Spruce Budworm (Choristoneura fumiferana Clem.) Defoliation Promotes Vertical Fuel Continuity in Ontario’s Boreal Mixedwood Forest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Site Selection
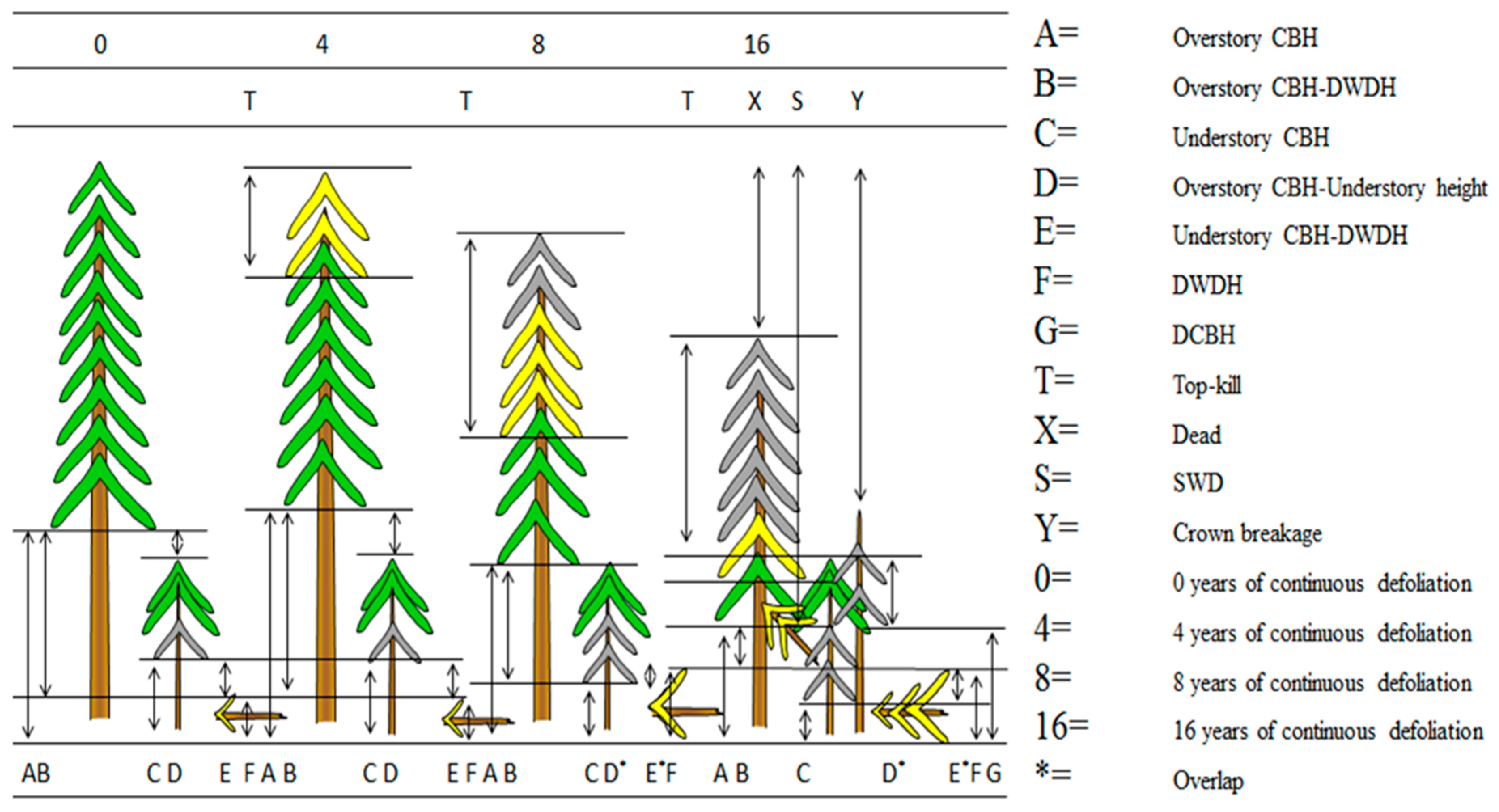
2.3. Data Collection
2.4. Statistical Methods
3. Results
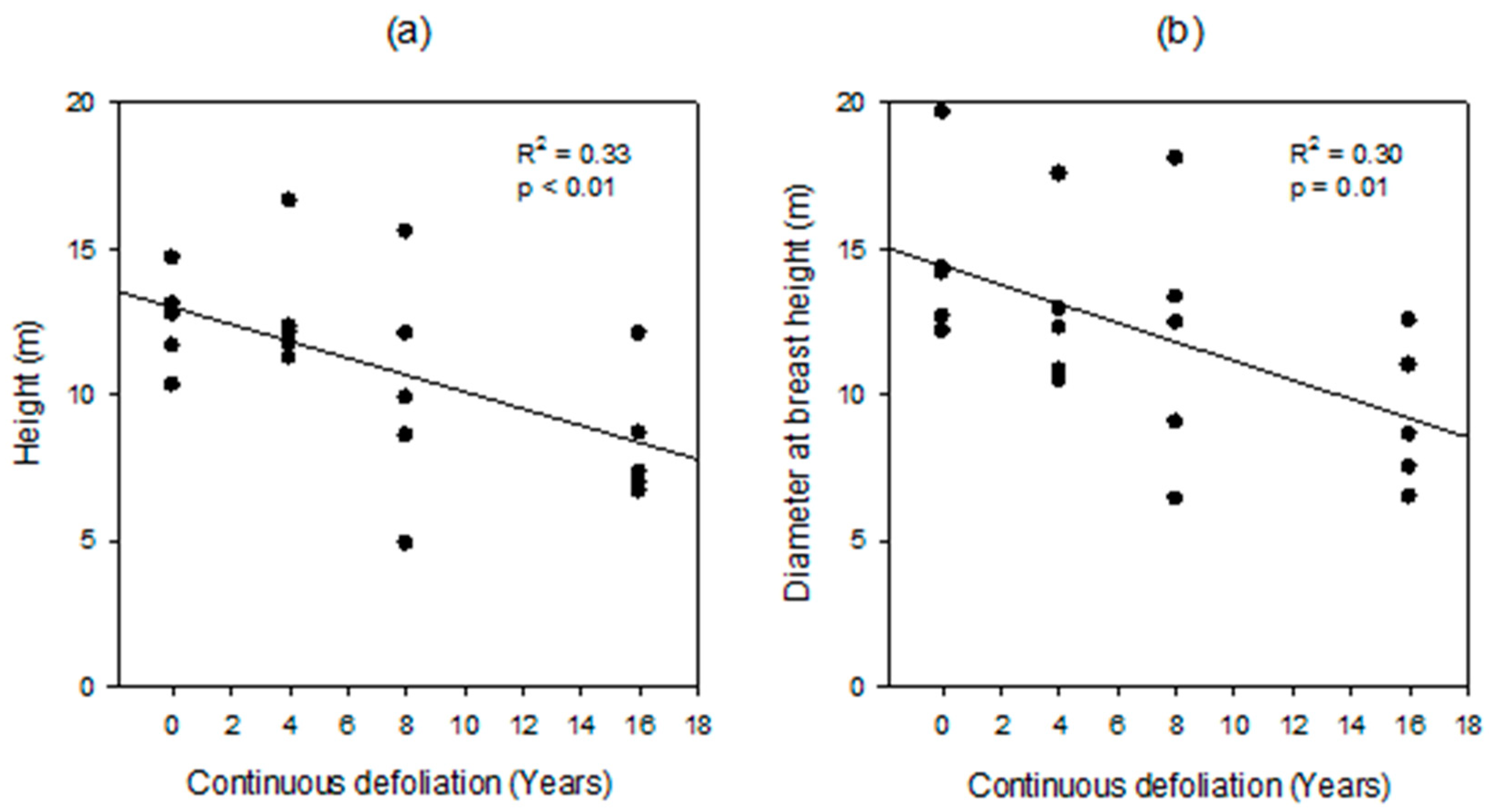
3.1. Forest Characteristics and Defoliation Duration
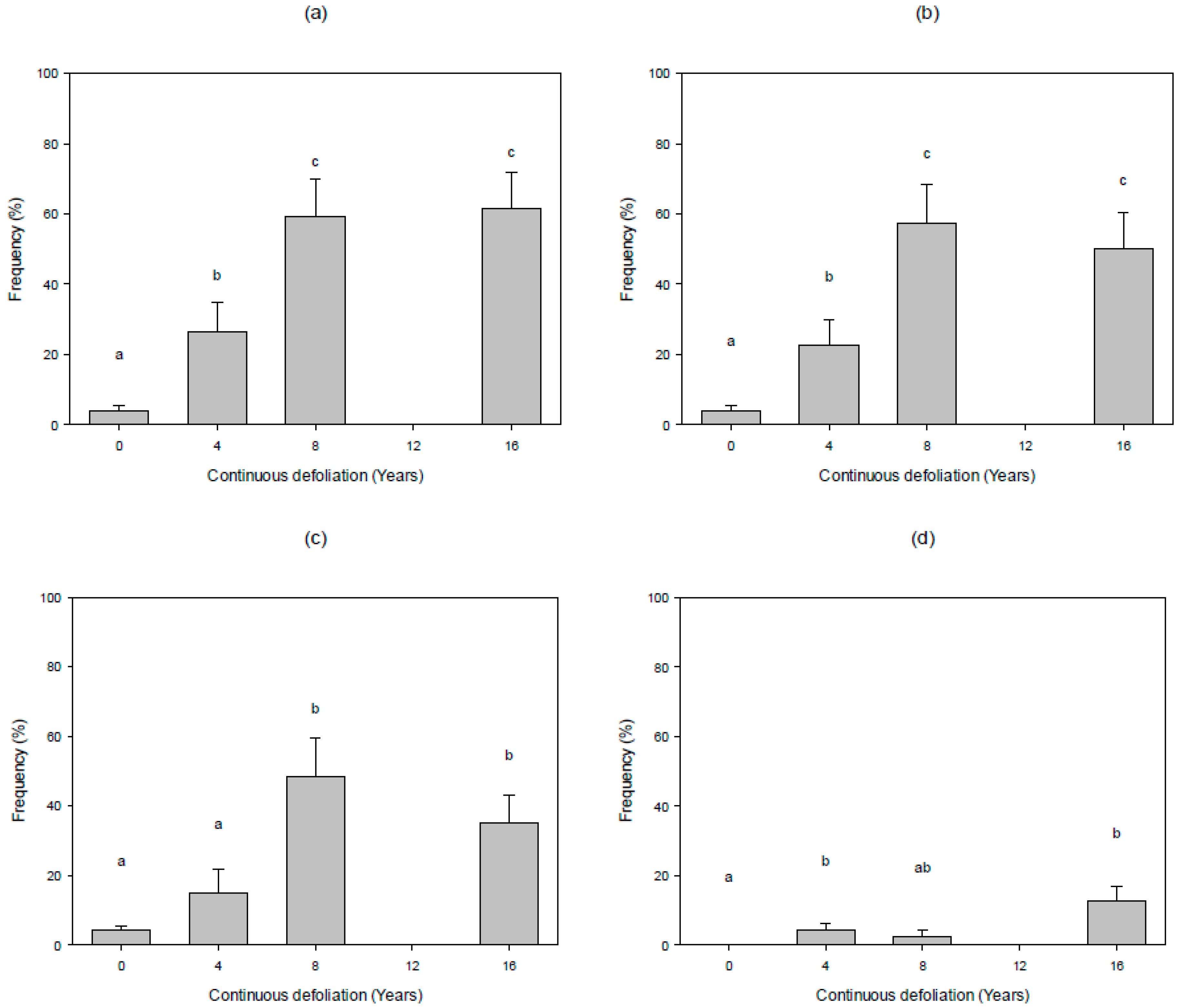
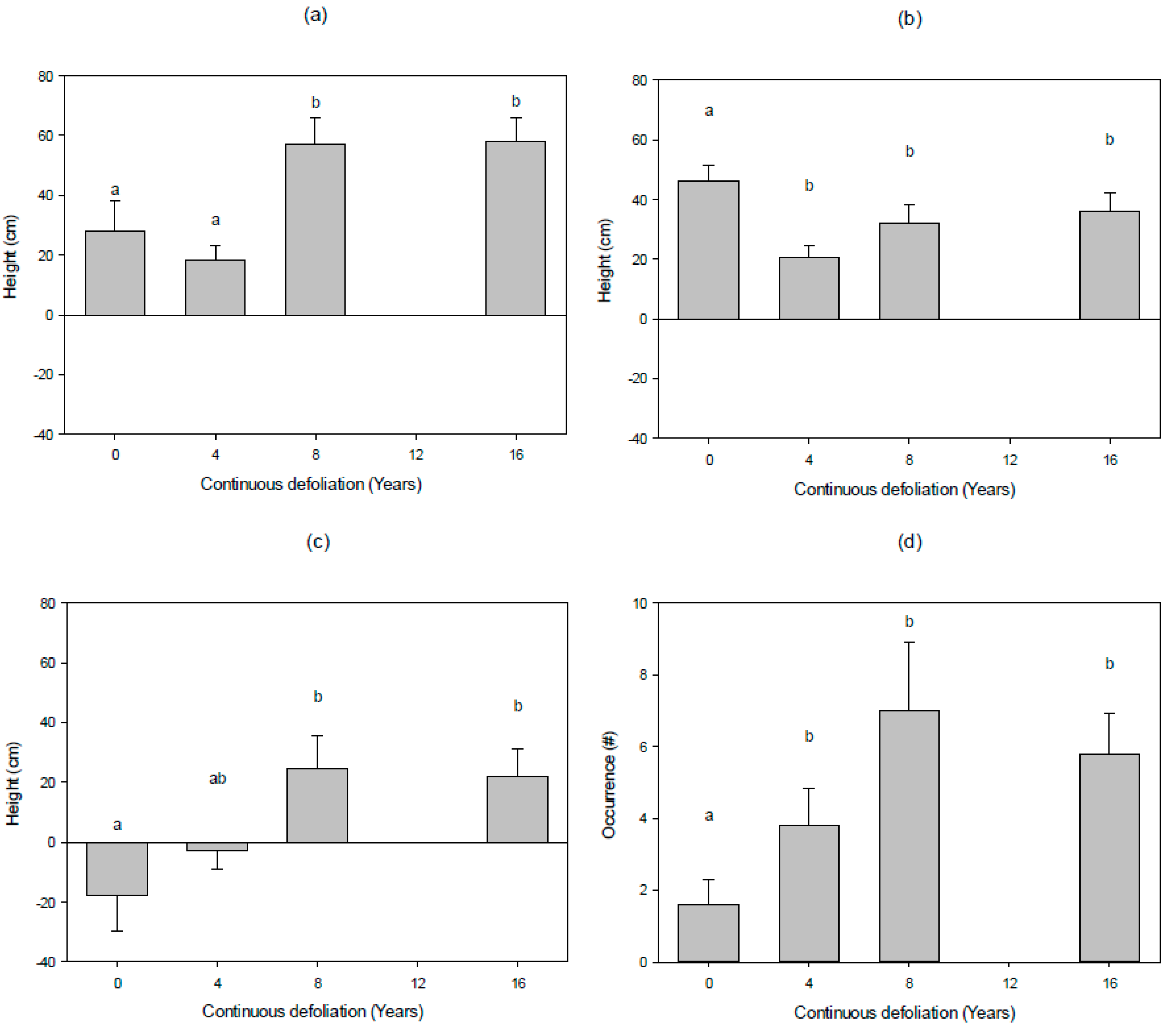
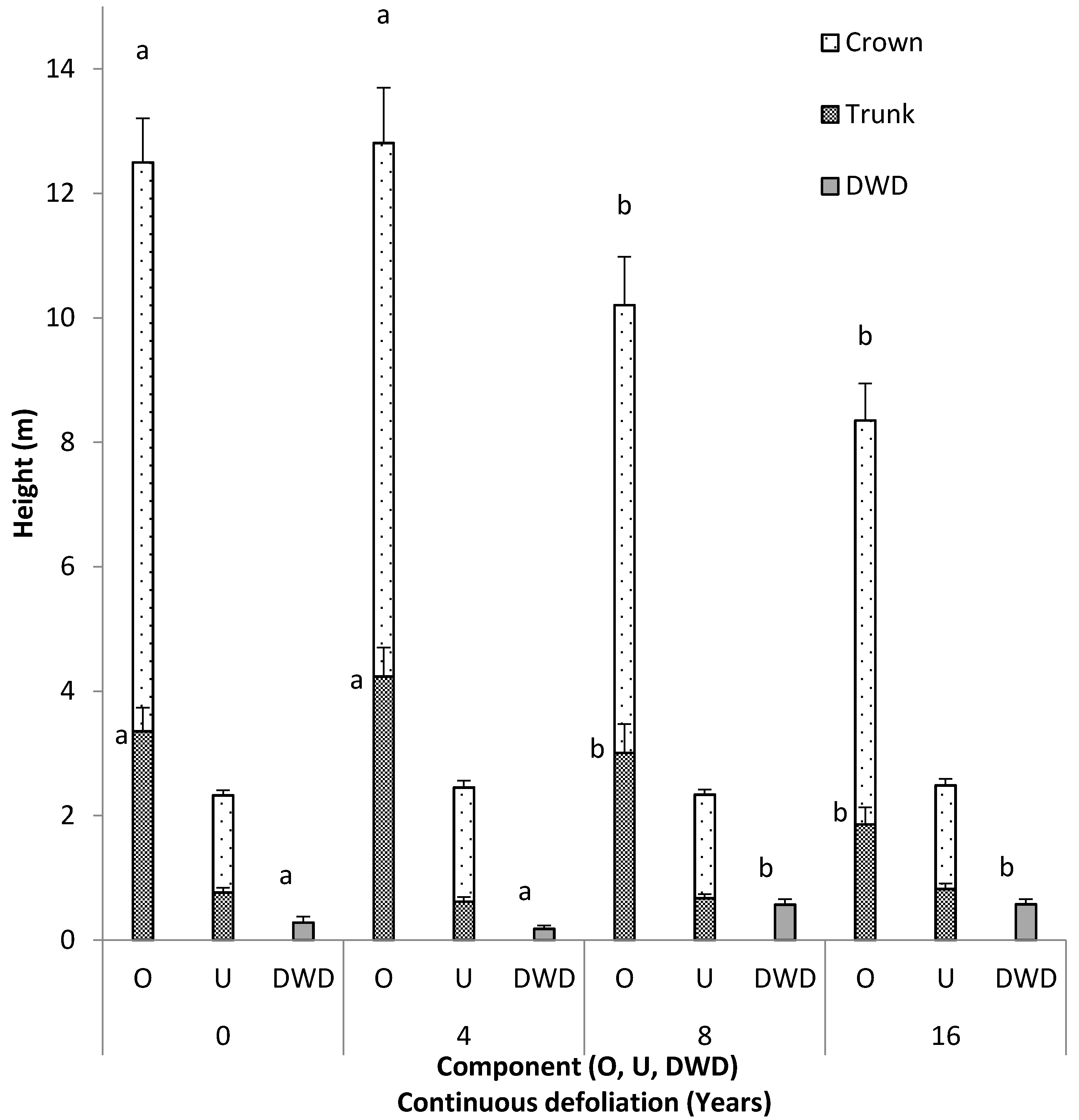
3.2. Vertical Fuel Continuity
4. Discussion
4.1. Forest Characteristics and Defoliation Duration
4.2. Vertical Fuel Continuity
4.3. Limitations
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Candau, J.; Fleming, R.; Hopkin, A. Spatiotemporal patterns of