Plant-Soil Properties Associated with Nitrogen Mineralization: Effect of Conversion of Natural Secondary Forests to Larch Plantations in a Headwater Catchment in Northeast China

Abstract
:1. Introduction
2. Materials and Methods
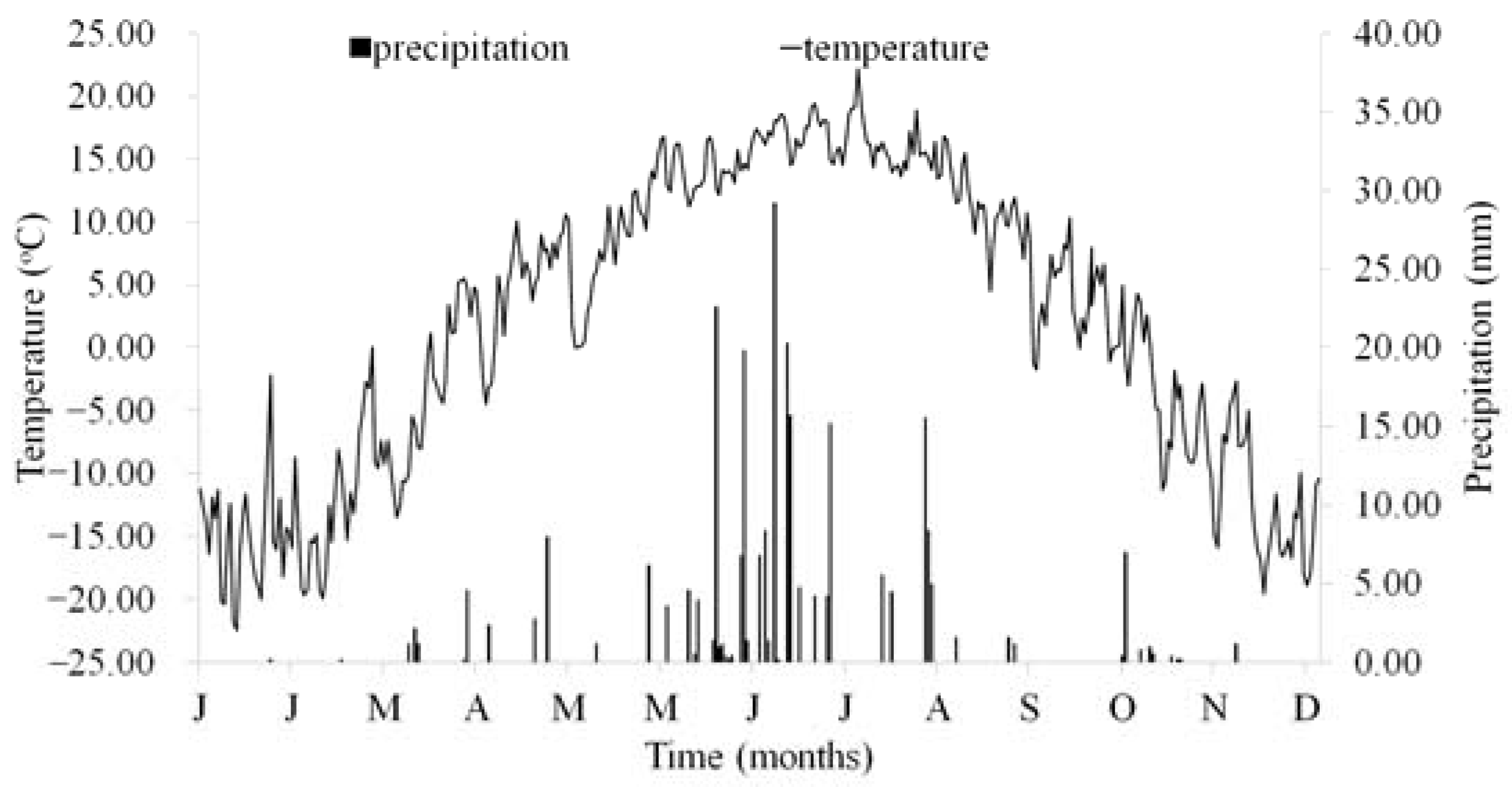
2.1. Study Area
2.2. Variables Assessed
2.2.1. Vegetation Survey
2.2.2. Soil Sampling and Incubation
2.2.3. Soil Chemical Properties
2.3. Statistical Analysis
3. Results
3.1. Seasonal Soil Mineral N
3.2. Net N Mineralization of Organic Matter
3.3. Plant and Soil Properties
3.4. Relationship of Soil Organic N Mineralization, Ammonification and Nitrification to Climate, Plant and Soil Variables
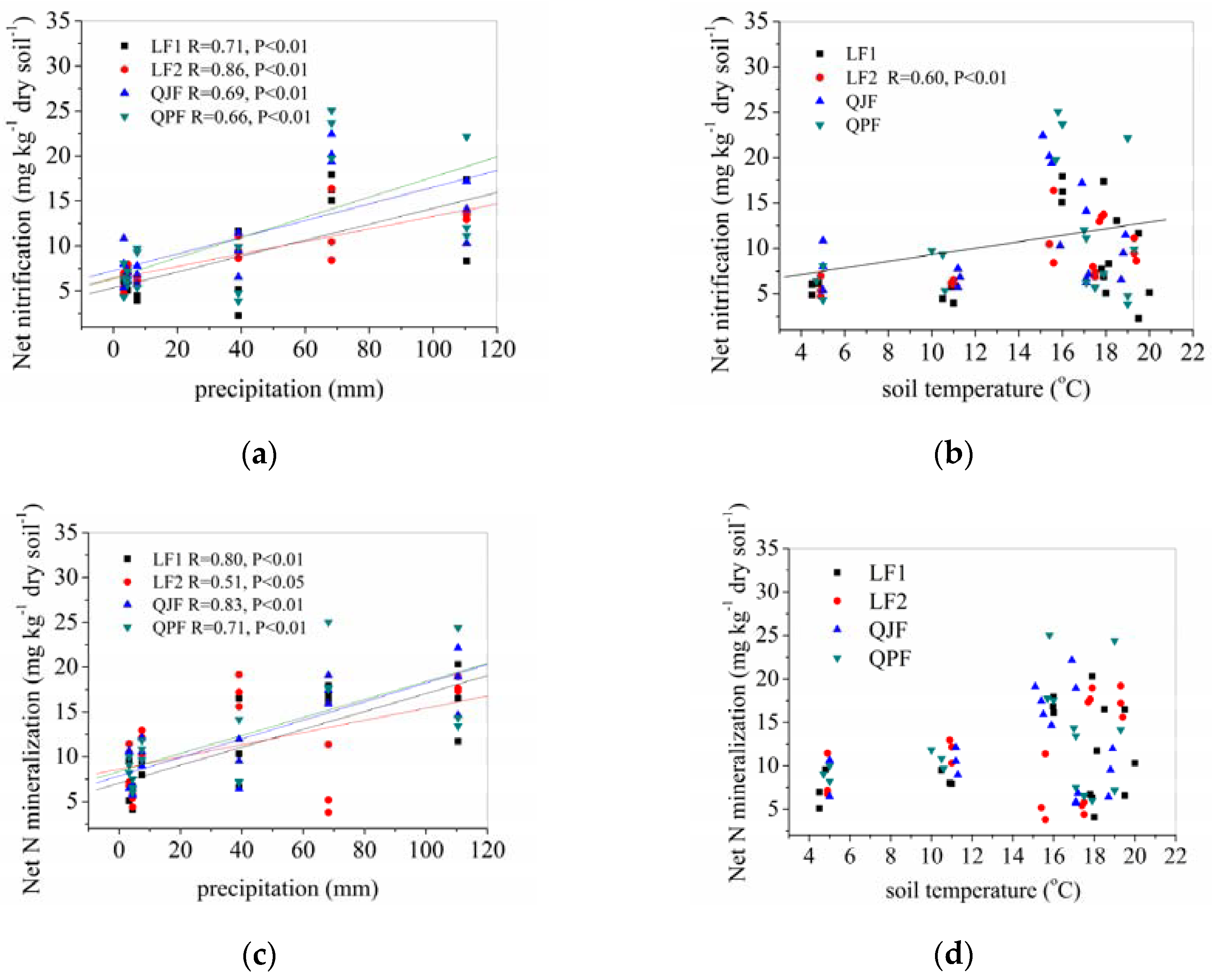
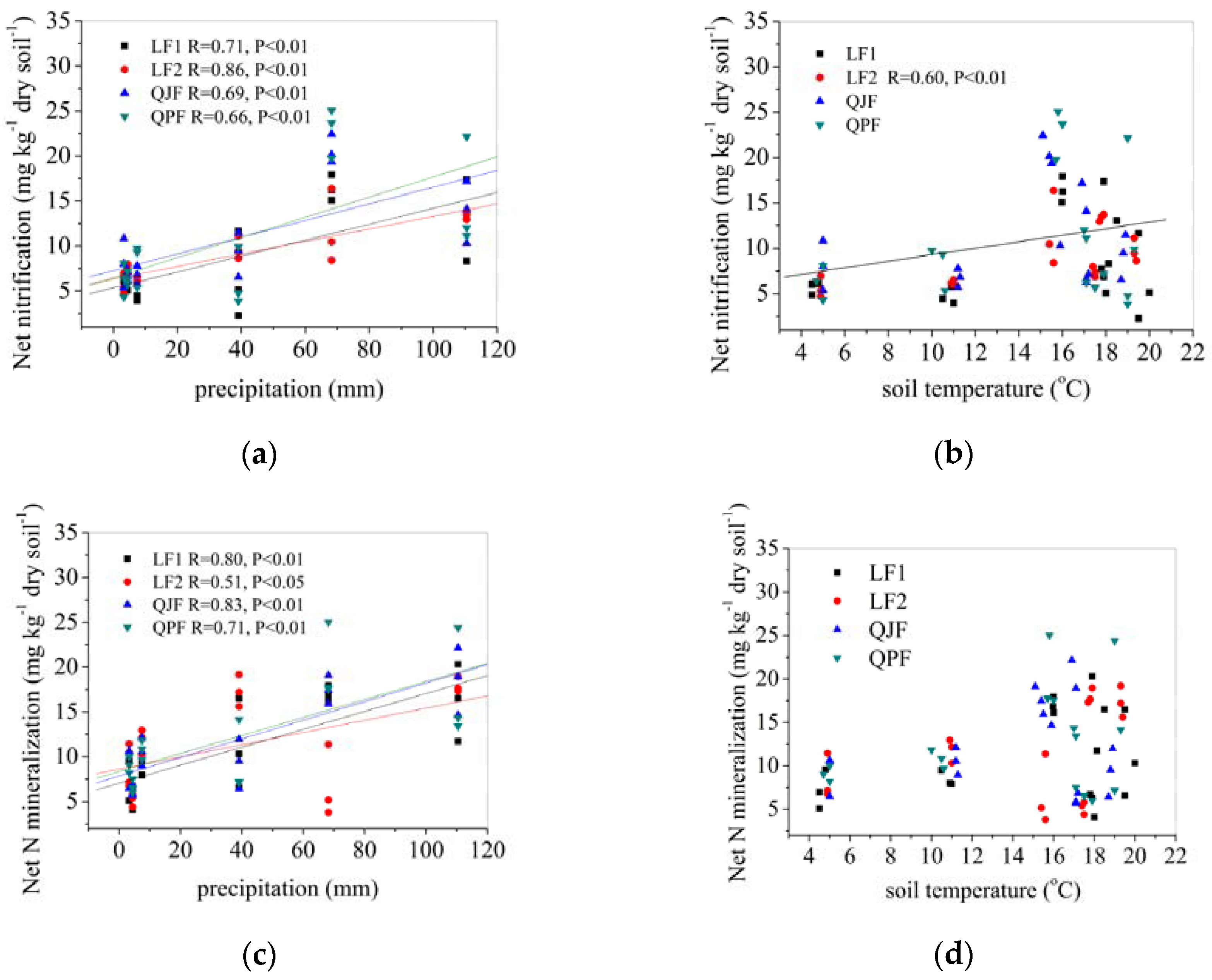
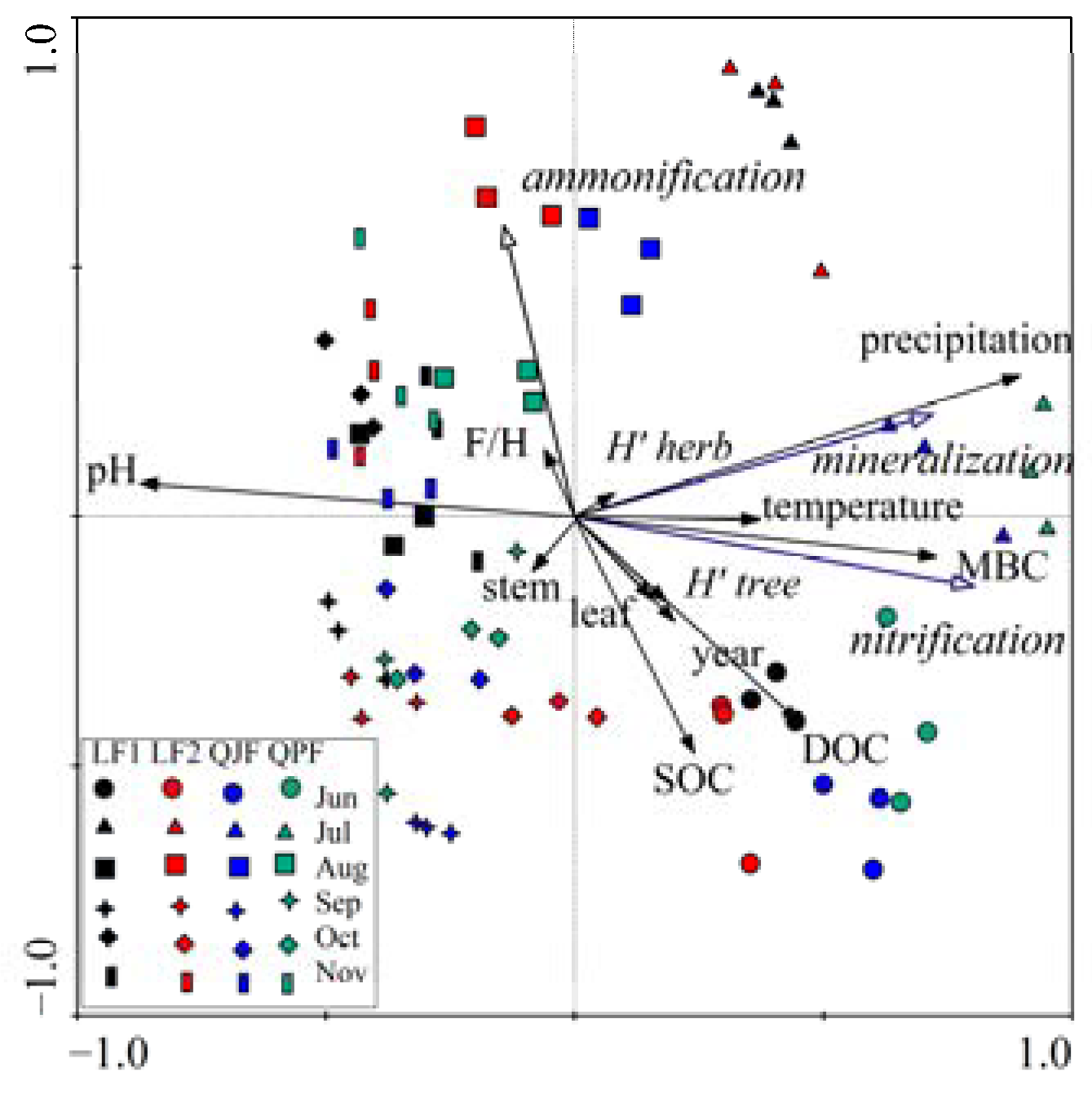
3.4.1. Precipitation and Temperature Effects on Net Organic N Mineralization and Nitrification Rates
3.4.2. Correlations between Soil Properties and Organic N Mineralization, Ammonification and Nitrification
3.4.3. Relationship between Vegetation Parameters and N Mineralization of Organic Matter
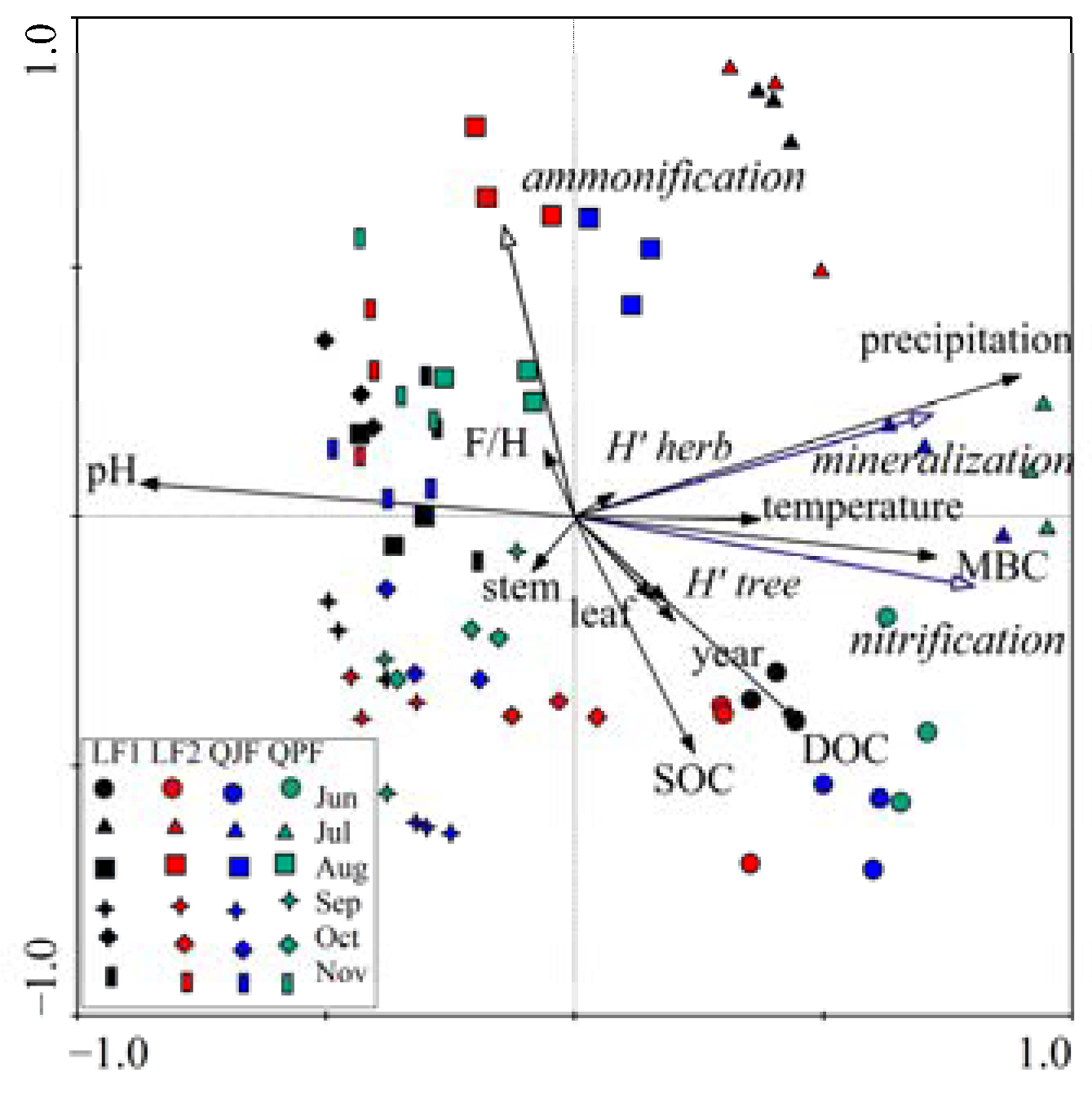
3.5. Multivariate Analysis (Redundancy Analysis)
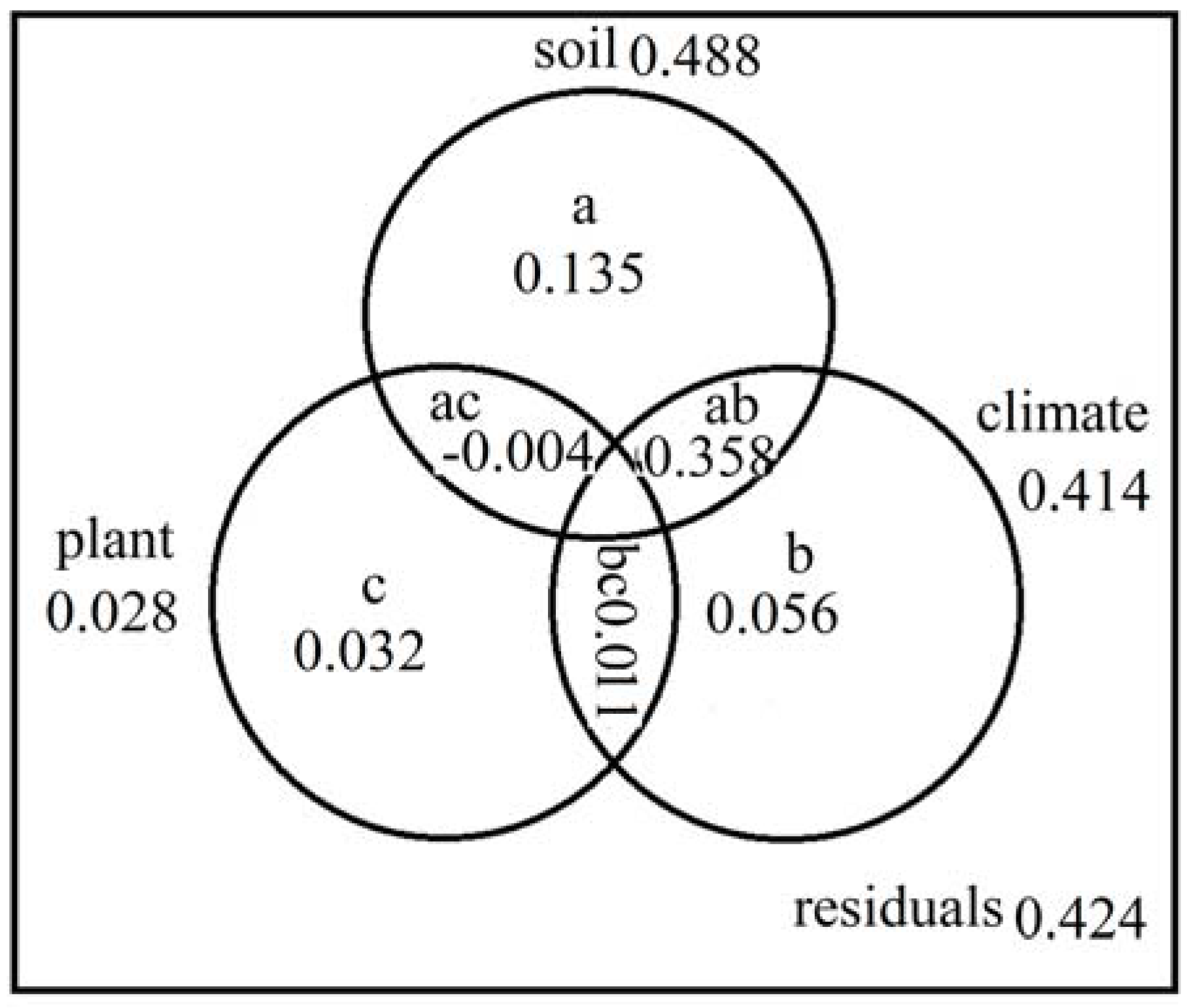
3.6. Variation Partitioning
4. Discussion
4.1. The Effects of Plant on N Mineralization
4.2. The Effects of Soil Properties on N Mineralization
4.3. Seasonality Effects on Soil N Transformation Patterns
4.4. Interaction of Climate, Plant and Soil Variables on N Transformation
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pampolino, M.F.; Urushiyama, T.; Hatano, R. Detection of nitrate leaching through bypass flow using pan lysimeter, suction cup, and resin capsule. Soil Sci.Plant Nutr. 2000, 46, 703–711. [Google Scholar] [CrossRef] [Green Version]
- Perakis, S.; Hedin, L. Nitrogen loss from unpolluted south american forests mainly via dissolved organic compounds. Nature 2002, 415, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Pandey, C.B.; Singh, G.B.; Singh, S.K.; Singh, R.K. Soil nitrogen and microbial biomass carbon dynamics in native forests and derived agricultural land uses in a humid tropical climate of india. Plant Soil 2010, 333, 453–467. [Google Scholar] [CrossRef]
