Analysis of Nitrogen Utilization Capability during the Proliferation and Maturation Phases of Norway Spruce (Picea abies (L.) H.Karst.) Somatic Embryogenesis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
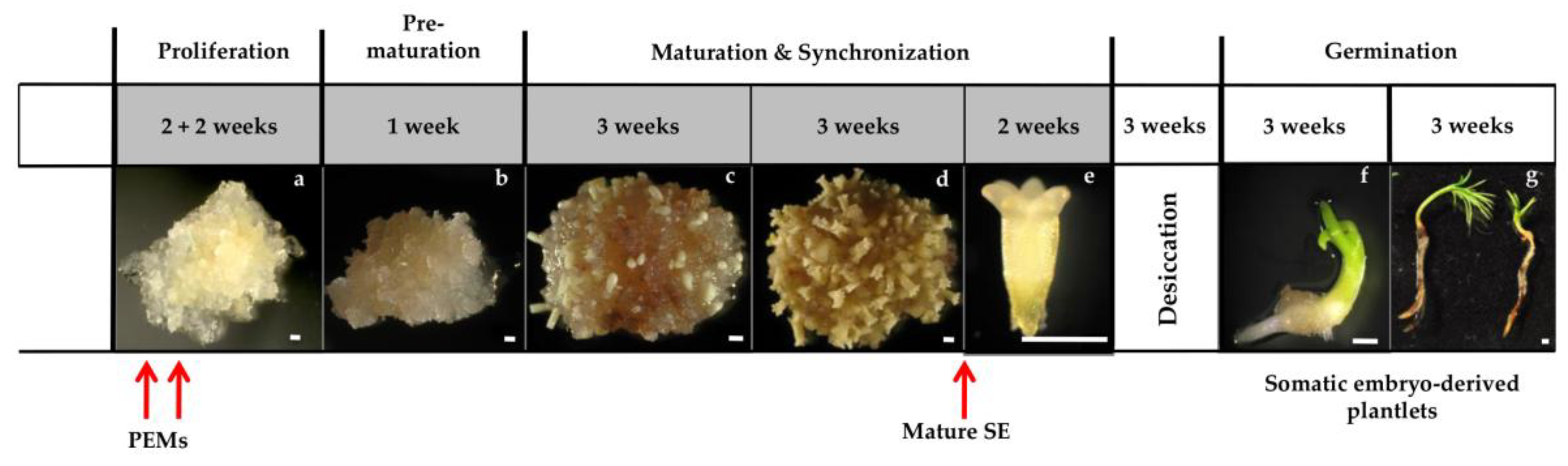
2.2. Experimental Set Up
2.3. Enzyme Assays
2.3.1. GS Assay
2.3.2. NR Assay
2.3.3. Arginase Assay
2.4. Ammonium and Amino Acid Content
2.5. Statistical Analysis
3. Results
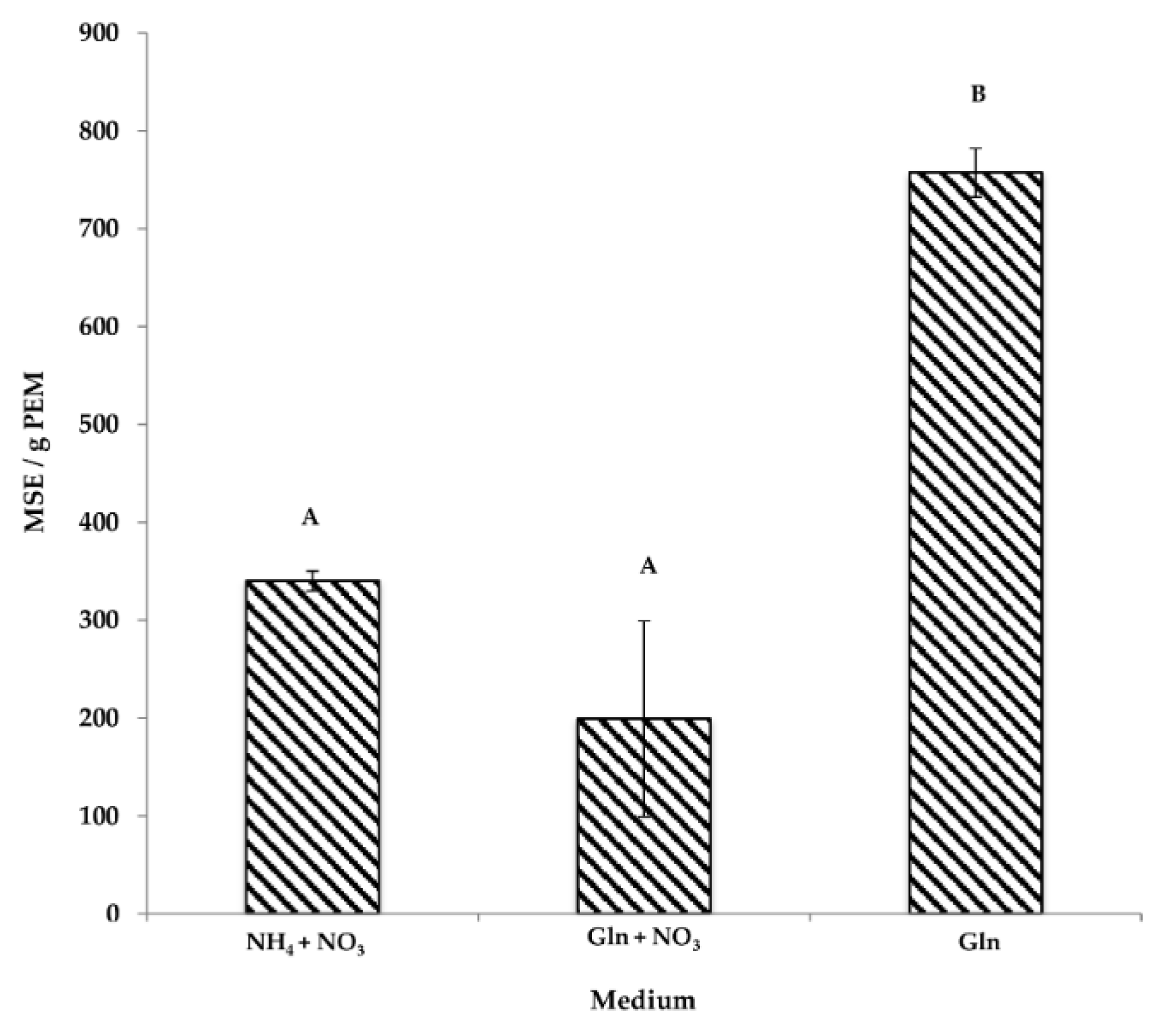
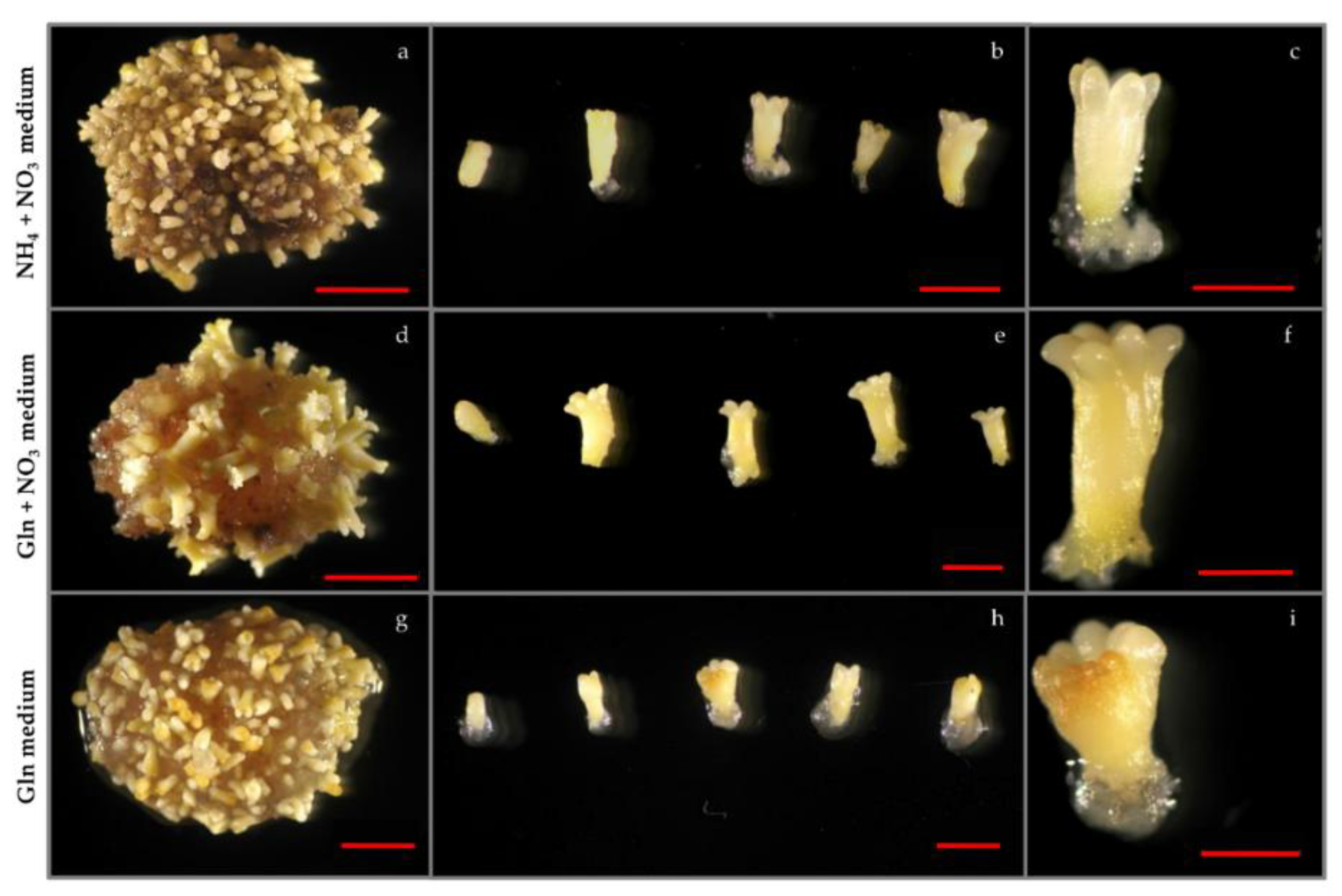
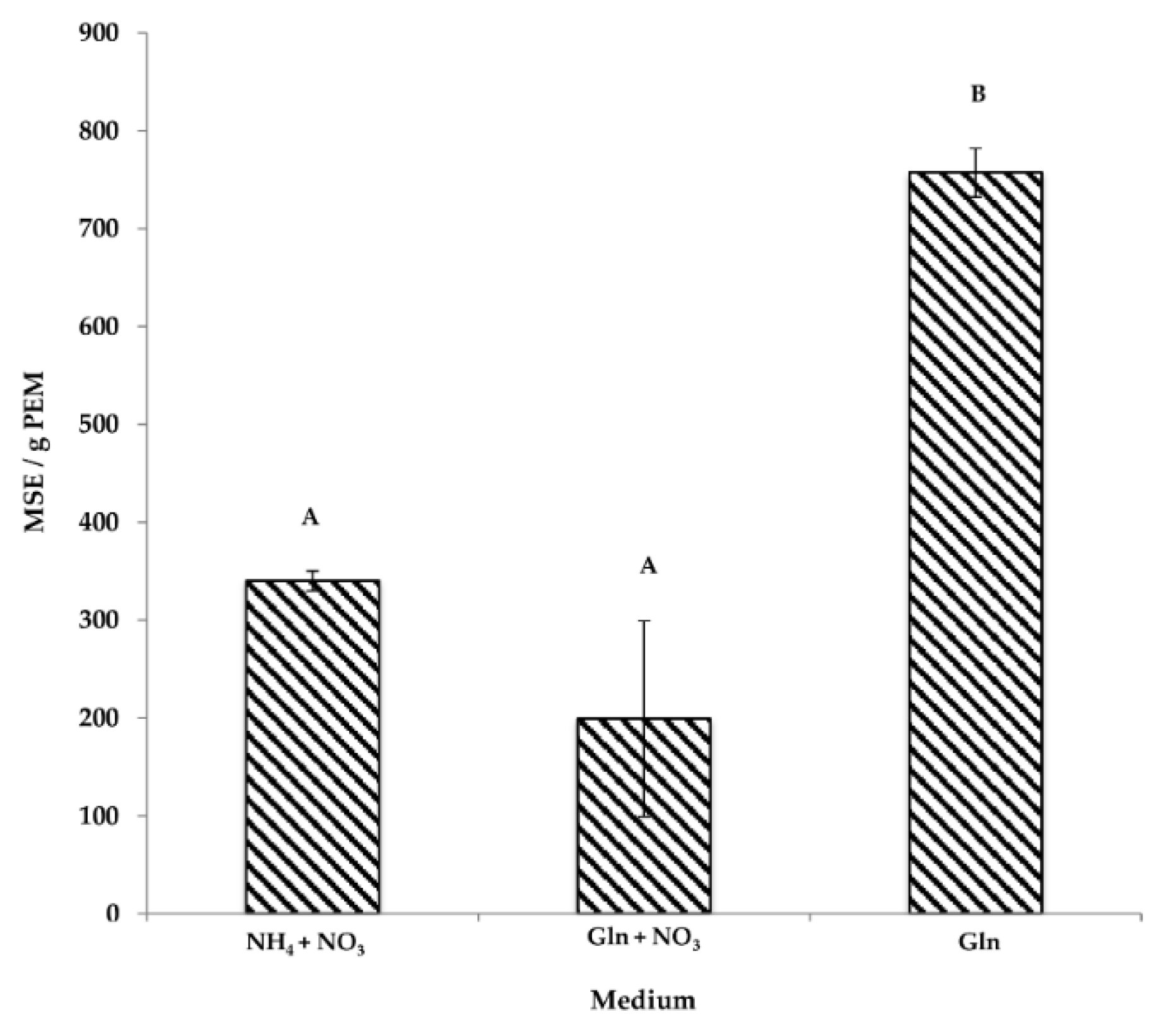
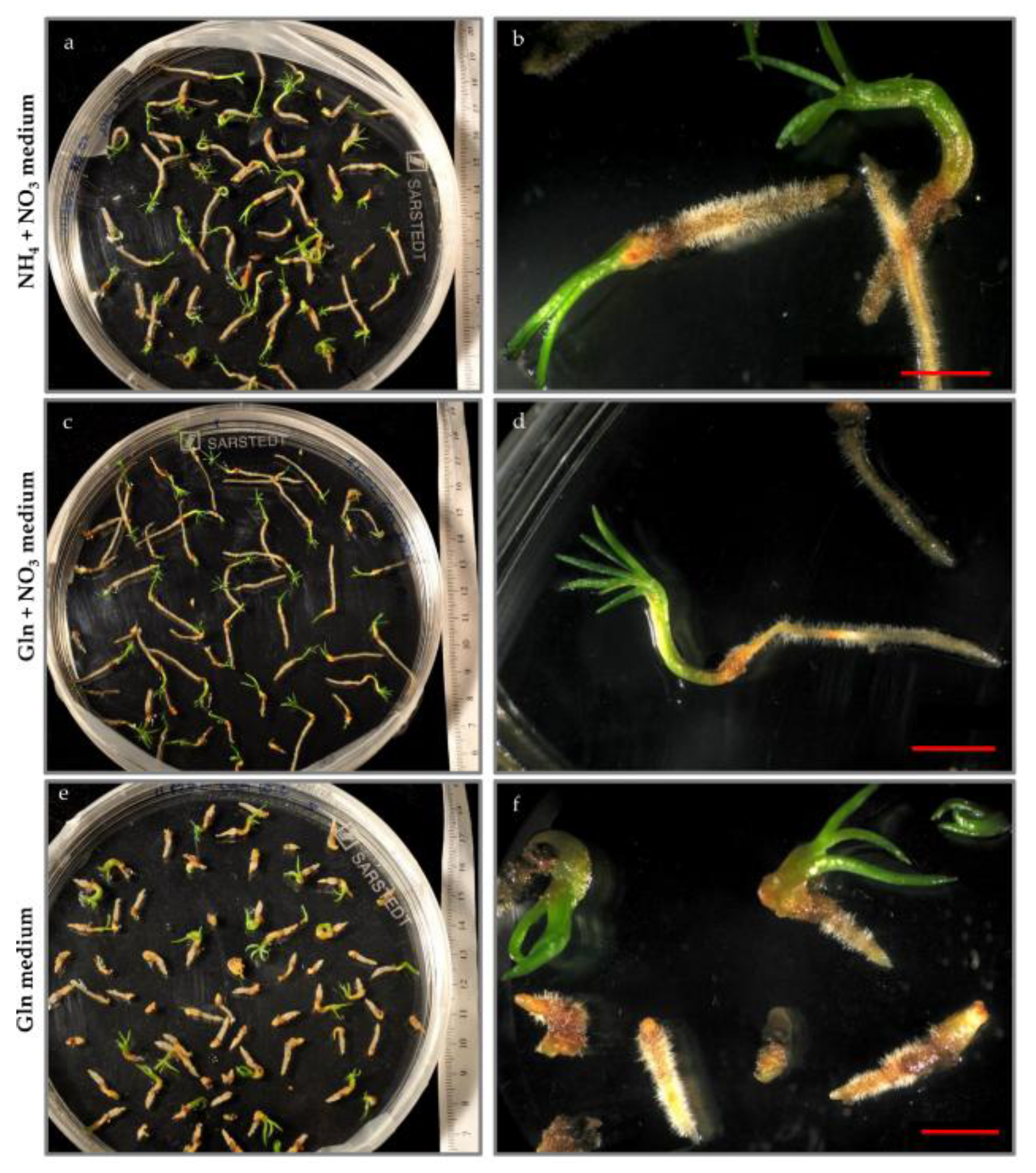
3.1. The Effect of Nitrogen Source on Somatic Embryogenesis
3.1.1. PEMs
3.1.2. Maturation
3.1.3. Germination
3.2. Enzyme Activities
3.2.1. GS Activity
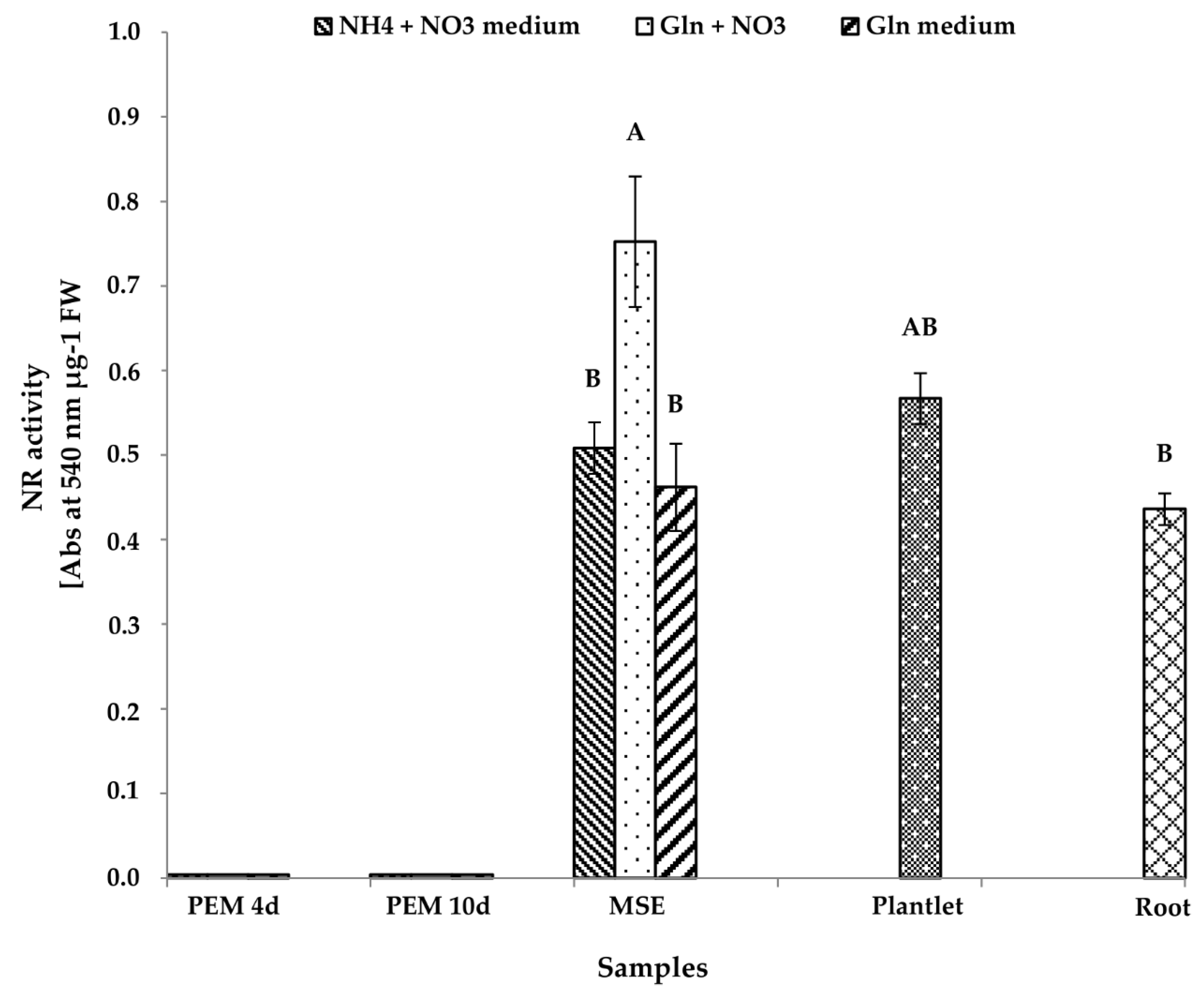
3.2.2. NR Activity
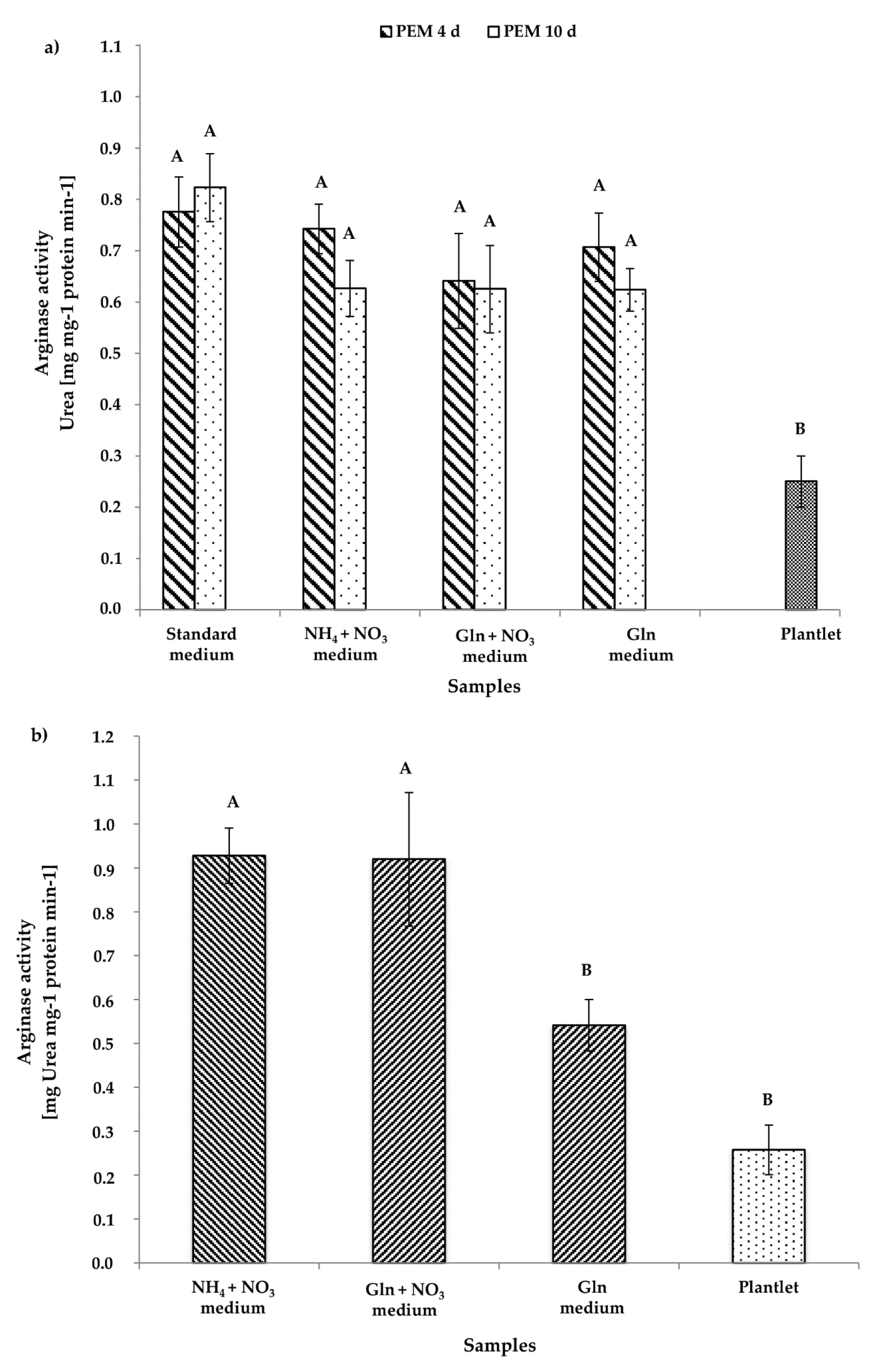
3.2.3. Arginase Activity
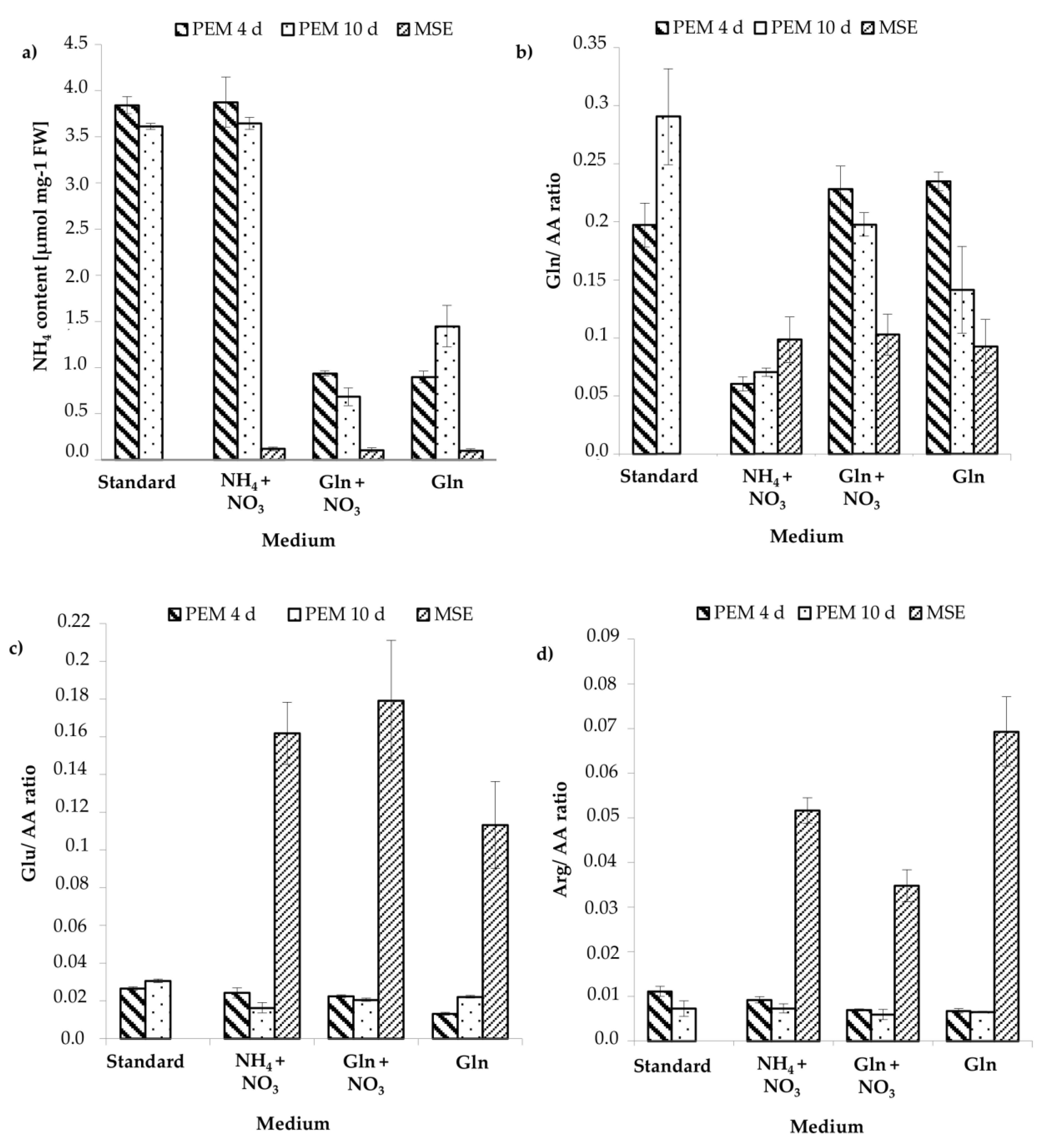
3.3. Analysis of the Content of Ammonium and Amino Acids
4. Discussion
4.1. Morphological and Developmental Characterization, and Nitrogen Content
4.2. Enzyme Activities
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Rosvall, O. Review of the Swedish Tree Breeding Programme; Forest Research Institute of Sweden: Uppsala, Sweden, 2011; pp. 1–88. [Google Scholar]
- Lindgren, D. A Way to Utilize the Advantages of Clonal Forestry for Norway Spruce? Working Papers of the Finnish Forest Research Institute: Vantaa, Finland, 2008; Volume 114, pp. 8–15. [Google Scholar]
- Funda, T.; Wennström, U.; Almqvist, C.; Andersson Gull, B.; Wang, X.-A. Mating dynamics of Scots pine in isolation tents. Tree Genet. Genomes 2016, 12, 112. [Google Scholar] [CrossRef]
