The Role of the Polymeric Immunoglobulin Receptor and Secretory Immunoglobulins during Mucosal Infection and Immunity


{kind=link}
{kind=link}
Abstract
:1. Introduction
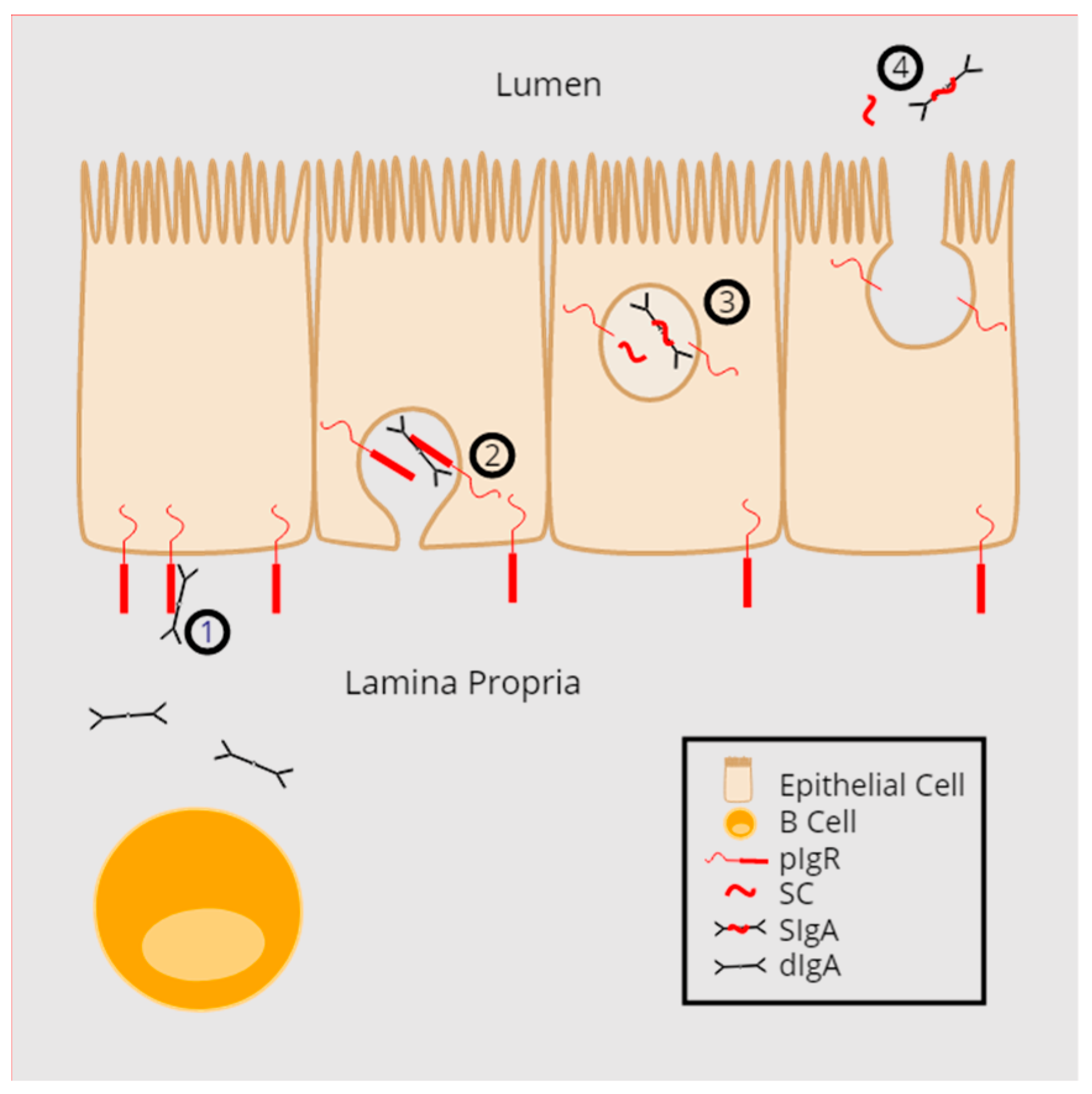
2. pIgR Structure and Function
3. The Multiple Functions of Secretory Component (SC)
4. Regulation of the pIgR/SIg System
5. Lessons from pIgR-Deficient Mice
6. Agglutination and Exclusion of Pathogens from Mucosal Surfaces (Immune Exclusion)
7. Intracellular Neutralization and Excretion of Pathogens
8. SIg-Mediated Immune Modulation during Infection
9. SIg-Induced Complement Activation and Immune Pathology
10. Subversion of the pIgR/SIg System by Pathogens
11. Conclusions
12. Outstanding Questions
- Are pathogenic infections modulated by natural non-specific SIg or SC and do pathogens modulate that response?
- Can non-bacterial mucosal infections be controlled by SIg-mediated agglutination?
- Does intracellular neutralization of viral infections by SIgA and basolateral to apical excretion of SIgA-immune complexes extend to non-viral infections, occur in vivo, and affect within or between host spread?
- What role does SIg-induced complement activation play during infection with mucosal pathogens and colonization of commensals?
- How common are pIgR/SIg subversion or evasion mechanisms among mucosal pathogens from different kingdoms and are the strategies shared or specific?
- What is the role of SIgM-immune complexes during mucosal homeostasis and pathogenesis?
- What is the identity of the SIg receptor on M cells?
- What breakthroughs will the future hold when organoid technology is applied to the study of the pIgR/SIg cycle?