- Lupon, A.; Gerber, S.; Sabater, F.; Bernal, S. Climate response of the soil nitrogen cycle in three forest types of a headwater mediterranean catchment: Climate response of soil nitrogen cycle. J. Geophys. Res. Biogeosci. 2015, 120, 2988–2999. [Google Scholar] [CrossRef]
- Kelly, C.N.; Schoenholtz, S.H.; Adams, M.B. Soil properties associated with net nitrification following watershed conversion from appalachian hardwoods to norway spruce. Plant Soil 2011, 344, 361–376. [Google Scholar] [CrossRef]
- Noe, G.B.; Hupp, C.R.; Rybicki, N.B. Hydrogeomorphology influences soil nitrogen and phosphorus mineralization in floodplain wetlands. Ecosystems 2013, 16, 75–94. [Google Scholar] [CrossRef]
- Chen, J.; Xiao, G.; Kuzyakov, Y.; Darrel Jenerette, G.; Ma, Y.; Liu, W.; Wang, Z.; Shen, W. Soil nitrogen transformation responses to seasonal precipitation changes are regulated by changes in functional microbial abundance in a subtropical forest. Biogeosciences 2017, 14, 2513–2525. [Google Scholar] [CrossRef]
- Wang, C.; Wan, S.; Xing, X.; Zhang, L.; Han, X. Temperature and soil moisture interactively affected soil net n mineralization in temperate grassland in northern china. Soil Biol. Biochem. 2006, 38, 1101–1110. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Huang, J. Net nitrogen mineralization and nitrification in three subtropical forests of southwestern china. Dyn. Soil Dyn. Plant 2008, 2, 33–40. [Google Scholar]
- Liu, X.R.; Dong, Y.S.; Ren, J.Q.; Li, S.G. Drivers of soil net nitrogen mineralization in the temperate grasslands in inner mongolia, china. Nutr. Cycl. Agroecosyst. 2010, 87, 59–69. [Google Scholar] [CrossRef]
- Hishi, T.; Urakawa, R.; Tashiro, N.; Maeda, Y.; Shibata, H. Seasonality of factors controlling n mineralization rates among slope positions and aspects in cool-temperate deciduous natural forests and larch plantations. Biol. Fertil. Soils 2014, 50, 343–356. [Google Scholar] [CrossRef]
- Mylliemngap, W.; Nath, D.; Barik, S.K. Changes in vegetation and nitrogen mineralization during recovery of a montane subtropical broadleaved forest in north-eastern india following anthropogenic disturbance. Ecol. Res. 2016, 31, 21–38. [Google Scholar] [CrossRef]
