Cellular hnRNP A1 Interacts with Nucleocapsid Protein of Porcine Epidemic Diarrhea Virus and Impairs Viral Replication

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus, Cells, Antibodies, and Plasmid
2.2. Animal Experiment and Proteome Study
2.3. Immunoprecipitation and Immunoblotting
2.4. Immunofluorescence and Confocal Microscopy
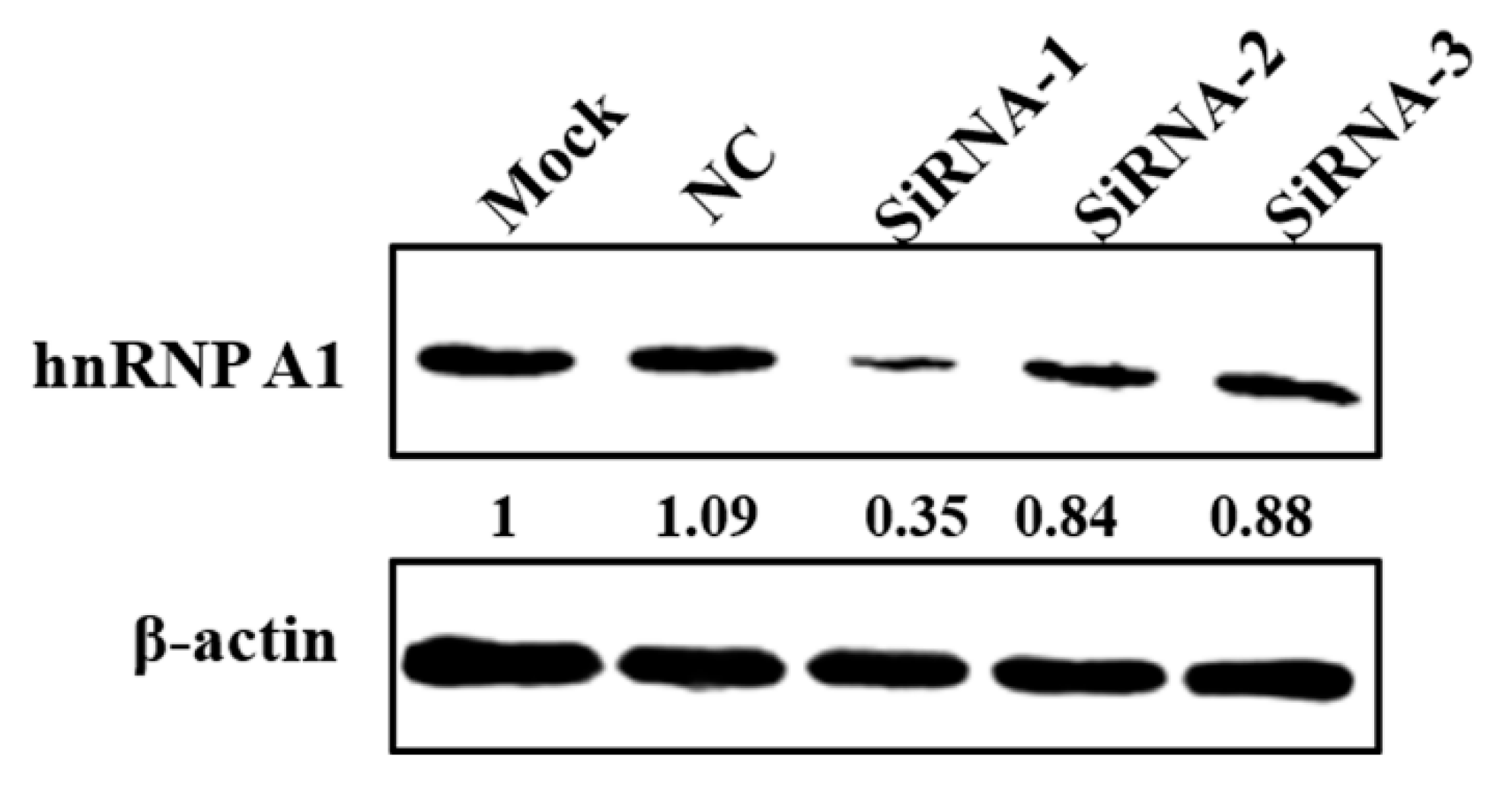
2.5. RNA Interference
2.6. Indirect Immunofluorescence Assay
2.7. Real-Time PCR
2.8. Western Blot
3. Result
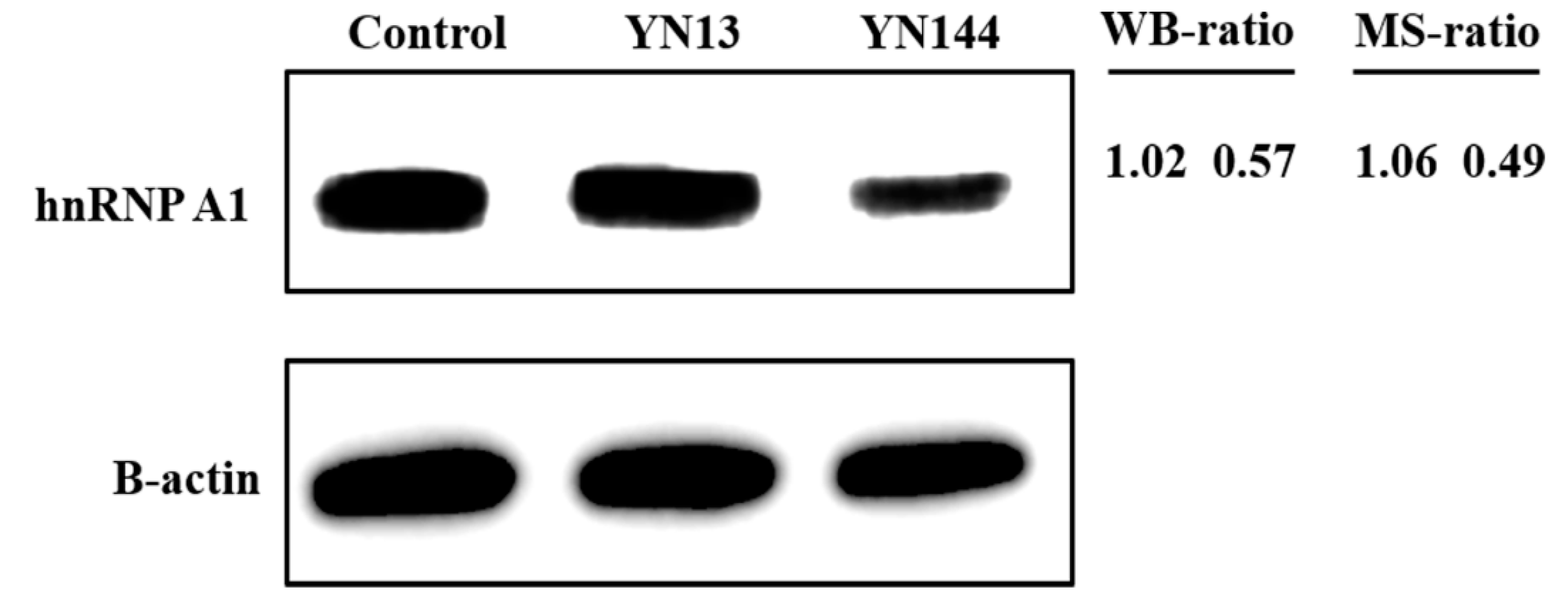
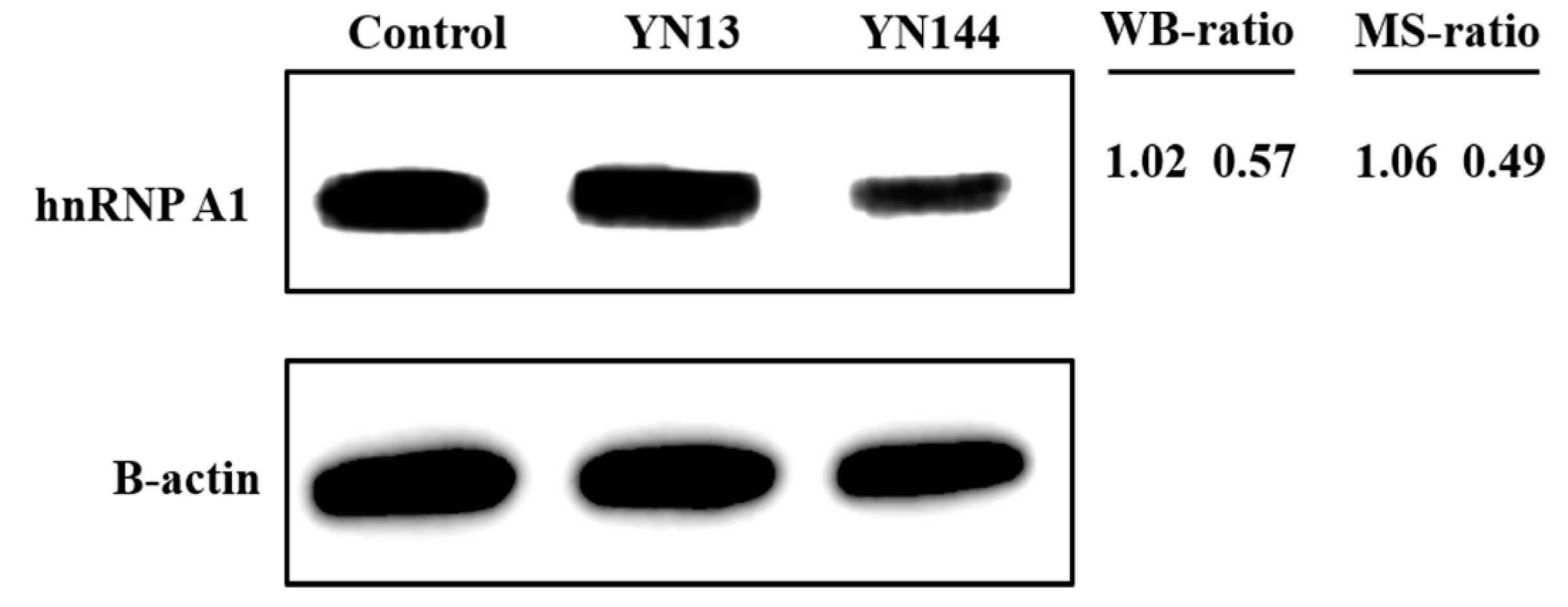
3.1. Different Regulations of hnRNP A1 in Jejunum Tissues Infected with PEDV YN13 Strain and YN144 Strain
3.2. PEDV N Protein Colocalizes with Cellular hnRNP A1 Protein during PEDV Infection
3.3. PEDV N Protein Interacts with Cellular hnRNP A1 Protein
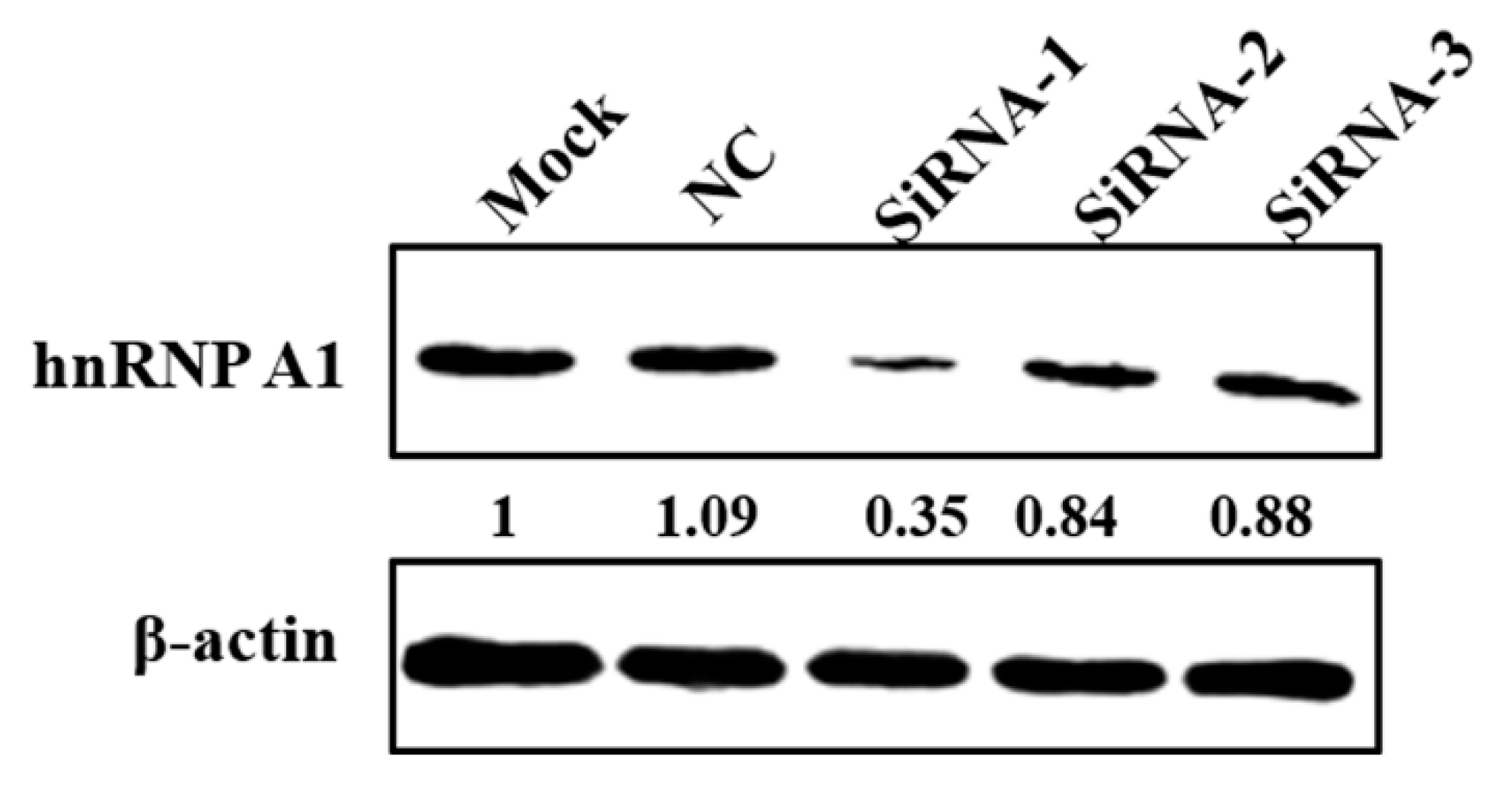
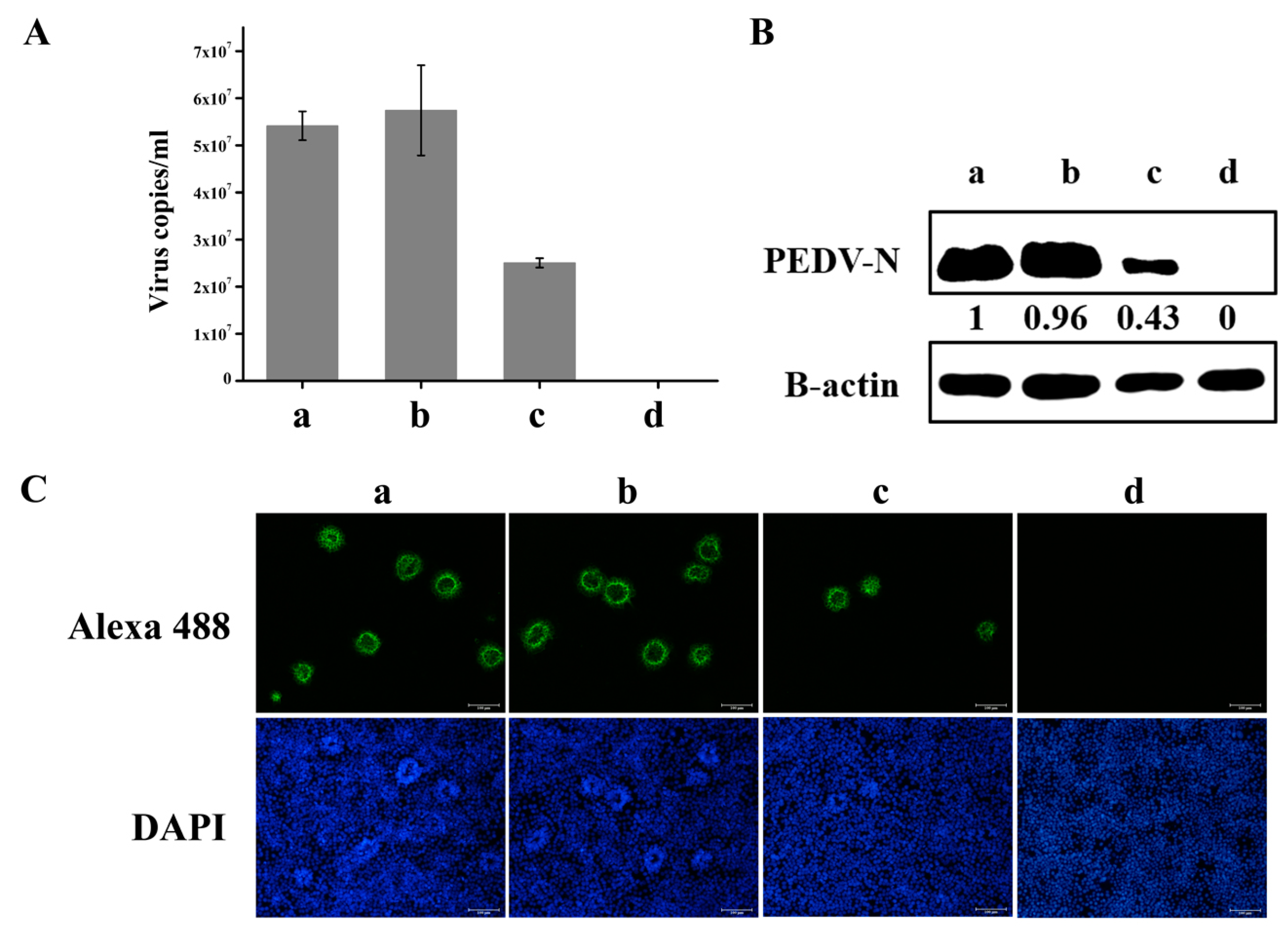
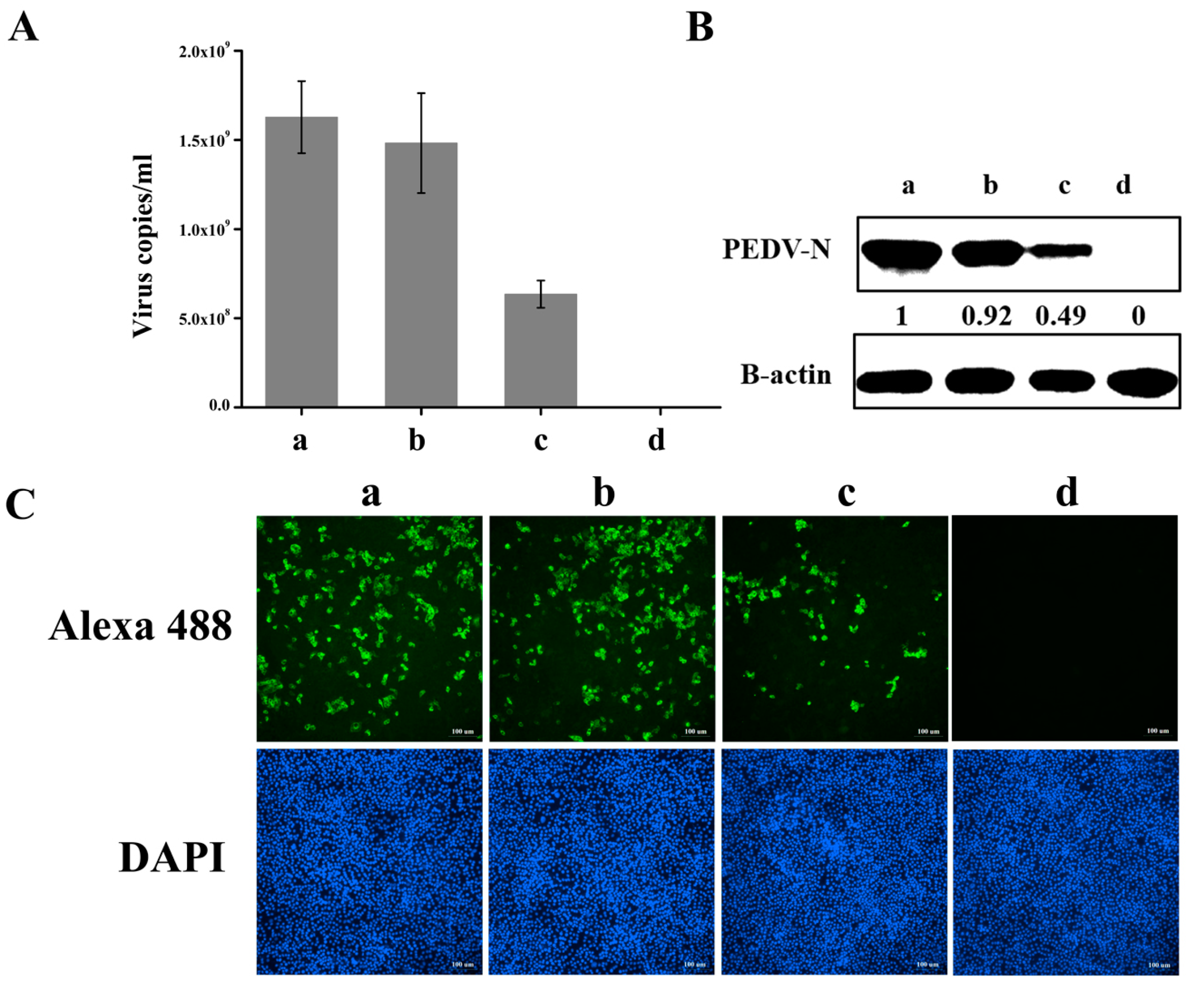
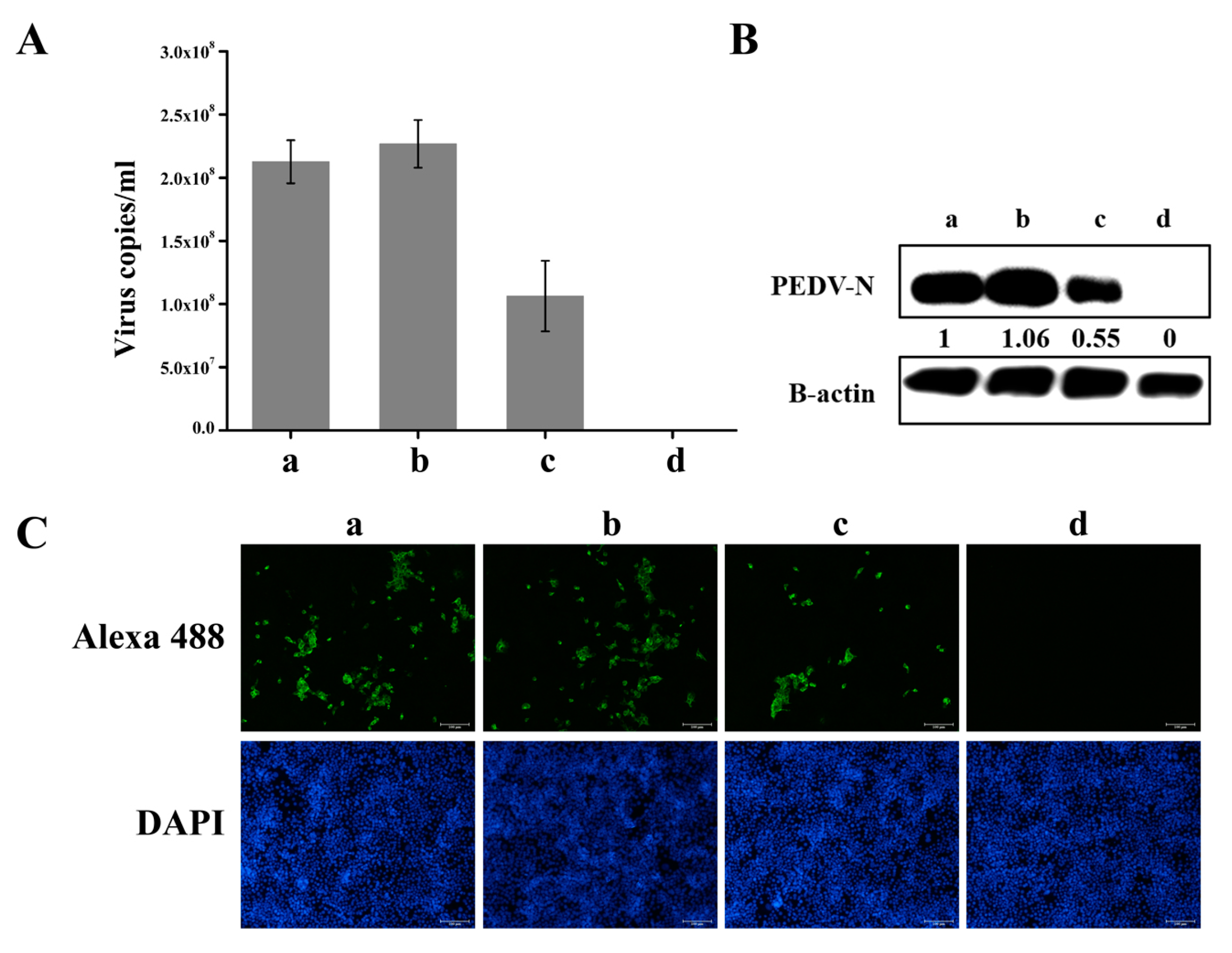
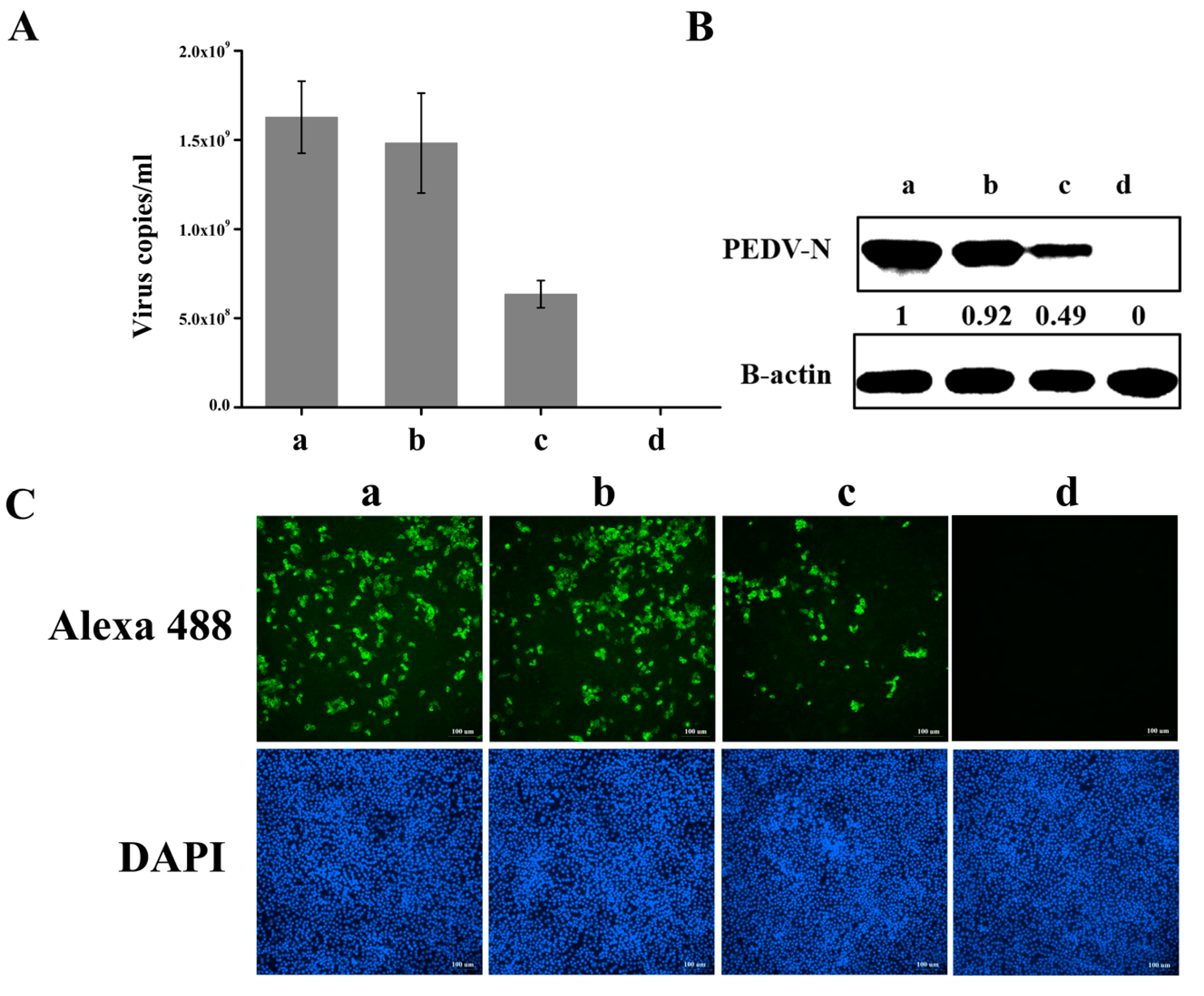
3.4. Knockdown of hnRNP A1 Expression Inhibits YN144 Replication
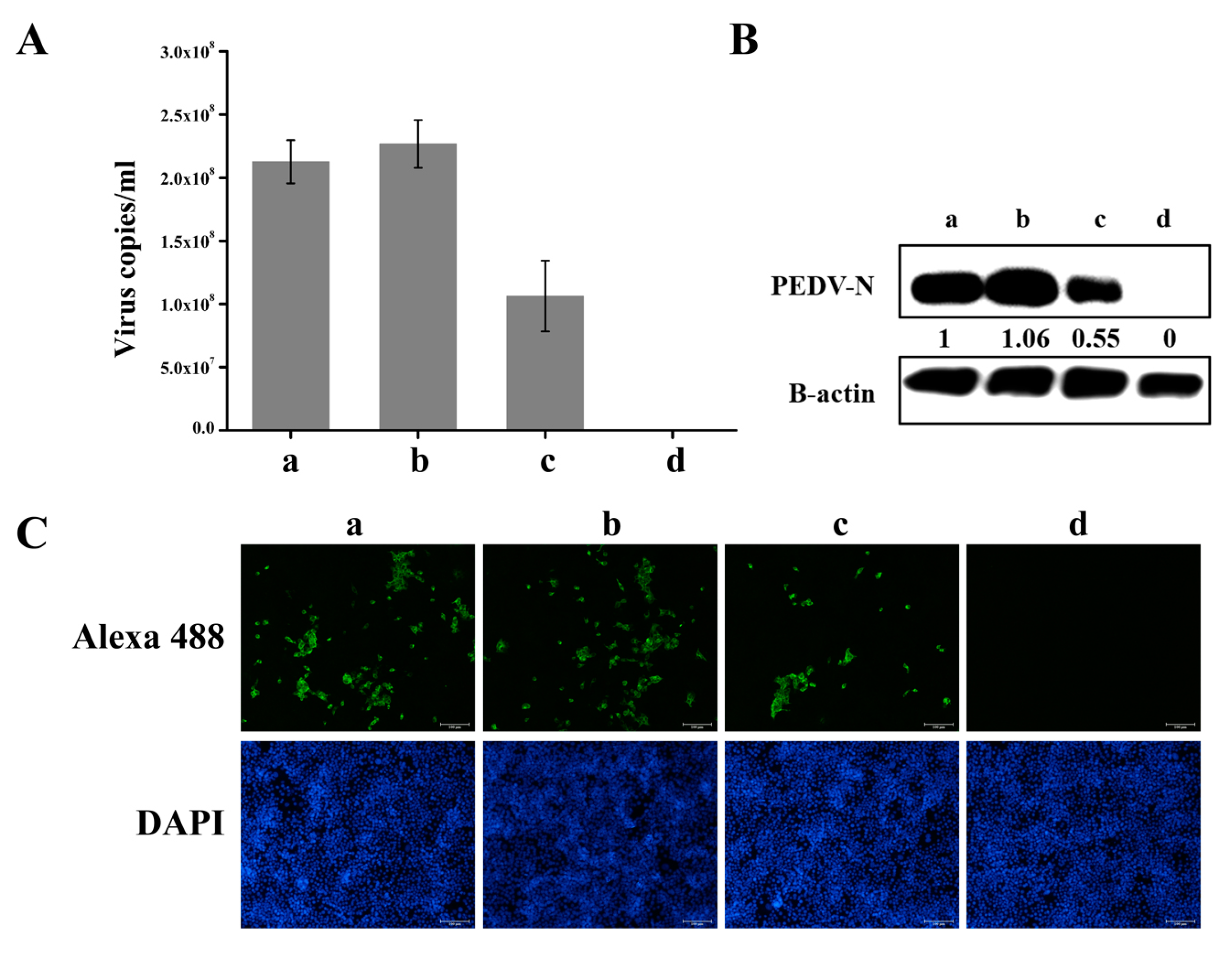
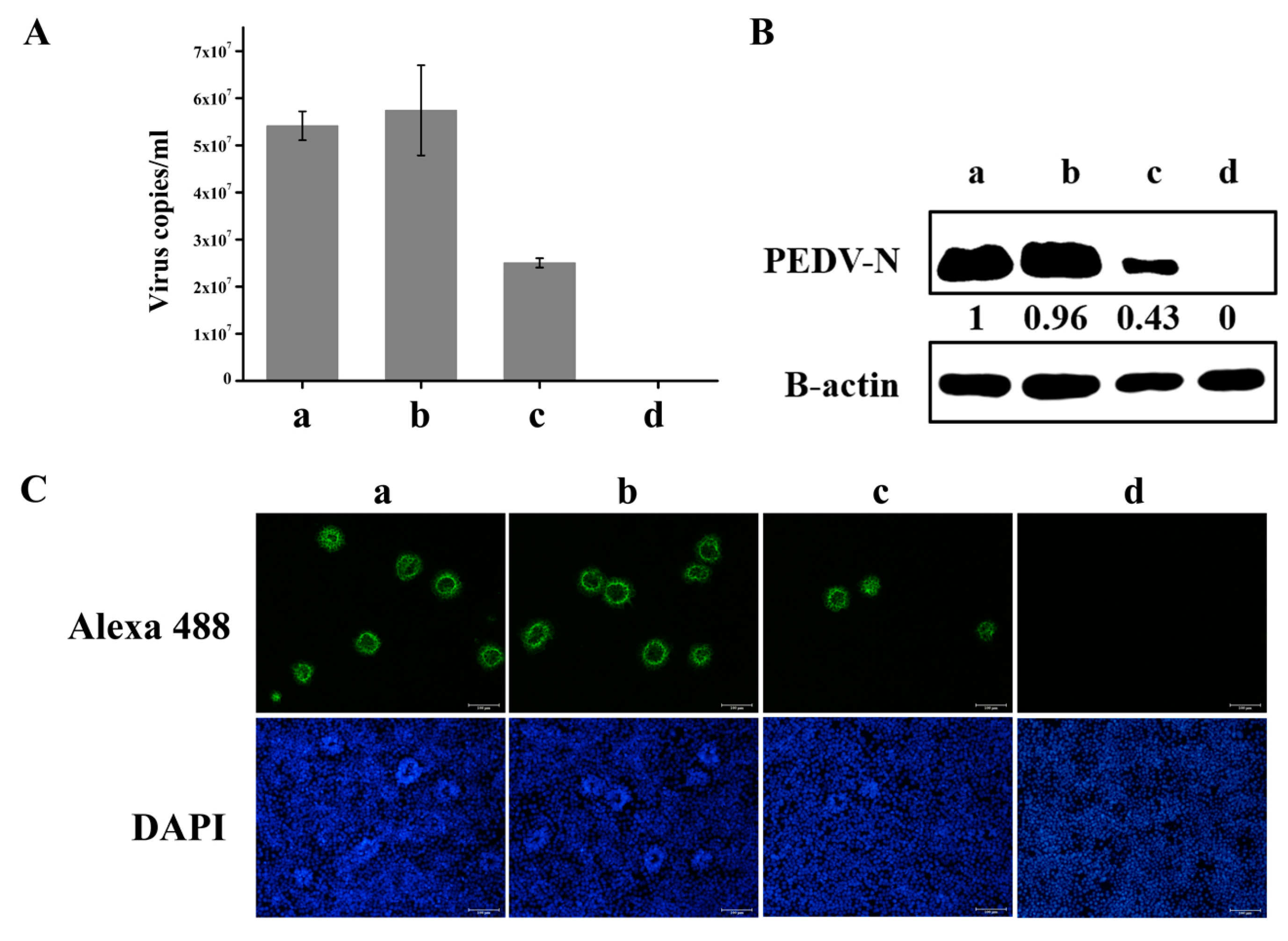