Acknowledgments
Conflicts of Interest
References
- Bunker, J.J.; Erickson, S.A.; Flynn, T.M.; Henry, C.; Koval, J.C.; Meisel, M.; Jabri, B.; Antonopoulos, D.A.; Wilson, P.C.; Bendelac, A.; et al. Natural polyreactive IgA antibodies coat the intestinal microbiota. Science 2017, 358. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, A.J.; Gatto, D.; Sainsbury, E.; Harriman, G.R.; Hengartner, H.; Zinkernagel, R.M. A primitive T cell-independent mechanism of intestinal mucosal IgA responses to commensal bacteria. Science 2000, 288, 2222–2226. [Google Scholar] [CrossRef] [PubMed]
- Quan, C.P.; Berneman, A.; Pires, R.; Avrameas, S.; Bouvet, J.P. Natural polyreactive secretory immunoglobulin A autoantibodies as a possible barrier to infection in humans. Infect. Immun. 1997, 65, 3997–4004. [Google Scholar] [PubMed]
- Pabst, O.; Cerovic, V.; Hornef, M. Secretory IgA in the Coordination of Establishment and Maintenance of the Microbiota. Trends Immunol. 2016, 37, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Mantis, N.J.; Cheung, M.C.; Chintalacharuvu, K.R.; Rey, J.; Corthesy, B.; Neutra, M.R. Selective adherence of IgA to murine Peyer’s patch M cells: Evidence for a novel IgA receptor. J. Immunol. 2002, 169, 1844–1851. [Google Scholar] [CrossRef] [PubMed]
- Stadtmueller, B.M.; Huey-Tubman, K.E.; Lopez, C.J.; Yang, Z.; Hubbell, W.L.; Bjorkman, P.J. The structure and dynamics of secretory component and its interactions with polymeric immunoglobulins. eLife 2016, 5, e10640. [Google Scholar] [CrossRef] [PubMed]
- Asano, M.; Komiyama, K. Polymeric immunoglobulin receptor. J. Oral. Sci. 2011, 53, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Schneeman, T.A.; Bruno, M.E.; Schjerven, H.; Johansen, F.E.; Chady, L.; Kaetzel, C.S. Regulation of the polymeric Ig receptor by signaling through TLRs 3 and 4: Linking innate and adaptive immune responses. J. Immunol. 2005, 175, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Mikami, Y.; Iwase, T.; Komiyama, Y.; Matsumoto, N.; Oki, H.; Komiyama, K. Secretory leukocyte protease inhibitor inhibits expression of polymeric immunoglobulin receptor via the NF-κB signaling pathway. Mol. Immunol. 2015, 67, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Kaetzel, C.S.; Robinson, J.K.; Chintalacharuvu, K.R.; Vaerman, J.P.; Lamm, M.E. The polymeric immunoglobulin receptor (secretory component) mediates transport of immune complexes across epithelial cells: A local defense function for IgA. Proc. Natl. Acad. Sci. USA 1991, 88, 8796–8800. [Google Scholar] [CrossRef] [PubMed]
- Johansen, F.E.; Braathen, R.; Brandtzaeg, P. The J chain is essential for polymeric Ig receptor-mediated epithelial transport of IgA. J. Immunol. 2001, 167, 5185–5192. [Google Scholar] [CrossRef] [PubMed]
- Bakos, M.A.; Kurosky, A.; Goldblum, R.M. Characterization of a critical binding site for human polymeric Ig on secretory component. J. Immunol. 1991, 147, 3419–3426. [Google Scholar] [PubMed]
- Pilette, C.; Ouadrhiri, Y.; Dimanche, F.; Vaerman, J.P.; Sibille, Y. Secretory component is cleaved by neutrophil serine proteinases but its epithelial production is increased by neutrophils through NF-κ B- and p38 mitogen-activated protein kinase-dependent mechanisms. Am. J. Respir. Cell Mol. Biol. 2003, 28, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.; Kaetzel, C.S. Secretory IgA is Concentrated in the Outer Layer of Colonic Mucus along with Gut Bacteria. Pathogens 2014, 3, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Obara, W.; Iida, A.; Suzuki, Y.; Tanaka, T.; Akiyama, F.; Maeda, S.; Ohnishi, Y.; Yamada, R.; Tsunoda, T.; Takei, T.; et al. Association of single-nucleotide polymorphisms in the polymeric immunoglobulin receptor gene with immunoglobulin A nephropathy (IgAN) in Japanese patients. J. Hum. Genet. 2003, 48, 293–299. [Google Scholar] [PubMed]
- Phalipon, A.; Corthesy, B. Novel functions of the polymeric Ig receptor: Well beyond transport of immunoglobulins. Trends Immunol. 2003, 24, 55–58. [Google Scholar] [CrossRef]
- Kaetzel, C.S. The polymeric immunoglobulin receptor: Bridging innate and adaptive immune responses at mucosal surfaces. Immunol. Rev. 2005, 206, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Lullau, E.; Heyse, S.; Vogel, H.; Marison, I.; von Stockar, U.; Kraehenbuhl, J.P.; Corthesy, B. Antigen binding properties of purified immunoglobulin A and reconstituted secretory immunoglobulin A antibodies. J. Biol. Chem. 1996, 271, 16300–16309. [Google Scholar] [CrossRef] [PubMed]
- Phalipon, A.; Cardona, A.; Kraehenbuhl, J.P.; Edelman, L.; Sansonetti, P.J.; Corthesy, B. Secretory component: A new role in secretory IgA-mediated immune exclusion in vivo. Immunity 2002, 17, 107–115. [Google Scholar] [CrossRef]