- Chu, H.; Grogan, P. Soil microbial biomass, nutrient availability and nitrogen mineralization potential among vegetation-types in a low arctic tundra landscape. Plant Soil 2010, 329, 411–420. [Google Scholar] [CrossRef]
- Trum, F.; Titeux, H.; Ranger, J.; Delvaux, B. Influence of tree species on carbon and nitrogen transformation patterns in forest floor profiles. Ann. For. Sci. 2011, 68, 837–847. [Google Scholar] [CrossRef]
- Xiong, Y.; Zeng, H.; Xia, H.; Guo, D. Interactions between leaf litter and soil organic matter on carbon and nitrogen mineralization in six forest litter-soil systems. Plant Soil 2014, 379, 217–229. [Google Scholar] [CrossRef]
- Yang, K.; Shi, W.; Zhu, J.J. The impact of secondary forests conversion into larch plantations on soil chemical and microbiological properties. Plant Soil 2013, 368, 535–546. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Khanna, P.K.; Fortmann, H.; Meesenburg, H.; Eichhorn, J.; Meiwes, K.J. Biomass and element content of foliage and aboveground litterfall on the three long-term experimental beech sites: Dynamics and significance. In Functioning and Management of European Beech Ecosystems; Brumme, R., Khanna, P.K., Eds.; Springer (Berlin Heidelberg): Berlin, Germany, 2009; pp. 183–205. [Google Scholar]
- Denton, C.S.; Bardgett, R.D.; Cook, R.; Hobbs, P.J. Low amounts of root herbivory positively influence the rhizosphere microbial community in a temperate grassland soil. Soil Biol. Biochem. 1999, 31, 155–165. [Google Scholar] [CrossRef]
- Mikola, J.; Barker, G.; Wardle, D. Linking above-ground and below-ground effects in autotrophic microcosms: Effects of shading and defoliation on plant and soil properties. Oikos 2000, 89, 577–587. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Hart, S.C.; Kaye, J.P.; Moore, M.M. Evidence for indirect effects of plant diversity and composition on net nitrification. Plant Soil 2010, 330, 435–445. [Google Scholar] [CrossRef]
- Alarcón-Gutiérrez, E.; Floch, C.; Ziarelli, F.; Albrecht, R.; Le Petit, J.; Augur, C.; Criquet, S. Characterization of a mediterranean litter by ¹³c cpmas nmr: Relationships between litter depth, enzyme activities and temperature. Eur. J. Soil Sci. 2008, 59, 486–495. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; Volume 27, p. 177. [Google Scholar]
