Effects of Increased Soil Scarification Intensity on Natural Regeneration of Scots Pine Pinus sylvestris L. and Birch Betula spp. L.


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Scarification Treatment
2.3. Field Procedures
2.4. Statistical Analyses
2.4.1. Data Exploration
2.4.2. Model Selection
- 1.
- Seedling Density—Scarification Intensity
- 2.
- Current Annual Growth—Scarification Intensity
3. Results
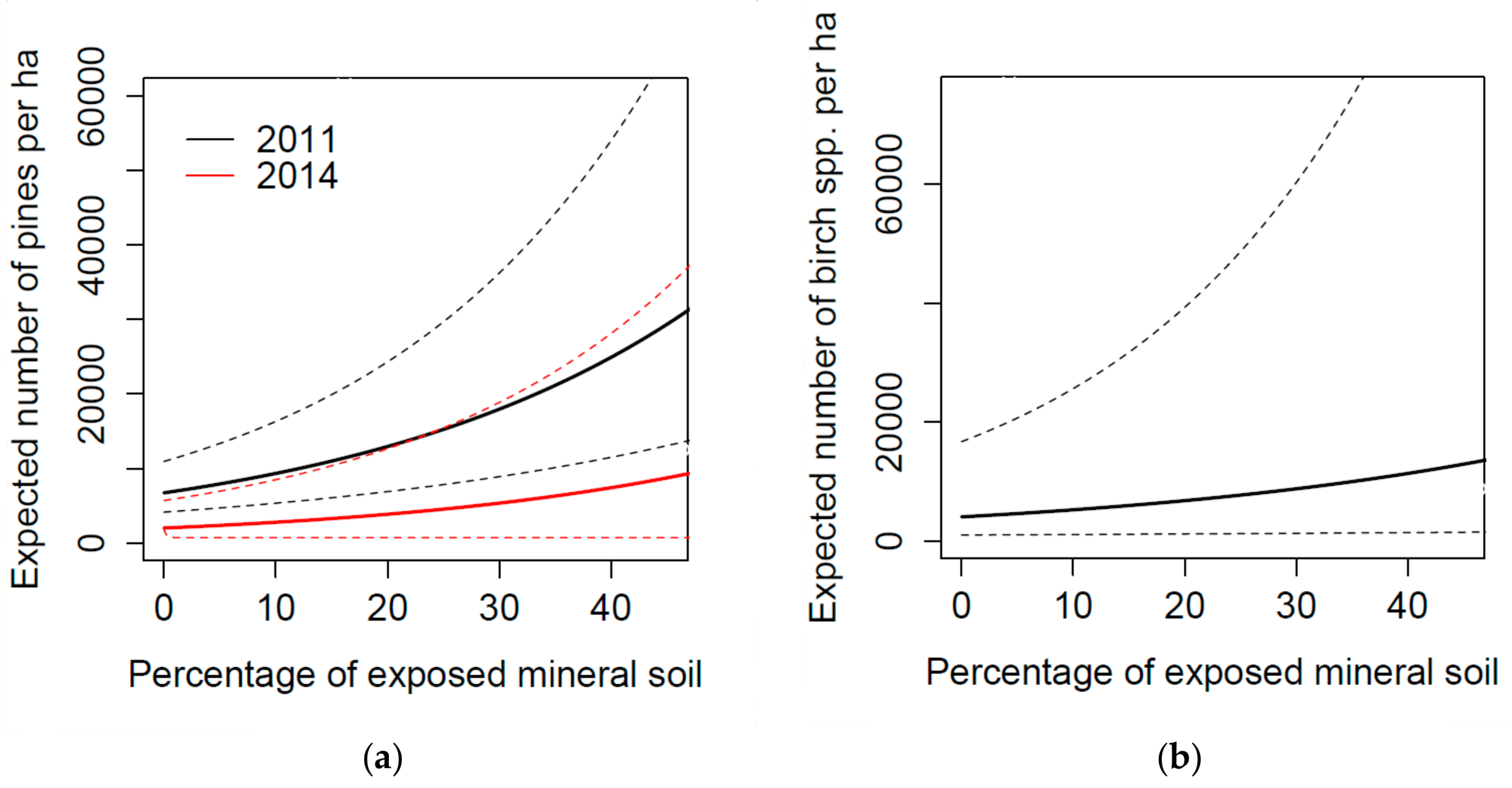
3.1. Tree Seedling Density
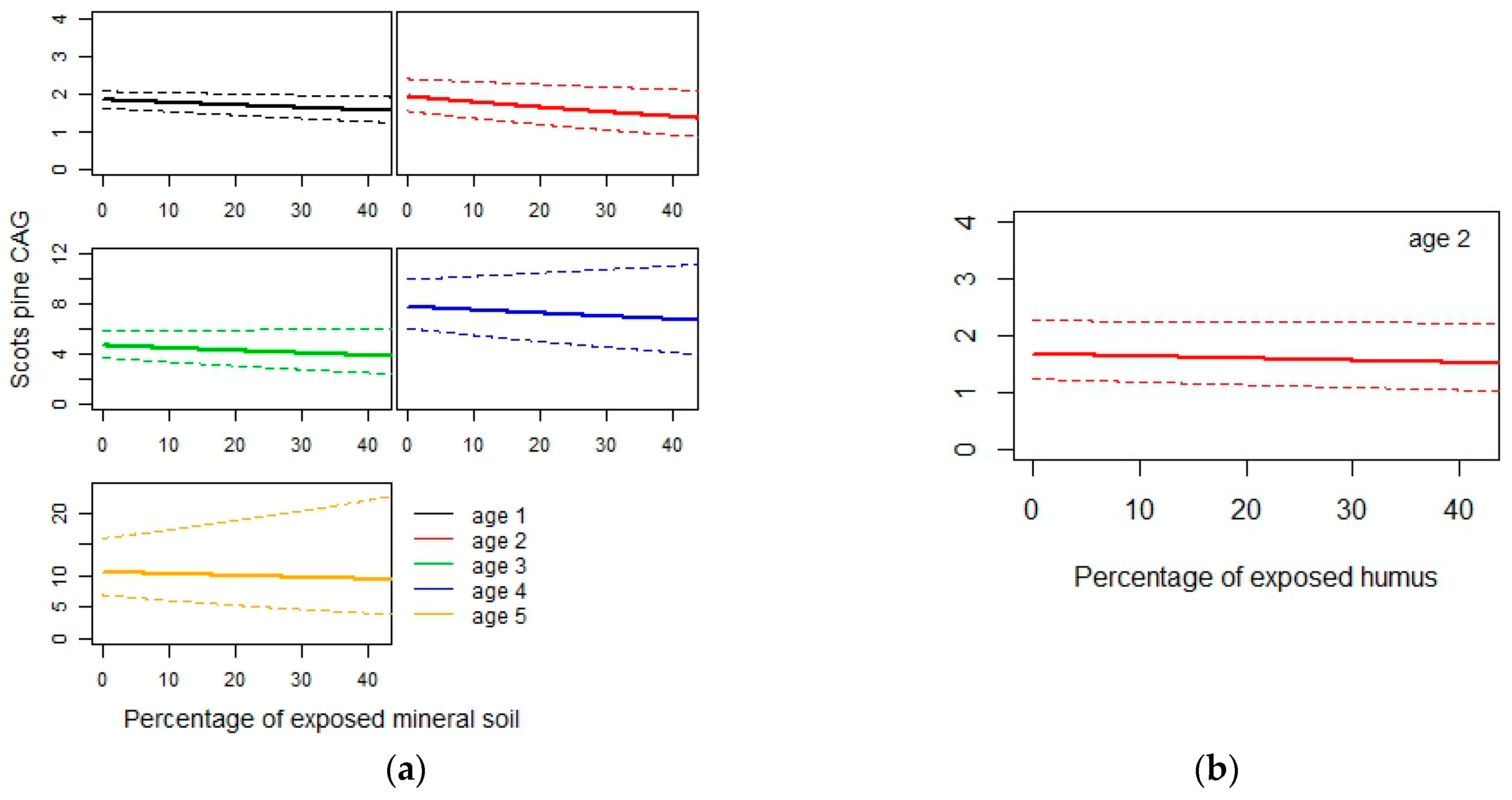
3.2. Current Annual Growth
4. Discussion
4.1. Seedling Density
4.2. Current Annual Growth
4.3. Management Implications
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bergström, R.; Hjeljord, O. Moose and vegetation interactions in northwestern Europe and Poland. Swed. Wildl. Res. Suppl. 1987, 1, 213–228. [Google Scholar]
- Månsson, J.; Bergström, R.; Pehrson, Å.; Skoglund, M.; Skarpe, C. Felled Scots pine (Pinus sylvestris) as supplemental forage for moose (Alces alces): Browse availability and utilization. Scand. J. For. Res. 2010, 25, 21–31. [Google Scholar] [CrossRef]
- Lavsund, S.; Nygrén, T.; Solberg, E.J. Status of moose populations and challenges to moose management in Fennoscandia. Alces 2003, 39, 109–130. [Google Scholar]
- Heikkilä, R.; Härkönen, S. Moose browsing in young Scots pine stands in relation to forest management. For. Ecol. Manag. 1996, 88, 179–186. [Google Scholar] [CrossRef]
- Bergqvist, G.; Bergström, R.; Edenius, L. Effects of moose (Alces alces) rebrowsing on damage development in young stands of Scots pine (Pinus sylvestris). For. Ecol. Manag. 2003, 176, 397–403. [Google Scholar] [CrossRef]
- Storaas, T.; Gundersen, H.; Henriksen, H.; Andreassen, H.P. The economic value of moose in Norway—A review. Alces 2001, 37, 97–107. [Google Scholar]
- Bærekraftig Hjorteviltforvaltning i Hedmark. Available online: https://www.hamar.kommune.no/getfile.php/13408296/Bilder/Hamar/Artikkelbilder/Organisasjonen/R%C3%A5dmann%20og%20staber/Vedlegg%201%20fra%20ERMG%20-%20B%C3%A6rekraftig%20hjorteviltforvaltning%20i%20Hedmark.pdf (accessed on 11 December 2017). (In Norwegian).