- Öhlund, J.; Näsholm, T. Growth of conifer seedlings on organic and inorganic nitrogen sources. Tree Physiol. 2001, 21, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Persson, J.; Gardeström, P.; Näsholm, T. Uptake, metabolism and distribution of organic and inorganic nitrogen sources by Pinus sylvestris. J. Exp. Bot. 2006, 57, 2651–2659. [Google Scholar] [CrossRef] [PubMed]
- Gruffman, L.; Ishida, T.; Nordin, A.; Näsholm, T. Cultivation of Norway spruce and Scots pine on organic nitrogen improves seedling morphology and field performance. For. Ecol. Manag. 2012, 276, 118–124. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Siddiqi, M.; Glass, A. Conifer root discrimination against soil nitrate and the ecology of forest succession. Nature 1997, 385, 59–61. [Google Scholar] [CrossRef]
- Gruffman, L.; Jämtgård, S.; Näsholm, T. Plant nitrogen status and co-occurrence of organic and inorganic nitrogen sources influence root uptake by Scots pine seedlings. Tree Physiol. 2014, 34, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Mérigout, P.; lelandais, M.; Bitton, F.; Renou, J.P.; Briand, X.; Meyer, C.; Daniel-Vedele, F. Physiological and transcriptomic aspects of urea uptake and assimilation in Arabidopsis plants. Plant Physiol. 2008, 147, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in nitrogen utilization of crops. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, M.J.; Suárez, M.F.; Heredia, R.; Ávila, C.; Breton, D.; Trontin, J.F.; Filonova, L.; Bozhkov, P.; von Arnold, S.; Harvengt, L.; et al. Expression patterns of two glutamine synthetase genes in zygotic and somatic pine embryos support specific roles in nitrogen metabolism during embryogenesis. New Phytol. 2006, 169, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Joy, R.W., IV; Vogel, H.J.; Thorpe, T.A. Inorganic nitrogen metabolism in embryogenic white spruce cultures: A nitrogen 14/15 NMR study. J. Plant Physiol. 1997, 151, 306–315. [Google Scholar] [CrossRef]
