Post-Fire Regeneration and Diversity Response to Burn Severity in Pinus halepensis Mill. Forests




Escuela Técnica Superior Ingenieros Agrónomos y Montes, Universidad de Castilla-La Mancha, Campus Universitario, 02071 Albacete, Spain
*
Author to whom correspondence should be addressed.
Forests 2018, 9(6), 299; https://doi.org/10.3390/f9060299
Submission received: 20 April 2018
/
Revised: 23 May 2018
/
Accepted: 24 May 2018
/
Published: 26 May 2018
(This article belongs to the Special Issue Ecological Management of Pine Forests)
Abstract
:In recent decades, fire regimes have been modified by various factors such as changes in land use, global change or forest management policies. The vulnerability of Mediterranean terrestrial ecosystems is increasing due to more severe and frequent droughts. This study aimed to determine the plant response of ecosystems during the short-term post-fire period by relating alpha diversity, floristic richness and tree recruitment dynamics to burn severity 5 years after a wildfire. Our results conclude that in the short term, Pinus halepensis Mill. stands in southeastern Spain quickly recovered alpha diversity values, mainly in areas burned with low severity. We observed that moderate and high severities affected the ecosystem more significantly, showing higher values for the Shannon Index but lower for the Simpson index. Pine recruitment was higher in burned areas, and we found the highest number of Aleppo pine seedlings under a moderate burn severity. Post-fire regeneration functional groups (obligate seeders and resprouters) were promoted under moderate and high burn severity, increasing their abundance. Annual species (mainly herbs) colonized burned areas, persisting with higher presence under moderate burn severity. Restoration tools should be focused on reducing fire severity, mainly in areas at high risk of desertification, and promoting resistance, vulnerability and resilience of these ecosystems.

1. Introduction
Forest fires have been described as natural processes inherent to terrestrial biomes as they modify ecosystems to shape the current general global distribution [1,2]. For this reason, forest fires and climate are key processes that condition vegetal dynamics, biome development, soil formation, and hydrological and erosive cycles [3]. The first signs of natural forest fires date back to the Carboniferous Era, and human activity managing fire was evidenced in the early Holocene in the Mediterranean Basin, changing the natural fire regime [4].
A fire regime is defined as the characteristic of wildfire activity that prevails in a given area, mainly determined by fuel consumption patterns, intensity and severity, fire frequency, patch size and seasonality [5]. The fire regime in the Mediterranean Basin has been affected by several factors, mainly by changes in land use, global warming, prolonged drought period, changes in forest policies, and fauna or invasive plant species [6].
Wildfires of the Mediterranean Basin are a dramatic hazard for dense populations but also for terrestrial ecosystems, since alteration of fire patterns modifies vegetation resilience to fire, and predictions point to a general rise in fire risk [7]. The main changes include increases in large forest fires, which promote the size of burned areas affected by high fire severity [8,9]. In this way, monitoring and data assessment of ecosystem response to disturbances is a research priority, and Mediterranean terrestrial ecosystems serve as exemplary natural laboratories in which to study impacts and drivers [10].
The predominant plant communities in the Mediterranean Basin are characterized by their rate of natural recovery to reach a previous equilibrium following disturbance, termed engineering resilience [11]. The two most common regeneration strategies after fire are resprouters (buds in soil or protected by tissues) or obligate seeders (seeds from unburned areas, buried in the soil bank or enclosed in aerial banks) [12,13]. In some cases, seed germination can be initiated or improved by heat, the presence of burnt wood or ash as scarifying agents [14,15]. High temperatures can also trigger the opening of serotinous cones or fruits to release seeds after a fire [16].
In the Mediterranean Basin, modal natural fire frequency ranges from 25 to 50 years, which promotes a high alpha diversity after disturbance, but shorter-lived species are rapidly excluded [17]. However, fire exclusion reduces fire frequency, and promotes fire severity, which could induce changes in post-fire recovery patterns related to burn severity [18]. To avoid confounding recovery patterns related to other fire regime components, monitoring of ecosystems in the same post-fire succession stage, but burned on different dates, is required [10]. However, the large spatial scales and time periods needed to establish a fire history in an ecosystem lead to pseudo-replication [19] and have been proven to be unavoidable in our research on burn severity and α-diversity.
There are several studies on Pinus halepensis Mill. (Aleppo pine) forests in the western Mediterranean Basin and on the effects that fire severity has on plant recovery and pine recruitment, diversity and time to recover ecosystem services (e.g., soil protection, vulnerability to new fire, nutrient cycling, etc.) [13,16,18,20,21,22]. However, the importance of burn severity and the short-term ecosystem changes are important for management decisions about potential intervention in burned areas [18,23,24]. Aleppo pine is one of the most widely distributed pine species throughout low altitudinal areas of the Mediterranean Basin; this is a tree species that has adapted to fire and drought [25,26]. This pine species is a pioneering and obligate seeder, shows precocious reproduction and a dual-life strategy, bearing partially non-serotinous cones releasing seeds and storing a canopy seed bank in serotinous cones to be released after fire [27,28]. However, the amount of serotinous cones increases with higher fire recurrence [29]. Burn severity, defined as the loss or change in organic matter both aboveground and belowground [28], influences the dynamics of the structure and vegetal composition after a fire [30]. In this way, the ecosystem’s ability to return to the pre-fire state, fulfilling similar functions and services as those prior to the disturbance, could be affected [11].
The objective of this research was to assess the effect of burn severity in the short term after fire on vegetation dynamics and community structure, including pine recruitment, plant regeneration and diversity performances following a diachronic approach. For this purpose, we carried out floristic inventories by recording the recruitment of Aleppo pine and the main companion species in four different burn severity classes in stands in the south-eastern part of the Iberian Peninsula with a semiarid climate. Our goal was to improve knowledge to support tools for post-fire planning and decision making for the stabilization, rehabilitation and restoration of burned areas depending on the fire damage.