- Margalef, R. Temporal Succession and Spatial Heterogeneity in Phytoplankton; University of California Press: Berkeley, CA, USA, 1958; pp. 323–349. [Google Scholar]
- Wang, C. Biomass allometric equations for 10 co-occurring tree species in chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Bhogal, A.; Hatch, D.J.; Shepherd, M.A.; Jarvis, S.C. Comparison of methodologies for field measurement of net nitrogen mineralisation in arable soils. Plant Soil 1999, 207, 15–28. [Google Scholar] [CrossRef]
- Hatch, D.J.; Jarvis, S.C.; Parkinson, R.J.; Lovell, R.D. Combining field incubation with nitrogen-15 labelling to examine nitrogen transformations in low to high intensity grassland management systems. Biol. Fertil. Soils 2000, 30, 492–499. [Google Scholar] [CrossRef]
- Hatch, D.J.; Jarvis, S.C.; Parkinson, R.J. Concurrent measurements of net mineralization, nitrification, denitrification and leaching from field incubated soil cores. Biol. Fertil. Soils 1998, 26, 323–330. [Google Scholar] [CrossRef]
- Carranca, C.; Oliveira, A.; Pampulha, E.; Torres, M.O. Temporal dynamics of soil nitrogen, carbon and microbial activity in conservative and disturbed fields amended with mature white lupine and oat residues. Geoderma 2009, 151, 50–59. [Google Scholar] [CrossRef]
- Raison, R.J.; Connell, M.J.; Khanna, P.K. Methodology for studying fluxes of soil mineral-n in situ. Soil Biol. Biochem. 1987, 19, 521–530. [Google Scholar] [CrossRef]
- Hübner, C.; Redl, G.; Wurst, F. In situ methodology for studying n-mineralization in soils using anion exchange resins. Soil Biol. Biochem. 1991, 23, 701–702. [Google Scholar] [CrossRef]
- Valenzuela-Solano, C.; Crohn, D.M.; Downer, J.A. Nitrogen mineralization from eucalyptus yardwaste mulch applied to young avocado trees. Biol. Fertil. Soils 2005, 41, 38–45. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass c. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Cabrera, M.L. Alkaline persulfate oxidation for determining total nitrogen in microbial biomass extracts. Soil Sci. Soc. Am. J. 1993, 57, 1007–1012. [Google Scholar] [CrossRef]
