Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
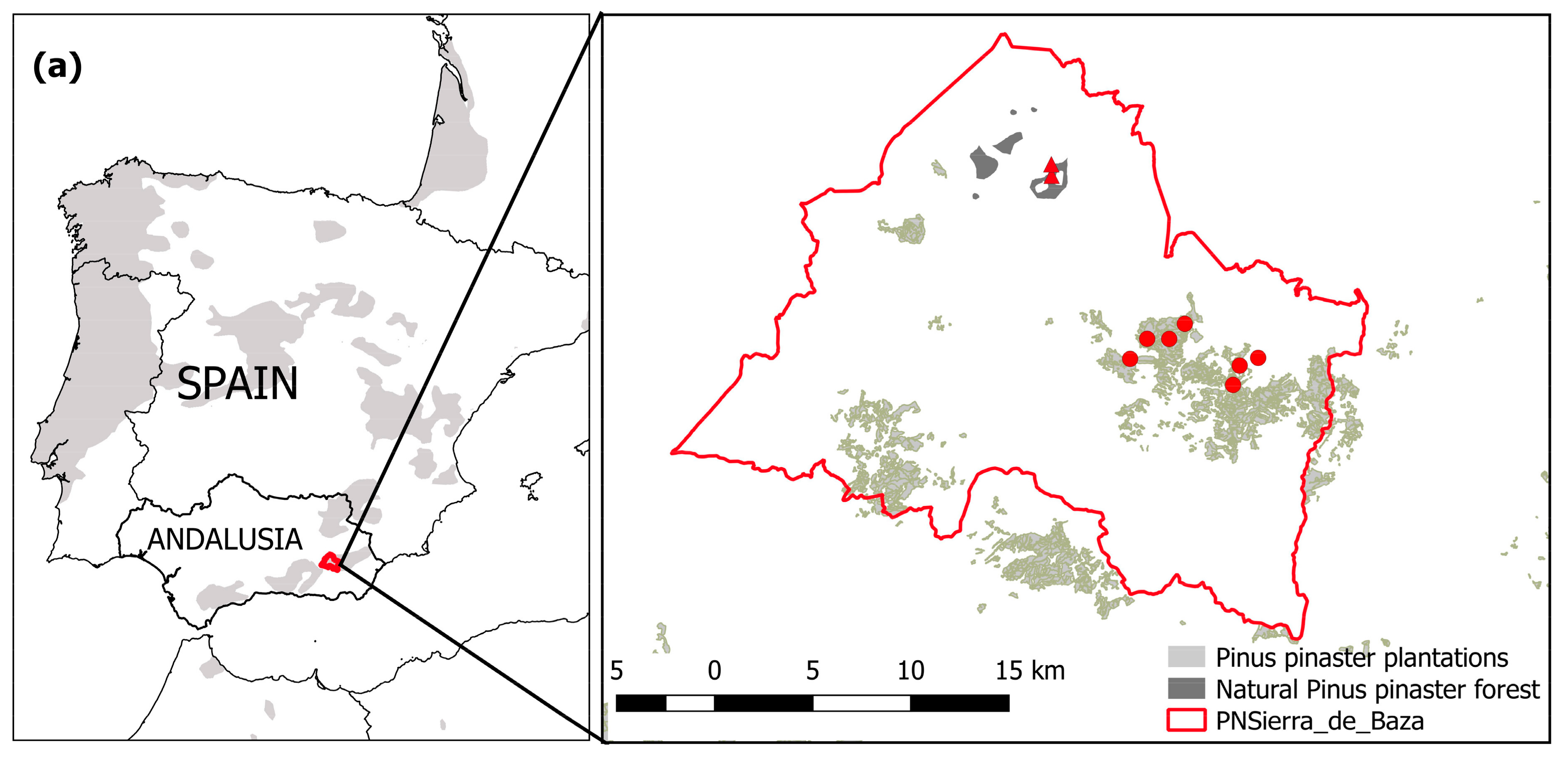
2.1. Study Sites and Tree Species
2.2. Field Sampling
2.3. Tree-Ring Methods
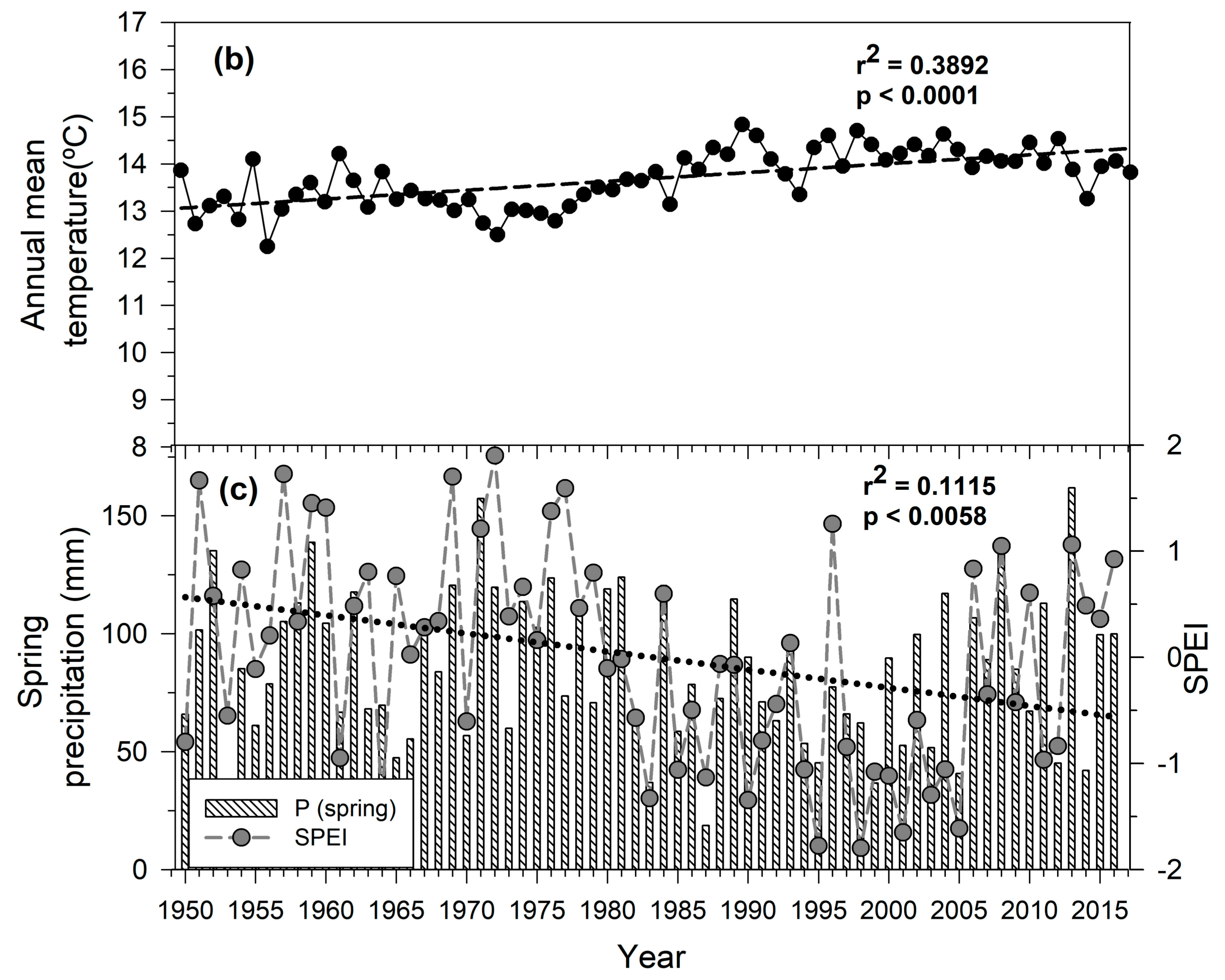
2.4. Climatic Data
2.5. Growth Resilience Indices
- The resistance index (CRT): this index quantifies the growth of the tree during the drought (Dr) with respect to the previous growth (PreDr):(CRT = Dr/PreDr)
- The recovery index (CRS): this is the response of the growth after the drought (PostDr) compared with Dr:(CRS = PostDr/Dr)