2. Materials and Methods
2.1. Study Area
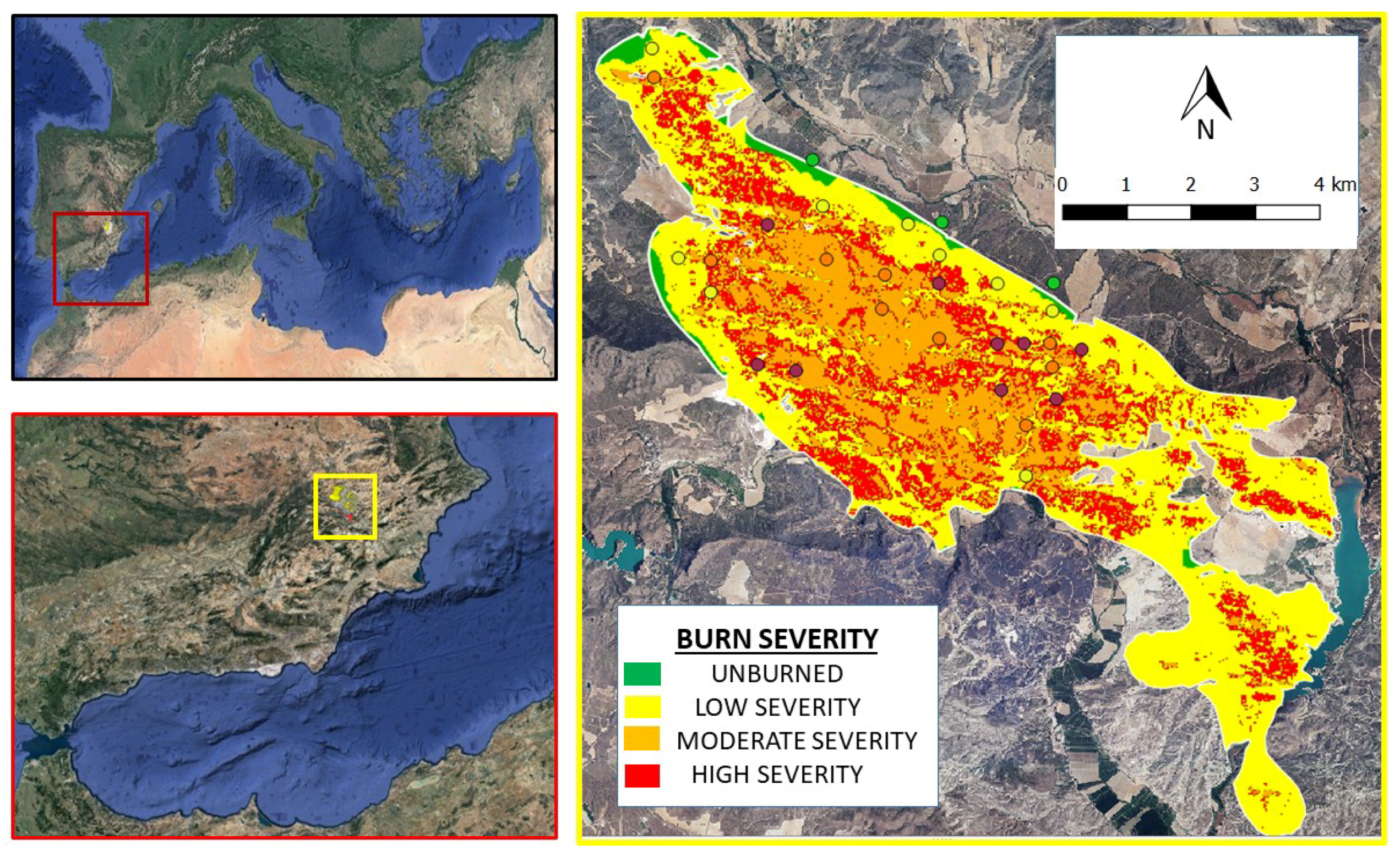
A wildfire from 1–6 July 2012 burned almost 6500 ha of the forests in “Sierra de los Donceles”, close to Hellin (the south-eastern area of the province of Albacete in Spain (ignition point: 30 S 611,807 E, 4,251,765 N)) (Figure 1) where no recurrent fires occurred for more than 80 years. During the wildfire, the maximum temperatures ranged from 31 to 37 °C (minimum average temperature was 16 °C), mean humidity ranged from 30 to 80% (although was less than 20% on the first day) and the mean wind speed was 1–3 m s−1, reaching gusts of 7–9 m s−1. The fire perimeter and weather conditions were provided by the Forest Services of the Regional Government of Castilla-La Mancha.
The study area is a semiarid Mediterranean climate, BSk, according to the Köppen-Geiger classification [31], which is in the upper meso-Mediterranean bioclimatic belt. The average precipitation and annual temperature values are 278.50 mm and 15.85 °C, respectively (based on the 25-year data period (1990–2014) provided by the National Meteorological Agency). The dry period extends from June to September, during which relative humidity is less than 50%. According to Garcia-Morote et al. [32], the total precipitation values recorded in the study area from early 2014 to late 2016 were slightly higher than the average values.
According to the Spanish Soil Map and following the USDA Soil Classification [33], the studied soils were classified as Aridisols (Lithic Haplocalcids), and as calcic soils with long periods (>90 days) of water not available for plants. According to Garcia-Morote et al. [32], they are alkaline soils with a clay content above 30% with a high cation exchange capacity (CEC) and low available P. The predominant landform is composed of dolomitic limestones forming rocky slopes that range from 500 to 700 m a.s.l.
The potential vegetation series belonged to Rhamno lycioidis-Querceto cocciferae sigmetum, but Pinus halepensis was the main tree species. The most common plant community in the area before fires occurred was Pinus halepensis Mill. (Aleppo pine) forests, accompanied by several scrub species and steppic grasslands with Macrochloa tenacissima (L.) Kunth (Alpha grass), Quercus coccifera L. (Kermes oak) and Pistacia lentiscus L. (Mastic tree).
The study site and burn severity were evaluated by using remote sensing and digital cartography provided by the Geographic Information National Centre, specifically, Digital Terrain Models (pixel size of 5 m, matrix ASCII file format, geodesic reference system ETRS89), and orthophotos (National Aviation Plan Orthophotography mosaics; ECW format, geodetic reference system ETRS89). The distribution units were set according to MTN50 (Spanish National Topographic Map, 1:50,000)) to create a map with GIS software (QGIS, GNU General Public License, June 1991, 2.8.2-Wien). Remote-sensing imagery from pre- and post-fire data (Enhanced Thematic Mapper Plus (ETM+) sensor on Landsat 7), after atmospheric correction undertaken by using the dark body subtraction technique and gap-filled, was used to create a burn severity map by calculating the dNBR (delta normalized burn ratio; [34]) by using pre- and post-fire images (23 June 2012 and 9 August 2012, respectively). We implemented a supervised classification by using real-ground data to identify and discriminate the categories of burn severity to be discriminated [35]. Following the Spanish technical guide for burned forest management [36], a systematic scheme overlapped the burned area (500 × 500 m grid cell) to validate the burn severity obtained. In 30 plots of the grid, we evaluated the burn severity in the field (November 2012) by recording ash color, soil affected and amount of burned biomass (in both the crown of trees and shrubs) [37].
2.2. Field Sampling
Following Gomez-Sanchez et al. [35], for our study area, we established four levels of burn severity (SEV hereafter): unburned for dNBR < 0.300 (UB), low severity for the values of dNBR ranging from 0.300 to 0.486 (Low), moderate severity for the dNBR threshold from 0.487 to 0.567 (Mod) and high severity for those higher than 0.568 (High).
We analyzed the response and dynamics of the plant community depending on the time after fire (TIME hereafter). A diachronic approach was followed, including three data collection campaigns (late spring in 2013, 2015 and 2017) to characterize 1, 3 and 5 years post-fire (short-term period after fire). To acquire information about the mature plant community, three control plots were sampled in undisturbed and mature stands. In the burned areas, we set nine circular plots (5-m radius; 78.5 m2) in each burn severity class, by using a Monterra GPS receiver (Garmin International, Inc., Olathe, KS, USA). A total of thirty plots were established and monitored in the spring of 2013, 2015 and 2017.
The selected plots had a similar orientation, slope and percentage of bare soil to reduce variability. Regarding vegetation, both structure and composition were evaluated by recording the coverage and plant abundance using a nested sampling technique [38]. Floristic inventory of all species in the plot followed the Braun-Blanquet approach, modified by Van der Maarel [39], and by the line transect method for plant surveys by recording species and the length intersecting the line (all on a perpendicular plane). Using the georeferenced center of the plot, three transects (3-m long) were arranged, with one oriented northward and forming 120° angles. We also established three square subplots (1 m2) in each transect to record and monitor pine seedlings. The plant survey and quantification of land cover type were carried out by the point intercept method because it is simple, unbiased, low-cost and suitable for our study area [40].
2.3. Plant Diversity and Regeneration
To characterize the diversity within a community, we used three indices related to α-diversity. We regarded the number of plant species or floristic richness (FR, number of species) and heterogeneity, including the measure of species abundance, i.e., the Shannon index (H′, nondimensional) [41] and the measure of species dominance, i.e., the Simpson index (D, nondimensional) [42].
FR was calculated as the number of plant species recorded on the sampling lines in each plot, including additional sampling in the entire plot to account for all those present in each one. H′ was measured as the relative abundance of the different species (1), and D evaluated the probability that two individuals chosen at random belonged to the same species (2).
where pi = coverage of each species (%)
where n = % coverage of each species and N = % total of the present organisms.
To characterize the Aleppo pine recruitment rate, we counted and recorded all the pine seedlings inside the three-square subplots (1 × 1 m, 1 m2). Pine recruitment was calculated as the mean pine seedling density (RECRUIT, pine m−2).