large-scale defoliation caused by the spruce budworm in Ontario since 1941. Can. J. For. Res. 1998, 28, 1733–1741. [Google Scholar] [CrossRef]
- Fleming, R.A. Climate change and insect disturbance regimes in Canada’s boreal forests. World Resour. Rev. 2000, 12, 520–554. [Google Scholar]
- Volney, W.J.; Fleming, R. Climate change and impacts of boreal forest insects. Agric. Ecosyst. Environ. 2000, 82, 283–294. [Google Scholar] [CrossRef]
- Brassard, B.; Chen, H. Stand structural dynamics of North American boreal forests. Crit. Rev. Plant. Sci. 2006, 25, 115–137. [Google Scholar] [CrossRef]
- MacLean, D.A. Effects of spruce budworm outbreaks on the productivity and stability of balsam fir forests. For. Chron. 1984, 60, 273–279. [Google Scholar] [CrossRef]
- MacLean, D.A. Vulnerability of fir-spruce stands during uncontrolled spruce budworm outbreaks: A review and discussion. For. Chron. 1980, 56, 213–221. [Google Scholar] [CrossRef]
- Taylor, S.; MacLean, D. Legacy of insect defoliators: Increased wind-related mortality two decades after a spruce budworm outbreak. For. Sci. 2009, 55, 256–267. [Google Scholar]
- Menning, K.; Stephens, S. Fire climbing in the forest: A semiqualitative approach to assessing ladder fuel hazards. West. J. Appl. For. 2007, 22, 88–93. [Google Scholar]
- Taylor, S.; MacLean, D. Dead wood dynamics in declining balsam fir and spruce stands in New Brunswick, Canada. Can. J. For. Res. 2007, 37, 750–762. [Google Scholar] [CrossRef]
- Stocks, B.J. Fire potential in the spruce budworm-damaged forests of Ontario. For. Chron. 1987, 63, 8–14. [Google Scholar] [CrossRef]