- Zeglin, L.H.; Stursova, M.; Sinsabaugh, R.L.; Collins, S.L. Microbial responses to nitrogen addition in three contrasting grassland ecosystems. Oecologia 2007, 154, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using Canoco; Cambridge University Press: Cambridge, UK, 2003; p. 269. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef]
- Scherer-Lorenzen, M.; Luis Bonilla, J.; Potvin, C. Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 2007, 116, 2108–2124. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Zhang, Y.; Chen, C.; Xu, Z.; Guo, X. Plant–soil interaction affects the mineralization of soil organic carbon: Evidence from 73-year-old plantations with three coniferous tree species in subtropical Australia. J. Soils Sediments 2017, 17, 985–995. [Google Scholar] [CrossRef]
- Fukushima, K.; Tateno, R.; Tokuchi, N. Soil nitrogen dynamics during stand development after clear-cutting of japanese cedar (cryptomeria japonica) plantations. J. For. Res. 2011, 16, 394. [Google Scholar] [CrossRef] [Green Version]
- Burns, D.A.; Murdoch, P.S. Effects of a clearcut on the net rates of nitrification and n mineralization in a northern hardwood forest, catskill mountains, new york, USA. Biogeochemistry 2005, 72, 123–146. [Google Scholar] [CrossRef]
- Koutika, LS.; Epron, D.; Bouillet, J.P.; Mareschal, L. Changes in n and c concentrations, soil acidity and p availability in tropical mixed acacia and eucalypt plantations on a nutrient-poor sandy soil. Plant Soil 2014, 379, 205–216. [Google Scholar] [CrossRef]
- Mulder, C.; Schouten, A.J.; Hund-Rinke, K.; Breure, A.M. The use of nematodes in ecological soil classification and assessment concepts. Ecotoxicol. Environ. Saf. 2005, 62, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Niklaus, P.A.; Kandeler, E.; Leadley, P.W.; Schmid, B.; Tscherko, D.; Körner, C. A link between plant diversity, elevated co2 and soil nitrate. Oecologia 2001, 127, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Buckeridge, K.M.; Zufelt, E.; Chu, H.; Grogan, P. Soil nitrogen cycling rates in low arctic shrub tundra are enhanced by litter feedbacks. Plant Soil 2010, 330, 407–421. [Google Scholar] [CrossRef]