- Solbraa, K. Elg i Atndal og Naboområder: Forvaltning av Elg og Skog; 4; Research Report; Hedmark University College: Terningen, Arena, 2008. (In Norwegian) [Google Scholar]
- Gundersen, H.; Andreassen, H.P.; Storaas, T. Supplemental feeding of migratory moose Alces alces: Forest damage at two spatial scales. Wildl. Biol. 2004, 10, 213–223. [Google Scholar]
- Heikkila, R.; Harkonen, S. Thinning residues as a source of browse for moose in managed forests in Finland. Alces 2000, 36, 85–92. [Google Scholar]
- Edenius, L.; Roberge, J.-M.; Månsson, J.; Ericsson, G. Ungulate-adapted forest management: Effects of slash treatment at harvest on forage availability and use. Eur. J. For. Res. 2014, 133, 191–198. [Google Scholar] [CrossRef]
- Edenius, L.; Månsson, J.; Hjortstråle, T.; Roberge, J.-M.; Ericsson, G. Browsing and damage inflicted by moose in young Scots pine stands subjected to high-stump precommercial thinning. Scand. J. For. Res. 2015, 30, 382–387. [Google Scholar] [CrossRef]
- Haug, F.; Haget, D.; Hekne, A.M.; Hegg, B.R. Elgbeite på Gran: Taksering i Stange og Våler kommuner 2013; Fylkesmannen i Hedmark, landbruksavdelingen: Hamar, Norway, 2014; p. 23. (In Norwegian) [Google Scholar]
- Lyly, O.; Saksa, T. The effect of stand density on moose damage in young Pinus sylvestris stands. Scand. J. For. Res. 1992, 7, 393–403. [Google Scholar] [CrossRef]
- Andren, H.; Angelstam, P. Moose browsing on Scots pine in relation to stand size and distance to forest edge. J. Appl. Ecol. 1993, 30, 133–142. [Google Scholar] [CrossRef]
- Wallgren, M.; Bergström, R.; Bergqvist, G.; Olsson, M. Spatial distribution of browsing and tree damage by moose in young pine forests, with implications for the forest industry. For. Ecol. Manag. 2013, 305, 229–238. [Google Scholar] [CrossRef]
- Bergström, R. Rebrowsing on birch (Betula pendula and B. pubescens) stems by moose. Alces 1984, 19, 3–13. [Google Scholar]