- Duc, M.; Johansen, F.E.; Corthesy, B. Antigen binding to secretory immunoglobulin A results in decreased sensitivity to intestinal proteases and increased binding to cellular Fc receptors. J. Biol. Chem. 2010, 285, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Crottet, P.; Corthesy, B. Secretory component delays the conversion of secretory IgA into antigen-binding competent F(ab’)2: A possible implication for mucosal defense. J. Immunol. 1998, 161, 5445–5453. [Google Scholar] [PubMed]
- Fasching, C.E.; Grossman, T.; Corthesy, B.; Plaut, A.G.; Weiser, J.N.; Janoff, E.N. Impact of the molecular form of immunoglobulin a on functional activity in defense against Streptococcus pneumoniae. Infect. Immun. 2007, 75, 1801–1810. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Guerrero, A.; Parker, E.; Strum, J.S.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Site-specific glycosylation of secretory immunoglobulin A from human colostrum. J. Proteom. Res. 2015, 14, 1335–1349. [Google Scholar] [CrossRef] [PubMed]
- Pierce-Cretel, A.; Pamblanco, M.; Strecker, G.; Montreuil, J.; Spik, G.; Dorland, L.; Van Halbeek, H.; Vliegenthart, J.F. Primary structure of the N-glycosidically linked sialoglycans of secretory immunoglobulins A from human milk. Eur. J. Biochem. 1982, 125, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Asano, M.; Ogura, Y.; Takenouchi-Ohkubo, N.; Chihaya, H.; Chung-Hsing, W.; Ishikawa, K.; Zhu, L.; Moro, I. Release of non-glycosylated polymeric immunoglobulin receptor protein. Scand. J. Immunol. 2003, 58, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Mathias, A.; Corthesy, B. Recognition of gram-positive intestinal bacteria by hybridoma- and colostrum-derived secretory immunoglobulin A is mediated by carbohydrates. J. Biol. Chem. 2011, 286, 17239–17247. [Google Scholar] [CrossRef] [PubMed]
- Dallas, S.D.; Rolfe, R.D. Binding of Clostridium difficile toxin A to human milk secretory component. J. Med. Microbiol. 1998, 47, 879–888. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, I.R.; de Araujo, A.N.; Bao, S.N.; Giugliano, L.G. Binding of lactoferrin and free secretory component to enterotoxigenic Escherichia coli. FEMS Microbiol. Lett. 2001, 203, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Motegi, Y.; Kita, H. Interaction with secretory component stimulates effector functions of human eosinophils but not of neutrophils. J. Immunol. 1998, 161, 4340–4346. [Google Scholar] [PubMed]
- Marshall, L.J.; Perks, B.; Ferkol, T.; Shute, J.K. IL-8 released constitutively by primary bronchial epithelial cells in culture forms an inactive complex with secretory component. J. Immunol. 2001, 167, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Kaetzel, C.S. Cooperativity among secretory IgA, the polymeric immunoglobulin receptor, and the gut microbiota promotes host-microbial mutualism. Immunol. Lett. 2014, 162, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.E.; Frantz, A.L.; Rogier, E.W.; Johansen, F.E.; Kaetzel, C.S. Regulation of the polymeric immunoglobulin receptor by the classical and alternative NF-κB pathways in intestinal epithelial cells. Mucosal Immunol. 2011, 4, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.E.; Rogier, E.W.; Frantz, A.L.; Stefka, A.T.; Thompson, S.N.; Kaetzel, C.S. Regulation of the polymeric immunoglobulin receptor in intestinal epithelial cells by Enterobacteriaceae: Implications for mucosal homeostasis. Immunol. Investig. 2010, 39, 356–382. [Google Scholar] [CrossRef] [PubMed]
- Frantz, A.L.; Rogier, E.W.; Weber, C.R.; Shen, L.; Cohen, D.A.; Fenton, L.A.; Bruno, M.E.; Kaetzel, C.S. Targeted deletion of MyD88 in intestinal epithelial cells results in compromised antibacterial immunity associated with downregulation of polymeric immunoglobulin receptor, mucin-2, and antibacterial peptides. Mucosal Immunol. 2012, 5, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Blanch, V.J.; Piskurich, J.F.; Kaetzel, C.S. Cutting edge: Coordinate regulation of IFN regulatory factor-1 and the polymeric Ig receptor by proinflammatory cytokines. J. Immunol. 1999, 162, 1232–1235. [Google Scholar] [PubMed]
- Moon, C.; VanDussen, K.L.; Miyoshi, H.; Stappenbeck, T.S. Development of a primary mouse intestinal epithelial cell monolayer culture system to evaluate factors that modulate IgA transcytosis. Mucosal Immunol. 2014, 7, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Kvale, D.; Brandtzaeg, P. Butyrate differentially affects constitutive and cytokine-induced expression of HLA molecules, secretory component (SC), and ICAM-1 in a colonic epithelial cell line (HT-29, clone m3). In Advances in Mucosal Immunology; Springer: Boston, MA, USA, 1995; pp. 183–188. [Google Scholar]