We classified the species identified in the field sampling according to their adaptive strategy to fire following the plant trait database for Mediterranean Basin species (BROT): non-adaptive species (R−S−); obligate seeders (R−S+); obligate resprouters (R+S−); facultative seeders (R+S+) [43]. To obtain life-form compositions, plant species were classified according to the life-form categories: trees (TREE), shrubs (SHRUB), dwarf shrubs (DWARF-S) and herbs (HERB) [44,45].
2.4. Statistical Analysis
Generalized linear models (GLMs) were used to evaluate the effects of categorical factors (TIME and SEV) and their interactions on the studied variables. Squared correlation coefficients and empirical p-values were calculated by one-way analysis of variance (ANOVA). We selected a critical p-value below 0.05 for the significant results. We also used the adjusted R-squared statistic, which is more suitable for comparing models than the R-squared value with different numbers of independent variables, and then the Durbin-Watson statistic to test residuals to determine whether there was any significant correlation based on the order in which they occurred in the data files. Log-transformed or rank-transformed data were used for those variables that did not meet the assumptions of normality of residuals and homogeneity of variance. The method currently used to discriminate among means is Tukey’s honestly significant difference (HSD) procedure.
Principal component analysis (PCA, varimax rotation method) was performed with all variables to obtain the structure of the dependence and correlation between variables and factors, at two levels (time after fire and burn severity). The PCA created a standardized correlation matrix covariance list-wise and reduced the variables used by employing linear combinations and accounted for most of the variability contained in the original data. Statistical analyses were performed using Statgraphics Centurion 18.1.03 (Statgraphics Technologies, Inc., Virginia, USA) [46] and the R software 3.2.1 (R Development Core Team) [47].
3. Results
In a first step, we verified that the control plots displayed no variations in the sampling period for structure and plant characteristics since the ANOVA for FR, H′, D and RECRUIT showed no significant differences. By running a GLM to analyze the result of the variables TIME, SEV and interactions were meaningful (Table 1). We found RECRUIT and FR significantly related to SE. H′ was significantly influenced by TIME and the interaction TIME*SEV, whereas D was significantly linked to TIME and SEV.
The ANOVA showed a significant influence of TIME and SEV on the studied variables (Figure S1). Independent of TIME, the values of variables studied in the control plots were not different, which implied that mature stands showed the same characteristics in the short term after the fire. One year after the fire, H′ and D were lower in burned plots than in unburned areas, whereas FR and RECRUIT were higher in burned areas. Three of the studied parameters (H′, RECRUIT and FR) increased three years after the fire, with all three SEV classes having higher values than UB, whereas D showed lower values. Five years after, H′ and FR (in all three burn severity classes) decreased to reach similar values to those found in mature stands. However, RECRUIT was significantly higher in the Low and Mod classes, but D showed lower values in Mod and High than in UB.
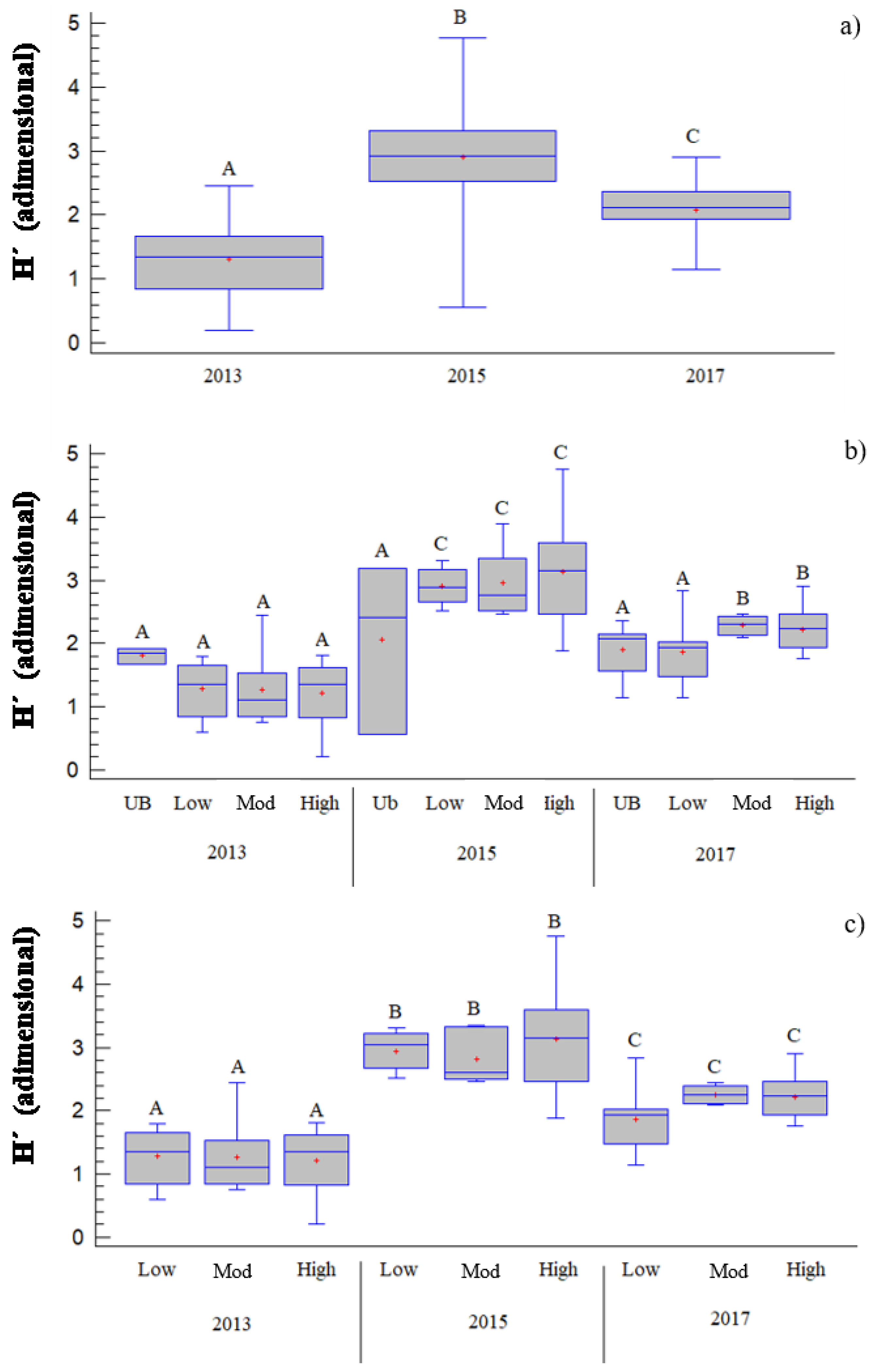
With the GLM, we ran ANOVAs for each variable according to the significant parameters. According to H′ (Figure 2), we observed that 1 year after the fire, H′ presented lower values in the burned plots than in the control plots, and roughly similar values among the severity values 3 years after the fire, H′ values increased, which were higher than the control plots with increases noted for SEV, and with higher values according to increasing burn severity. However, H′ showed a maximum value 3 years after the fire, whereas the values decreased 5 years later, with no differences between unburned plots and those burned with low severity.