- Buchmann, N.; Schulze, E.D.; Gebauer, G. 15N-ammonium and 15N-nitrate uptake of a 15-year-old Picea abies plantation. Oecologia 1995, 102, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Von Arnold, S.; Clapham, D. Spruce embryogenesis. In Plant Embryogenesis: Methods in Molecular Biology; Suarez, M.F., Bozhkov, P.V., Eds.; Human Press: Totowa, NJ, USA, 2008; Volume 427, pp. 31–47. [Google Scholar]
- Carlsson, J.; Svennerstam, H.; Moritz, T.; Egertsdotter, U.; Ganeteg, U. Nitrogen uptake and assimilation in proliferating embryogenic cultures of Norway spruce—Investigating the specific role of glutamine. PLoS ONE 2017, 12, e0181785. [Google Scholar] [CrossRef] [PubMed]
- Larsson, E.; Sundström, J.F.; Sitbon, F.; von Arnold, S. Expression of PaNAC01, a Picea abies CUP-SHAPED COTYLEDON orthologue, is regulated by polar auxin transport and associated with differentiation of the shoot apical meristem and formation of separated cotyledons. Ann. Bot. 2012, 110, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Kvaalen, H.; Appelgren, M. Light quality influences germination, root growth and hypocotyl elongation in somatic embryos but not in seedlings of Norway spruce. In Vitro Cell. Dev. Biol. Plant 1999, 35, 437–441. [Google Scholar] [CrossRef]
- Canovas, F.M.; Canton, F.R.; Gallardo, F.; García-Gutiérrez, A.; de Vicente, A. Accumulation of glutamine synthestase during early development of maritime pine (Pinus pinaster) seedlings. Planta 1991, 185, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Canovas, F.; Valpuesta, V.; Castro, I.N. Characterization of tomato leaf glutamine synthetase. Plant Sci. Lett. 1984, 37, 79–85. [Google Scholar] [CrossRef]