- The resilience index (CRC): this is the ratio of the growth values after (PostDr) and before (PreDr) the drought:(CRC = PostDr/PreDr)
2.6. Statistical Analyses
2.7. Environmental and Silvicultural Variables Related to Tree Death
3. Results
3.1. Regional Climatic Variability
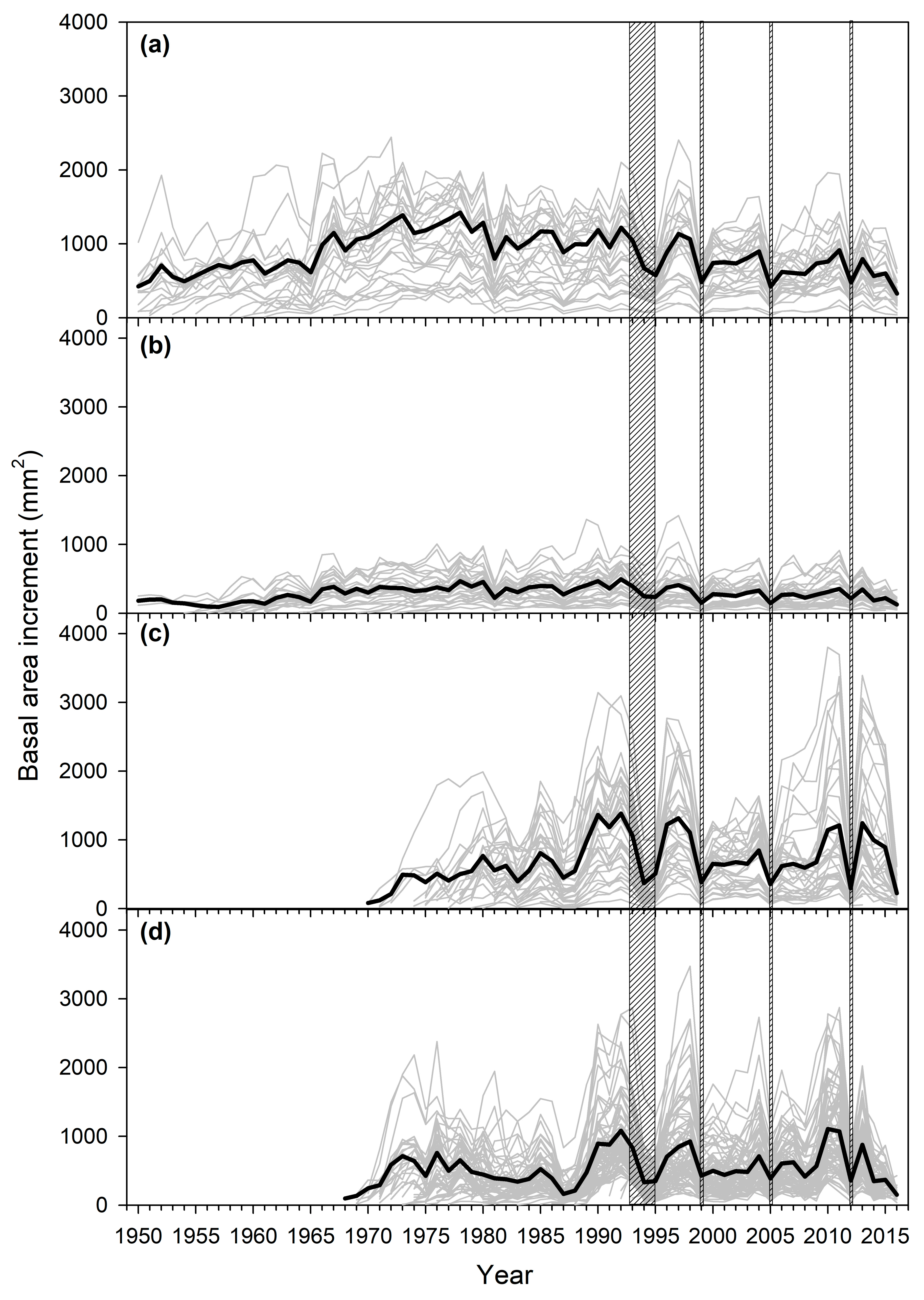
3.2. Mortality and Growth Patterns
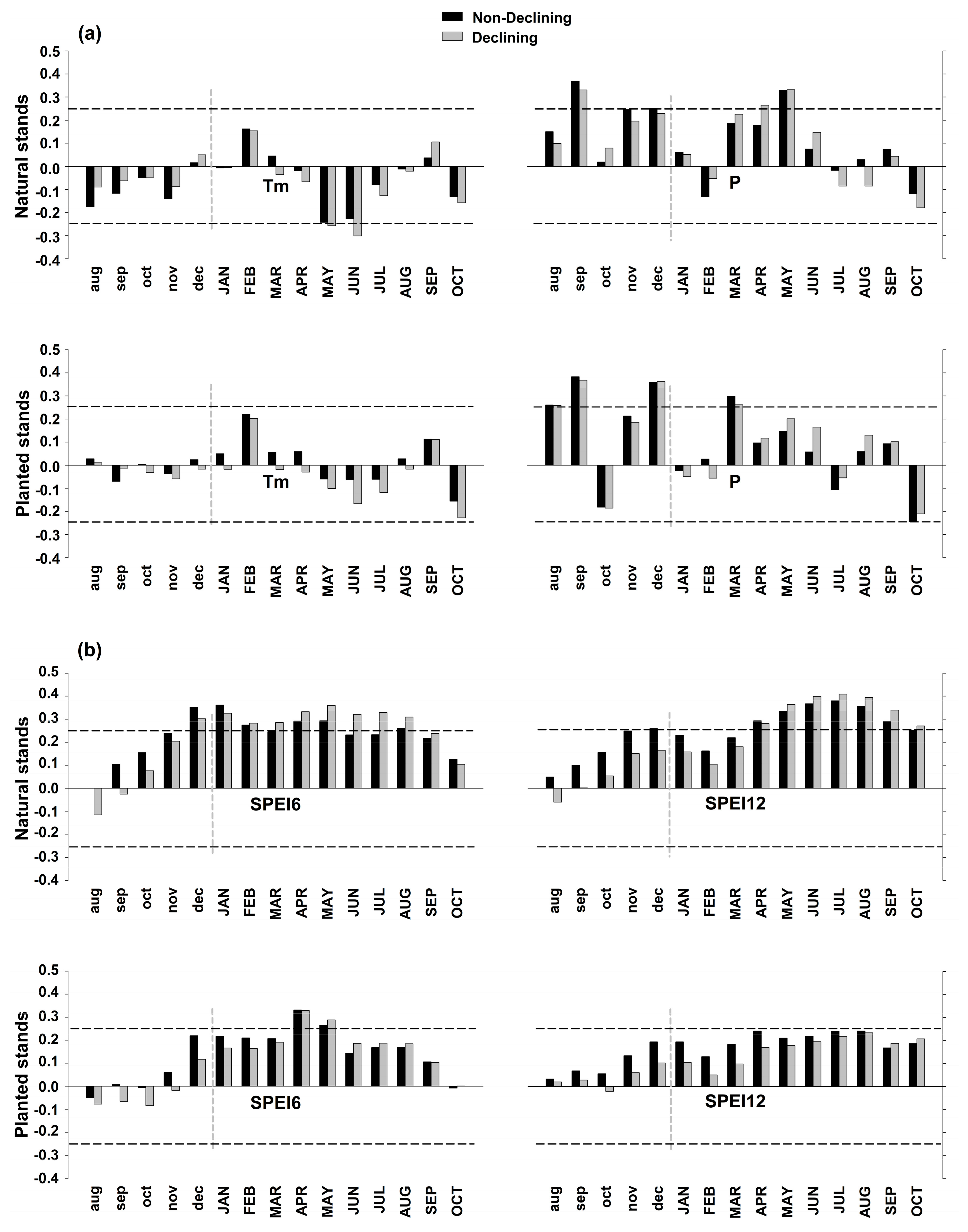
3.3. Climate-Growth Relationships
3.4. Post-Drought Resilience Indices
3.5. Environmental and Silvicultural Drivers of Stand Responses to Drought
4. Discussion
4.1. Climate- and Drought-Growth Relationships in Planted and Naturally Regenerated Stands
4.2. Response of the Resilience Indices of Pine to Droughts
4.3. Responses of Growth and Climatic Indices to the Environment and Stand Structure
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013.
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Swetnam, T.W.; Zavala, M.A. Is drought the main decline factor at the rear edge of Europe? The case of southern Iberian pine plantations. For. Ecol. Manag. 2012, 271, 158–169. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Camarero, J.J.; Fernández-Cancio, Á. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Zhu, J.J.; Fan, Z.P.; Zeng, D.H.; Jiang, F.Q.; Matsuzaki, T. Comparison of stand structure and growth between artificial and naturally regenerated forests of Pinus sylvestiris var. mongolica on sandy land. J. For. Res. 2003, 14, 103–111. [Google Scholar] [CrossRef]
- Song, L.; Li, M.; Zhu, J.; Zhang, J. Comparisons of radial growth and tree-ring cellulose δ13C for Pinus sylvestris var. mongolica in naturally regenerated and plantation forests on sandy lands. J. For. Res. 2017, 22, 160–168. [Google Scholar] [CrossRef]
- FAO Global Forest Resources Assessment 2010; FAO: Rome, Italy, 2010.