3.5. Knockdown of hnRNP A1 Expression Inhibits YN13 and CV777 Replication
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Choudhury, B.; Dastjerdi, A.; Doyle, N.; Frossard, J.P.; Steinbach, F. From the field to the lab—An European view on the global spread of PEDV. Virus Res. 2016, 226, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Saif, L.J. Porcine epidemic diarrhea virus infection: Etiology, epidemiology, pathogenesis and immunoprophylaxis. Vet. J. 2015, 204, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Park, B. Porcine epidemic diarrhoea virus: A comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes 2012, 44, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, C. Outbreak-Related Porcine Epidemic Diarrhea Virus Strains Similar to US Strains, South Korea, 2013. Emerg. Infect. Dis. 2014, 20, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Marthaler, D.; Wang, Q.; Culhane, M.R.; Rossow, K.D.; Rovira, A.; Collins, J.; Saif, L.J. Distinct Characteristics and Complex Evolution of PEDV Strains, North America, May 2013–February 2014. Emerg. Infect. Dis. 2014, 20, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Adolf, S.; Sandra, R.F.; Friedrich, S.; Jutta, P.; Tatjana, S.; Friedrich, S. First detection, clinical presentation and phylogenetic characterization of Porcine epidemic diarrhea virusin Austria. BMC Vet. Res. 2015, 11, 310. [Google Scholar]
- Mcbride, R.; van, Z.M.; Fielding, B.C. The coronavirus nucleocapsid is a multifunctional protein. Viruses 2014, 6, 2991–3018. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Lv, M.; Chen, J.; Shi, H.; Zhang, S.; Zhang, X.; Feng, L. Molecular characterizations of subcellular localization signals in the nucleocapsid protein of porcine epidemic diarrhea virus. Viruses 2014, 6, 1253–1273. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xie, C.; Zhang, J.; Zhang, W.; Yang, D.; Yu, L.; Jiang, Y.; Yang, S.; Gao, F.; Yang, Z. The Identification and Characterization of Two Novel Epitopes on the Nucleocapsid Protein of the Porcine Epidemic Diarrhea Virus. Sci. Rep. 2016, 6, 39010. [Google Scholar] [CrossRef] [PubMed]
- Da, S.; Shi, H.; Sun, D.; Chen, J.; Xin, Z.; Wang, X.; Zhang, J.; Ji, Z.; Liu, J.; Cao, L. Nucleocapsid Interacts with NPM1 and Protects it from Proteolytic Cleavage, Enhancing Cell Survival, and is Involved in PEDV Growth. Sci. Rep. 2017, 7, 39700. [Google Scholar]