- Park, B.S.; Song, J.T.; Seo, H.S. Arabidopsis nitrate reductase activity is stimulated by the E3 SUMO ligase AtSIZ1. Nat. Commun. 2011, 2, 400. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M. The partitioning of nitrate assimilation between root and shoot of higher plants. Plant Cell Environ. 1986, 9, 511–519. [Google Scholar]
- Geyer, J.W.; Dabich, D. Rapid method for determination of arginase activity in tissue homogenates. Anal. Biochem. 1971, 39, 412–417. [Google Scholar] [CrossRef]
- Martín-Falquina, A.; Legaz, M.E. Purification and properties of constitutive arginase of Evernia prunastri. Plant Physiol. 1984, 76, 1065–1069. [Google Scholar] [CrossRef] [PubMed]
- Miflin, B.J.; Lea, P.J. Amino acid metabolism. Ann. Rev. Plant Physiol. 1977, 28, 299–329. [Google Scholar] [CrossRef]
- Durzan, D.J. Nitrogen metabolism and vegetative propagation of forest trees. In Tissue Culture in Forestry; Bonga, J.M., Durzan, D.J., Eds.; Martinus Nijhoff/Dr. W. Junk Publishers: Dordrecht, The Netherlands; Boston, MA, USA; Lancaster, UK, 1981; pp. 256–324. [Google Scholar]
- Zerihun, A.; McKenzie, B.A.; Morton, J.D. Photosynthate costs associated with the utilization of different nitrogen-forms: Influence on the carbon balance of plants and shoot–root biomass partitioning. New Phytol. 1998, 138, 1–11. [Google Scholar] [CrossRef]
- Leustek, T.; Kirkby, E.G. The influence of glutamine on growth and viability of cell suspension cultures of Douglas-fir after exposure to polyethylene glycol. Tree Physiol. 1987, 4, 371–380. [Google Scholar] [CrossRef]