- Pal, K.; Kaetzel, C.S.; Brundage, K.; Cunningham, C.A.; Cuff, C.F. Regulation of polymeric immunoglobulin receptor expression by reovirus. J. Gen. Virol. 2005, 86, 2347–23457. [Google Scholar] [CrossRef] [PubMed]
- Armitage, C.W.; O’Meara, C.P.; Beagley, K.W. Chlamydial infection enhances expression of the polymeric immunoglobulin receptor (pIgR) and transcytosis of IgA. Am. J. Reprod. Immunol. 2017, 77, e12611. [Google Scholar] [CrossRef] [PubMed]
- Godinez-Victoria, M.; Cruz-Hernandez, T.R.; Reyna-Garfias, H.; Barbosa-Cabrera, R.E.; Drago-Serrano, M.E.; Sanchez-Gomez, M.C.; Campos-Rodriguez, R. Modulation by bovine lactoferrin of parameters associated with the IgA response in the proximal and distal small intestine of BALB/c mice. Immunopharmacol. Immunotoxicol. 2017, 39, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Rincheval-Arnold, A.; Belair, L.; Djiane, J. Developmental expression of pIgR gene in sheep mammary gland and hormonal regulation. J. Dairy Res. 2002, 69, 13–26. [Google Scholar] [PubMed]
- Godinez-Victoria, M.; Campos-Rodriguez, R.; Rivera-Aguilar, V.; Lara-Padilla, E.; Pacheco-Yepez, J.; Jarillo-Luna, R.A.; Drago-Serrano, M.E. Intermittent fasting promotes bacterial clearance and intestinal IgA production in Salmonella typhimurium-infected mice. Scand. J. Immunol. 2014, 79, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Kurimoto, Y.; Saruta, J.; To, M.; Yamamoto, Y.; Kimura, K.; Tsukinoki, K. Voluntary exercise increases IgA concentration and polymeric Ig receptor expression in the rat submandibular gland. Biosci. Biotechnol. Biochem. 2016, 80, 2490–2496. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.C. Chronic alcohol consumption regulates the expression of poly immunoglobulin receptor (pIgR) and secretory IgA in the gut. Toxicol. Appl. Pharmacol. 2017, 333, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Rusznak, C.; Sapsford, R.J.; Devalia, J.L.; Wang, J.H.; Shah, S.S.; Mills, P.R.; Davies, R.J.; Lozewicz, S. Cigarette smoke decreases the expression of secretory component in human bronchial epithelial cells, in vitro. Acta Microbiol. Immunol. Hung. 2001, 48, 81–94. [Google Scholar] [PubMed]
- Qi, X.; Li, X.; Sun, X. Reduced expression of polymeric immunoglobulin receptor (pIgR) in nasopharyngeal carcinoma and its correlation with prognosis. Tumour. Biol. 2016, 37, 11099–11104. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, P.; Bhattacharya, S.; Chin-Aleong, J.; Capaso, M.; Kocher, H.M. Expression of polymeric immunoglobulin receptor and stromal activity in pancreatic ductal adenocarcinoma. Pancreatology 2017, 17, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Ai, J.; Xu, Y.; Chen, Y.; Huang, M.; Yang, X.; Hu, B.; Zhang, H.; He, C.; Yang, X.; et al. Polymeric immunoglobulin receptor promotes tumor growth in hepatocellular carcinoma. Hepatology 2017, 65, 1948–1962. [Google Scholar] [CrossRef] [PubMed]
- Shimada, S.; Kawaguchi-Miyashita, M.; Kushiro, A.; Sato, T.; Nanno, M.; Sako, T.; Matsuoka, Y.; Sudo, K.; Tagawa, Y.; Iwakura, Y.; et al. Generation of polymeric immunoglobulin receptor-deficient mouse with marked reduction of secretory IgA. J. Immunol. 1999, 163, 5367–5373. [Google Scholar] [PubMed]
- Johansen, F.E.; Pekna, M.; Norderhaug, I.N.; Haneberg, B.; Hietala, M.A.; Krajci, P.; Betsholtz, C.; Brandtzaeg, P. Absence of epithelial immunoglobulin A transport, with increased mucosal leakiness, in polymeric immunoglobulin receptor/secretory component-deficient mice. J. Exp. Med. 1999, 190, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Uren, T.K.; Johansen, F.E.; Wijburg, O.L.; Koentgen, F.; Brandtzaeg, P.; Strugnell, R.A. Role of the polymeric Ig receptor in mucosal B cell homeostasis. J. Immunol. 2003, 170, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Turula, H.; Bragazzi-Cunha, J.; Ramakrishnan, S.; Wilke, C.; Gonzalez-Hernandez, M.; Pry, A.; Fava, J.; Svoboda, S.; Shah, Y.; Corthesy, B.; et al. Natural Secretory Immunoglobulins Enhance Norovirus Infection. bioRxiv 2018, 253286. [Google Scholar] [CrossRef]
- Yamazaki, K.; Shimada, S.; Kato-Nagaoka, N.; Soga, H.; Itoh, T.; Nanno, M. Accumulation of intestinal intraepithelial lymphocytes in association with lack of polymeric immunoglobulin receptor. Eur. J. Immunol. 2005, 35, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Richmond, B.W.; Du, R.H.; Han, W.; Benjamin, J.T.; Meer, R.V.; Gleaves, L.; Guo, M.; McKissack, A.; Zhang, Y.; Cheng, D.S.; et al. Bacterial-derived Neutrophilic Inflammation Drives Lung Remodeling in a Mouse Model of COPD. Am. J. Respir. Cell Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Sait, L.; Galic, M.; Strugnell, R.A.; Janssen, P.H. Secretory antibodies do not affect the composition of the bacterial microbiota in the terminal ileum of 10-week-old mice. Appl. Environ. Microbiol. 2003, 69, 2100–2109. [Google Scholar] [CrossRef] [PubMed]