- Ding, Z.; Fang, L.; Jing, H.; Zeng, S.; Wang, D.; Liu, L.; Zhang, H.; Luo, R.; Chen, H.; Xiao, S. Porcine epidemic diarrhea virus nucleocapsid protein antagonizes β interferon production by sequestering the interaction between IRF3 and TBK1. J. Virol. 2014, 88, 8936–8945. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.K.; Chen, C.J.; Wu, C.C.; Chen, S.W.; Shih, S.R.; Kuo, R.L. Cellular hnRNP A2/B1 interacts with the NP of influenza A virus and impacts viral replication. PLoS ONE 2017, 12, e0188214. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Chen, Q.; Chen, J.; Chen, K.; Shen, X.; Jiang, H. The nucleocapsid protein of SARS coronavirus has a high binding affinity to the human cellular heterogeneous nuclear ribonucleoprotein A1. FEBS Lett. 2005, 579, 2623–2628. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.C.; Will, C.L.; Lührmann, R. The Spliceosome: Design Principles of a Dynamic RNP Machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Bekenstein, U.; Soreq, H. Heterogeneous nuclear ribonucleoprotein A1 in health and neurodegenerative disease: From structural insights to post-transcriptional regulatory roles. Mol. Cell. Neurosci. 2013, 56, 436. [Google Scholar] [CrossRef] [PubMed]
