11 | |
| slope | 15.5 | 0.01 | −0.03 | −0.02 | −0.80 | |
| intercept | 154 | 0.11 | 0.72 | 1.86 | 13.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mau, A.C.; Reed, S.C.; Wood, T.E.; Cavaleri, M.A. Temperate and Tropical Forest Canopies are Already Functioning beyond Their Thermal Thresholds for Photosynthesis. Forests 2018, 9, 47. https://doi.org/10.3390/f9010047
AMA Style
Mau AC, Reed SC, Wood TE, Cavaleri MA. Temperate and Tropical Forest Canopies are Already Functioning beyond Their Thermal Thresholds for Photosynthesis. Forests. 2018; 9(1):47. https://doi.org/10.3390/f9010047
Chicago/Turabian StyleMau, Alida C., Sasha C. Reed, Tana E. Wood, and Molly A. Cavaleri. 2018. "Temperate and Tropical Forest Canopies are Already Functioning beyond Their Thermal Thresholds for Photosynthesis" Forests 9, no. 1: 47. https://doi.org/10.3390/f9010047
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.