- Heikkilä, R. Moose browsing in a Scots pine plantation mixed with deciduous tree species. Acta For. Fenn. 1991, 224, 1–13. [Google Scholar] [CrossRef]
- Mathisen, K.M.; Milner, J.M.; Skarpe, C. Moose–tree interactions: Rebrowsing is common across tree species. BMC Ecol. 2017, 17, 12. [Google Scholar] [CrossRef] [PubMed]
- Edenius, L. Browsing by moose on Scots pine in relation to plant resource availability. Ecology 1993, 74, 2261–2269. [Google Scholar] [CrossRef]
- Löyttyniemi, K. On repeated browsing of Scots pine saplings by moose (Alces alces). Silva Fenn. 1985, 19, 387–391. [Google Scholar] [CrossRef]
- Danell, K.; Bergström, R.; Edenius, L.; Ericsson, G. Ungulates as drivers of tree population dynamics at module and genet levels. For. Ecol. Manag. 2003, 181, 67–76. [Google Scholar] [CrossRef]
- Siipilehto, J.; Heikkilä, R. The effect of moose browsing on the height structure of Scots pine saplings in a mixed stand. For. Ecol. Manag. 2005, 205, 117–126. [Google Scholar] [CrossRef]
- Karlsson, A.; Albrektson, A.; Forsgren, A.; Svensson, L. An analysis of successful natural regeneration of downy and silver birch on abandoned farmland in Sweden. Silva Fenn. 1998, 32, 229–240. [Google Scholar] [CrossRef]
- Nygaard, P. Rett tid for markberedning. Aktuelt fra Skogforskningen 2001, 6, 25. [Google Scholar]
- Øyen, B.-H. Litt om markberedning i furuskog. Aktuelt fra Skogforskningen 2002, 3, 47–49. [Google Scholar]
- Karlsson, C.; Örlander, G. Soil scarification shortly before a rich seed fall improves seedling establishment in seed tree stands of Pinus sylvestris. Scand. J. For. Res. 2000, 15, 256–266. [Google Scholar] [CrossRef]
- Aleksandrowicz-Trzcinska, M.; Drozdowski, S.; Brzeziecki, B.; Rutkowska, P.; Jablonska, B. Effects of different methods of site preparation on natural regeneration of Pinus sylvestris in Eastern Poland. Dendrobiology 2014, 71, 73–81. [Google Scholar] [CrossRef]
- Örlander, G.; Egnell, G.; Albrektson, A. Long-term effects of site preparation on growth in Scots pine. For. Ecol. Manag. 1996, 86, 27–37. [Google Scholar] [CrossRef]