- Biamonti, G.; Buvoli, M.; Bassi, M.T.; Morandi, C.; Cobianchi, F.; Riva, S. Isolation of an active gene encoding human hnRNP protein A1: Evidence for alternative splicing. J. Mol. Biol. 1989, 207, 491. [Google Scholar] [CrossRef]
- Jean-Philippe, J.; Paz, S.; Caputi, M. hnRNP A1: The Swiss army knife of gene expression. Int. J. Mol. Sci. 2013, 14, 18999. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Lokugamage, K.G.; Makino, S. Viral and Cellular mRNA Translation in Coronavirus-Infected Cells. Adv. Virus Res. 2016, 96, 165. [Google Scholar] [PubMed]
- Li, Z.; Chen, F.; Ye, S.; Guo, X.; Memon, A.M.; Wu, M.; He, Q. Comparative Proteome Analysis of Porcine Jejunum Tissues in Response to a Virulent Strain of Porcine Epidemic Diarrhea Virus and Its Attenuated Strain. Viruses 2016, 8, 323. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Hu, H.; Chen, F.; Li, Z.; Ye, S.; Cheng, S.; Zhang, M.; He, Q. iTRAQ-based comparative proteomic analysis of Vero cells infected with virulent and CV777 vaccine strain-like strains of porcine epidemic diarrhea virus. J. Proteom. 