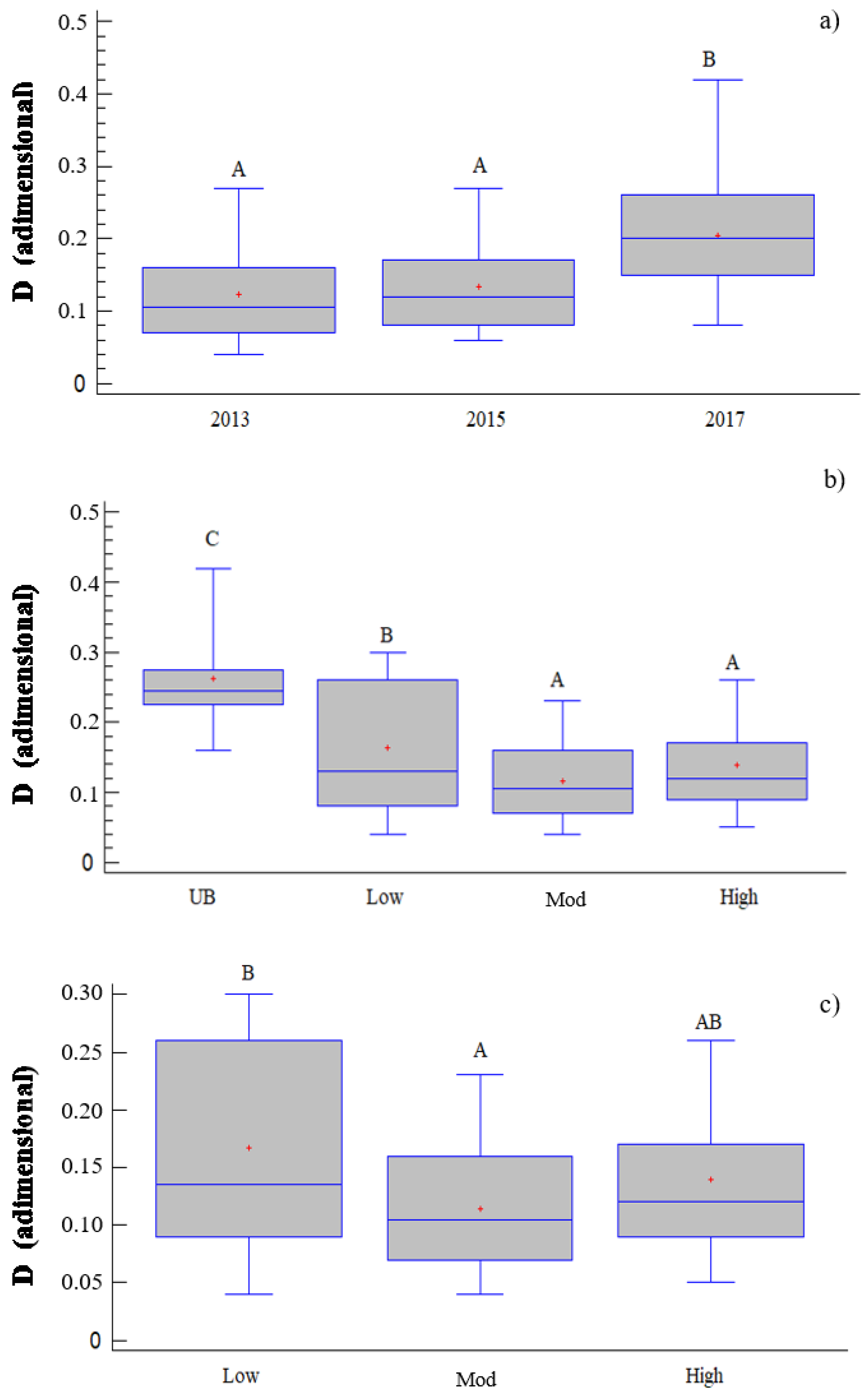
According to the analysis of the Simpson Index (Figure 3), we found that within 1 and 3 years after the fire, the D values were more similar than 5 years after the fire, which presented slightly higher values. However, when considering severity in the short term, the D values of the burned plots were always below the values presented by the control plots, and severity had a significant effect and resulted in the highest D values under low severities.
When analyzing the results obtained from the analysis without the control plots, we found that the plots affected by low severities presented slightly higher values than those affected by moderate severities, with two significantly different homogeneous groups appeared, while the plots affected by high severities presented medium values that belonged to both groups.
According to the ANOVA carried out to analyze RECRUIT (Figure 4) related to SEV, we observed that RECRUIT was lower in the UB plots than in burned areas, with the exception of High SEV, and the highest RECRUIT was found in Mod SEV (Figure 4a). We ran an ANOVA with only data from burned areas (Figure 4c) and found no significant differences for Low and High SEV, but values were lower than those observed for Mod. Regarding FR (Figure 4b), the study showed that in the short term, the values of this parameter were always higher than for the control plots, and these values were significantly similar in severity terms, although the plots affected by moderate severities had slightly higher values. When we ran a one-way ANOVA with no control plots, we observed no significant differences in the FR values according to severity.
When we carried out a descriptive analysis to represent the presence of species according to their adaptive strategy, we represented the mean values related to TIME and SEV (Table 2). We found that 1 year after the fire, the most representative adaptive strategy, independent of the burn severity level, was R−S+ (with almost 50% of the species present). During our study period, R−S+ maintained high percentages of species’ presence (always >40%) with the highest values for the Mod and High severities. However, these values slightly decreased with time. We found no changes in the percentage of the presence of R+S−, showing approximately 20% regardless of burn severity. Finally, the percentages of R+S+ were close to 20–25%, which corresponded to an increase 5 years after the fire, with similar values to the mature stands (30%).
According to growth forms and after another descriptive analysis, we observed that 1 year after the fire (in 2013), the most representative growth forms were HERB because these species had the highest presence percentages. For SEV, HERB showed a higher presence in Low SEV, decreasing according to increasing SEV. However, 3 years after the fire, the HERB percentage decreased and was not related to SEV because we found similar values in all the burned plots, with the predominant growth shown for DWARF-S. Five years after the fire, the presence of HERB and Dwarf-S was poor in the burned plots, with SHRUB as the most common growth form. This stage showed similar percentages for each growth form as the mature and unburned stands. TREE values had the lowest percentage values, which were similar when comparing TIME and SEV.
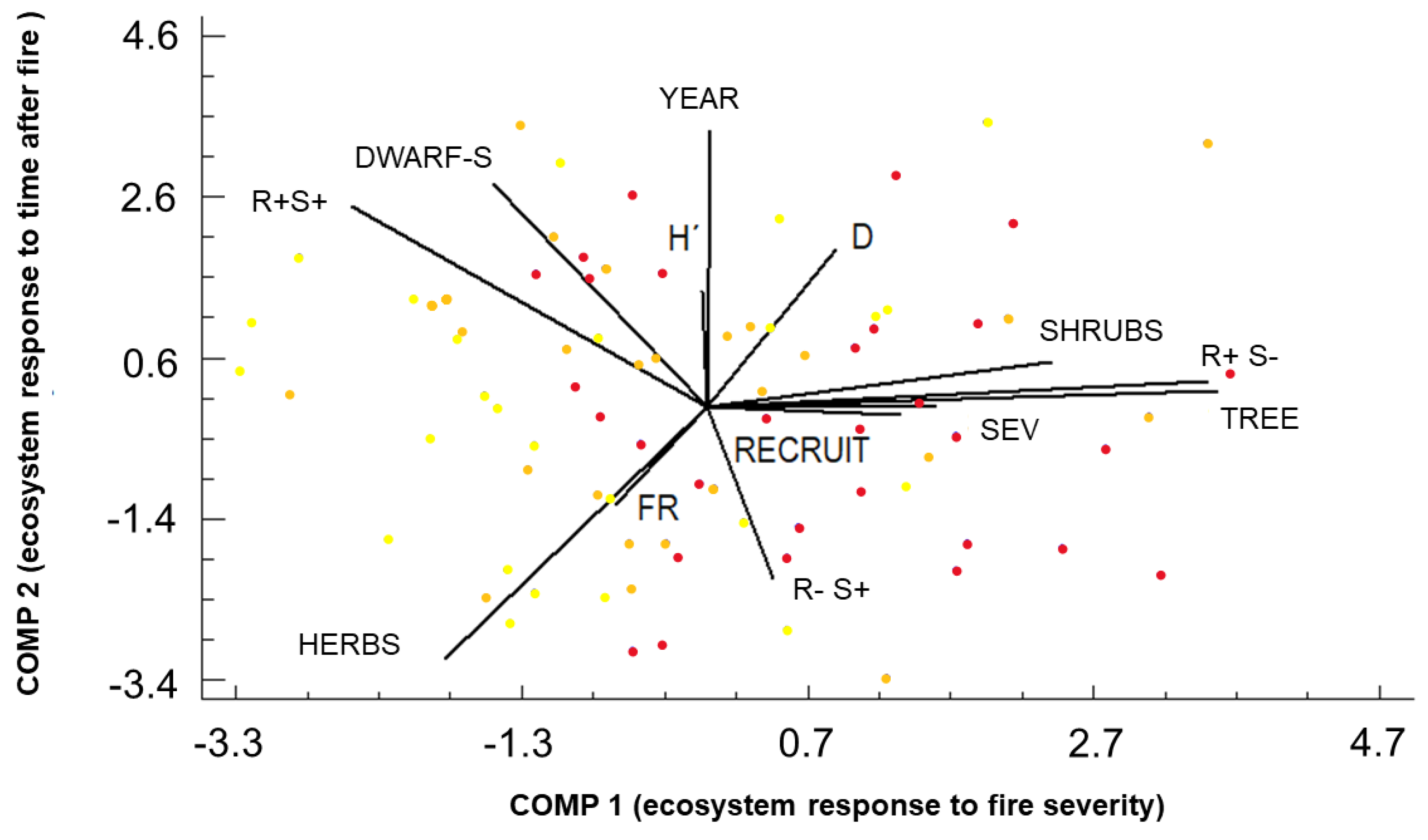
A PCA (Figure 5), which included the results for the burned area, reduced the variables to a five-component solution (eigenvalues > 1) and explained more than 73.84% of variability but this was simplified to a two-component solution which included 39.17% of the variability in the original data. The first component (COMP1) accounted for 21.17%, while the second explained 18.00% (COMP2). The ecosystem responses to burn severity (SEV) were clustered along the COMP1 axis, while that related to time after the fire (YEAR) was clustered on the COMP2 axis.