- Van de Perre, P. Transfer of antibody via mother’s milk. Vaccine 2003, 21, 3374–3376. [Google Scholar] [CrossRef]
- Reikvam, D.H.; Derrien, M.; Islam, R.; Erofeev, A.; Grcic, V.; Sandvik, A.; Gaustad, P.; Meza-Zepeda, L.A.; Jahnsen, F.L.; Smidt, H.; et al. Epithelial-microbial crosstalk in polymeric Ig receptor deficient mice. Eur. J. Immunol. 2012, 42, 2959–2970. [Google Scholar] [CrossRef] [PubMed]
- Kato-Nagaoka, N.; Shimada, S.; Yamakawa, Y.; Tsujibe, S.; Naito, T.; Setoyama, H.; Watanabe, Y.; Shida, K.; Matsumoto, S.; Nanno, M. Enhanced differentiation of intraepithelial lymphocytes in the intestine of polymeric immunoglobulin receptor-deficient mice. Immunology 2015, 146, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Polosukhin, V.V.; Cates, J.M.; Lawson, W.E.; Zaynagetdinov, R.; Milstone, A.P.; Massion, P.P.; Ocak, S.; Ware, L.B.; Lee, J.W.; Bowler, R.P.; et al. Bronchial secretory immunoglobulin a deficiency correlates with airway inflammation and progression of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2011, 184, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Gohy, S.T.; Detry, B.R.; Lecocq, M.; Bouzin, C.; Weynand, B.A.; Amatngalim, G.D.; Sibille, Y.M.; Pilette, C. Polymeric immunoglobulin receptor down-regulation in chronic obstructive pulmonary disease. Persistence in the cultured epithelium and role of transforming growth factor-beta. Am. J. Respir. Crit. Care Med. 2014, 190, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Richmond, B.W.; Brucker, R.M.; Han, W.; Du, R.H.; Zhang, Y.; Cheng, D.S.; Gleaves, L.; Abdolrasulnia, R.; Polosukhina, D.; Clark, P.E.; et al. Airway bacteria drive a progressive COPD-like phenotype in mice with polymeric immunoglobulin receptor deficiency. Nat. Commun. 2016, 7, e11240. [Google Scholar] [CrossRef] [PubMed]
- Frantz, A.L.; Bruno, M.E.; Rogier, E.W.; Tuna, H.; Cohen, D.A.; Bondada, S.; Chelvarajan, R.L.; Brandon, J.A.; Jennings, C.D.; Kaetzel, C.S. Multifactorial patterns of gene expression in colonic epithelial cells predict disease phenotypes in experimental colitis. Inflamm. Bowel Dis. 2012, 18, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Arsenescu, R.; Bruno, M.E.; Rogier, E.W.; Stefka, A.T.; McMahan, A.E.; Wright, T.B.; Nasser, M.S.; de Villiers, W.J.; Kaetzel, C.S. Signature biomarkers in Crohn’s disease: Toward a molecular classification. Mucosal Immunol. 2008, 1, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Bruno, M.E.; Rogier, E.W.; Arsenescu, R.I.; Flomenhoft, D.R.; Kurkjian, C.J.; Ellis, G.I.; Kaetzel, C.S. Correlation of Biomarker Expression in Colonic Mucosa with Disease Phenotype in Crohn's Disease and Ulcerative Colitis. Dig. Dis. Sci. 2015, 60, 2976–2984. [Google Scholar] [CrossRef] [PubMed]
- Wijburg, O.L.; Uren, T.K.; Simpfendorfer, K.; Johansen, F.E.; Brandtzaeg, P.; Strugnell, R.A. Innate secretory antibodies protect against natural Salmonella typhimurium infection. J. Exp. Med. 2006, 203, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Davids, B.J.; Palm, J.E.; Housley, M.P.; Smith, J.R.; Andersen, Y.S.; Martin, M.G.; Hendrickson, B.A.; Johansen, F.E.; Svard, S.G.; Gillin, F.D.; et al. Polymeric immunoglobulin receptor in intestinal immune defense against the lumen-dwelling protozoan parasite Giardia. J. Immunol. 2006, 177, 6281–6290. [Google Scholar] [CrossRef] [PubMed]
- Testerman, T.L.; Morris, J. Beyond the stomach: An updated view of Helicobacter pylori pathogenesis, diagnosis, and treatment. World J. Gastroenterol. 2014, 20, 12781–12808. [Google Scholar] [CrossRef] [PubMed]
- Gorrell, R.J.; Wijburg, O.L.; Pedersen, J.S.; Walduck, A.K.; Kwok, T.; Strugnell, R.A.; Robins-Browne, R.M. Contribution of secretory antibodies to intestinal mucosal immunity against Helicobacter pylori. Infect. Immun. 2013, 81, 3880–3893. [Google Scholar] [CrossRef] [PubMed]
- Falk, P.; Roth, K.A.; Boren, T.; Westblom, T.U.; Gordon, J.I.; Normark, S. An in vitro adherence assay reveals that Helicobacter pylori exhibits cell lineage-specific tropism in the human gastric epithelium. Proc. Natl. Acad. Sci. USA 1993, 90, 2035–2039. [Google Scholar] [CrossRef] [PubMed]
- Ahlstedt, I.; Lindholm, C.; Lonroth, H.; Hamlet, A.; Svennerholm, A.M.; Quiding-Jarbrink, M. Role of local cytokines in increased gastric expression of the secretory component in Helicobacter pylori infection. Infect. Immun. 1999, 67, 4921–4925. [Google Scholar] [PubMed]