- David, H.; Jarlet, E.; David, A. Effects of nitrogen source, calcium concentration and osmotic stress on protoplasts and protoplast-derived cell cultures of Pinus pinaster cotyledons. Physiol. Plant. 1984, 61, 477–482. [Google Scholar] [CrossRef]
- Kirby, E.G. The effects of organic nitrogen sources on growth of cell cultures of Douglas-fir. Physiol. Plant. 1982, 56, 114–117. [Google Scholar] [CrossRef]
- Barett, J.D.; Park, Y.S.; Bonga, J.M. The effectiveness of various nitrogen sources in white spruce [Picea glauca (Moench) Voss] somatic embryogenesis. Plant Cell Rep. 1997, 16, 411–415. [Google Scholar]
- Forsum, O.; Svennerstam, H.; Ganeteg, U.; Näsholm, T. Capacities and constraints of amino acid utilization in Arabidopsis. New Phytol. 2008, 179, 1058–1069. [Google Scholar] [PubMed]
- Higashi, K.; Kamada, H.; Harada, H. The effects of reduced nitrogenous compounds suggests that glutamine synthetase activity is involved in the development of somatic embryos in carrot. Plant Cell Tissue Organ Cult. 1996, 45, 109–114. [Google Scholar] [CrossRef]
- Stuart, D.; Strickland, S. Somatic embryogenesis from cell cultures of Medicago sativa L. II. The interaction of amino acids with ammonium. Plant Sci. Lett. 1984, 34, 175–181. [Google Scholar]
- Muralidharan, E.M.; Gupta, P.K.; Mascarenhas, A.F. Plantlet production through high frequency somatic embryogenesis in long term cultures of Eucalyptus citriodora. Plant Cell Rep. 1989, 8, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Khlifi, S.; Tremblay, F.M. Maturation of black spruce somatic embryos. Part I. Effect of l-glutamine on the number and germinability of somatic embryos. Plant Cell Tissue Organ Cult. 1995, 41, 23–32. [Google Scholar]