4. Discussion
Changes in fire regimes and the effects on plant communities, mainly in terms of plant (alpha) diversity and recovery time of Mediterranean plant communities, are a major trend to manage natural and seminatural landscapes in ecosystem conservation [48]. Thus, our approach demonstrated a relationship between burn severity, time after fire passage and the recovery/regeneration of the plant community in P. halepensis stands in the SE Iberian Peninsula.
The results indicated that the α-diversity values were lower in the burned area than in the control plots, mainly in the areas burned by high fire severity, which reduced seedbanks and increased colonizer species cover [49,50]. Lopez-Poma and Bautista [22] found different effects of fire severity, depending on soil damage, which we linked to species and adaptive strategies (related to growth forms). We also found higher and increasing floristic richness in burned areas, but the highest values were recorded for the Moderate severity, which also had the highest RECRUIT value that would encourage the plant community toward a more mature successional stage in the short term. On the contrary, H′ increased according to the time after fire and burn severity, showing a maximum value under the high burn severity in 2017. To avoid an interaction of the unburned plots, we found marked differences in the specific diversity values for years 5 after the fire, with values remaining above those of the more severely burned plots. This indicated that there is a sustainable fire frequency (more than 25 years) since the high alpha diversity decreases rapidly in the short term [51] although a high burn severity could induce changes in post-fire recovery patterns [17,22].
Post-fire succession chrono-sequences in the Mediterranean Basin suggest that post-fire succession is dominated by herbaceous taxa in the short term [52], according to the initial floristic composition model of Egler [20], which concluded that herbaceous plants rapidly appeared and colonized bare ground after fire. In our assessment, the highest cumulative number of species was found 3 years after the fire due to the presence of herbaceous species, heliophiles and pioneer species, established due to low interspecific competition. These colonizer and pioneer species disappeared 3 years after the fire, probably due to successional dynamics, including successive species removal related to intra- and inter-specific competition [53].
The plant community included no fire-intolerant plant species, with post-fire obligate seeders predominating, mainly those comprising persistent seed banks enclosed in soil (Cistus) or in the canopy (Pinus). Nonetheless, some species (HERBS in early years) may come from unburned patches or outside the fire perimeter (metapopulation dynamics) [54]. The facultative seeders showed their highest percentages under the moderate burn severity and obligate resprouters were not apparently affected by burn severity, similar to other Mediterranean communities [55]. According to time after fire, heliophiles and pioneer species were replaced with shrubs, with increasing tree cover, but obligate resprouters were not affected by burn severity. We found no clear evidence that variation in burn severity modulates the seeders to resprouters abundance ratio, which contrasts with other findings [23,56,57]. However, RECRUIT was promoted by fire, with lower values for Low and High severities than for Moderate severity, and where the conditions had been related to the killing of seeds enclosed in cones [58], competition of germinated seedlings [59] or diminished regeneration capacity [21].
5. Conclusions
Adaptive strategies developed by plants evolved in Mediterranean climates provide resilience to regenerate after disturbances such as wildfires (time variation according to natural productivity and sustainable fire regime) [60]. However, it is important to acquire knowledge about the ecological effects of fire severity and the influence on the time extension for the immaturity risk [61] to apply a proper adaptive forest management under changing scenarios.
A diachronic study was used to improve the assessment of fire effects in Mediterranean forests, mainly in the short term. In Aleppo pine forests in semiarid stands, alpha diversity quickly recovers after fire, but is higher in the plots burned according to moderate and high burn severity in terms of species complexity and abundance. Floristic richness and pine recruitment are promoted by moderate burn severity. When considering the adaptive strategies and growth forms, we conclude that 5 years after the fire, plant composition apparently follows the mature successional stage and shows similar values to closer mature stands, but moderate and high severity will induce higher risk of fire due to the remaining percentages of herbs and dwarf shrubs.
Therefore, fire prevention should not focus only on prevention of fire risk but also on promoting the frequency and severity to similar levels as in a sustainable fire regime [28]. Once fire occurs, it is important to bear in mind to include in post-fire management and restoration actions that moderately burned areas will have a high natural potential recovery, but those under high severity could show a lack of pine recruitment and dominance of species.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/9/6/299/s1, Figure S1: Descriptive comparison procedure to determine patterns of (a) Shannon index (H′, nondimensional), (b) Simpson index (D, nondimensional), (c) Aleppo pine recruitment rate (RECRUIT, pine seedlings m−2) and (d) Floristic richness index (FR, number of species) related to time after fire (TIME, years after fire (one (2013), three (2015) and five (2017))) and burn severity classes (SEV, UB = Unburned (green line)).
Author Contributions
All authors participated and significantly contributed to the editing of the manuscript. J.D.l.H. supervised and coordinated the experiment and improved the experimental design. D.M. and S.G.-D.V. oversaw the experimental design, conducted the field experiments and analyzed the results, ran statistical analyses, and compiled the Tables and Figures. All authors contributed to manuscript writing and figure drafting.
Funding
This research was funded by the Excma. Diputación Albacete with a grant for research initiation (DIPU4-AB2015), the Spanish National Institute for Agricultural and Food Research and Technology (INIA), i.e., the GEPRIF project (RTA2014-00011-C06-05) and the Universidad de Castilla-La Mancha with support to the Forest Ecology Research Group.