- Kaneko, T.; Ota, H.; Hayama, M.; Akamatsu, T.; Katsuyama, T. Helicobacter pylori infection produces expression of a secretory component in gastric mucous cells. Virchows Arch. 2000, 437, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Fubara, E.S.; Freter, R. Protection against enteric bacterial infection by secretory IgA antibodies. J. Immunol. 1973, 111, 395–403. [Google Scholar] [PubMed]
- Tokuhara, D.; Yuki, Y.; Nochi, T.; Kodama, T.; Mejima, M.; Kurokawa, S.; Takahashi, Y.; Nanno, M.; Nakanishi, U.; Takaiwa, F.; et al. Secretory IgA-mediated protection against V. cholerae and heat-labile enterotoxin-producing enterotoxigenic Escherichia coli by rice-based vaccine. Proc. Natl. Acad. Sci. USA 2010, 107, 8794–8799. [Google Scholar] [CrossRef] [PubMed]
- Winner, L., 3rd; Mack, J.; Weltzin, R.; Mekalanos, J.J.; Kraehenbuhl, J.P.; Neutra, M.R. New model for analysis of mucosal immunity: Intestinal secretion of specific monoclonal immunoglobulin A from hybridoma tumors protects against Vibrio cholerae infection. Infect. Immun. 1991, 59, 977–982. [Google Scholar] [PubMed]
- Boullier, S.; Tanguy, M.; Kadaoui, K.A.; Caubet, C.; Sansonetti, P.; Corthesy, B.; Phalipon, A. Secretory IgA-mediated neutralization of Shigella flexneri prevents intestinal tissue destruction by down-regulating inflammatory circuits. J. Immunol. 2009, 183, 5879–5885. [Google Scholar] [CrossRef] [PubMed]
- Mathias, A.; Longet, S.; Corthesy, B. Agglutinating secretory IgA preserves intestinal epithelial cell integrity during apical infection by Shigella flexneri. Infect. Immun. 2013, 81, 3027–3034. [Google Scholar] [CrossRef] [PubMed]
- Longet, S.; Vonarburg, C.; Lotscher, M.; Miescher, S.; Zuercher, A.; Corthesy, B. Reconstituted human polyclonal plasma-derived secretory-like IgM and IgA maintain the barrier function of epithelial cells infected with an enteropathogen. J. Biol. Chem. 2014, 289, 21617–21626. [Google Scholar] [CrossRef] [PubMed]
- Armitage, C.W.; O'Meara, C.P.; Harvie, M.C.; Timms, P.; Wijburg, O.L.; Beagley, K.W. Evaluation of intra- and extra-epithelial secretory IgA in chlamydial infections. Immunology 2014, 143, 520–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrier, C.; Sprenger, N.; Corthesy, B. Glycans on secretory component participate in innate protection against mucosal pathogens. J. Biol. Chem. 2006, 281, 14280–14287. [Google Scholar] [CrossRef] [PubMed]
- Wold, A.E.; Mestecky, J.; Tomana, M.; Kobata, A.; Ohbayashi, H.; Endo, T.; Eden, C.S. Secretory immunoglobulin A carries oligosaccharide receptors for Escherichia coli type 1 fimbrial lectin. Infect. Immun. 1990, 58, 3073–3077. [Google Scholar] [PubMed]
- Murthy, A.K.; Chaganty, B.K.; Troutman, T.; Guentzel, M.N.; Yu, J.J.; Ali, S.K.; Lauriano, C.M.; Chambers, J.P.; Klose, K.E.; Arulanandam, B.P. Mannose-containing oligosaccharides of non-specific human secretory immunoglobulin A mediate inhibition of Vibrio cholerae biofilm formation. PLoS ONE 2011, 6, e16847. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, A.P.; Top, J.; Bayjanov, J.R.; Kemperman, H.; Rogers, M.R.; Paganelli, F.L.; Bonten, M.J.; Willems, R.J. Antibiotic-Driven Dysbiosis Mediates Intraluminal Agglutination and Alternative Segregation of Enterococcus faecium from the Intestinal Epithelium. MBio 2015, 6, e01346-15. [Google Scholar] [CrossRef] [PubMed]
- Bioley, G.; Monnerat, J.; Lotscher, M.; Vonarburg, C.; Zuercher, A.; Corthesy, B. Plasma-Derived Polyreactive Secretory-Like IgA and IgM Opsonizing Salmonella enterica Typhimurium Reduces Invasion and Gut Tissue Inflammation through Agglutination. Front. Immunol. 2017, 8, e1043. [Google Scholar] [CrossRef] [PubMed]
- Mazanec, M.B.; Kaetzel, C.S.; Lamm, M.E.; Fletcher, D.; Nedrud, J.G. Intracellular neutralization of virus by immunoglobulin A antibodies. Proc. Natl. Acad. Sci. USA 1992, 89, 6901–6905. [Google Scholar] [CrossRef] [PubMed]
- Mazanec, M.B.; Coudret, C.L.; Fletcher, D.R. Intracellular neutralization of influenza virus by immunoglobulin A anti-hemagglutinin monoclonal antibodies. J. Virol. 1995, 69, 1339–1343. [Google Scholar] [PubMed]
- Yan, H.; Lamm, M.E.; Bjorling, E.; Huang, Y.T. Multiple functions of immunoglobulin A in mucosal defense against viruses: An in vitro measles virus model. J. Virol. 2002, 76, 10972–10979. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.; Yan, H.; Lamm, M.E.; Huang, Y.T. Immunoglobulin A antibodies against internal HIV-1 proteins neutralize HIV-1 replication inside epithelial cells. Virology 2006, 356, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Corthesy, B.; Benureau, Y.; Perrier, C.; Fourgeux, C.; Parez, N.; Greenberg, H.; Schwartz-Cornil, I. Rotavirus anti-VP6 secretory immunoglobulin A contributes to protection via intracellular neutralization but not via immune exclusion. J. Virol. 2006, 80, 10692–10699. [Google Scholar] [CrossRef] [PubMed]
