Molecular Diagnosis of Felis catus Gammaherpesvirus 1 (FcaGHV1) Infection in Cats of Known Retrovirus Status with and without Lymphoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Samples
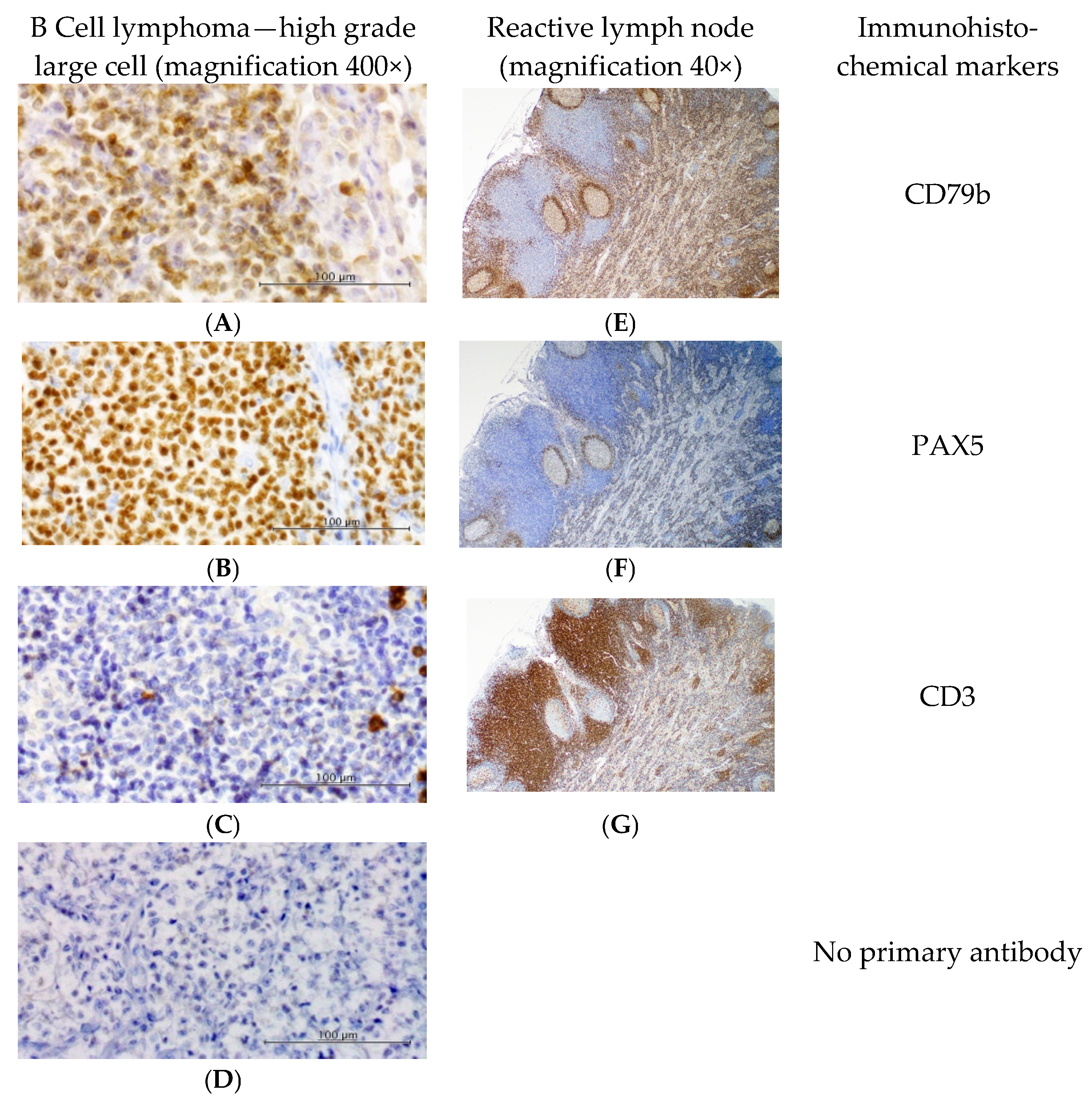
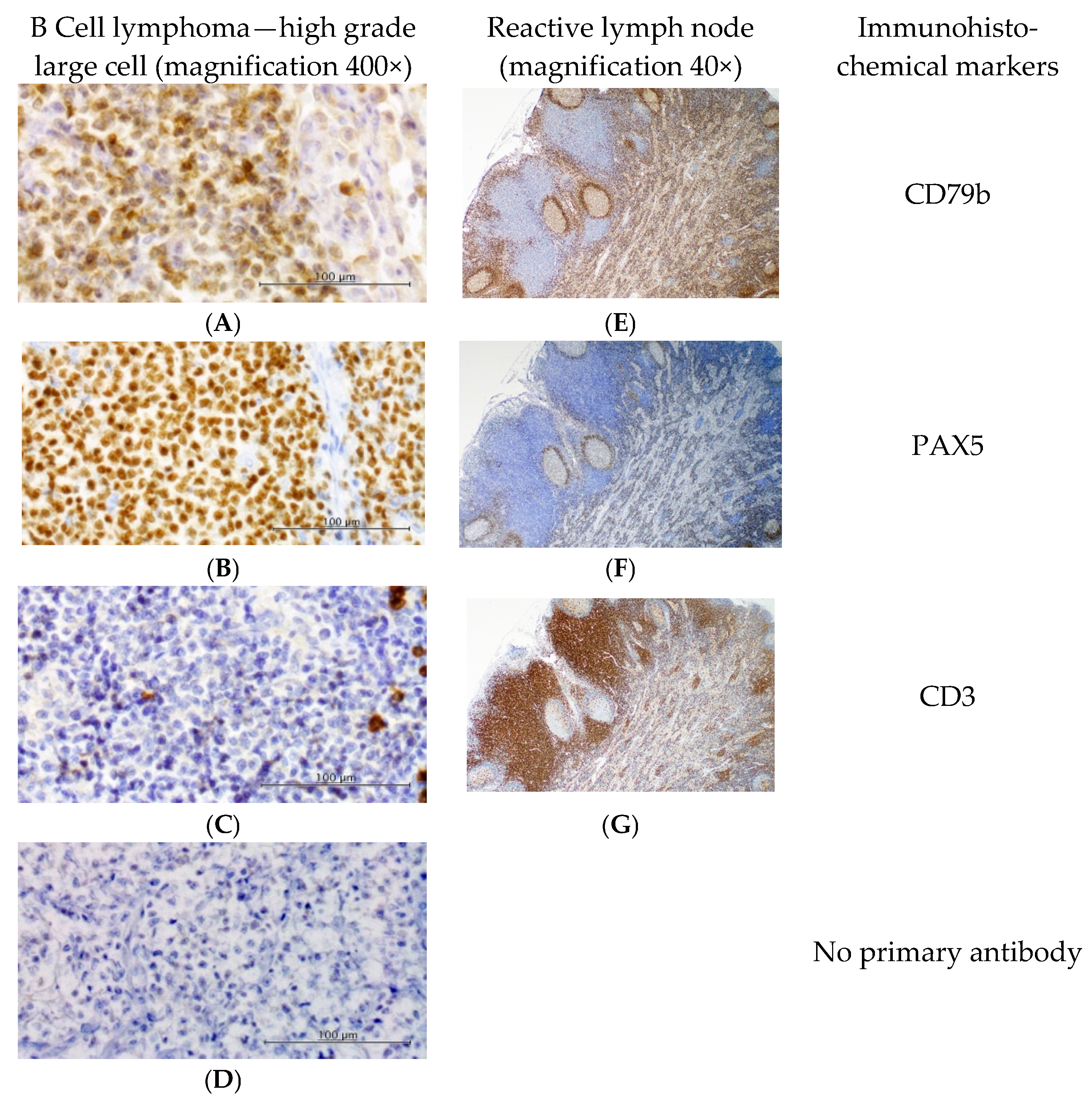
2.3. Immunophenotyping
2.4. Virus DNA Detection
2.4.1. DNA Extraction
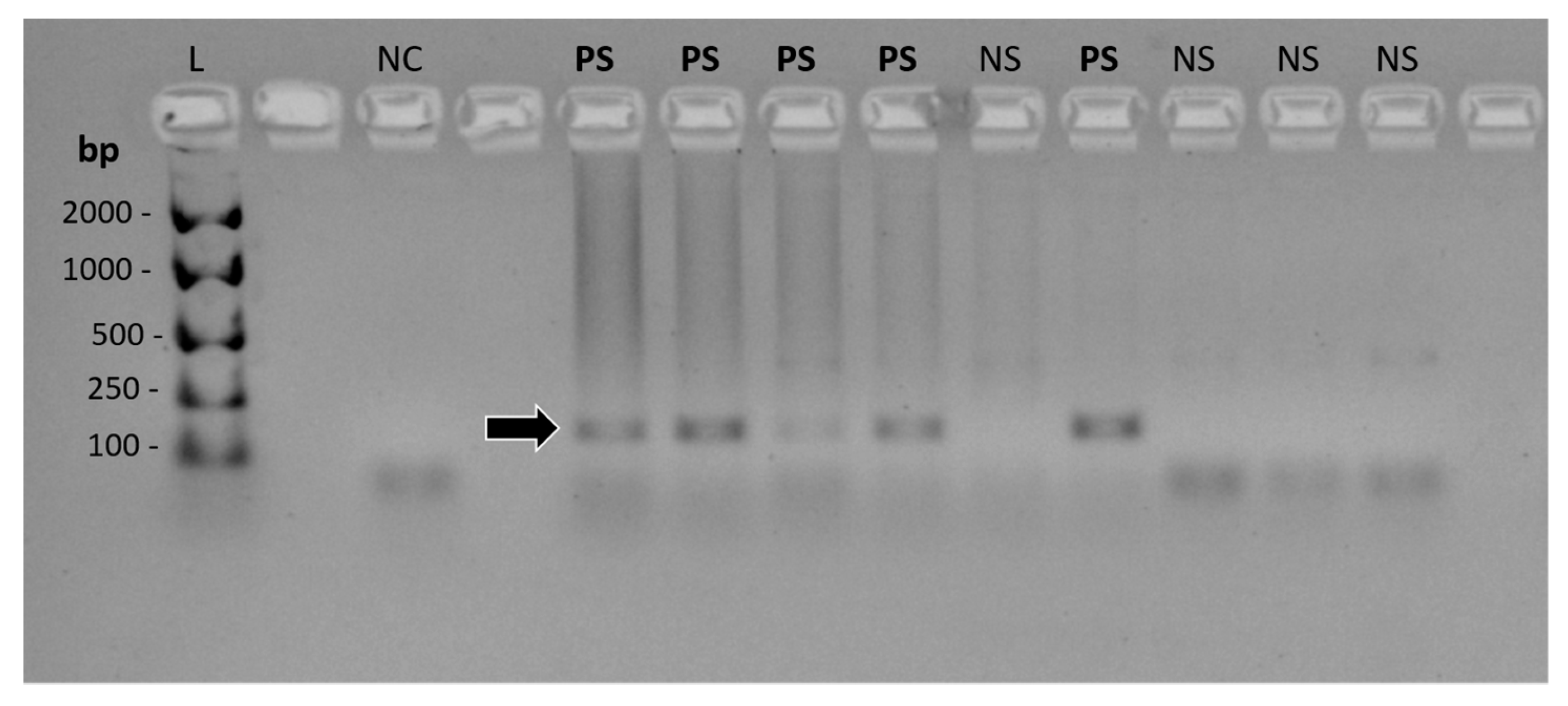
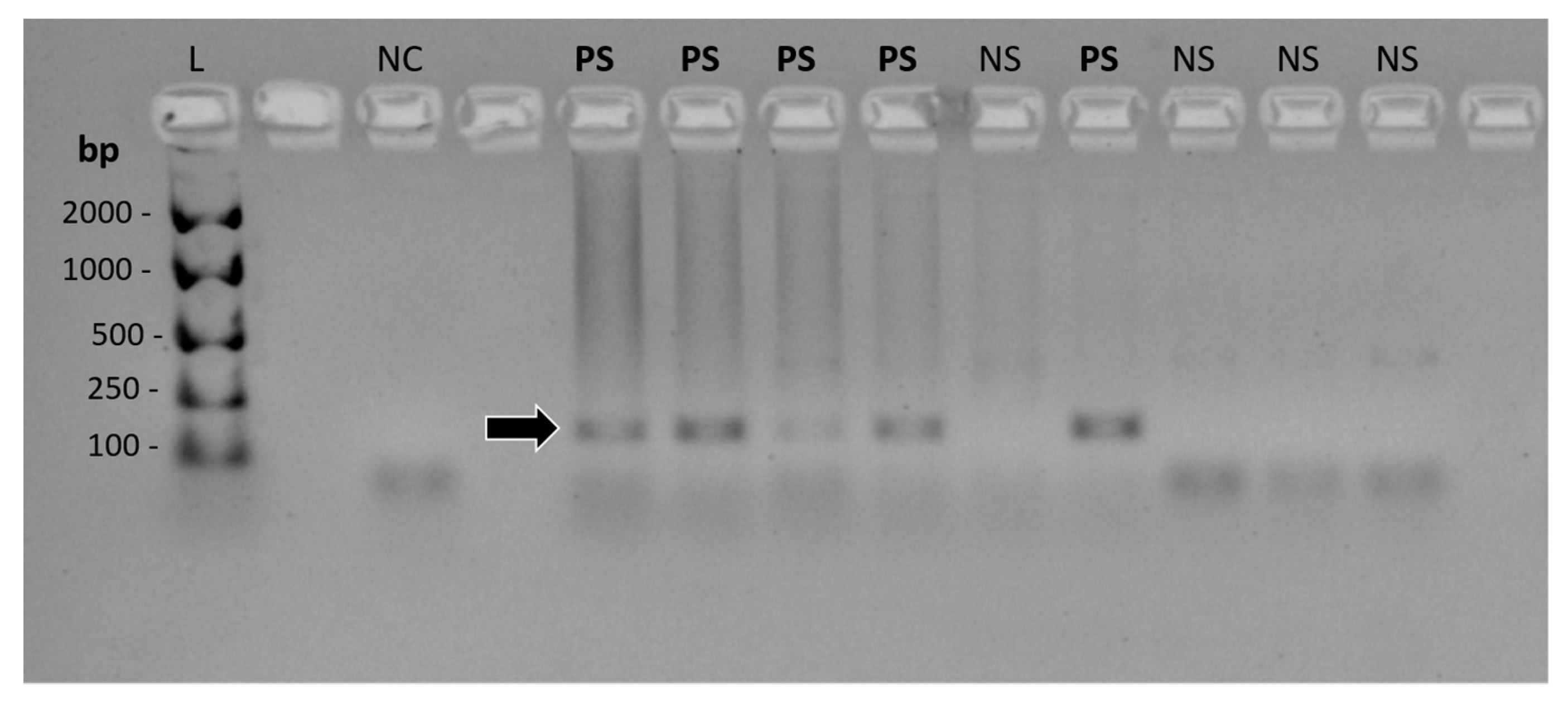
2.4.2. FcaGHV1 PCR
2.4.3. FIV and FeLV cPCR
2.4.4. cPCR Product Detection and Confirmation
2.5. Individual Infection Status
2.6. Age- and Sex-Matching
2.7. Statistical Analysis
3. Results
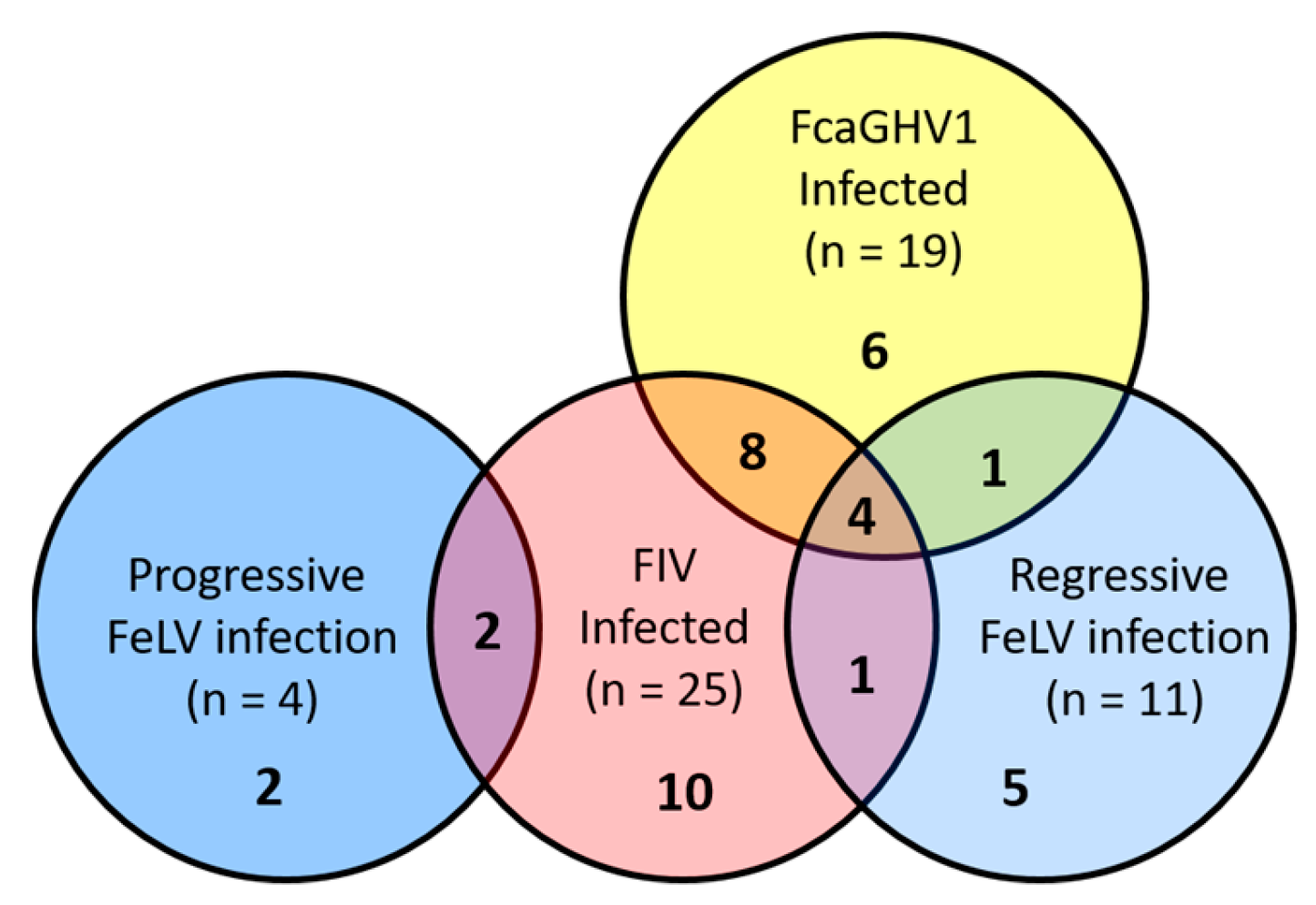
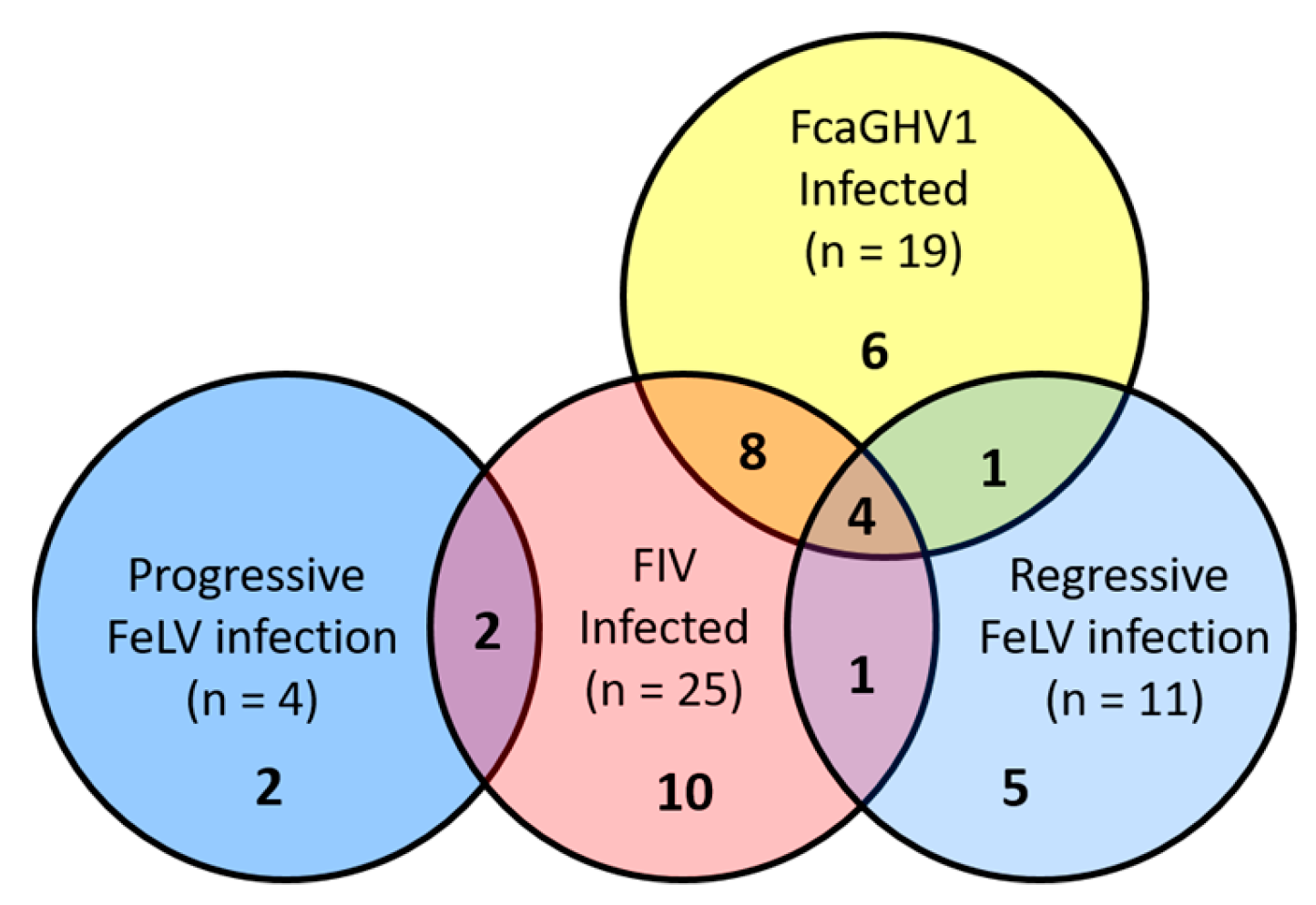
3.1. Viral Infection Status and Lymphoma Classification
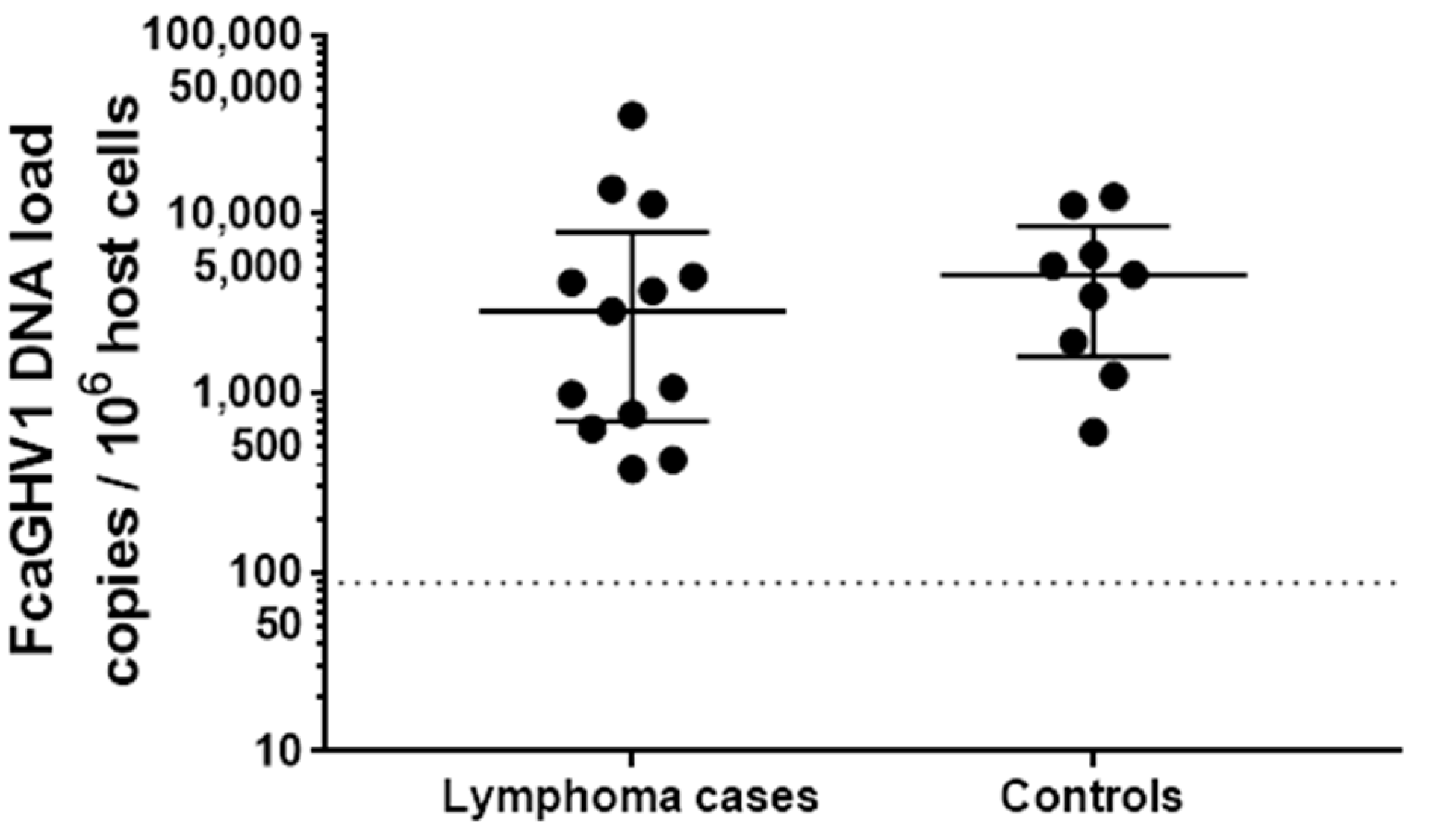
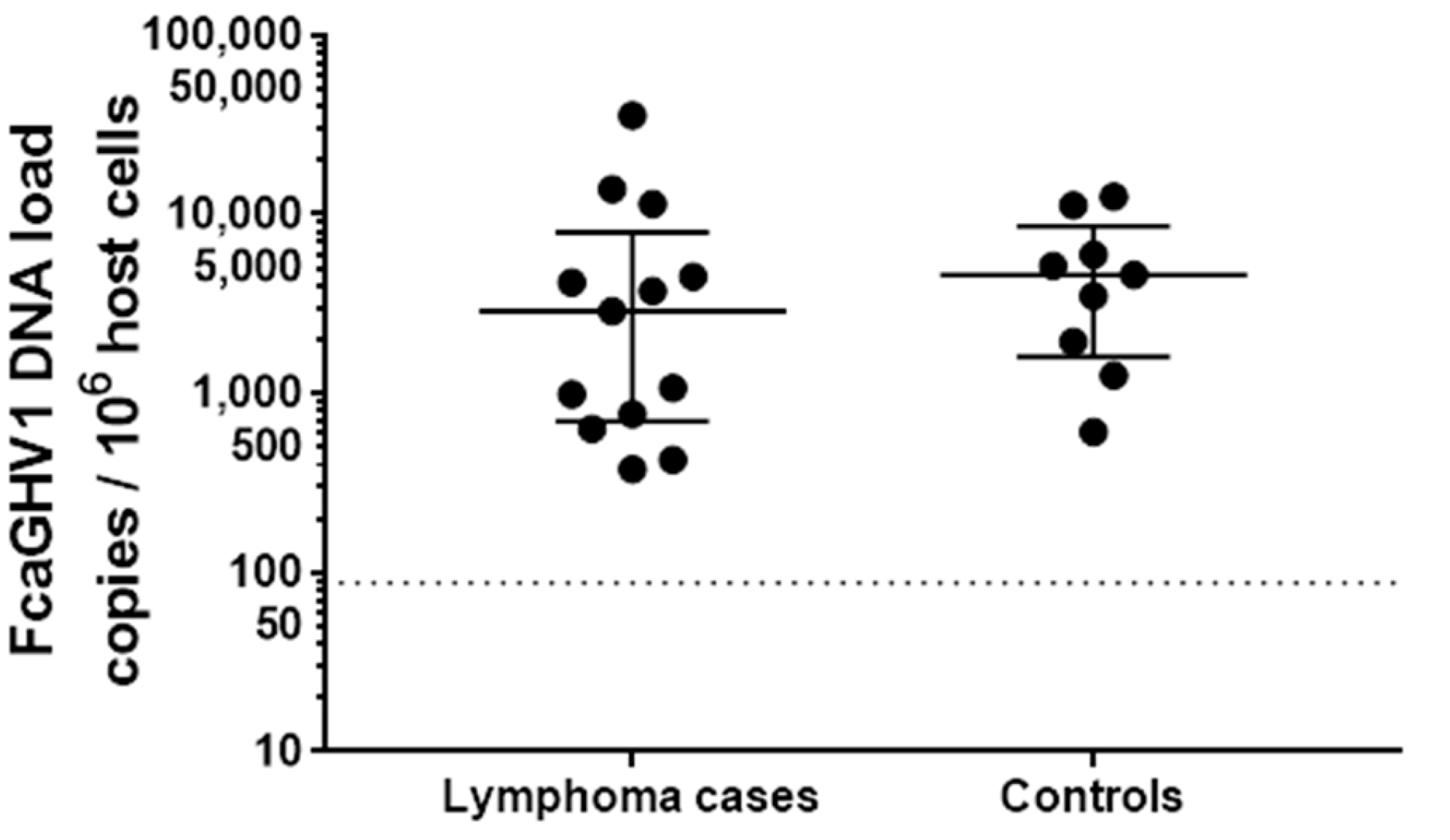
3.2. Association between FcaGHV1 DNA Detection and Lymphoma
3.3. FeLV Infection
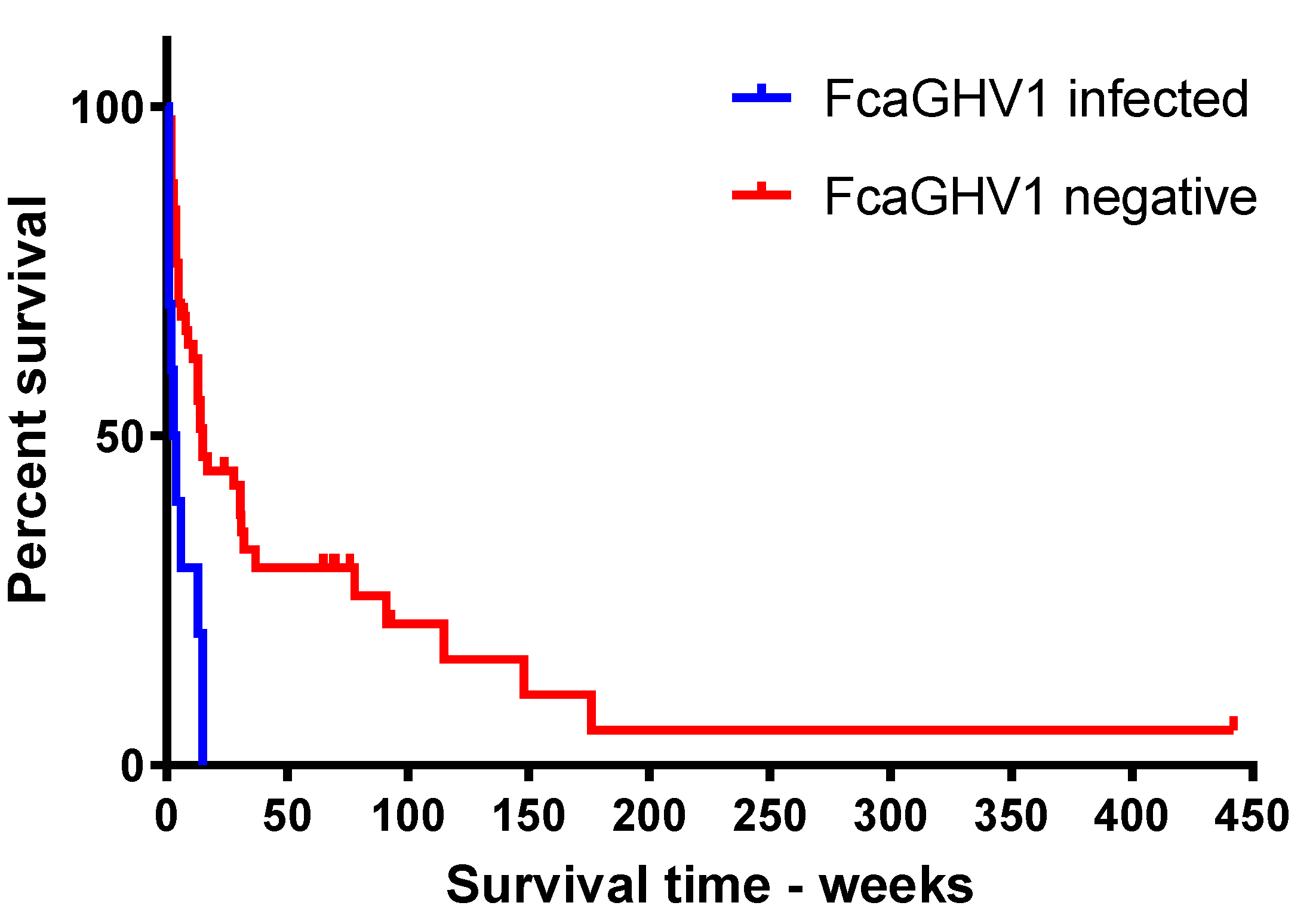
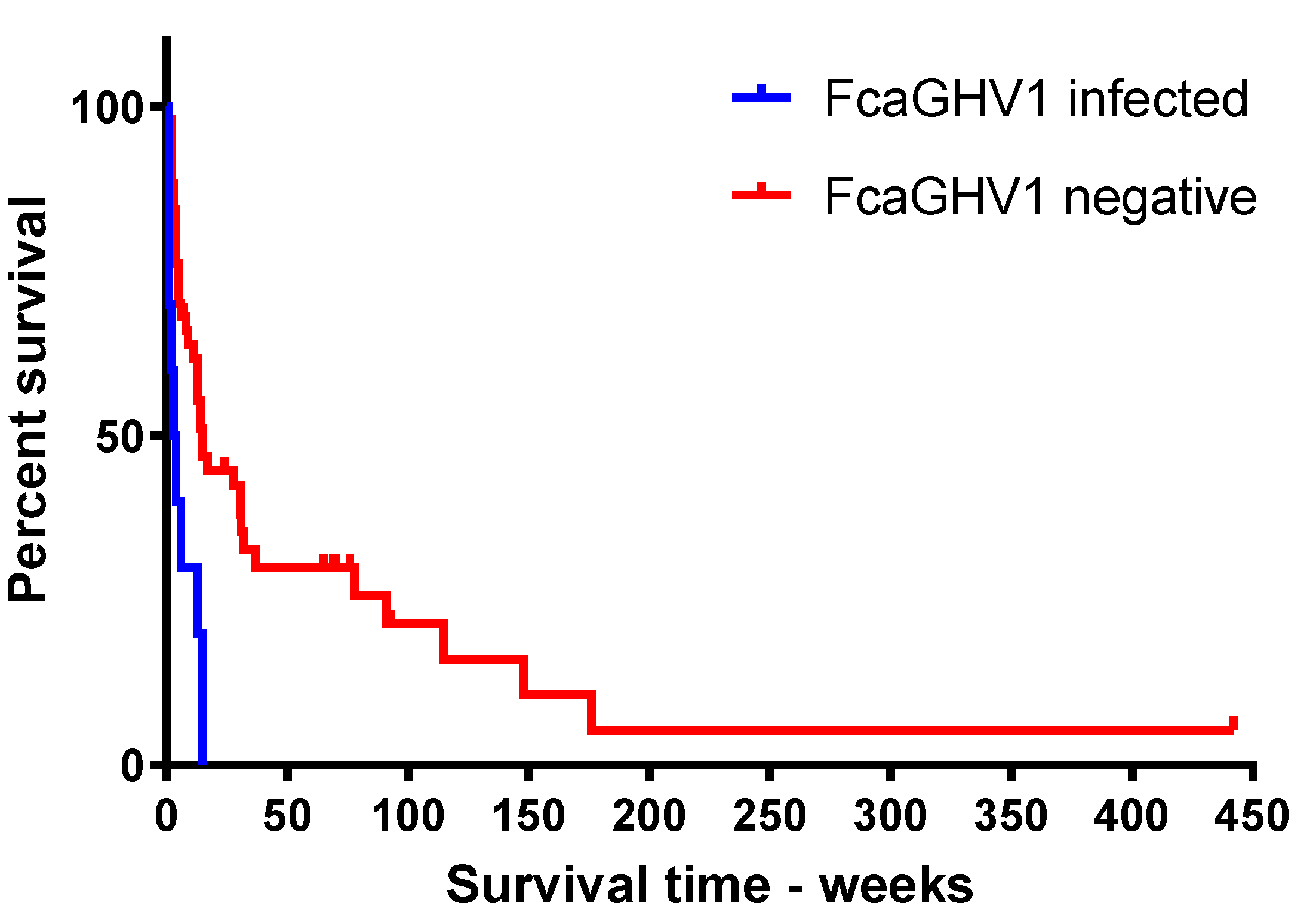
3.4. FcaGHV1 DNAemia and Survival
3.5. Outdoor Access and Fighting History as Risk Factors for FcaGHV1 Detection
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ackermann, M.; Engels, M.; Fraefel, C.; Metzler, A.; Schwyzer, M.; Suter, M.; Tobler, K. Herpesviruses: Balance in power and powers imbalanced. Vet. Microbiol. 2002, 86, 175–181. [Google Scholar] [CrossRef]
- Ackermann, M. Pathogenesis of gammaherpesvirus infections. Vet. Microbiol. 2006, 113, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Lankester, F.; Lugelo, A.; Kazwala, R.; Keyyu, J.; Cleaveland, S.; Yoder, J. The economic impact of malignant catarrhal fever on pastoralist livelihoods. PLoS ONE 2015, 10, e0116059. [Google Scholar] [CrossRef] [PubMed]
- Plummer, M.; de Martel, C.; Vignat, J.; Ferlay, J.; Bray, F.; Franceschi, S. Global burden of cancers attributable to infections in 2012: A synthetic analysis. Lancet Glob. Health 2016, 4, e609–e616. [Google Scholar] [CrossRef]
- Parkin, D.M. The global health burden of infection-associated cancers in the year 2002. Int. J. Cancer 2006, 118, 3030–3044. [Google Scholar] [CrossRef] [PubMed]