2016, 130, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhu, Y.; Wu, M.; Ku, X.; Ye, S.; Li, Z.; Guo, X.; He, Q. Comparative Genomic Analysis of Classical and Variant Virulent Parental/Attenuated Strains of Porcine Epidemic Diarrhea Virus. Viruses 2015, 7, 5525–5538. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X. The nucleocapsid protein of coronavirus mouse hepatitis virus interacts with the cellular heterogeneous nuclear ribonucleoprotein A1 in vitro and in vivo. Virology 1999, 265, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Cammas, A.; Lacroix-Triki, M.; Pierredon, S.; Bras, M.L.; Iacovoni, J.S.; Teulade-Fichou, M.P.; Favre, G.; Roché, H.; Filleron, T.; Millevoi, S. hnRNP A1-mediated translational regulation of the G quadruplex-containing RON receptor tyrosine kinase mRNA linked to tumor progression. Oncotarget 2016, 7, 16793–16805. [Google Scholar] [CrossRef] [PubMed]
- Cammas, A.; Pileur, F.; Bonnal, S.; Lewis, S.M.; Lévêque, N.; Holcik, M.; Vagner, S. Cytoplasmic relocalization of heterogeneous nuclear ribonucleoprotein A1 controls translation initiation of specific mRNAs. Mol. Biol. Cell 2007, 18, 5048. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.T.; Huang, P.; Li, H.P.; Lai, M.M. Heterogeneous nuclear ribonucleoprotein A1 regulates RNA synthesis of a cytoplasmic virus. Embo J. 2000, 19, 4701–4711. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.T.; Yu, G.Y.; Lai, M.M.C. Multiple Type A/B Heterogeneous Nuclear Ribonucleoproteins (hnRNPs) Can Replace hnRNP A1 in Mouse Hepatitis Virus RNA Synthesis. J. Virol. 2003, 77, 10584–10593. [Google Scholar] [CrossRef] [PubMed]
- Cencic, R.; Desforges, M.; Hall, D.R.; Kozakov, D.; Du, Y.; Min, J.; Dingledine, R.; Fu, H.; Vajda, S.; Talbot, P.J. Blocking eIF4E–eIF4G Interaction as a Strategy To Impair Coronavirus Replication. J. Virol. 2011, 85, 6381. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, R.E. Nuclear Proteins Hijacked by Mammalian Cytoplasmic Plus Strand RNA Viruses. Virology 2015, 457, 479–480. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequences (5′-3′) |
|---|---|
| SiRNA-1 | GGAAGAGUUGUGGAACCAATT |
| SiRNA-2 | GGAUUUGGUAAUGAUGGAATT |
| SiRNA-3 | GCGGUGGAGGUCAAUACUUTT |