- Ruggeri, F.M.; Johansen, K.; Basile, G.; Kraehenbuhl, J.P.; Svensson, L. Antirotavirus immunoglobulin A neutralizes virus in vitro after transcytosis through epithelial cells and protects infant mice from diarrhea. J. Virol. 1998, 72, 2708–2714. [Google Scholar] [PubMed]
- Wright, A.; Lamm, M.E.; Huang, Y.T. Excretion of human immunodeficiency virus type 1 through polarized epithelium by immunoglobulin A. J. Virol. 2008, 82, 11526–11535. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.W.; Siadat-Pajouh, M.; Krishnaney, A.A.; Greenberg, H.B. Protective effect of rotavirus VP6-specific IgA monoclonal antibodies that lack neutralizing activity. Science 1996, 272, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Cornil, I.; Benureau, Y.; Greenberg, H.; Hendrickson, B.A.; Cohen, J. Heterologous protection induced by the inner capsid proteins of rotavirus requires transcytosis of mucosal immunoglobulins. J. Virol. 2002, 76, 8110–8117. [Google Scholar] [CrossRef] [PubMed]
- Rey, J.; Garin, N.; Spertini, F.; Corthesy, B. Targeting of secretory IgA to Peyer’s patch dendritic and T cells after transport by intestinal M cells. J. Immunol. 2004, 172, 3026–3033. [Google Scholar] [CrossRef] [PubMed]
- Kadaoui, K.A.; Corthesy, B. Secretory IgA mediates bacterial translocation to dendritic cells in mouse Peyer’s patches with restriction to mucosal compartment. J. Immunol. 2007, 179, 7751–7757. [Google Scholar] [CrossRef] [PubMed]
- Diana, J.; Moura, I.C.; Vaugier, C.; Gestin, A.; Tissandie, E.; Beaudoin, L.; Corthesy, B.; Hocini, H.; Lehuen, A.; Monteiro, R.C. Secretory IgA induces tolerogenic dendritic cells through SIGNR1 dampening autoimmunity in mice. J. Immunol. 2013, 191, 2335–2343. [Google Scholar] [CrossRef] [PubMed]
- Baumann, J.; Park, C.G.; Mantis, N.J. Recognition of secretory IgA by DC-SIGN: Implications for immune surveillance in the intestine. Immunol. Lett. 2010, 131, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Mikulic, J.; Bioley, G.; Corthesy, B. SIgA-Shigella Immune Complexes Interact with Dectin-1 and SIGNR3 to Differentially Regulate Mouse Peyer’s Patch and Mesenteric Lymph Node Dendritic Cell’s Responsiveness. J. Mol. Biol. 2017, 429, 2387–2400. [Google Scholar] [CrossRef] [PubMed]
- Hansen, I.S.; Hoepel, W.; Zaat, S.A.J.; Baeten, D.L.P.; den Dunnen, J. Serum IgA Immune Complexes Promote Proinflammatory Cytokine Production by Human Macrophages, Monocytes, and Kupffer Cells through FcalphaRI-TLR Cross-Talk. J. Immunol. 2017, 199, 4124–4131. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.L.; Suscovich, T.J.; Fortune, S.M.; Alter, G. Beyond binding: Antibody effector functions in infectious diseases. Nat. Rev. Immunol. 2018, 18, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, T.E.; Emilsen, S.; Sandin, R.H.; Granerud, B.K.; Bratlie, D.; Ihle, O.; Sandlie, I. Human Secretory IgM Antibodies Activate Human Complement and Offer Protection at Mucosal Surface. Scand. J. Immunol. 2017, 85, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, B.A.; Guo, J.; Brown, I.; Dennis, K.; Marcellino, D.; Hetzel, J.; Herold, B.C. Decreased vaginal disease in J-chain-deficient mice following herpes simplex type 2 genital infection. Virology 2000, 271, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Salerno-Goncalves, R.; Safavie, F.; Fasano, A.; Sztein, M.B. Free and complexed-secretory immunoglobulin A triggers distinct intestinal epithelial cell responses. Clin. Exp. Immunol. 2016, 185, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Church, D.; Vanderkooi, O.G.; Low, D.E.; Pillai, D.R. Streptococcus pneumoniae infection: A Canadian perspective. Expert Rev. Anti-Infect. Ther. 2013, 11, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, S.; Talay, S.R.; Brandtzaeg, P.; Chhatwal, G.S. SpsA, a novel pneumococcal surface protein with specific binding to secretory immunoglobulin A and secretory component. Mol. Microbiol. 1997, 25, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.R.; Mostov, K.E.; Lamm, M.E.; Nanno, M.; Shimida, S.; Ohwaki, M.; Tuomanen, E. The polymeric immunoglobulin receptor translocates pneumococci across human nasopharyngeal epithelial cells. Cell 2000, 102, 827–837. [Google Scholar] [CrossRef]