- Fleming, R.; Candau, J.; McAlpine, R. Landscape-scale analysis of interaction between insect defoliation and forest fire in central Canada. Clim. Chang. 2002, 55, 251–272. [Google Scholar] [CrossRef]
- James, P.M.; Robert, L.E.; Wotton, B.M.; Martell, D.L.; Fleming, R.A. Lagged cumulative spruce budworm defoliation affects the risk of fire ignition in Ontario, Canada. Ecol. Appl. 2017, 27, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Candau, J.; Fleming, R.; Wang, X. Ecoregional patterns of spruce budworm-wildfire interactions in central Canada’s forests. Forests 2018, 9, 137. [Google Scholar] [CrossRef]
- Stocks, B.J. The extent and impact of forest fires in northern circumpolar countries. In Global Biomass Burning: Atmospheric, Climatic, and Biospheric Implications; Levine, J.S., Ed.; MIT Press: Cambridge, MA, USA, 1991; pp. 197–202. [Google Scholar]
- Van Wagner, C.E. Conditions for the start and spread of crown fire. Can. J. For. Res. 1977, 7, 23–34. [Google Scholar] [CrossRef]
- Scarr, T.; Ryall, K.; Hodge, P. (Eds.) Forest Health Conditions in Ontario, 2011; Queen’s Printer for Ontario: Sault Ste. Marie, ON, Canada, 2012; pp. 1–100. [Google Scholar]
- Lavoie, N.; Alexander, M.; Macdonald, S. Photo guide for qualitatively assessing the characteristics of forest fuels in a jack pine—Black spruce chronosequence in the Northwest Territories. Nat. Res. Can. 2010, 1–51. Available online: http://cfs.nrcan.gc.ca/publications/download-pdf/31785 (accessed on 19 April 2013).