- Troyer, R.M.; Beatty, J.A.; Stutzman-Rodriguez, K.R.; Carver, S.; Lozano, C.C.; Lee, J.S.; Lappin, M.R.; Riley, S.P.D.; Serieys, L.E.K.; Logan, K.A.; et al. Novel gammaherpesviruses in north american domestic cats, bobcats, and pumas: Identification, prevalence, and risk factors. J. Virol. 2014, 88, 3914–3924. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J.A.; Troyer, R.M.; Carver, S.; Barrs, V.R.; Espinasse, F.; Conradi, O.; Stutzman-Rodriguez, K.; Chan, C.C.; Tasker, S.; Lappin, M.R.; et al. Felis catus gammaherpesvirus 1; a widely endemic potential pathogen of domestic cats. Virology 2014, 460, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Ertl, R.; Korb, M.; Langbein-Detsch, I.; Klein, D. Prevalence and risk factors of gammaherpesvirus infection in domestic cats in central europe. Virol. J. 2015, 12, 1–4. [Google Scholar] [CrossRef] [PubMed]
- McLuckie, A.; Tasker, S.; Dhand, N.K.; Spencer, S.; Beatty, J.A. High prevalence of felis catus gammaherpesvirus 1 infection in haemoplasma-infected cats supports co-transmission. Vet. J. 2016, 214, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Tateno, M.; Takahashi, M.; Miyake, E.; Nishigaki, K.; Tsujimoto, H.; Endo, Y. Molecular epidemiological study of gammaherpesvirus in domestic cats in Japan. J. Vet. Med. Sci. 2017, 79, 1735–1740. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, C.A.; Menotti-Raymond, M.; Roca, A.L.; Hupe, K.; Johnson, W.E.; Geffen, E.; Harley, E.H.; Delibes, M.; Pontier, D.; Kitchener, A.C.; et al. The near eastern origin of cat domestication. Science (N. Y.) 2007, 317, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Luria, B.J.; Levy, J.K.; Lappin, M.R.; Breitschwerdt, E.B.; Legendre, A.M.; Hernandez, J.A.; Gorman, S.P.; Lee, I.T. Prevalence of infectious diseases in feral cats in northern florida. J. Feline Med. Surg. 2004, 6, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Georges, K.; Ezeokoli, C.; Auguste, T.; Seepersad, N.; Pottinger, A.; Sparagano, O.; Tasker, S. A comparison of real-time pcr and reverse line blot hybridization in detecting feline haemoplasmas of domestic cats and an analysis of risk factors associated with haemoplasma infections. BMC Vet. Res. 2012, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- McLuckie, A.J.; Barrs, V.R.; Smith, A.L.; Beatty, J.A. Detection of felis catus gammaherpesvirus 1 (Fcaghv1) in peripheral blood B- and T-lymphocytes in asymptomatic, naturally-infected domestic cats. Virology 2016, 497, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Dorn, C.R.; Taylor, D.O.N.; Schneider, R.; Hibbard, H.H.; Klauber, M.R. Survey of animal neoplasms in alameda and contra costa counties, california. Ii. Cancer morbidity in dogs and cats from alameda county. J. Natl. Cancer Inst. 1968, 40, 307–318. [Google Scholar] [PubMed]