Acknowledgments
We acknowledge the Forest Services of the Regional Government of Castilla-La Mancha (Junta de Comunidades de Castilla-La Mancha) for field support, mainly Elena Gomez. We also thank Helen Warburton for the professional English editing and suggestions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Keeley, J.E. A burning story: The role of fire in the history of life. BioScience 2009, 59, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Cerdá, A.; Mataix-Solera, J. Efectos de los Incendios Forestales Sobre los Suelos en España: El Estado de la Cuestión visto por los Científicos Españoles; FUEGORED, Cátedra Divulgación de la Ciencia (Universitat de Valencia): Valencia, Spain, 2009; 529p. [Google Scholar]
- Scott, A.C. The Pre-Quaternary history of fire. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 164, 281–329. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Abrupt climate-independent fire regime changes. Ecosystems 2014, 17, 1109–1120. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Doblas-Miranda, E.; Martínez-Vilalta, J.; Lloret, F.; Álvarez, A.; Ávila, A.; Bonet, F.J.; Brotons, L.; Castro, J.; Curiel Yuste, J.; Díaz, M.; et al. Reassessing global change research priorities in Mediterranean terrestrial ecosystems: How far have we come and where do we go from here? Glob. Ecol. Biog. 2015, 24, 25–43. [Google Scholar] [CrossRef]
- Moriondo, M.; Good, P.; Durao, R.; Bindi, M.; Giannakopoulos, C.; Corte-Real, J. Potential impact of climate change on fire risk in the Mediterranean area. Clim. Res. 2006, 31, 85–95. [Google Scholar] [CrossRef]
- San-Miguel-Ayanz, J.; Schulte, E.; Schmuck, G.; Camia, A.; Strobl, P.; Liberta, G.; McInerney, D. Comprehensive monitoring of wildfires in Europe: The European forest fire information system (EFFIS). Approaches to Managing Disaster-Assessing Hazards, Emergencies and Disaster Impacts. Intech Open. 2012. Available online: https://www.intechopen.com/books/approaches-to-managing-disaster-assessing-hazards-emergencies-and-disaster-impacts/comprehensive-monitoring-of-wildfires-in-europe-the-european-forest-fire-information-system-effis- (accessed on 25 May 2018).
- Doblas-Miranda, E.; Alonso, R.; Arnan, X.; Bermejo, V.; Brotons, L.; de las Heras, J.; Estiarte, M.; Hódar, J.A.; Llorens, P.; Lloret, F.; et al. A review of the combination among global change factors in forests, shrublands and pastures of the Mediterranean Region: Beyond drought effects. Glob. Planet. Chang. 2017, 148, 42–54. [Google Scholar] [CrossRef]
- Newton, A.C.; Cantarello, E. Restoration of forest resilience: An achievable goal? New For. 2015, 46, 645–668. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.J.; Midgley, J.J.; Lamont, B.B.; Ojeda, F.; Burrows, G.E.; Knox, K.J.E. Resprouting as a key functional trait: How buds, protection and resources drive persistence after fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of South-Eastern Spain: A comparison between Pinus halepensis Mill.; P. pinaster Ait.; P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- Reyes, O.; Trabaud, L. Germination behaviour of 14 Mediterranean species in relation to fire factors: Smoke and heat. Plant Ecol. 2009, 202, 113. [Google Scholar] [CrossRef]
- Moreira, F.; Catry, F.X.; Rego, F.; Bacao, F. Size-dependent pattern of wildfire ignitions in Portugal: When do ignitions turn into big fires? Landsc. Ecol. 2010, 25, 1405–1417. [Google Scholar] [CrossRef]
- Moya, D.; De las Heras, J.; López-Serrano, F.R.; Leone, V. Optimal intensity and age of management in young Aleppo pine stands for post-fire resilience. For. Ecol. Manag. 2008, 255, 3270–3280. [Google Scholar] [CrossRef]
- González-De Vega, S.; De Las Heras, J.; Moya, D. Resilience of Mediterranean terrestrial ecosystems and fire severity in semiarid areas: Responses of Aleppo pine forests in the short, mid and long term. Sci. Total Environ. 2016, 573, 1171. [Google Scholar] [CrossRef] [PubMed]
- Viana-Soto, A.; Aguado, I.; Martínez, S. Assessment of Post-Fire Vegetation Recovery Using Fire Severity and Geographical Data in the Mediterranean Region (Spain). Environments 2017, 4, 9. [Google Scholar] [CrossRef]
- Schwilk, D.W.; Keeley, J.E.; Bond, W.J. The intermediate disturbance hypothesis does not explain fire and diversity pattern in Fynbos. Plant Ecol. 1997, 132, 77–84. [Google Scholar] [CrossRef]
- Verroios, G.; Georgiadis, T. Post-fire vegetation succession: The case of Aleppo pine (Pinus halepensis Miller) forests of Northern Achaia (Greece). Fresenius Environ. Bull. 2002, 11, 186–193. [Google Scholar]
- García-Jiménez, R.; Palmero-Iniesta, M.; Espelta, J.M. Contrasting effects of fire severity on the regeneration of Pinus halepensis Mill. and resprouter species in recently thinned thickets. Forests 2017, 8, 55. [Google Scholar] [CrossRef]
- López-Poma, R.; Bautista, S. Plant regeneration functional groups modulate the response to fire of soil enzyme activities in a Mediterranean shrubland. Soil Biol. Biochem. 2014, 79, 5–13. [Google Scholar] [CrossRef]
- Pereira, P.; Francos, M.; Brevik, E.C.; Ubeda, X.; Bogunovic, I. Post-fire soil management. Curr. Opin. Environ. Sci. Health 2018, 5, 26–32. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteor. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Quezel, P. Taxonomy and biogeography of Mediterranean pines (Pinus halepensis and P. brutia). In Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Trabaud, L., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 1–12. [Google Scholar]
- Sarris, D.; Christodoulakis, D.; Korner, C. Impact of recent climatic change on growth of low elevation eastern Mediterranean forest trees. Clim. Chang. 2011, 106, 203–223. [Google Scholar] [CrossRef]
- Goubitz, S.; Nathan, R.; Roitemberg, D.; Shmida, A.; Ne’eman, G. Canopy seed bank structure in relation to: Fire, tree size and density. Plant Ecol. 2004, 173, 191–201. [Google Scholar] [CrossRef]
- Keeley, J.E.; Bond, W.J.; Bradstock, R.A.; Pausas, J.G.; Rundel, P.W. Fire in Mediterranean Ecosystems: Ecology, Evolution and Management; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Hernández-Serrano, A.; Verdú, M.; González-Martínez, S.C.; Pausas, J.G. Fire structures pine serotiny at different scales. Am. J. Bot. 2013, 100, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- DeRose, R.J.; Long, J.N. Resistance and resilience: A conceptual framework for silviculture. For. Sci. 2014, 60, 1205–1212. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Rieser, C.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Koppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- García-Morote, F.A.; Martínez-García, E.; Andrés-Abellán, M.; Caballero, E.R.; Miettinen, H.; López-Serrano, F.R. Direct seeding of Pinus halepensis Mill. for recovery of burned Semi-Arid Forests: Implications for post-fire management for improving natural regeneration. Forests 2017, 8, 353. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Miller, J.D.; Thode, A.E. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta normalized burn ratio (dNBR). Remote Sens. Environ. 2007, 109, 66–80. [Google Scholar] [CrossRef]
- Gómez-Sánchez, E.; de las Heras, J.; Lucas-Borja, M.; Moya, D. Assessing fire severity in semi-arid environments: Application in Donceles 2012 wildfire (SE Spain). Revista Teledetección 2017, 49, 103–113. [Google Scholar] [CrossRef]
- Alloza, J.A.; García, S.; Gimeno, T.; Baeza, M.J.; Vallejo, V.R. Guía Técnica Para la Gestión de Montes Quemados; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2014; 188p. [Google Scholar]
- Ryan, K.; Noste, N. Evaluating prescribed fires. Proceedings of the Symposium and Workshop on Wilderness Fire, Missoula, Montana, 15 November 1983. USDA Forest Service Intermountain Forest and Range Experiment Station, General Technical Report INT 182. 1985, pp. 230–238. Available online: https://www.fs.fed.us/rm/publications/titles/int_gtr.html (accessed on 25 May 2018).