- Cottam, G.; Curtis, J. The use of distance measures in phytosociological sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Maser, C.; Anderson, R.G.; Cromack, K.; Williams, J.T.; Martin, R.E. Dead and down woody material. In Wildlife Habitats in Managed Forests: The Blue Mountains of Oregon and Washington; Thomas, J.W., Ed.; USDA: Portland, OR, USA, 1979; Volume 553, pp. 78–95. [Google Scholar]
- Van Wagner, C.E. The line intersect method in forest fuel sampling. For. Sci. 1968, 14, 20–26. [Google Scholar]
- Péch, G. Fire hazard in budworm-killed balsam fir stands on Cape Breton Highlands. For. Chron. 1993, 69, 178–186. [Google Scholar] [CrossRef]
- Greenbank, D.O.; Schaefer, G.W.; Rainey, R.C. Spruce budworm (Lepidoptera: Tortricidae) moth flight and dispersal: New understanding from canopy observations, radar, and aircraft. Mem. Entomol. Soc. Can. 1980, 110, 1–49. [Google Scholar] [CrossRef]
- Fleming, R.; Piene, H. Spruce budworm defoliation and growth loss in young balsam fir: Cohort models of needlefall schedules for spaced trees. For. Sci. 1992, 38, 287–304. [Google Scholar]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Bouchard, M.; Kneeshaw, D.; Bergeron, Y. Mortality and stand renewal patterns following the last spruce budworm outbreak in mixed forests of western Quebec. For. Ecol. Manag. 2005, 204, 297–313. [Google Scholar] [CrossRef]
- MacLean, D.A. Impact of insect outbreaks on tree mortality, productivity, and stand development. Can. Entomol. 2015, 148, S138–S159. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| July | Forest Resource Inventory Data (1989) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | Structure | Composition | |||||||||
| Continuous Defoliation | Site | Coordinate | Height (m) | Age (Years) | Stocking (0–1) | Host (%) | |||||
| (Years) | (#) | N (°) | W (°) | Value | SEM | Value | SEM | Value | SEM | Value | SEM |
| 0 | 1 | 46 29 52.9999 | 79 09 39.5012 | 21 | 90 | 0.8 | 20 | ||||