- Jarrett, W.F.H.; Davie, F.; Martin, W.B.; Crawford, E.M. A Virus-like particle associated with leukaemia (lymphosarcoma). Nature 1964, 202, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Cotter, S.M.; Hardy, W.D.; Essex, M. Association of feline leukemia-virus with lymphosarcoma and other disorders in the cat. J. Am. Vet. Med. Assoc. 1975, 166, 449–454. [Google Scholar]
- Meichner, K.; Kruse, B.D.; Hirschberger, J.; Hartmann, K. Changes in prevalence of progressive feline leukaemia virus infection in cats with lymphoma in germany. Vet. Rec. 2012, 171, 348. [Google Scholar] [CrossRef] [PubMed]
- Louwerens, M.; London, C.A.; Pedersen, N.C.; Lyons, L.A. Feline lymphoma in the Post—Feline leukemia virus era. J. Vet. Intern. Med. 2005, 19, 329–335. [Google Scholar] [PubMed]
- Sarid, R.; Gao, S.-J. Viruses and human cancer: From detection to causality. Cancer Lett. 2011, 305, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Shelton, G.H.; Grant, C.K.; Cotter, S.M.; Gardner, M.B.; Hardy, W.D., Jr.; DiGiacomo, R.F. Feline immunodeficiency virus and feline leukemia virus infections and their relationships to lymphoid malignancies in cats: A retrospective study (1968–1988). J. Acquir. Immune Defic. Syndr. 1990, 3, 623–630. [Google Scholar] [PubMed]
- Hoffmann-Fezer, G.; Thum, J.; Ackley, C.; Herbold, M.; Mysliwietz, J.; Thefeld, S.; Hartmann, K.; Kraft, W. Decline in CD4+ cell numbers in cats with naturally acquired feline immunodeficiency virus infection. J. Virol. 1992, 66, 1484–1488. [Google Scholar] [PubMed]
- Cesarman, E. Gammaherpesvirus and lymphoproliferative disorders in immunocompromised patients. Cancer Lett. 2011, 305, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Valli, V.E.; Jacobs, R.M.; Norris, A.; Couto, C.G.; Morrison, W.B.; McCaw, D.; Cotter, S.; Ogilvie, G.; Moore, A. The histologic classification of 602 cases of feline lymphoproliferative disease using the national cancer institute working formulation. J. Vet. Diagn. Investig. 2000, 12, 295–306. [Google Scholar] [CrossRef] [PubMed]