- Stohlgren, T.J.; Falkner, M.; Schell, D.L. A Modified-Whittaker nested vegetation sampling method. Plant Ecol. 1995, 117, 113–121. [Google Scholar] [CrossRef]
- Van der Maarel, E. Transformation of cover-abundance values for appropriate numerical treatment–Alternatives to the proposals by Podani. J. Veg. Sci. 2007, 18, 767–770. [Google Scholar]
- Buckland, S.T.; Borchers, D.L.; Johnston, A.; Henrys, P.A.; Marques, T.A. Line transect methods for plant surveys. Biometrics 2007, 63, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C. A mathematical theory of communication. Bell. Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Paula, S.; Arianoutsou, M.; Kazanis, D.; Tavsanoglu, Ç.; Lloret, F.; Buhk, C.; Ojeda, F.; Luna, B.; Moreno, J.M.; Rodrigo, A.; et al. Fire-related traits for plant species of the Mediterranean Basin. Ecology 2009, 90, 1420. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea; Cambridge University Press: Cambridge, UK, 1964. [Google Scholar]
- Castroviejo, S. Flora ibérica. Available online: http://www.floraiberica.es/ (accessed on 24 May 2018).
- Statgraphics Centurion 18.1.03; Statpoint Technologies Inc.: Warrenton, VA, USA, 2018.
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Kruger, F.J. Plant Community Diversity and Dynamics in Relation to Fire. In Mediterranean-Type Ecosystems; Kruger, F.J., Mitchell, D.T., Jarvis, J.U.M., Eds.; Ecological Studies (Analysis and Synthesis); Springer: Berlin/Heidelberg, Germany, 1983. [Google Scholar]
- Ghermandi, L.; Gonzalez, S.; Lescano, M.N.; Oddi, F. Effects of fire severity on early recovery of Patagonian steppes. Int. J. Wildland Fire 2013, 22, 1055–1062. [Google Scholar] [CrossRef]
- Zedler, P.H.; Gautier, C.R.; McMaster, G.S. Vegetation change in response to extreme events: The effect of a short interval between fires in California chaparral and coastal scrub. Ecology 1983, 64, 809–818. [Google Scholar] [CrossRef]
- Cowling, R.M.; Rundel, P.W.; Lamont, B.B.; Arroyo, M.K.; Arianoutsou, M. Plant diversity in mediterranean-climate regions. Trends Ecol. Evol. 1996, 11, 362–366. [Google Scholar] [CrossRef]
- Kazanis, D.; Arianoutsou, M. Vegetation Composition in a Post-Fire Successional Gradient of Pinus halepensis Forests in Attica, Greece. Int. J. Wildland Fire 1996, 6, 83–91. [Google Scholar] [CrossRef]
- Bond, W.J.; Van Wilgen, B.W. Fire and Plants (population and community biology series 14). Popul. community Biol. 1996, 263. [Google Scholar]
- Pausas, J.G.; Keeley, J. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Marais, K.E.; Pratt, R.B.; Jacobs, S.M.; Jacobsen, A.L.; Esler, K.J. Postfire regeneration of resprouting mountain fynbos shrubs: Differentiating obligate resprouters and facultative seeders. Plant Ecol. 2014, 215, 195–208. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E. Plant Functional traits in relation to fire in crown-fire ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef]
- Hollingsworth, T.N.; Johnstone, J.F.; Bernhardt, E.L.; Chapin, F.S., III. Fire severity filters regeneration traits to shape community assembly in Alaska’s boreal forest. PLoS ONE 2013, 8, e56033. [Google Scholar] [CrossRef] [PubMed]
- Moya, D.; De las Heras, J.; Salvatore, R.; Valero, E.; Leone, V. Fire intensity and serotiny: Response of germination and enzymatic activity in seeds of Pinus halepensis Mill. from southern Italy. Ann. For. Sci. 2013, 70, 49–59. [Google Scholar] [CrossRef]
- Sagra, J.; Plaza-Álvarez, P.A.; Moya, D.; Lucas-Borja, M.E.; Alfaro Sánchez, R.; de las Heras Ibáñez, J.; Ferrandis, P. Predation influence on seedling recruitment in Mediterranean forests after prescribed burning. Forests 2017, 8, 243. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E.; Schwilk, D.W. Flammability as an ecological and evolutionary driver. J. Ecol. 2017, 105, 289–297. [Google Scholar] [CrossRef]
- Ne’eman, G.; Goubitz, S.; Nathan, R. Reproductive traits of Pinus halepensis in the light of fire-A critical review. Plant Ecol. 2004, 171, 69–79. [Google Scholar] [CrossRef]
Figure 1.
Sierra de Los Donceles wildfire, close to Hellin (province of Albacete, Spain). Burn severity levels were: unburned (uncolored), low severity (yellow), moderate severity (orange) and high severity (red) obtained from [35]. The sampling plots were represented according burn severity: unburned in green, low severity in yellow, moderate severity in orange and high severity in red.
Figure 1.
Sierra de Los Donceles wildfire, close to Hellin (province of Albacete, Spain). Burn severity levels were: unburned (uncolored), low severity (yellow), moderate severity (orange) and high severity (red) obtained from [35]. The sampling plots were represented according burn severity: unburned in green, low severity in yellow, moderate severity in orange and high severity in red.

Figure 2.
Box and Whisker plot of the one-way ANOVA relating the Shannon Index (H′, nondimensional) to (a) time after fire (TIME, years) and (b) the interaction of time after fire and burn severity classes (SEV, UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity). In addition, we checked the significance between (c) time after fire and burn severity classes, but we excluded the unburned plots. The different capital letters indicate significant differences between means of groups (ANOVA according to Tukey’s HSD method).
Figure 2.
Box and Whisker plot of the one-way ANOVA relating the Shannon Index (H′, nondimensional) to (a) time after fire (TIME, years) and (b) the interaction of time after fire and burn severity classes (SEV, UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity). In addition, we checked the significance between (c) time after fire and burn severity classes, but we excluded the unburned plots. The different capital letters indicate significant differences between means of groups (ANOVA according to Tukey’s HSD method).

Figure 3.
Box and Whisker plot of the one-way ANOVA relating the Simpson Index (D, nondimensional) to (a) time after fire (TIME, years) and (b) burn severity classes (SEV, UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity). In addition, (c) burn severity classes and the Simpson Index were related after excluding the unburned plots. The different capital letters indicate significant differences between the means of groups (ANOVA according Tukey’s HSD method).
Figure 3.
Box and Whisker plot of the one-way ANOVA relating the Simpson Index (D, nondimensional) to (a) time after fire (TIME, years) and (b) burn severity classes (SEV, UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity). In addition, (c) burn severity classes and the Simpson Index were related after excluding the unburned plots. The different capital letters indicate significant differences between the means of groups (ANOVA according Tukey’s HSD method).

Figure 4.
Box and Whisker plot of the one-way ANOVA relating the burn severity classes (SEV, UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity) to (a) the Aleppo pine recruitment rate, calculated as the mean pine seedling density (RECRUIT, pine m−2) and (b) floristic richness (FR, number of species). In addition, (c) burn severity classes and floristic richness were related when the unburned plots were excluded. The different capital letters indicate significant differences between means of groups (ANOVA according Tukey’s HSD method).
Figure 4.
Box and Whisker plot of the one-way ANOVA relating the burn severity classes (SEV, UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity) to (a) the Aleppo pine recruitment rate, calculated as the mean pine seedling density (RECRUIT, pine m−2) and (b) floristic richness (FR, number of species). In addition, (c) burn severity classes and floristic richness were related when the unburned plots were excluded. The different capital letters indicate significant differences between means of groups (ANOVA according Tukey’s HSD method).

Figure 5.