| 0 | 2 | 46 32 25.9689 | 79 09 09.1302 | 19 | 110 | 0.7 | 40 | ||||
| 0 | 3 | 46 32 26.2641 | 79 08 48.2574 | 15 | 90 | 0.5 | 80 | ||||
| 0 | 4 | 46 33 53.6288 | 79 09 29.5137 | 14 | 90 | 0.7 | 50 | ||||
| 0 | 5 | 46 33 50.3132 | 79 09 34.8001 | 11 | 95 | 0.7 | 30 | ||||
| Mean | 16 | ±2 | 95 | ±4 | 0.7 | ±0.0 | 44 | ±10 | |||
| 4 | 6 | 46 47 49.6233 | 79 48 08.4162 | 13 | 110 | 0.9 | 50 | ||||
| 4 | 7 | 46 37 35.6161 | 79 48 04.3366 | 13 | 150 | 0.5 | 20 | ||||
| 4 | 8 | 46 45 36.4558 | 79 48 25.0054 | 11 | 103 | 0.4 | 20 | ||||
| 4 | 9 | 46 44 40.7008 | 79 49 16.8733 | 16 | 95 | 0.7 | 30 | ||||
| 4 | 10 | 46 44 35.7205 | 79 49 09.5751 | 16 | 95 | 0.7 | 30 | ||||
| Mean | 14 | ±1 | 111 | ±10 | 0.6 | ±0.1 | 30 | ±5 | |||
| 8 | 11 | 46 37 58.8879 | 80 00 18.5172 | 22 | 85 | 0.8 | 30 | ||||
| 8 | 12 | 46 37 18.0186 | 80 00 38.8205 | 21 | 80 | 0.0 | 50 | ||||
| 8 | 13 | 46 37 02.8668 | 80 00 26.1413 | 20 | 80 | 0.8 | 30 | ||||
| 8 | 14 | 46 35 50.8079 | 80 00 26.7027 | 13 | 65 | 0.7 | 60 | ||||
| 8 | 15 | 46 35 34.8847 | 80 00 32.9673 | 15 | 62 | 0.9 | 30 | ||||
| Mean | 18 | ±2 | 74 | ±5 | 0.6 | ±0.2 | 40 | ±6 | |||
| 16 | 16 | 46 30 32.7797 | 80 16 03.1156 | 13 | 75 | 0.6 | 50 | ||||
| 16 | 17 | 46 29 50.5380 | 80 15 14.1664 | 7 | 50 | 0.5 | 60 | ||||
| 16 | 18 | 46 30 34.4934 | 80 16 09.8633 | 13 | 70 | 0.5 | 70 | ||||
| 16 | 19 | 46 30 50.4948 | 80 16 03.5893 | 13 | 75 | 0.6 | 50 | ||||
| 16 | 20 | 46 31 56.0937 | 80 76 04.3965 | 14 | 70 | 0.5 | 20 | ||||
| Mean | 12 | ±1 | 68 | ±5 | 0.5 | ±0.0 | 50 | ±8 | |||
| Continuous Defoliation | n | Density (Stems/ha) | Composition (%) | ||||
|---|---|---|---|---|---|---|---|
| (Years) | (# of Sites) | Overall | SEM | Host | SEM | Balsam Fir | SEM |
| (of Total) | (of Total) | (of Host) | (of Host) | ||||
| Data from July 2011 | |||||||
| 0 | 5 | 3142 | ±702 | 72 | ±31 | 93 | ±3 |
| 4 | 5 | 2664 | ±412 | 51 | ±29 | 57 | ±12 |
| 8 | 5 | 2847 | ±337 | 94 | ±24 | 48 | ±16 |