- Karlsson, A. Site preparation of abandoned fields and early establishment of naturally and direct-seeded birch in Sweden. Stud. For. Suec. 1996, 199, 25. [Google Scholar]
- Bedford, L.; Sutton, R. Site preparation for establishing lodgepole pine in the sub-boreal spruce zone of interior British Columbia: The Bednesti trial, 10-year results. For. Ecol. Manag. 2000, 126, 227–238. [Google Scholar] [CrossRef]
- Oleskog, G.; Sahlén, K. Effects of seedbed substrate on moisture conditions and germination of Scots pine (Pinus sylvestris) seeds in a mixed conifer stand. New For. 2000, 20, 119–133. [Google Scholar] [CrossRef]
- Oleskog, G.; Sahlén, K. Effects of seedbed substrate on moisture conditions and germination of Pinus sylvestris seeds in a clearcut. Scand. J. For. Res. 2000, 15, 225–236. [Google Scholar] [CrossRef]
- Piirainen, S.; Finér, L.; Mannerkoski, H.; Starr, M. Carbon, nitrogen and phosphorus leaching after site preparation at a boreal forest clear-cut area. For. Ecol. Manag. 2007, 243, 10–18. [Google Scholar] [CrossRef]
- Chantal, M.D.; Leinonen, K.; Ilvesniemi, H.; Westman, C.J. Combined effects of site preparation, soil properties, and sowing date on the establishment of Pinus sylvestris and Picea abies from seeds. Can. J. For. Res. 2003, 33, 931–945. [Google Scholar] [CrossRef]
- Gaudio, N.; Balandier, P.; Perret, S.; Ginisty, C. Growth of understorey Scots pine (Pinus sylvestris L.) saplings in response to light in mixed temperate forest. Forestry 2011, 84, 187–195. [Google Scholar] [CrossRef]
- Hytönen, J. Allelopathic potential of peatland plant species on germination and early seedling growth of Scots pine, silver birch and downy birch. Silva Fenn. 1992, 26, 63–73. [Google Scholar] [CrossRef]
- Hille, M.; Den Ouden, J. Charcoal and activated carbon as adsorbate of phytotoxic compounds—A comparative study. Oikos 2005, 108, 202–207. [Google Scholar] [CrossRef]
- Bryant, J.P.; Chapin, F.S., III; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef]
- Price, P.W. The plant vigor hypothesis and herbivore attack. Oikos 1991, 62, 244–251. [Google Scholar] [CrossRef]