- McLuckie, A.; Barrs, V.; Wilson, B.; Westman, M.; Beatty, J. Felis catus gammaherpesvirus 1 DNAemia in whole blood from therapeutically immunosuppressed or retrovirus-infected cats. Vet. Sci. 2017, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Whitney, J.L.; Krockenberger, M.B.; Day, M.J.; Beatty, J.A.; Dhand, N.K.; Barrs, V.R. Immunohistochemical analysis of leucocyte subsets in the sinonasal mucosa of cats with upper respiratory tract aspergillosis. J. Comp. Pathol. 2016, 155, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, K.A.; Krockenberger, M.; Beatty, J.A.; Crowley, A.; Dennis, M.M.; Canfield, P.J.; Dhand, N.; Lingard, A.E.; Barrs, V.R. Histopathological and immunohistochemical evaluation of 53 cases of feline lymphoplasmacytic enteritis and low-grade alimentary lymphoma. J. Comp. Pathol. 2011, 145, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Kidney, B.A.; Ellis, J.A.; Haines, D.M.; Jackson, M.L. Evaluation of formalin-fixed paraffin-embedded tissues obtained from vaccine site-associated sarcomas of cats for DNA of feline immunodeficiency virus. Am. J. Vet. Res. 2000, 61, 1037–1041. [Google Scholar] [CrossRef] [PubMed]
- Tandon, R.; Cattori, V.; Gomes-Keller, M.A.; Meli, M.L.; Golder, M.C.; Lutz, H.; Hofmann-Lehmann, R. Quantitation of feline leukaemia virus viral and proviral loads by taqman real-time polymerase chain reaction. J. Virol. Methods 2005, 130, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Liem, B.P.; Dhand, N.K.; Pepper, A.E.; Barrs, V.R.; Beatty, J.A. Clinical findings and survival in cats naturally infected with feline immunodeficiency virus. J. Vet. Intern. Med. 2013, 27, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.B. The environment and disease: Association or causation? Reprinted from JRSM, vol. 58, 1965. J. R. Soc. Med. 2015, 108, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Bouvard, V.; Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; Ghissassi, F.E.; Benbrahim-Tallaa, L.; Guha, N.; Freeman, C.; Galichet, L.; et al. A review of human carcinogens—Part B: Biological agents. Lancet Oncol. 2009, 10, 321–322. [Google Scholar] [CrossRef]
- Dunmire, S.K.; Hogquist, K.A.; Balfour, H.H. Infectious mononucleosis. Curr. Top. Microbiol. Immunol. 2015, 390, 211–240. [Google Scholar] [PubMed]