- Navarro Cerrillo, R.M.; Varo, M.A.; Lanjeri, S.; Hernandez Clemente, R. Cartografía de defoliación en los pinares de pino silvestre (Pinus sylvestris L.) y pino salgareño (Pinus nigra Arnold.) en la Sierra de los Filabres. Rev. Ecosistemas 2007, 16. [Google Scholar] [CrossRef]
- Dorman, M.; Perevolotsky, A.; Sarris, D.; Svoray, T. The effect of rainfall and competition intensity on forest response to drought: Lessons learned from a dry extreme. Oecologia 2015, 177, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- De la Serrana, R.G.; Vilagrosa, A.; Alloza, J.A. Pine mortality in southeast Spain after an extreme dry and warm year: Interactions among drought stress, carbohydrates and bark beetle attack. Trees 2015, 29, 1791–1804. [Google Scholar] [CrossRef]
- Andreu, L.; Gutiérrez, E.; Macias, M.; Ribas, M.; Bosch, O.; Camarero, J.J. Climate increases regional tree-growth variability in Iberian pine forests. Glob. Chang. Biol. 2007, 13, 804–815. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Christopoulou, A.; Fulé, P.Z.; Andriopoulos, P.; Sarris, D.; Arianoutsou, M. Dendrochronology-based fire history of Pinus nigra forests in Mount Taygetos, Southern Greece. For. Ecol. Manag. 2013, 293, 132–139. [Google Scholar] [CrossRef]
- De Luis, M.; Čufar, K.; Filippo, A.D.; Novak, K.; Papadopoulos, A.; Piovesan, G.; Rathgeber, C.B.K.; Raventós, J.; Saz, M.A.; Smith, K.T. Plasticity in Dendroclimatic Response across the Distribution Range of Aleppo Pine (Pinus halepensis). PLoS ONE 2013, 8, e83550. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Manzanedo, R.D.; Sanchez-Salguero, R.; Navarro-Cerrillo, R.M. Growth response to climate and drought change along an aridity gradient in the southernmost Pinus nigra relict forests. Ann. For. Sci. 2013, 70, 769–780. [Google Scholar] [CrossRef]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sánchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Salguero, R.; Navarro, R.M.; Camarero, J.J.; Fernández-Cancio, Á. Drought-induced growth decline of Aleppo and maritime pine forests in south-eastern Spain. For. Syst. 2010, 19, 458–470. [Google Scholar] [CrossRef]
- Herguido, E.; Granda, E.; Benavides, R.; García-Cervigón, A.I.; Camarero, J.J.; Valladares, F. Contrasting growth and mortality responses to climate warming of two pine species in a continental Mediterranean ecosystem. For. Ecol. Manag. 2016, 363, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Hereş, A.-M.; Martínez-Vilalta, J.; López, B.C. Growth patterns in relation to drought-induced mortality at two Scots pine (Pinus sylvestris L.) sites in NE Iberian Peninsula. Trees 2012, 26, 621–630. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Sterba, H.; del Río, M.; Bravo, F. Competition-induced mortality for Mediterranean Pinus pinaster Ait. and P. sylvestris L. For. Ecol. Manag. 2006, 222, 88–98. [Google Scholar] [CrossRef]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Anderegg, W.R.L.; Kane, J.M.; Anderegg, L.D.L. Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang. 2013, 3, 30–36. [Google Scholar] [CrossRef]
- Galiano, L.; Martínez-Vilalta, J.; Lloret, F. Carbon reserves and canopy defoliation determine the recovery of Scots pine 4 yr after a drought episode. New Phytol. 2011, 190, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderegg, W.R.L.; Berry, J.A.; Field, C.B. Linking definitions, mechanisms, and modeling of drought-induced tree death. Trends Plant Sci. 2012, 17, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Caminero, L.; Génova, M.; Camarero, J.J.; Sánchez-Salguero, R. Growth responses to climate and drought at the southernmost European limit of Mediterranean Pinus pinaster forests. Dendrochronologia 2018, 48, 20–29. [Google Scholar] [CrossRef]