- De Dios, V.R.; Yoshida, T.; Iga, Y. Effects of topsoil removal by soil-scarification on regeneration dynamics of mixed forests in Hokkaido, Northern Japan. For. Ecol. Manag. 2005, 215, 138–148. [Google Scholar] [CrossRef]
- MacKenzie, M.; Schmidt, M.; Bedford, L. Soil microclimate and nitrogen availability 10 years after mechanical site preparation in northern British Columbia. Can. J. For. Res. 2005, 35, 1854–1866. [Google Scholar] [CrossRef]
- Zaczek, J.J. Composition, diversity, and height of tree regeneration, 3 years after soil scarification in a mixed-oak shelterwood. For. Ecol. Manag. 2002, 163, 205–215. [Google Scholar] [CrossRef]
- Löf, M.; Dey, D.C.; Navarro, R.M.; Jacobs, D.F. Mechanical site preparation for forest restoration. New For. 2012, 43, 825–848. [Google Scholar] [CrossRef]
- Mäkitalo, K. Effect of site preparation and reforestation method on survival and height growth of Scots pine. Scand. J. For. Res. 1999, 14, 512–525. [Google Scholar] [CrossRef]
- Moen, A. Vegetasjonsatlas for Norge: Vegetasjon; Lillethun, A., Ed.; Norwegian Mapping Authority: Hønefoss, Norway, 1998. [Google Scholar]
- Climate Data, Norwegian Meteorological Institute. Available online: https://www.met.no/frie-meteorologiske-data (accessed on 2 March 2016).
- Karlsson, C.; Sikström, U.; Orlander, G.; Hannerz, M.; Hånell, B. Naturlig föryngring Av Tall Och Gran; 4; Research Report; Skogsstyrelsen: Jonkoping, Sweden, 2009. (In Swedish) [Google Scholar]
- Statistics Norway. Klassifisering av Produktivt Skogareal Etter Bonitet (H40) 1997. Available online: http://www.ssb.no/klass/klassifikasjoner/71 (accessed on 26 April 2016). (In Norwegian).
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2009, 1, 3–14. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer Science & Business Media: Berlin, Germany, 2002. [Google Scholar]
- R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 23 January 2016).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-121. Available online: http://CRAN.R-project.org/package=nlme (accessed on 23 January 2016).