- Zeng, Y.; Xiang, W.; Deng, X.; Fang, X.; Liu, C.; Peng, C. Soil n forms and gross transformation rates in chinese subtropical forests dominated by different tree species. Plant Soil 2014, 384, 231–242. [Google Scholar] [CrossRef]
- Yan, E.R.; Wang, X.H.; Guo, M.; Zhong, Q.; Zhou, W.; Li, Y.F. Temporal patterns of net soil n mineralization and nitrification through secondary succession in the subtropical forests of eastern China. Plant Soil 2009, 320, 181–194. [Google Scholar] [CrossRef]
- Hart, S.C.; Stark, J.M.; Davidson, E.A.; Firestone, M.K. Nitrogen mineralization, immobilization, and nitrification. In Methods of Soil Analysis, Part 2; Soil Sci. Society of America, Inc.: Madison, WI, USA, 1994; pp. 985–1019. [Google Scholar]
- Taylor, P.G.; Townsend, A. Stoichiometric control of organic carbon-nitrate relationships from soils to the sea. Nature 2010, 464, 1178–1181. [Google Scholar] [CrossRef] [PubMed]
- Jongkind, A.G.; Velthorst, E.; Buurman, P. Soil chemical properties under kauri (agathis australis) in the waitakere ranges, new zealand. Geoderma 2007, 141, 320–331. [Google Scholar] [CrossRef]
- Johnson, D.W.; Miegroet, H.; Lindberg, S.E.; Harrison, R.; Todd, D.E. Nutrient cycling in red spruce forests of the great smoky mountains. Can. J. For. Res. 2011, 21, 769–787. [Google Scholar] [CrossRef]
- Pérez, C.A.; Carmona, M.R.; Aravena, J.C.; Armesto, J.J. Successional changes in soil nitrogen availability, non-symbiotic nitrogen fixation and carbon/nitrogen ratios in southern chilean forest ecosystems. Oecologia 2004, 140, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Yan, E.R.; Wang, X.H.; Huang, J.J.; Li, G.Y.; Zhou, W. Decline of soil nitrogen mineralization and nitrification during forest conversion of evergreen broad-leaved forest to plantations in the subtropical area of eastern china. Biogeochemistry 2008, 89, 239–251. [Google Scholar] [CrossRef]
- Li, Q.; Wang, X.; Jiang, M.; Wu, Y.; Yang, X.; Liao, C.; Liu, F. How environmental and vegetation factors affect spatial patterns of soil carbon and nitrogen in a subtropical mixed forest in central china. J. Soils Sediments 2017, 17, 2296–2304. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | LF1 | LF2 | QJF | QPF |
|---|---|---|---|---|
| Representative plants | Larix kaempferi | Larix kaempferi | Quercus mongolica Juglans mandshurica | Quercus mongolica Populus davidiana |