- Corcuera, L.; Gil-Pelegrín, E.; Notivol, E. Differences in hydraulic architecture between mesic and xeric Pinus pinaster populations at the seedling stage. Tree Physiol. 2012, 32, 1442–1457. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Queyrens, A.; Costa, P.; Loustau, D.; Plomion, C. Osmotic adjustment in Pinus pinaster cuttings in response to a soil drying cycle. Ann. For. Sci. 2002, 59, 795–799. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Bogino, S.M.; Bravo, F. Growth response of Pinus pinaster Ait. to climatic variables in central Spanish forests. Ann. For. Sci. 2008, 65, 506. [Google Scholar] [CrossRef]
- Smiley, T. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AR, USA, 1968. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating Crossdating Accuracy: A Manual and Tutorial for the Computer Program COFECHA. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 2001. [Google Scholar]
- Haylock, M.R.; Hofstra, N.; Klein-Tank, A.; Klok, E.J.; Jones, P.D.; New, M. A European daily high-resolution gridded data set of surface temperature and precipitation for 1950–2006. J. Geophys. Res. Atmos. 2008, 113, D20. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2009, 23, 1696–1718. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, K.; Eckstein, D.; Holmes, R.L. The Dendrochronological Signal of Pine Trees (Pinus spp.) in Spain. Tree-Ring Bull. 1991, 51, 1–13. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics; W.H. Free Company: New York, NY, USA, 1995. [Google Scholar]
- Zang, C.; Biondi, F. Dendroclimatic calibration in R: The bootRes package for response and correlation function analysis. Dendrochronologia 2013, 31, 68–74. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Gandullo, J.M.; Sánchez Palomares, O. Estaciones Ecológicas de los Pinares Españoles; ICONA: Madrid, Spain, 1994. [Google Scholar]
- Chung, D.; Chun, H.; Keles, S. An Introduction to the ‘spls’ Package; Version 1.0; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Hair, J.F.; Ringle, C.M.; Sarstedt, M. Editorial-Partial Least Squares: The Better Approach to Structural Equation Modeling? Long Range Plan. 2012, 45, 312–319. [Google Scholar] [CrossRef]
- Adams, H.D.; Guardiola-Claramonte, M.; Barron-Gafford, G.A.; Villegas, J.C.; Breshears, D.D.; Zou, C.B.; Troch, P.A.; Huxman, T.E. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. USA 2009, 106, 7063–7066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, J.; Campelo, F.; Nabais, C. Age-dependent responses of tree-ring growth and intra-annual density fluctuations of Pinus pinaster to Mediterranean climate. Trees 2009, 23, 257–265. [Google Scholar] [CrossRef]
- Rozas, V.; García-González, I.; Zas, R. Climatic control of intra-annual wood density fluctuations of Pinus pinaster in NW Spain. Trees 2011, 25, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Candel-Pérez, D.; Linares, J.C.; Viñegla, B.; Lucas-Borja, M.E. Assessing climate–growth relationships under contrasting stands of co-occurring Iberian pines along an altitudinal gradient. For. Ecol. Manag. 2012, 274, 48–57. [Google Scholar] [CrossRef]
- Campelo, F.; Vieira, J.; Nabais, C. Tree-ring growth and intra-annual density fluctuations of Pinus pinaster responses to climate: Does size matter? Trees 2013, 27, 763–772. [Google Scholar] [CrossRef]