| Negative control siRNA (NC) | UUCUCCGAACGUGUCACGUTT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zeng, W.; Ye, S.; Lv, J.; Nie, A.; Zhang, B.; Sun, Y.; Han, H.; He, Q. Cellular hnRNP A1 Interacts with Nucleocapsid Protein of Porcine Epidemic Diarrhea Virus and Impairs Viral Replication. Viruses 2018, 10, 127. https://doi.org/10.3390/v10030127
Li Z, Zeng W, Ye S, Lv J, Nie A, Zhang B, Sun Y, Han H, He Q. Cellular hnRNP A1 Interacts with Nucleocapsid Protein of Porcine Epidemic Diarrhea Virus and Impairs Viral Replication. Viruses. 2018; 10(3):127. https://doi.org/10.3390/v10030127
Chicago/Turabian StyleLi, Zhonghua, Wei Zeng, Shiyi Ye, Jian Lv, Axiu Nie, Bingzhou Zhang, Yumei Sun, Heyou Han, and Qigai He. 2018. "Cellular hnRNP A1 Interacts with Nucleocapsid Protein of Porcine Epidemic Diarrhea Virus and Impairs Viral Replication" Viruses 10, no. 3: 127. https://doi.org/10.3390/v10030127
APA StyleLi, Z., Zeng, W., Ye, S., Lv, J., Nie, A., Zhang, B., Sun, Y., Han, H., & He, Q. (2018). Cellular hnRNP A1 Interacts with Nucleocapsid Protein of Porcine Epidemic Diarrhea Virus and Impairs Viral Replication. Viruses, 10(3), 127. https://doi.org/10.3390/v10030127