Identification of Felis catus Gammaherpesvirus 1 in Tsushima Leopard Cats (Prionailurus bengalensis euptilurus) on Tsushima Island, Japan


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
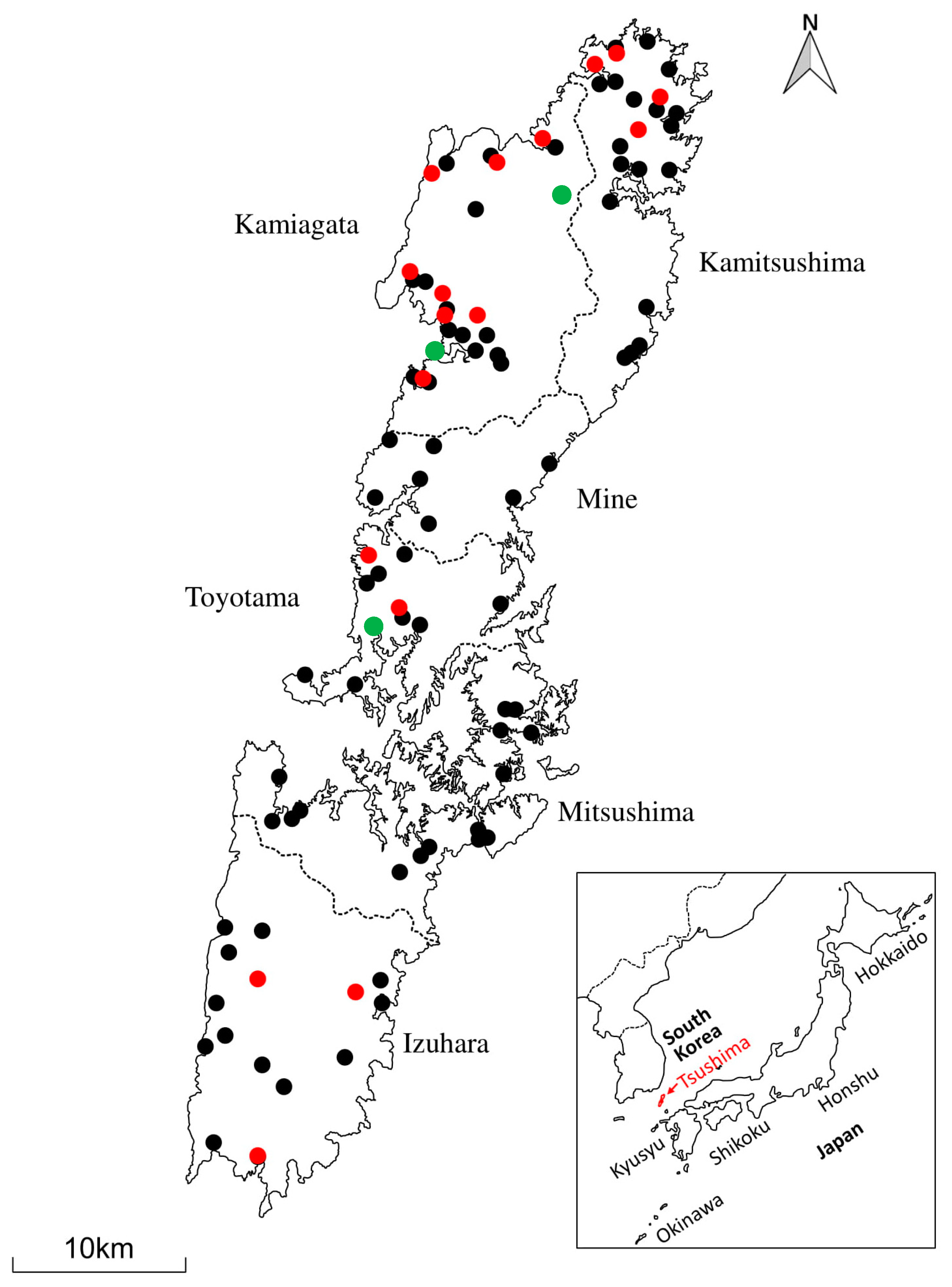
2.2. Study Area