- Laroche, C.; Drouet, E.B.; Brousset, P.; Pain, C.; Boibieux, A.; Biron, F.; Icart, J.; Denoyel, G.A.; Niveleau, A. Measurement by the polymerase chain reaction of the epstein-barr virus load in infectious mononucleosis and aids-related non-hodgkin’s lymphomas. J. Med. Virol. 1995, 46, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Kim, S.C.; Chima, C.O.; Israel, B.F.; Lawless, K.M.; Eagan, P.A.; Elmore, S.; Moore, D.T.; Schichman, S.A.; Swinnen, L.J.; et al. Epstein–barr viral load as a marker of lymphoma in aids patients. J. Med. Virol. 2005, 75, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Gartner, B.; Preiksaitis, J.K. Ebv viral load detection in clinical virology. J. Clin. Virol. 2010, 48, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.; Miller, G.; Atchison, R.W.; Breinig, M.K.; Dummer, J.S.; Andiman, W.; Starzl, T.E.; Eastman, R.; Griffith, B.P.; Hardesty, R.L.; et al. Epstein-barr virus infections and DNA hybridization studies in posttransplantation lymphoma and lymphoproliferative lesions: The role of primary infection. J. Infect. Dis. 1985, 152, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Stutzman-Rodriguez, K.; Rovnak, J.; VandeWoude, S.; Troyer, R.M. Domestic cats seropositive for felis catus gammaherpesvirus 1 are often qPCR negative. Virology 2016, 498, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the world health organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Fujino, Y.; Chino, J.; Takahashi, M.; Fukushima, K.; Goto-Koshino, Y.; Uchida, K.; Ohno, K.; Tsujimoto, H. Prognostic analyses on anatomical and morphological classification of feline lymphoma. J. Vet. Med. Sci. 2014, 76, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Collette, S.A.; Allstadt, S.D.; Chon, E.M.; Vernau, W.; Smith, A.N.; Garrett, L.D.; Choy, K.; Rebhun, R.B.; Rodriguez, C.O., Jr.; Skorupski, K.A. Treatment of feline intermediate- to high-grade lymphoma with a modified university of wisconsin-madison protocol: 119 cases (2004–2012). Vet. Comp. Oncol. 2016, 14, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Russell, K.J.; Beatty, J.A.; Dhand, N.; Gunew, M.; Lingard, A.E.; Baral, R.M.; Barrs, V.R. Feline low-grade alimentary lymphoma: How common is it? J. Feline Med. Surg. 2012, 14, 910–912. [Google Scholar] [CrossRef] [PubMed]