The PCA that included the recorded variables in the burned area: H′ (Shannon Diversity Index, nondimensional), FR (Floristic Richness, number of species), SEV (burn severity, quantitative levels), TIME (period of time since the wildfire event to the sampling date, years), adaptive strategies (R−S+, R+S− and R+S+) and growth form (trees (TREE), shrubs (SHRUB), dwarf shrubs (DWARF-S) and herbs (HERB)). Dots represent the standardized values for the short-term period after the wildfire event (1, 3 and 5 years after) in light orange (low burn severity), dark orange (moderate burn severity) and red (high burn severity). COMP1: first component (ecosystem response to burn severity) accounts for 21.17% of the variance; COMP2: second component (ecosystem response to time after fire) explains 18.00% of the variance (both accounted for 39.17% of the total variability contained in the original data when added together).
Figure 5.
The PCA that included the recorded variables in the burned area: H′ (Shannon Diversity Index, nondimensional), FR (Floristic Richness, number of species), SEV (burn severity, quantitative levels), TIME (period of time since the wildfire event to the sampling date, years), adaptive strategies (R−S+, R+S− and R+S+) and growth form (trees (TREE), shrubs (SHRUB), dwarf shrubs (DWARF-S) and herbs (HERB)). Dots represent the standardized values for the short-term period after the wildfire event (1, 3 and 5 years after) in light orange (low burn severity), dark orange (moderate burn severity) and red (high burn severity). COMP1: first component (ecosystem response to burn severity) accounts for 21.17% of the variance; COMP2: second component (ecosystem response to time after fire) explains 18.00% of the variance (both accounted for 39.17% of the total variability contained in the original data when added together).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
General Linear Models relating α-diversity variables: H′ (Shannon Index, nondimensional), D (Simpson Index, nondimensional), RECRUIT (recruitment, pine seedlings m−2) and FR (floristic richness, number of species).
Table 1.
General Linear Models relating α-diversity variables: H′ (Shannon Index, nondimensional), D (Simpson Index, nondimensional), RECRUIT (recruitment, pine seedlings m−2) and FR (floristic richness, number of species).
| p-Value | F-Ratio | Adj R2 | Durbin Watson | Factor | F-Ratio | p-Value | |
|---|---|---|---|---|---|---|---|
| H′ | <0.01 | 18.78 | 27.66 | 1.20 | TIME | 28.96 | <0.01 |
| TIME*SEV | 6.42 | 0.01 | |||||
| D | <0.01 | 29.39 | 37.91 | 1.77 | TIME | 17.45 | <0.01 |
| SEV | 37.39 | <0.01 | |||||
| RECRUIT | 0.01 | 6.42 | 5.56 | 2.18 | SEV | 6.42 | 0.01 |
| FR | 0.02 | 5.26 | 4.34 | 1.51 | SEV | 5.26 | 0.02 |
Factors studied were years after fire (TIME), burn severity level (SEV) and the interaction of both (TIME*SEV).
Table 2.
Descriptive analysis including the mean values of reproductive strategies (non-adapted species (R−S−), obligate seeders (R−S+), obligate resprouters (R+S−) and facultative seeders (R+S+)) and growth form categories (trees (TREE), shrubs (SHRUB), dwarf shrubs (DWARF-S) and herbs (HERB) related to both time after fire (1, 3 and 5 years after fire, i.e. sampling carried out in 2013, 2015 and 2017 respectively) and the burn severity classes (UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity).
Table 2.
Descriptive analysis including the mean values of reproductive strategies (non-adapted species (R−S−), obligate seeders (R−S+), obligate resprouters (R+S−) and facultative seeders (R+S+)) and growth form categories (trees (TREE), shrubs (SHRUB), dwarf shrubs (DWARF-S) and herbs (HERB) related to both time after fire (1, 3 and 5 years after fire, i.e. sampling carried out in 2013, 2015 and 2017 respectively) and the burn severity classes (UB = unburned; Low = low burn severity; Mod = moderate burn severity; High = high burn severity).
| Regeneration Strategy | Growth Form | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| R−S− | R−S+ | R+S− | R+S+ | TREE | SHRUB | DWARF-S | HERB | ||
| 2013 | UB | 0.00 ± 0.00 | 50.00 ± 5.77 | 16.11 ± 4.55 | 33.89 ± 3.89 | 8.89 ± 1.11 | 23.89 ± 4.55 | 39.44 ± 5.30 | 27.78 ± 9.09 |
| Low | 0.00 ± 0.00 | 54.54 ± 7.06 | 16.08 ± 2.29 | 23.54 ± 1.33 | 11.56 ± 1.35 | 16.07 ± 2.35 | 19.37 ± 1.44 | 50.66 ± 1.80 | |
| Mid | 1.04 ± 0.68 | 54.10 ± 2.40 | 18.03 ± 2.25 | 26.83 ± 1.73 | 12.12 ± 2.03 | 25.49 ± 1.66 | 16.23 ± 1.88 | 46.15 ± 3.79 | |
| High | 0.35 ± 0.35 | 56.39 ± 1.81 | 18.30 ± 1.28 | 25.91 ± 2.04 | 11.82 ± 1.63 | 26.74 ± 1.89 | 19.01 ± 2.74 | 42.44 ± 3.03 | |
| 2015 | UB | 2.38 ± 2.38 | 36.96 ± 6.19 | 22.67 ± 4.85 | 37.99 ± 3.96 | 11.08 ± 1.97 | 20.19 ± 2.56 | 38.92 ± 4.15 | 29.81 ± 4.79 |
| Low | 0.41 ± 0.41 | 46.34 ± 6.13 | 14.40 ± 1.86 | 32.62 ± 2.27 | 9.52 ± 1.44 | 16.00 ± 2.34 | 33.54 ± 2.53 | 38.71 ± 2.99 | |
| Mid | 0.00 ± 0.00 | 54.88 ± 2.35 | 14.88 ± 1.86 | 30.24 ± 2.70 | 8.54 ± 0.94 | 24.01 ± 1.97 | 27.13 ± 1.73 | 40.33 ± 2.20 | |
| High | 0.82 ± 0.52 | 53.25 ± 2.32 | 19.45 ± 1.89 | 26.49 ± 2.95 | 9.44 ± 1.23 | 24.01 ± 1.29 | 29.72 ± 1.72 | 36.83 ± 2.04 | |
| 2017 | UB | 0.00 ± 0.00 | 46.85 ± 8.59 | 17.60 ± 5.07 | 38.33 ± 9.22 | 11.44 ± 3.45 | 22.66 ± 3.45 | 35.62 ± 7.10 | 30.28 ± 7.52 |
| Low | 0.41 ± 0.41 | 44.08 ± 6.38 | 17.55 ± 2.22 | 33.10 ± 2.89 | 12.97 ± 1.76 | 20.40 ± 2.70 | 28.78 ± 2.33 | 34.38 ± 2.10 | |
| Mid | 0.00 ± 0.00 | 46.38 ± 3.73 | 18.61 ± 2.99 | 35.01 ± 3.09 | 10.49 ± 2.15 | 28.73 ± 1.99 | 23.51 ± 2.49 | 37.28 ± 5.07 | |
| High | 0.34 ± 0.34 | 45.13 ± 1.53 | 20.84 ± 2.29 | 33.69 ± 3.30 | 11.05 ± 1.35 | 26.10 ± 2.10 | 28.08 ± 2.66 | 34.77 ± 3.09 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
González-De Vega, S.; De las Heras, J.; Moya, D. Post-Fire Regeneration and Diversity Response to Burn Severity in Pinus halepensis Mill. Forests. Forests 2018, 9, 299. https://doi.org/10.3390/f9060299
AMA Style
González-De Vega S, De las Heras J, Moya D. Post-Fire Regeneration and Diversity Response to Burn Severity in Pinus halepensis Mill. Forests. Forests. 2018; 9(6):299. https://doi.org/10.3390/f9060299
Chicago/Turabian StyleGonzález-De Vega, Sonsoles, Jorge De las Heras, and Daniel Moya. 2018. "Post-Fire Regeneration and Diversity Response to Burn Severity in Pinus halepensis Mill. Forests" Forests 9, no. 6: 299. https://doi.org/10.3390/f9060299
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.