| Slope (o) | 18 | 18 | 16 | 18 |
| Elevation (m) | 656 | 684 | 672 | 663 |
| Forest age (a) | 43 | 43 | 43 | 46 |
| No. of tree species | 3 | 8 | 12 | 10 |
| Tree density (stems/hm2) | 150 | 410 | 383 | 312 |
| Canopy density | 0.6 | 0.8 | 0.9 | 0.9 |
| Diameter at breast height (cm) | 24.57 ± 5.98 | 24.82 ± 3.74 | 24.53 ± 9.73 | 25.37 ± 8.00 |
| Tree height (m) | 20.16 ± 2.44 | 20.22 ± 1.53 | 19.00 ± 3.56 | 19.88 ± 3.90 |
| Factor | NO3–-N | NH4+-N | Mineral N | Nnit | Namm | Nmin |
|---|---|---|---|---|---|---|
| Forest types | 54.44 *** | 30.31 ** | 37.54 *** | 14.57 ** | 16.86 ** | 4.03 |
| Months | 94.10 ** | 263.11 *** | 81.71 ** | 56.26 ** | 164.12 *** | 88.80 *** |
| Forest types × Months | 2.35 | 15.65 *** | 3.54 ** | 1.02 | 10.30 *** | 1.08 |
| Item | LF1 | LF2 | QJF | QPF |
|---|---|---|---|---|
| Biomass for tree (t/hm2) | ||||
| Foliage | 0.71 ± 0.07c | 2.31 ± 0.43b | 3.05 ± 0.88a | 1.96 ± 0.23b |
| Branches | 4.87 ± 0.60b | 16.28 ± 3.31ab | 34.30 ± 18.5a | 15.10 ± 2.55ab |
| Stems | 31.20 ± 1.27c | 90.66 ± 8.67a | 83.43 ± 25.88ab | 75.70 ± 10.52b |
| Roots | 7.29 ± 0.39c | 21.79 ± 2.69ab | 25.20 ± 7.87a | 18.24 ± 2.32b |
| Total | 44.06 ± 2.33c | 131.03 + 15.04a | 145.98 ± 51.57a | 111.01 ± 15.29b |
| Biomass for herb (t/hm2) | ||||
| Above ground | 0.93 ± 0.57c | 1.57 ± 0.21a | 1.11 ± 0.47b | 0.96 ± 0.39c |
| Below ground | 1.81 ± 0.49a | 2.36 ± 0.59a | 2.80 ± 1.04a | 2.27 ± 0.75a |
| total | 2.75 ± 1.06a | 3.93 ± 0.66a | 3.91 ± 1.46a | 3.23 ± 0.99a |
| Forest floor litter mass (t/hm2) | ||||
| L layer | 0.82 ± 0.16a | 0.87 ± 0.14a | 0.77 ± 0.18a | 0.74 ± 0.23a |
| F+H layer | 3.73 ± 1.62b | 4.46 ± 0.92a | 2.64 ± 0.50b | 2.42 ± 0.42b |
| Total | 4.53 ± 1.76b | 5.33 ± 1.05a | 3.14 ± 0.63b | 3.61 ± 0.64b |
| Annual litterfall mass (kg/hm2) | ||||
| 30.67 ± 1.53c | 44.64 ± 9.16b | 61.33 ± 10.21a | 62.13 ± 2.64a | |
| H’ and MI for tree layer | ||||
| H’ | 0.42 ± 0.07d | 0.89 ± 0.20c | 1.31 ± 0.03b | 2.16 ± 0.25a |
| MI | 0.53 ± 0.07b | 0.63 ± 0.27b | 1.72 ± 0.24a | 1.76 ± 0.18a |
| H’ and MI for herb layer | ||||
| H’ | 2.28 ± 0.13b | 2.07 ± 0.21b | 1.99 ± 0.29b | 2.89 ± 0.54a |