- Béland, M.; Agestam, E.; Ekö, P.; Gemmel, P.; Nilsson, U. Scarification and seedfall affects natural regeneration of Scots pine under two shelterwood densities and a clear-cut in southern Sweden. Scand. J. For. Res. 2000, 15, 247–255. [Google Scholar] [CrossRef]
- Hörnfeldt, R.; Hu, B.; Chiriacò, M.V. Optimum timing of soil scarification for the natural regeneration of Pinus sylvestris in Central Sweden. Scand. J. For. Res. 2012, 27, 424–431. [Google Scholar] [CrossRef]
- Sarvas, R. Investigations on the flowering and seed crop of Pinus silvestris. Metsät Tutkimuslait Julk 1962, 53, 198. [Google Scholar]
- Tegelmark, D. Site factors as multivariate predictors of the success of natural regeneration in Scots pine forests. For. Ecol. Manag. 1998, 109, 231–239. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Pukkala, T. Effect of Scots pine seed trees on the density of ground vegetation and tree seedlings. Silva Fenn. 1989, 23, 159–167. [Google Scholar] [CrossRef]
- Yazdani, R.; Lindgren, D. Gene Dispersion after Natural Regeneration under a Widely-Spaced Seed-Tree Stand of Pinus sylvestris. Silvae Genet. 1992, 41, 1–5. [Google Scholar]
- Ackzell, L. Natural regeneration on planted clear-cuts in boreal Sweden. Scand. J. For. Res. 1994, 9, 245–250. [Google Scholar] [CrossRef]
- Kinnaird, J. Effect of site conditions on the regeneration of birch (Betula pendula Roth and B. pubescens Ehrh.). J. Ecol. 1974, 62, 467–472. [Google Scholar] [CrossRef]
- Paavilainen, E.; Päivänen, J. Peatland Forestry: Ecology and Principles; Springer Science & Business Media: Berlin, Germany, 1995; pp. 134–165. [Google Scholar]
- Marquis, D.A. Silvical requirements for natural birch regeneration. In Birch Symposium Proceedings; Northeastern Forest Experiment Station: Pennsylvania, PA, USA, 1969; pp. 36–48. [Google Scholar]
- Morris, L.; Ludovici, K.; Torreano, S.; Carter, E.; Lincoln, M.; Will, R. An approach for using general soil physical condition–root growth relationships to predict seedling growth response to site preparation tillage in loblolly pine plantations. For. Ecol. Manag. 2006, 227, 169–177. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Importance of root growth in overcoming planting stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Davies, W. Control of water balance in transplanting trees. J. Arboric 1975, 1, 1–10. [Google Scholar]
- Kozlowski, T.T.; Clausen, J.J. Shoot growth characteristics of heterophyllous woody plants. Can. J. Bot. 1966, 44, 827–843. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Shoot growth in woody plants. Bot. Rev. 1964, 30, 335–392. [Google Scholar] [CrossRef]
- Odland, A. Bjørkeartenes Spredning, Etablering og Samspill med Naturmiljøet; Nina Report 292; Norwegian Institute for Nature Research: Trondheim, Norway, 1994; pp. 1–50, (In Norwegian, abstract in English). [Google Scholar]
- Johansson, K.; Ring, E.; Hogbom, L. Effects of pre-harvest fertilization and subsequent soil scarification on the growth of planted Pinus Sylvestris seedlings and ground vegetation after clear-felling. Silva Fenn. 2013, 47, 18. [Google Scholar] [CrossRef]
- Hille, M.; Den Ouden, J. Improved recruitment and early growth of Scots pine (Pinus sylvestris L.) seedlings after fire and soil scarification. Eur. J. For. Res. 2004, 123, 213–218. [Google Scholar] [CrossRef]