| 16 | 5 | 4630 | ±280 | 90 | ±40 | 67 | ±18 |
| Pooled data from May and July 2011 | |||||||
| 0 | 10 | 3203 | ±720 | 72 | ±37 | - | - |
| 4 | 10 | 3242 | ±744 | 59 | ±33 | - | - |
| 8 | 10 | 2895 | ±392 | 94 | ±53 | - | - |
| 16 | 10 | 3793 | ±1056 | 87 | ±48 | - | - |
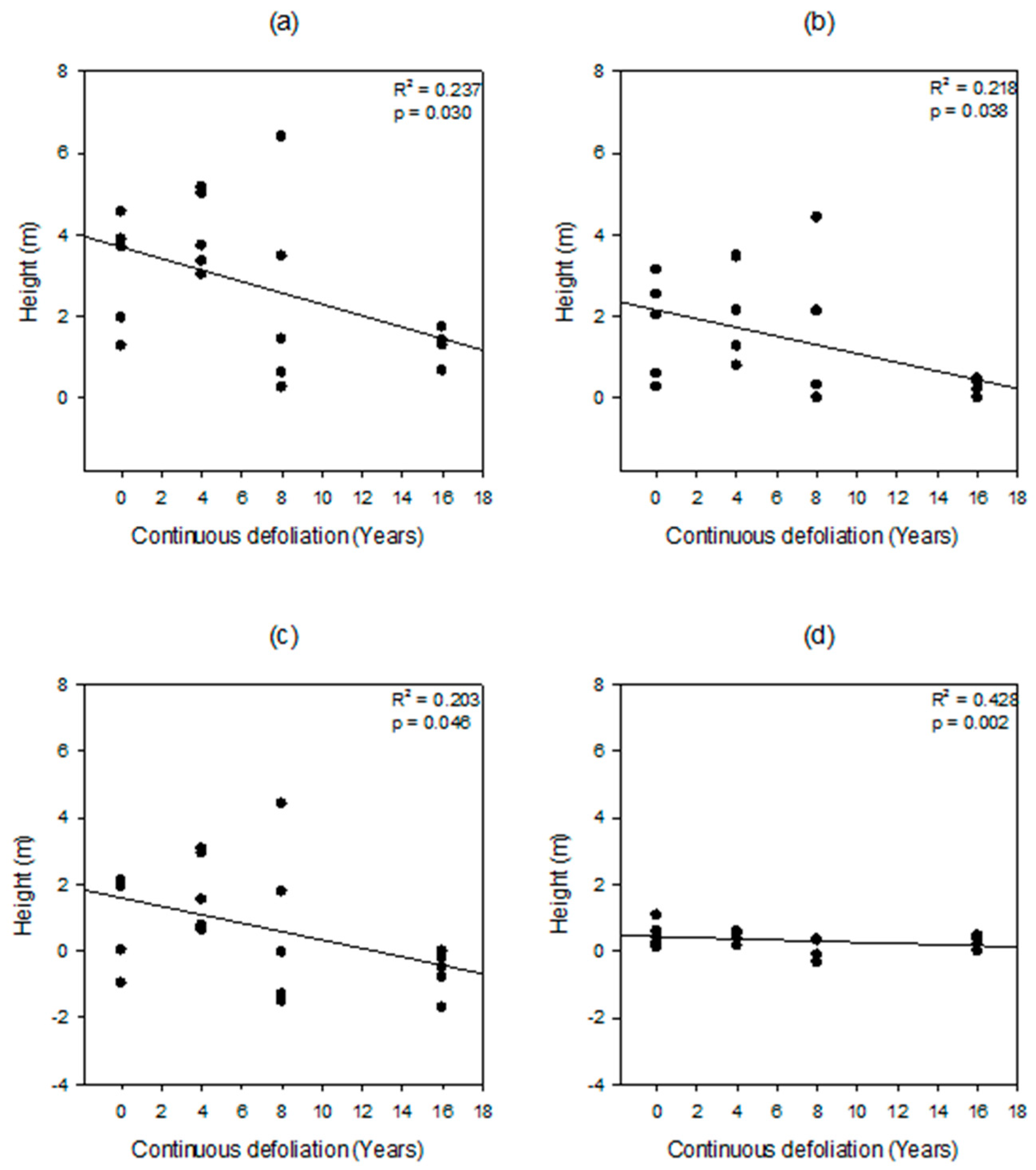
| Dependent Variable | Parameter | Estimate | R2 | SEM | t stat | p | |
|---|---|---|---|---|---|---|---|
| Overstory CBH − DWDH (m) | Intercept | 3.698 | 0.237 | 0.544 | 6.803 | <0.001 | |
| Overstory CBH − DWDH (m) | Slope | −0.140 | 0.059 | −2.364 | 0.030 | ||
| Overstory CBH − Understory H + Understory CBH - DWDH (m) | Intercept | 2.153 | 0.218 | 0.438 | 4.916 | <0.001 | |
| Overstory CBH − Understory H + Understory CBH - DWDH (m) | Slope | −0.107 | 0.048 | −2.240 | 0.038 | ||
| Overstory CBH − Understory H (m) | Intercept | 1.594 | 0.203 | 0.541 | 2.948 | 0.009 | |
| Overstory CBH − Understory H (m) | Slope | −0.126 | 0.059 | −2.140 | 0.046 | ||
| Understory DCBH − DWDH (m) | Intercept | −0.037 | 0.428 | 0.075 | −0.499 | 0.624 | |
| Understory DCBH − DWDH (m) | Slope | −0.030 | 0.008 | −3.669 | 0.002 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watt, G.A.; Fleming, R.A.; Smith, S.M.; Fortin, M.-J. Spruce Budworm (Choristoneura fumiferana Clem.) Defoliation Promotes Vertical Fuel Continuity in Ontario’s Boreal Mixedwood Forest. Forests 2018, 9, 256. https://doi.org/10.3390/f9050256
Watt GA, Fleming RA, Smith SM, Fortin M-J. Spruce Budworm (Choristoneura fumiferana Clem.) Defoliation Promotes Vertical Fuel Continuity in Ontario’s Boreal Mixedwood Forest. Forests. 2018; 9(5):256. https://doi.org/10.3390/f9050256
Chicago/Turabian StyleWatt, Graham A., Richard A. Fleming, Sandy M. Smith, and Marie-Josée Fortin. 2018. "Spruce Budworm (Choristoneura fumiferana Clem.) Defoliation Promotes Vertical Fuel Continuity in Ontario’s Boreal Mixedwood Forest" Forests 9, no. 5: 256. https://doi.org/10.3390/f9050256