| MI | 4.11 ± 0.38ab | 4.04 ± 0.96b | 3.9 ± 0.53b | 4.93 ± 0.42a |
| Item | LF1 | LF2 | QJF | QPF |
|---|---|---|---|---|
| SOC (g/kg dry soil) | 60.69 ± 6.97ab | 59.17 ± 11.94b | 66.52 ± 9.04a | 63.22 ± 5.24ab |
| TN (g/kg dry soil) | 4.99 ± 0.70ab | 4.46 ± 1.02b | 4.90 ± 1.57ab | 5.26 ± 0.85a |
| C: N | 12.47 ± 2.91a | 10.35 ± 5.42a | 10.40 ± 5.33a | 12.27 ± 1.94a |
| MBC (mg/kg dry soil) | 403.79 ± 13.33b | 406.81 ± 27.18b | 539.28 ± 47.51a | 543.83 ± 15.99a |
| MBN (mg/kg dry soil) | 167.66 ± 38.29a | 169.62 ± 38.80a | 176.52 ± 40.51a | 183.30 ± 30.83a |
| MBC: MBN | 2.54 ± 0.63b | 2.51 ± 0.52b | 3.15 ± 0.90a | 2.97 ± 0.66ab |
| DOC (mg/kg dry soil) | 129.24 ± 23.05a | 138.07 ± 50.85a | 128.89 ± 43.84a | 135.15 ± 40.89a |
| Soil pH | 6.39 ± 0.24a | 6.37 ± 0.24a | 6.67 ± 0.03a | 6.41 ± 0.31a |
| Item | Above Ground | Below Ground | L | F + H | Annual Literfall | Foliage | Branches | Stems | Roots | H’tree | MItree | H’herb | MIherb |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nnit | −0.09 | 0.53 | 0.20 | −0.32 | 0.88 ** | 0.68 * | 0.60 * | 0.55 | 0.63 ** | 0.79 ** | 0.89 ** | 0.27 | 0.42 |
| Namm | 0.26 | −0.31 | 0.15 | 0.51 | −0.79 ** | −0.61 * | −0.59 * | −0.38 | −0.52 | −0.65 * | −0.87 * | 0.10 | −0.17 |
| Nmin | −0.02 | 0.57 | 0.41 | −0.14 | 0.74 ** | 0.55 * | 0.46 | 0.51 | 0.53 | 0.71 ** | 0.72 ** | 0.52 * | 0.53 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Li, F.; Rong, X.; Fan, Z. Plant-Soil Properties Associated with Nitrogen Mineralization: Effect of Conversion of Natural Secondary Forests to Larch Plantations in a Headwater Catchment in Northeast China. Forests 2018, 9, 386. https://doi.org/10.3390/f9070386
Wang Q, Li F, Rong X, Fan Z. Plant-Soil Properties Associated with Nitrogen Mineralization: Effect of Conversion of Natural Secondary Forests to Larch Plantations in a Headwater Catchment in Northeast China. Forests. 2018; 9(7):386. https://doi.org/10.3390/f9070386
Chicago/Turabian StyleWang, Qiong, Fayun Li, Xiangmin Rong, and Zhiping Fan. 2018. "Plant-Soil Properties Associated with Nitrogen Mineralization: Effect of Conversion of Natural Secondary Forests to Larch Plantations in a Headwater Catchment in Northeast China" Forests 9, no. 7: 386. https://doi.org/10.3390/f9070386