- Nilsson, U.; Allen, H.L. Short-and long-term effects of site preparation, fertilization and vegetation control on growth and stand development of planted loblolly pine. For. Ecol. Manag. 2003, 175, 367–377. [Google Scholar] [CrossRef]
- Nordborg, F.; Nilsson, U. Growth, damage and net nitrogen uptake in Picea abies (L.) Karst. seedlings, effects of site preparation and fertilisation. Ann. For. Sci 2003, 60, 657–666. [Google Scholar]
- Prévost, M.; Dumais, D. Long-term growth response of black spruce advance regeneration (layers), natural seedlings and planted seedlings to scarification: 25th year update. Scand. J. For. Res. 2018. [Google Scholar] [CrossRef]
- Bergh, J.; Linder, S.; Lundmark, T.; Elfving, B. The effect of water and nutrient availability on the productivity of Norway spruce in northern and southern Sweden. For. Ecol. Manag. 1999, 119, 51–62. [Google Scholar] [CrossRef]
- Fremming, O.R.; Knudsen, M.F.; Storaas, T. Elgbeiteskader på Ungfuru: Avstem Fôrbehov og Fôrproduksjon; 3; Hjortevilttoget Report; Oplandske Bokforlag: Vallset, Norway, 2014; pp. 107–128. (In Norwegian) [Google Scholar]
- Kubin, E. Leaching of nitrate nitrogen into the groundwater after clear felling and site preparation. Boreal Environ. Res. 1998, 3, 3–8. [Google Scholar]
- Knudsen, M.F. Effects of Different Scarification Intensities on Bilberry Abundance. Master’s Thesis, Hedmark University College, Easthern, Norway, 2014. [Google Scholar]
- Parlane, S.; Summers, R.; Cowie, N.; Van Gardingen, P. Management proposals for bilberry in Scots pine woodland. For. Ecol. Manag. 2006, 222, 272–278. [Google Scholar] [CrossRef]
- Storch, I. Habitat selection by capercaillie in summer and autumn: Is bilberry important? Oecologia 1993, 95, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Conen, F.; Zerva, A.; Arrouays, D.; Jolivet, C.; Jarvis, P.G.; Grace, J.; Mencuccini, M. The Carbon Balance of Forest Soils: Detectability of Changes in Soil Carbon Stocks in Temperate and Boreal Forests; Taylor & Francis e-Library: Oxford, UK, 2005; pp. 59–66. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Year | Gravberget | Plassen | Ljørdalen | Total |
|---|---|---|---|---|
| 2011 | 0 | 9 | 0 | 9 |
| 2012 | 29 | 1 | 0 | 30 |
| 2013 | 15 | 1 | 4 | 20 |
| 2014 | 6 | 0 | 2 | 8 |
| Total | 50 | 11 | 6 | 67 |
| Species | Latin Name | Counts |
|---|---|---|
| Scots pine | Pinus sylvestris | 3084 |
| Downy birch | Betula pubescens | 7358 |
| Silver birch | Betula pendula | 666 |
| Young unspecified birch | Betula spp. | 4312 |
| Norway spruce | Picea abies | 170 |
| Common juniper | Juniperus communis | 1 |
| Willows | Salix spp. | 56 |
| Aspen | Populus tremula | 16 |
| Rowan | Sorbus aucuparia | 27 |
| Total | 15,690 |
| Response Variable | Parameter | Estimate | SE |
|---|---|---|---|
| Pine | Intercept | 1.8271 | 0.2002 |
| Mineral soil | 0.0328 | 0.0037 | |
| Other cover | −0.0166 | 0.0056 | |
| Year 2012 | −0.5627 | 0.2133 | |
| Year 2013 | −1.0981 | 0.2227 | |
| Year 2014 | −1.2122 | 0.2730 | |
| Birch | Intercept | 0.9833 | 0.6491 |
| Mineral soil | 0.0258 | 0.0070 | |
| Other cover | 0.0217 | 0.0087 |
| Pine Seedling Density | Df | AIC | ΔAIC |
| mineral + other + year + year × mineral | 11 | 3036.59 | 0.00 |
| mineral + other + year | 8 | 3036.79 | 0.20 |
| mineral + humus + other + year | 9 | 3038.64 | 2.05 |
| mineral + humus + other + year + prod | 11 | 3041.50 | 4.91 |