- Feirer, R. The biochemistry of conifer embryo development: Amino acids, polyamines and storage proteins. In Somatic Embryogenesis in Woody Plants; Jain, S., Gupta, P., Newton, R., Eds.; Kluwer Academic Press: Dordrecht, The Netherlands, 1995; Volume 1, Chapter 15; pp. 317–336. [Google Scholar]
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Krouk, G.; Crawford, G.; Coruzzi, G.M.; Tsay, Y.F. Nitrate signalling: Adaptation to fluctuating environments. Curr. Opin. Plant Biol. 2010, 13, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Takei, K.; Hirose, N. Interactions between nitrogen and cytokinin in the regulation of metabolism and development. Trends Plant Sci. 2006, 11, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 135–189. [Google Scholar]
- Cruz, C.; Bio, A.F.M.; Dominguez-Valdivia, M.D.; Aparicio-Tejo, P.M.; Lamsfus, C.; Martins-Loução, M.A. How does glutamine synthetase activity determine plant tolerance to ammonium? Planta 2006, 223, 1068–1080. [Google Scholar] [CrossRef] [PubMed]
- Kaul, K.; Hoffman, S.A. Ammonium ion inhibition of Pinus strobus L. callus growth. Plant Sci. 1993, 88, 169–173. [Google Scholar] [CrossRef]
- Chen, S.J.; Kao, C.H. Polyamines in relation to ammonium-inhibited growth in suspension-cultured rice cells. Bot. Bull. Acad. Sin. 1996, 37, 197–200. [Google Scholar]
- Niedz, R.P. Growth of embryogenic sweet organge callus on media varying in the ratio of nitrate to ammonium nitrogen. Plant Cell Tissue Organ Cult. 1994, 39, 1–5. [Google Scholar] [CrossRef]
- Chen, S.J.; Kao, C.H. Ammonium-inhibited growth of suspension-cultured rice cells as affected by medium pH. Plant Growth Regul. 1997, 21, 1–6. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Gruissem, W.; Jones, L. Interaction between nitrate assimilation and carbon metabolism. In Biochemistry and Molecular Biology of Plants; Wiley: Hoboken, NJ, USA, 2000; Chapter 16; pp. 786–849. [Google Scholar]