- Génova, M.; Caminero, L.; Dochao, J. Resin tapping in Pinus pinaster: Effects on growth and response function to climate. Eur. J. For. Res. 2014, 133, 323–333. [Google Scholar] [CrossRef]
- Kurz-Besson, C.B.; Lousada, J.L.; Gaspar, M.J.; Correia, I.E.; David, T.S.; Soares, P.M.M.; Cardoso, R.M.; Russo, A.; Varino, F.; Mériaux, C.; et al. Effects of Recent Minimum Temperature and Water Deficit Increases on Pinus pinaster Radial Growth and Wood Density in Southern Portugal. Front. Plant Sci. 2016, 7, 1170. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Sinoga, J.D.; Garcia Marin, R.; Martinez Murillo, J.F.; Gabarron Galeote, M.A. Precipitation dynamics in southern Spain: trends and cycles. Int. J. Climatol. 2011, 31, 2281–2289. [Google Scholar] [CrossRef]
- Spinoni, J.; Naumann, G.; Vogt, J.V.; Barbosa, P. The biggest drought events in Europe from 1950 to 2012. J. Hydrol. Reg. Stud. 2015, 3, 509–524. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Dobbertin, M.; Fernández-Cancio, Á.; Vilà-Cabrera, A.; Manzanedo, R.D.; Zavala, M.A.; Navarro-Cerrillo, R.M. Contrasting vulnerability and resilience to drought-induced decline of densely planted vs. naturally regenerated rear-edge Pinus nigra forests. For. Ecol. Manag. 2013, 310, 956–967. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Viguera, B.; Cabrera, M.; Cañellas, I. Drought induced decline could portend widespread pine mortality at the xeric ecotone in managed mediterranean pine-oak woodlands. For. Ecol. Manag. 2014, 320, 70–82. [Google Scholar] [CrossRef]
- Tyree, M.T.; Davis, S.D.; Cochard, H. Biophysical Perspectives of Xylem Evolution: Is there a Tradeoff of Hydraulic Efficiency for Vulnerability to Dysfunction? IAWA J. 1994, 15, 335–360. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; López, B.C.; Loepfe, L.; Lloret, F. Stand- and tree-level determinants of the drought response of Scots pine radial growth. Oecologia 2012, 168, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Correia, I.; Almeida, M.H.; Aguiar, A.; Alía, R.; David, T.S.; Pereira, J.S. Variations in growth, survival and carbon isotope composition (δ13C) among Pinus pinaster populations of different geographic origins. Tree Physiol. 2008, 28, 1545–1552. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Recio, C.; Martín-García, J.; Bravo, F.; Diez, J.J. Unravelling the associations between climate, soil properties and forest management in Pinus pinaster decline in the Iberian Peninsula. For. Ecol. Manag. 2015, 356, 74–83. [Google Scholar] [CrossRef]
- Di Matteo, G.; Voltas, J. Multienvironment Evaluation of Pinus pinaster Provenances: Evidence of Genetic Trade-Offs between Adaptation to Optimal Conditions and Resistance to the Maritime Pine Bast Scale (Matsucoccus feytaudi). For. Sci. 2016, 62, 553–563. [Google Scholar] [CrossRef]
- Shanahan, E.; Irvine, K.M.; Thoma, D.; Wilmoth, S.; Ray, A.; Legg, K.; Shovic, H. Whitebark pine mortality related to white pine blister rust, mountain pine beetle outbreak, and water availability. Ecosphere 2016, 7, e01610. [Google Scholar] [CrossRef]
- Löf, M.; Dey, D.C.; Navarro, R.M.; Jacobs, D.F. Mechanical site preparation for forest restoration. New For. 2012, 43, 825–848. [Google Scholar] [CrossRef]
- Rueda, B.; Rodriguez-Alleres, M.; Varela Tejero, E. Environmental Factors Governing Soil Water Repellency Dynamics in a Pinus Pinaster Plantation in NW Spain. Land Degrad. Dev. 2015, 27, 719–728. [Google Scholar] [CrossRef]