- Asmat, T.M.; Agarwal, V.; Rath, S.; Hildebrandt, J.P.; Hammerschmidt, S. Streptococcus pneumoniae infection of host epithelial cells via polymeric immunoglobulin receptor transiently induces calcium release from intracellular stores. J. Biol. Chem. 2011, 286, 17861–17869. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Johansen, F.E.; Eckmann, L.; Metzger, D.W. An important role for polymeric Ig receptor-mediated transport of IgA in protection against Streptococcus pneumoniae nasopharyngeal carriage. J. Immunol. 2004, 173, 4576–4581. [Google Scholar] [CrossRef] [PubMed]
- Boehme, J.D.; Stegemann-Koniszewski, S.; Autengruber, A.; Peters, N.; Wissing, J.; Jansch, L.; Jeron, A.; Bruder, D. Chronic lung inflammation primes humoral immunity and augments antipneumococcal resistance. Sci. Rep. 2017, 7, 4972. [Google Scholar] [CrossRef] [PubMed]
- ovino, F.; Engelen-Lee, J.Y.; Brouwer, M.; van de Beek, D.; van der Ende, A.; Valls Seron, M.; Mellroth, P.; Muschiol, S.; Bergstrand, J.; Widengren, J.; et al. pIgR and PECAM-1 bind to pneumococcal adhesins RrgA and PspC mediating bacterial brain invasion. J. Exp. Med. 2017, 214, 1619–1630. [Google Scholar]
- Poulain, D. Candida albicans, plasticity and pathogenesis. Crit. Rev. Microbiol. 2015, 41, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.R.; Bandara, B.M.; Cannon, R.D. Saliva promotes Candida albicans adherence to human epithelial cells. J. Dent. Res. 2002, 81, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Van der Wielen, P.A.; Holmes, A.R.; Cannon, R.D. Secretory component mediates Candida albicans binding to epithelial cells. Oral. Dis. 2016, 22, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Vetsika, E.K.; Callan, M. Infectious mononucleosis and Epstein-Barr virus. Expert Rev. Mol. Med. 2004, 6, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sixbey, J.W.; Yao, Q.Y. Immunoglobulin A-induced shift of Epstein-Barr virus tissue tropism. Science 1992, 255, 1578–1580. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Lin, C.R.; Tan, G.K.; Chen, W.; Dee, A.N.; Chan, W.Y. The mechanism of Epstein-Barr virus infection in nasopharyngeal carcinoma cells. Am. J. Pathol. 1997, 150, 1745–1756. [Google Scholar] [PubMed]
- Gan, Y.J.; Chodosh, J.; Morgan, A.; Sixbey, J.W. Epithelial cell polarization is a determinant in the infectious outcome of immunoglobulin A-mediated entry by Epstein-Barr virus. J. Virol. 1997, 71, 519–526. [Google Scholar] [PubMed]
- Liu, G.; Ren, W.; Fang, J.; Hu, C.A.; Guan, G.; Al-Dhabi, N.A.; Yin, J.; Duraipandiyan, V.; Chen, S.; Peng, Y.; et al. l-Glutamine and l-arginine protect against enterotoxigenic Escherichia coli infection via intestinal innate immunity in mice. Amino Acids 2017, 49, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, F.J.; Yu, L.; Yao, W.R.; Cui, Y.F.; Yang, G.B. Expression of pIgR in the tracheal mucosa of SHIV/SIV-infected rhesus macaques. Zool Res. 2017, 38, 44–48. [Google Scholar] [PubMed]
- Wang, Y.; Yang, G.B. Alteration of Polymeric Immunoglobulin Receptor and Neonatal Fc Receptor Expression in the Gut Mucosa of Immunodeficiency Virus-Infected Rhesus Macaques. Scand. J. Immunol. 2016, 83, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Aurora, M.; Spence, J.R. hPSC-derived lung and intestinal organoids as models of human fetal tissue. Dev. Biol. 2016, 420, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Noel, G.; Baetz, N.W.; Staab, J.F.; Donowitz, M.; Kovbasnjuk, O.; Pasetti, M.F.; Zachos, N.C. A primary human macrophage-enteroid co-culture model to investigate mucosal gut physiology and host-pathogen interactions. Sci. Rep. 2017, 7, 45270. [Google Scholar] [CrossRef] [PubMed]
- Skardal, A.; Murphy, S.V.; Devarasetty, M.; Mead, I.; Kang, H.W.; Seol, Y.J.; Shrike Zhang, Y.; Shin, S.R.; Zhao, L.; Aleman, J.; et al. Multi-tissue interactions in an integrated three-tissue organ-on-a-chip platform. Sci. Rep. 2017, 7, 8837. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turula, H.; Wobus, C.E. The Role of the Polymeric Immunoglobulin Receptor and Secretory Immunoglobulins during Mucosal Infection and Immunity. Viruses 2018, 10, 237. https://doi.org/10.3390/v10050237
Turula H, Wobus CE. The Role of the Polymeric Immunoglobulin Receptor and Secretory Immunoglobulins during Mucosal Infection and Immunity. Viruses. 2018; 10(5):237. https://doi.org/10.3390/v10050237
Chicago/Turabian StyleTurula, Holly, and Christiane E. Wobus. 2018. "The Role of the Polymeric Immunoglobulin Receptor and Secretory Immunoglobulins during Mucosal Infection and Immunity" Viruses 10, no. 5: 237. https://doi.org/10.3390/v10050237
APA StyleTurula, H., & Wobus, C. E. (2018). The Role of the Polymeric Immunoglobulin Receptor and Secretory Immunoglobulins during Mucosal Infection and Immunity. Viruses, 10(5), 237. https://doi.org/10.3390/v10050237