- King, J.E.; Gifford, D.J. Amino acid utilization in seeds of loblolly pine during germination and early seedling growth (I. Arginine and arginase activity). Plant Physiol. 1997, 113, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Bedell, J.P.; Chalot, M.; Garnier, A.; Botton, B. Effects of nitrogen source on growth and activity of nitrogen-assimilating enzymes in Douglas-fir seedlings. Tree Physiol. 1999, 19, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Tritsch, G.L.; Moore, G.E. Spontaneous decomposition of glutamine in cell culture media. Exp. Cell Res. 1962, 28, 360–364. [Google Scholar] [CrossRef]
- Cantón, F.R.; Suarez, M.F.; José-Estanyol, M.; Cánovas, F.M. Expression analysis of a cytosolic glutamine synthetase gene in cotyledons of Scots pine seedlings: Developmental light-dark regulation and spatial distribution of specific transcripts. Plant Mol. Biol. 1999, 40, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Slocum, R.D. Genes, enzymes and regulation of arginine biosynthesis in plants. Plant Physiol. Biochem. 2005, 43, 729–745. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.M.; Senaratna, T.; McKersie, B.D. Glutamine enhances storage protein synthesis in Medicago sativa L. somatic embryos. Plant Sci. 1992, 87, 69–77. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Media | ||||
|---|---|---|---|---|
| Constituent | Standard | NH4 + NO3 | Gln + NO3 | Gln |
| Concentration, mM | ||||
| KNO3 | 9.40 | 0 | 0 | 0 |
| NH4NO3 | 3.75 | 0 | 0 | 0 |
| NaNO3 | 0 | 26.58 | 26.58 | 0 |
| NH4Cl | 0 | 3.46 | 0 | 0 |
| K2SO4 | 0 | 0.72 | 0.72 | 0.72 |