- Cerrillo, R.M.N.; Rumbaó, I.C.; Vidaña, A.L.; Pérez, J.L.Q.; Duque-Lazo, J. Integración de datos de inventario y modelos de hábitat para predecir la regeneración de especies leñosas mediterráneas en repoblaciones forestales. Rev. Ecosistemas 2016, 25, 6–21. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
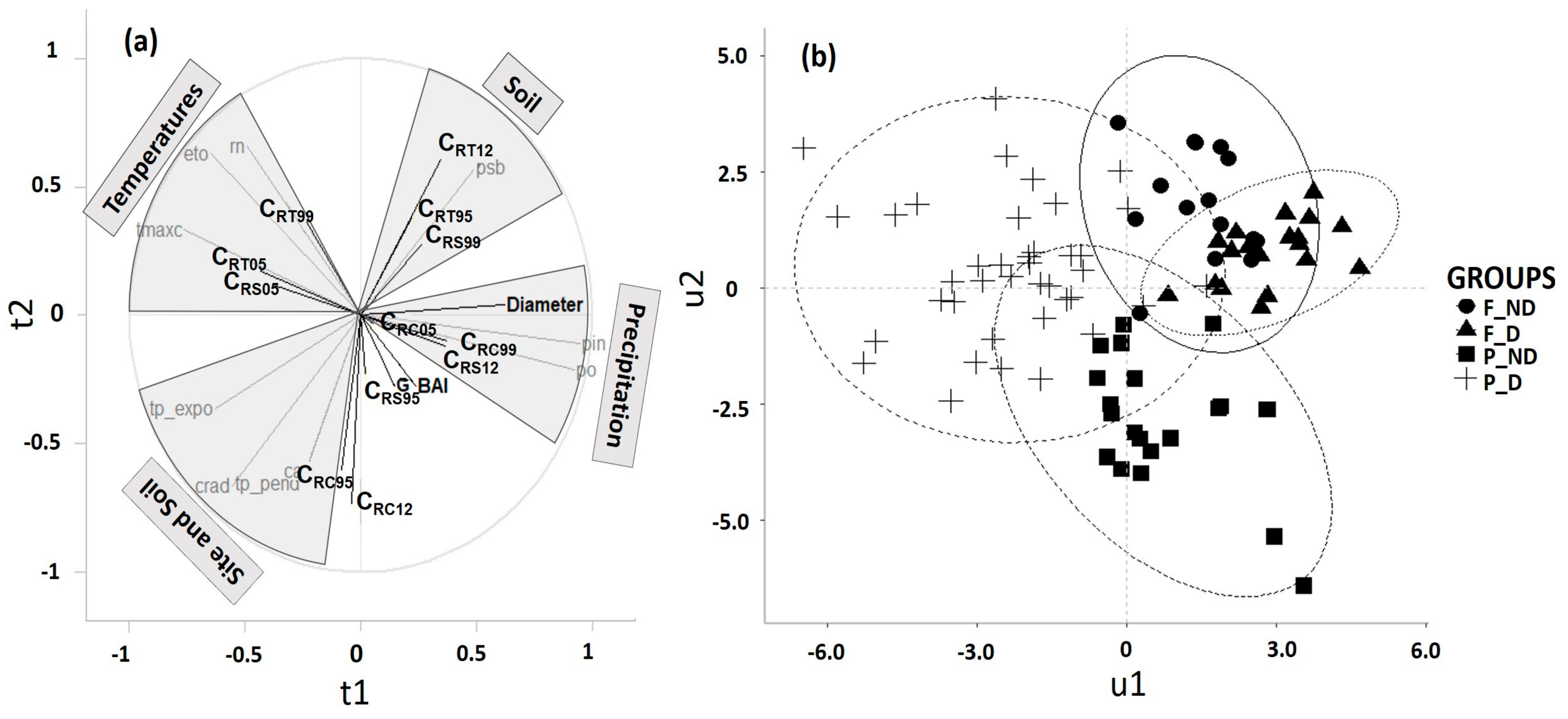{kind=link}
| Stand Type | Tree Vigor Class | Site Code | Elevation (m) | Slope (%) | Basal Area (m2 ha−1) | Diameter at 1.3 m (cm) | Height (m) |
|---|---|---|---|---|---|---|---|
| Forest (naturally regenerated stand) | Non-declining trees | F-ND | 1430 | 30–40 | 22.5 | 29.7 ± 0.6 | 8.3 ± 0.9 |
| Declining trees | F-D | 1300 | 20–30 | 19.5 | 22.9 ± 0.8 | 6.4 ± 0.4 | |
| Planted stand | Non-declining trees | P-ND | 1380 | 20–30 | 24.0 | 24.07 ± 0.7 | 7.9 ± 0.3 |
| P-ND | 1505 | 20-30 | 22.5 | 23.76 ± 0.6 | 8.3± 0.3 | ||
| Declining trees | P-D | 1286 | 10–20 | 21.0 | 22.84± 0.5 | 6.2 ± 0.2 | |
| P-D | 1340 | 10–20 | 23.5 | 20.78± 0.6 | 6.4 ± 0.2 | ||
| P-D | 1338 | 20-30 | 22.0 | 21.01± 0.6 | 8.8 ± 0.3 | ||
| P-D | 1256 | 10–20 | 22.5 | 24.34± 0.7 | 7.7 ± 0.3 | ||
| P-D | 1231 | 20-30 | 20.0 | 24.07± 0.6 | 7.1 ± 0.3 |
| Stand Type | Vigor Class | Code | No. Trees (No. Cores) | Age at 1.3 m (years) | Timespan | TRW (mm) | BAI20 (mm2) | MS | AC1 |
|---|---|---|---|---|---|---|---|---|---|
| Naturally regenerated | ND | F-ND | 18 (36) | 62 ± 3 | 1941–2016 | 2.04 ± 0.19 b | 702 ± 46 b | 0.27 ± 0.01 | 0.81 ± 0.01 |
| D | F-D | 18 (30) | 57 ± 2 | 1930–2016 | 1.26 ± 0.11 a | 262 ± 17 a | 0.30 ± 0.01 | 0.76 ± 0.02 | |
| Planted | ND | P-ND | 20 (39) | 39 ± 1 | 1970–2016 | 2.43 ± 0.30 | 753 ± 72 b | 0.44 ± 0.01 | 0.69 ± 0.01 |
| D | P-D | 20 (40) | 37 ± 1 | 1969–2016 | 2.28 ± 0.20 | 578 ± 61 a | 0.43 ± 0.01 | 0.64 ± 0.02 |
| Factor | 1995 Drought | 1999 Drought | 2005 Drought | 2012 Drought | BAI20 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CRT | CRC | CRS | CRT | CRC | CRS | CRT | CRC | CRS | CRT | CRC | CRS | ||
| Stand type (S) | 24.070 | 30.06 | 0.28 | 0.20 | 15.64 | 15.48 | 6.54 | 0.15 | 3.39 | 112.11 | 40.18 | 8.05 | 0.56 |
| <0.001 | <0.01 | 0.60 | 0.63 | <0.01 | <0.01 | 0.03 | 0.69 | 0.07 | <0.01 | <0.01 | 0.01 | 0.46 | |
| Decline (D) | 0.54 | 14.26 | 2.62 | 9.39 | 8.01 | 0.44 | 11.15 | 0.02 | 8.51 | 14.02 | 21.57 | 2.87 | 101.43 |
| 0.44 | <0.01 | 0.11 | 0.01 | 0.01 | 0.51 | 0.01 | 0.87 | 0.01 | <0.01 | <0.01 | 0.09 | <0.01 | |
| S*D | 3.70 | 7.18 | 0.02 | 20.83 | 36.99 | 0.02 | 9.45 | 1.12 | 2.13 | 0.01 | 14.02 | 8.24 | 36.81 |
| 0.06 | 0.01 | 0.97 | <0.01 | <0.01 | 0.89 | 0.02 | 0.29 | 0.15 | 0.91 | <0.01 | 0.01 | <0.01 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-Cerrillo, R.M.; Rodriguez-Vallejo, C.; Silveiro, E.; Hortal, A.; Palacios-Rodríguez, G.; Duque-Lazo, J.; Camarero, J.J. Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands. Forests 2018, 9, 358. https://doi.org/10.3390/f9060358
Navarro-Cerrillo RM, Rodriguez-Vallejo C, Silveiro E, Hortal A, Palacios-Rodríguez G, Duque-Lazo J, Camarero JJ. Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands. Forests. 2018; 9(6):358. https://doi.org/10.3390/f9060358
Chicago/Turabian StyleNavarro-Cerrillo, Rafael M., Carlos Rodriguez-Vallejo, Emidio Silveiro, Antonio Hortal, Guillermo Palacios-Rodríguez, Joaquín Duque-Lazo, and J. Julio Camarero. 2018. "Cumulative Drought Stress Leads to a Loss of Growth Resilience and Explains Higher Mortality in Planted than in Naturally Regenerated Pinus pinaster Stands" Forests 9, no. 6: 358. https://doi.org/10.3390/f9060358