| mineral + humus + other + year + prod + year × mineral + year × humus + prod × mineral + prod × humus | 21 | 3041.88 | 5.29 |
| Birch Seedling Density | Df | AIC | ΔAIC |
| mineral + other | 6 | 4404.72 | 0.00 |
| mineral + humus + other | 7 | 4405.69 | 0.96 |
| mineral + other + year | 9 | 4406.03 | 1.31 |
| mineral + humus + other + year | 10 | 4406.36 | 1.64 |
| mineral | 5 | 4409.58 | 4.86 |
| Response Variable | Parameter | Estimate | SE |
|---|---|---|---|
| Pine | Intercept | 0.6304 | 0.0506 |
| Mineral soil | −0.0040 | 0.0012 | |
| Humus | −0.0023 | 0.0010 | |
| Age 2 | 0.0456 | 0.0517 | |
| Age 3 | 0.9298 | 0.0547 | |
| Age 4 | 1.4411 | 0.0658 | |
| Age 5 | 1.7571 | 0.1505 | |
| Age 2 × mineral soil | −0.0041 | 0.0014 | |
| Age 3 × mineral soil | −0.0006 | 0.0015 | |
| Age 4 × mineral soil | 0.0006 | 0.0019 | |
| Age 5 × mineral soil | 0.0013 | 0.0043 | |
| Downy birch | Intercept | 0.2472 | 0.0608 |
| Mineral soil | −0.0040 | 0.0014 | |
| Humus | −0.0018 | 0.0016 | |
| Age 2 | 1.2288 | 0.0463 | |
| Age 3 | 1.8562 | 0.0482 | |
| Age 4 | 2.3371 | 0.0578 | |
| Mineral soil × age 2 | −0.0046 | 0.0016 | |
| Mineral soil × age 3 | 0.0013 | 0.0018 | |
| Mineral soil × age 4 | 0.0023 | 0.0021 | |
| Humus × age 2 | −0.0025 | 0.0016 | |
| Humus × age 3 | 0.0067 | 0.0020 | |
| Humus × age 4 | 0.0075 | 0.0028 | |
| Silver birch | Intercept | 0.7419 | 0.1963 |
| Age 2 | 1.1246 | 0.1884 | |
| Age 3 | 1.9009 | 0.1870 | |
| Age 4 | 2.1541 | 0.1945 |
| CAG for Scots Pine | Df | AIC | ΔAIC |
| mineral + humus + age + age × mineral | 15 | 5751.42 | 0.00 |
| mineral + humus + prod + age + age × mineral | 17 | 5753.77 | 2.35 |
| mineral + age + age × mineral | 14 | 5755.25 | 3.83 |
| mineral + humus + prod + age + age × mineral + prod × mineral | 19 | 5757.59 | 6.17 |
| mineral + humus + age | 11 | 5757.88 | 6.46 |
| CAG for Downy Birch | Df | AIC | ΔAIC |
| mineral + humus + age + age × mineral + age × humus | 16 | 11,193.54 | 0.00 |
| mineral + humus + prod + age + age × mineral + age × humus + prod × mineral | 20 | 11,198.26 | 4.72 |
| mineral + humus + other + prod + age + age × mineral + age × humus + prod × mineral + prod × humus | 23 | 11,203.75 | 10.21 |
| mineral + humus + age + age × humus | 13 | 11,215.04 | 21.5 |
| mineral + humus + prod + age + age × humus + prod × mineral | 17 | 11,220.62 | 27.08 |
| CAG for Silver Birch | Df | AIC | ΔAIC |
| age | 8 | 1242.48 | 0.00 |
| humus + age | 9 | 1243.25 | 0.77 |
| mineral + age | 9 | 1243.73 | 1.25 |
| humus + age + age × humus | 12 | 1244.89 | 2.41 |
| mineral + humus + age + age × humus | 13 | 1245.94 | 3.46 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saursaunet, M.; Mathisen, K.M.; Skarpe, C. Effects of Increased Soil Scarification Intensity on Natural Regeneration of Scots Pine Pinus sylvestris L. and Birch Betula spp. L. Forests 2018, 9, 262. https://doi.org/10.3390/f9050262
Saursaunet M, Mathisen KM, Skarpe C. Effects of Increased Soil Scarification Intensity on Natural Regeneration of Scots Pine Pinus sylvestris L. and Birch Betula spp. L. Forests. 2018; 9(5):262. https://doi.org/10.3390/f9050262
Chicago/Turabian StyleSaursaunet, Mona, Karen Marie Mathisen, and Christina Skarpe. 2018. "Effects of Increased Soil Scarification Intensity on Natural Regeneration of Scots Pine Pinus sylvestris L. and Birch Betula spp. L." Forests 9, no. 5: 262. https://doi.org/10.3390/f9050262