| KCl | 0 | 2.56 | 2.56 | 2.56 |
| MgSO4·7H2O | 0.75 | 0.77 | 0.77 | 0.77 |
| KH2PO4 | 1.25 | 1.86 | 1.86 | 1.86 |
| CaCl2·2H2O | 1.50 | 0.74 | 0.74 | 0.74 |
| Concentration, µM | ||||
| FeSO4·7H2O | 45.75 | 91.5 | 91.5 | 91.5 |
| Na2EDTA | 25.02 | 50.03 | 50.03 | 50.03 |
| Zn-EDTA | 5.004 | 0 | 0 | 0 |
| ZnSO4·7H2O | 0 | 29.91 | 29.91 | 29.91 |
| KI | 2.26 | 5.00 | 5.00 | 5.00 |
| H3BO3 | 5.09 | 100.26 | 100.26 | 100.26 |
| MnSO4·7H2O | 5.00 | 108.26 | 108.26 | 108.26 |
| NaMoO4·2H2O | 0.007 | 1.37 | 1.37 | 1.37 |
| CuSO4·5H2O | 0.001 | 0.16 | 0.16 | 0.16 |
| CoCl2·6H2O | 0.001 | 0.19 | 0.19 | 0.19 |
| Pyridoxine | 2.95 | 0 | 0 | 0 |
| Nicotinic acid | 8.12 | 0 | 0 | 0 |
| Thiamine·HCl | 7.41 | 2.9 | 2.9 | 2.9 |
| Concentration, mM | ||||
| Myo-inositol | 1.25 | 0.28 | 0.28 | 0.28 |
| l-Glutamine | 3.08 | 0 | 6.20 | 6.20 |
| Casein hydrolysate | 0 | 3.90 | 3.90 | 3.90 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahrendorf, J.; Clapham, D.; Egertsdotter, U. Analysis of Nitrogen Utilization Capability during the Proliferation and Maturation Phases of Norway Spruce (Picea abies (L.) H.Karst.) Somatic Embryogenesis. Forests 2018, 9, 288. https://doi.org/10.3390/f9060288
Dahrendorf J, Clapham D, Egertsdotter U. Analysis of Nitrogen Utilization Capability during the Proliferation and Maturation Phases of Norway Spruce (Picea abies (L.) H.Karst.) Somatic Embryogenesis. Forests. 2018; 9(6):288. https://doi.org/10.3390/f9060288
Chicago/Turabian StyleDahrendorf, Julia, David Clapham, and Ulrika Egertsdotter. 2018. "Analysis of Nitrogen Utilization Capability during the Proliferation and Maturation Phases of Norway Spruce (Picea abies (L.) H.Karst.) Somatic Embryogenesis" Forests 9, no. 6: 288. https://doi.org/10.3390/f9060288