- Stein, T.J.; Pellin, M.; Steinberg, H.; Chun, R. Treatment of feline gastrointestinal small-cell lymphoma with chlorambucil and glucocorticoids. J. Am. Anim. Hosp. Assoc. 2010, 46, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.T.A.; Klopfleisch, R.; Gruber, A.D. Prevalence of feline leukaemia provirus DNA in feline lymphomas. J. Feline Med. Surg. 2010, 12, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Gabor, L.J.; Canfield, P.J.; Malik, R. Immunophenotypic and histological characterisation of 109 cases of feline lymphosarcoma. Aust. Vet. J. 1999, 77, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Cesarman, E.; Pessin, M.S.; Lee, F.; Culpepper, J.; Knowles, D.M.; Moore, P.S. Identification of herpesvirus-like DNA-sequences in aids-associated kaposis-sarcoma. Science 1994, 266, 1865–1869. [Google Scholar] [CrossRef] [PubMed]
- Tisi, M.C.; Cupelli, E.; Santangelo, R.; Maiolo, E.; Alma, E.; Giachelia, M.; Martini, M.; Bellesi, S.; D’Alò, F.; Voso, M.T.; et al. Whole blood EBV-DNA predicts outcome in diffuse large b-cell lymphoma. Leuk. Lymphoma 2016, 57, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Gabor, L.J.; Jackson, M.L.; Trask, B.; Malik, R.; Canfield, P.J. Feline leukaemia virus status of australian cats with lymphosarcoma. Aust. Vet. J. 2001, 79, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Westman, M.E.; Paul, A.; Malik, R.; McDonagh, P.; Ward, M.P.; Hall, E.; Norris, J.M. Seroprevalence of feline immunodeficiency virus and feline leukaemia virus in australia: Risk factors for infection and geographical influences (2011–2013). J. Feline Med. Surg. Open Rep. 2016, 2. [Google Scholar] [CrossRef] [PubMed]
- Reinacher, M. Diseases associated with spontaneous feline leukemia virus (FeLV) infection in cats. Vet. Immunol. Immunopathol. 1989, 21, 85–95. [Google Scholar] [CrossRef]
- Neil, J.C.; Hughes, D.; McFarlane, R.; Wilkie, N.M.; Onions, D.E.; Lees, G.; Jarrett, O. Transduction and rearrangement of the myc gene by feline leukemia-virus in naturally-occurring T-cell leukemias. Nature 1984, 308, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, K. Clinical aspects of feline retroviruses: A review. Viruses 2012, 4, 2684–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatty, J. Viral causes of feline lymphoma: Retroviruses and beyond. Vet. J. 2014, 201, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J.A.; Tasker, S.; Jarrett, O.; Lam, A.; Gibson, S.; Noe-Nordberg, A.; Phillips, A.; Fawcett, A.; Barrs, V.R. Markers of feline leukaemia virus infection or exposure in cats from a region of low seroprevalence. J. Feline Med. Surg. 2011, 13, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Westman, M.E. Epidemiology and Diagnosis of Feline Retroviruses (FIV and FeLV) in Australia and a Trial of Fiv Vaccine Effectiveness in the Field. Ph.D. Thesis, The University of Sydney, Camperdown, NSW, Australia, 2016. [Google Scholar]
- Englert, T.; Lutz, H.; Sauter-Louis, C.; Hartmann, K. Survey of the feline leukemia virus infection status of cats in southern germany. J. Feline Med. Surg. 2012, 14, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Lehmann, R.; Huder, J.B.; Gruber, S.; Boretti, F.; Sigrist, B.; Lutz, H. Feline leukaemia provirus load during the course of experimental infection and in naturally infected cats. J. Gen. Virol. 2001, 82, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Gomes-Keller, M.A.; Gonczi, E.; Tandon, R.; Riondato, F.; Hofmann-Lehmann, R.; Meli, M.L.; Lutz, H. Detection of feline leukemia virus rna in saliva from naturally infected cats and correlation of pcr results with those of current diagnostic methods. J. Clin. Microbiol. 2006, 44, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Pinches, M.D.G.; Helps, C.R.; Gruffydd-Jones, T.J.; Egan, K.; Jarrett, O.; Tasker, S. Diagnosis of feline leukaemia virus infection by semi-quantitative real-time polymerase chain reaction. J. Feline Med. Surg. 2007, 9, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Lehmann, R.; Cattori, V.; Tandon, R.; Boretti, F.S.; Meli, M.L.; Riond, B.; Lutz, H. How molecular methods change our views of felv infection and vaccination. Vet. Immunol. Immunopathol. 2008, 123, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.L.; Haines, D.M.; Meric, S.M.; Misra, V. Feline leukemia virus detection by immunohistochemistry and polymerase chain-reaction in formalin-fixed, paraffin-embedded tumor tissue from cats with lymphosarcoma. Can. J. Vet. Res. 1993, 57, 269–276. [Google Scholar] [PubMed]
- Stutzer, B.; Simon, K.; Lutz, H.; Majzoub, M.; Hermanns, W.; Hirschberger, J.; Sauter-Louis, C.; Hartmann, K. Incidence of persistent viraemia and latent feline leukaemia virus infection in cats with lymphoma. J. Feline Med. Surg. 2011, 13, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Suntz, M.; Failing, K.; Hecht, W.; Schwartz, D.; Reinacher, M. High prevalence of non-productive felv infection in necropsied cats and significant association with pathological findings. Vet. Immunol. Immunopathol. 2010, 136, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Museux, K.; Boretti, F.S.; Willi, B.; Riond, B.; Hoelzle, K.; Hoelzle, L.E.; Wittenbrink, M.M.; Tasker, S.; Wengi, N.; Reusch, C.E.; et al. In Vivo transmission studies of ‘candidatus mycoplasma turicensis’ in the domestic cat. Vet. Res. 2009, 40, 45. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Total | FcaGHV1 Positive (%) | |
|---|---|---|---|
| Lymphoma cases | FFPE lymphoma tissue | 67 | 10 (14.9) |
| Blood | 93 | 14 (15.1) | |
| Controls | FFPE lymph node | 19 | 2 (10.5) |
| Blood | 52 | 9 (17.3) |
| Lymphoma Tissue FcaGHV1 Positive | Autologous Tissue FcaGHV1 Positive | Number of Paired Samples |
|---|---|---|
| Yes | Yes | 0 |
| Yes | No | 3 |
| No | Yes | 3 |
| No | No | 19 |
| Treatment Group | FcaGHV1 DNAemic n = 10 (%) | FcaGHV1 Negative n = 51 (%) |
|---|---|---|
| Multiagent chemotherapy | 7 (70.0%) | 43 (84.3%) |
| Prednisolone | 1 (10.0%) | 3 (5.9%) |
| No treatment | 1 (10.0%) | 3 (5.9%) |
| Unknown | 1 (10.0%) | 2 (3.9%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McLuckie, A.J.; Barrs, V.R.; Lindsay, S.; Aghazadeh, M.; Sangster, C.; Beatty, J.A. Molecular Diagnosis of Felis catus Gammaherpesvirus 1 (FcaGHV1) Infection in Cats of Known Retrovirus Status with and without Lymphoma. Viruses 2018, 10, 128. https://doi.org/10.3390/v10030128
McLuckie AJ, Barrs VR, Lindsay S, Aghazadeh M, Sangster C, Beatty JA. Molecular Diagnosis of Felis catus Gammaherpesvirus 1 (FcaGHV1) Infection in Cats of Known Retrovirus Status with and without Lymphoma. Viruses. 2018; 10(3):128. https://doi.org/10.3390/v10030128
Chicago/Turabian StyleMcLuckie, Alicia J., Vanessa R. Barrs, Scott Lindsay, Mahdis Aghazadeh, Cheryl Sangster, and Julia A. Beatty. 2018. "Molecular Diagnosis of Felis catus Gammaherpesvirus 1 (FcaGHV1) Infection in Cats of Known Retrovirus Status with and without Lymphoma" Viruses 10, no. 3: 128. https://doi.org/10.3390/v10030128