2.3. Sample Collection and DNA Preparation
2.4. GAPDH PCR
2.5. PCR Amplification and Sequencing
2.6. Determination of PCR Sensitivity
2.7. Phylogenetic Analyses
2.8. Statistical Analyses
3. Results
3.1. Frequency and Distribution of FcaGHV1 on Tsushima Island
3.2. Variations of the FcaGHV1 Sequence
3.3. Sex and FIV/FeLV Status as Risk Factors for FcaGHV1 Infection in Domestic Cats
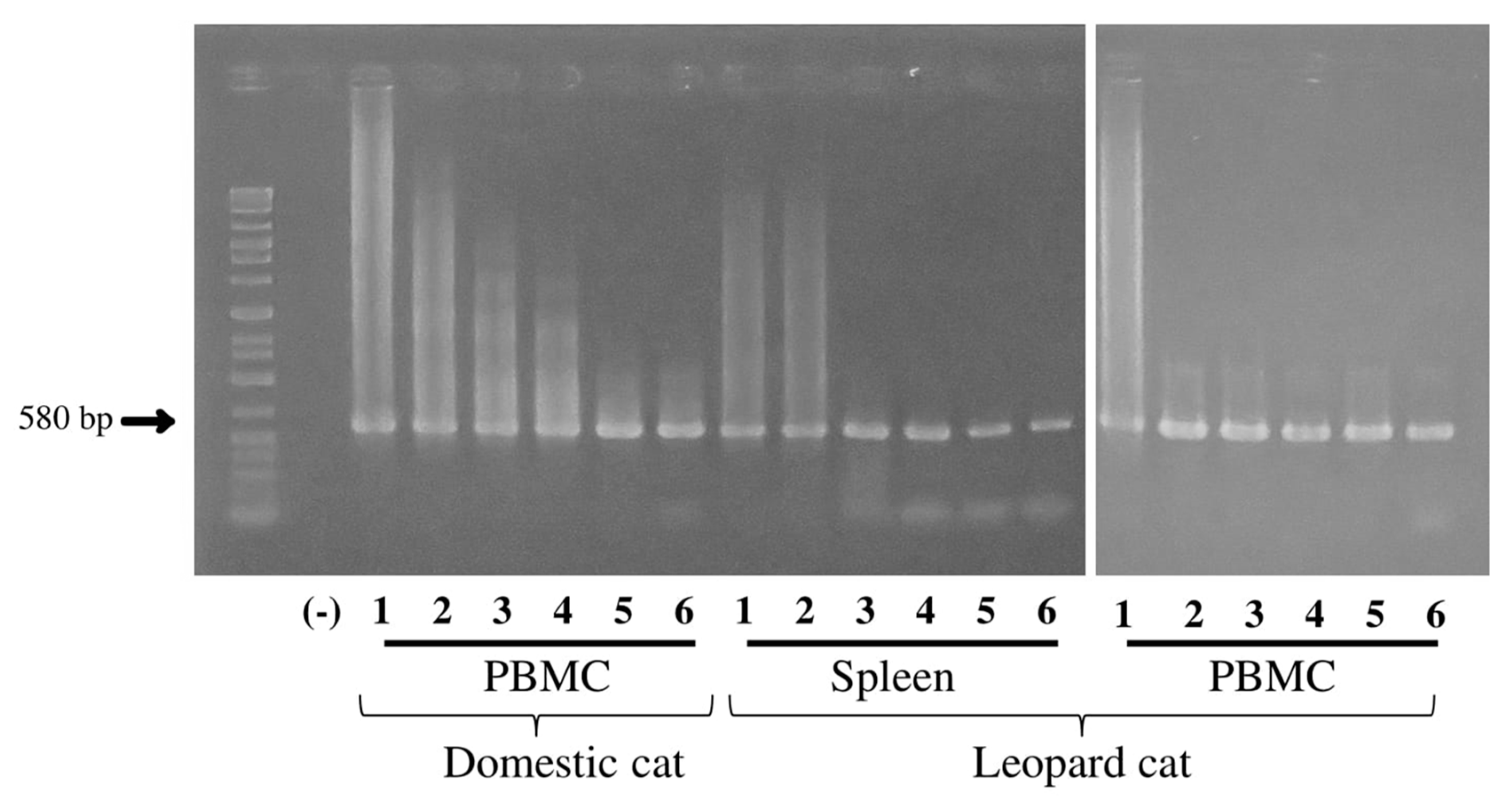
3.4. Sensitivity of FcaGHV1 gB PCR
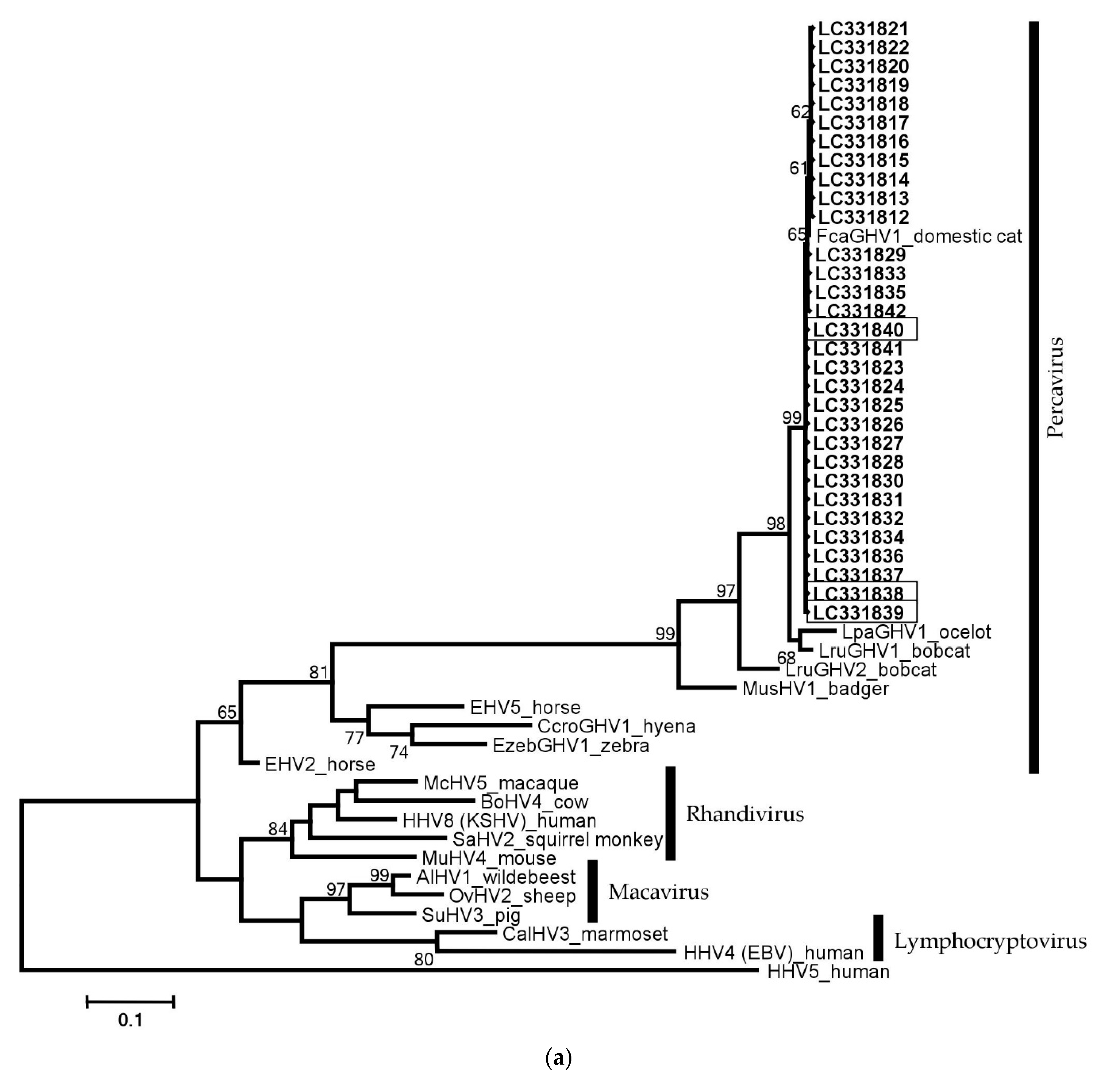
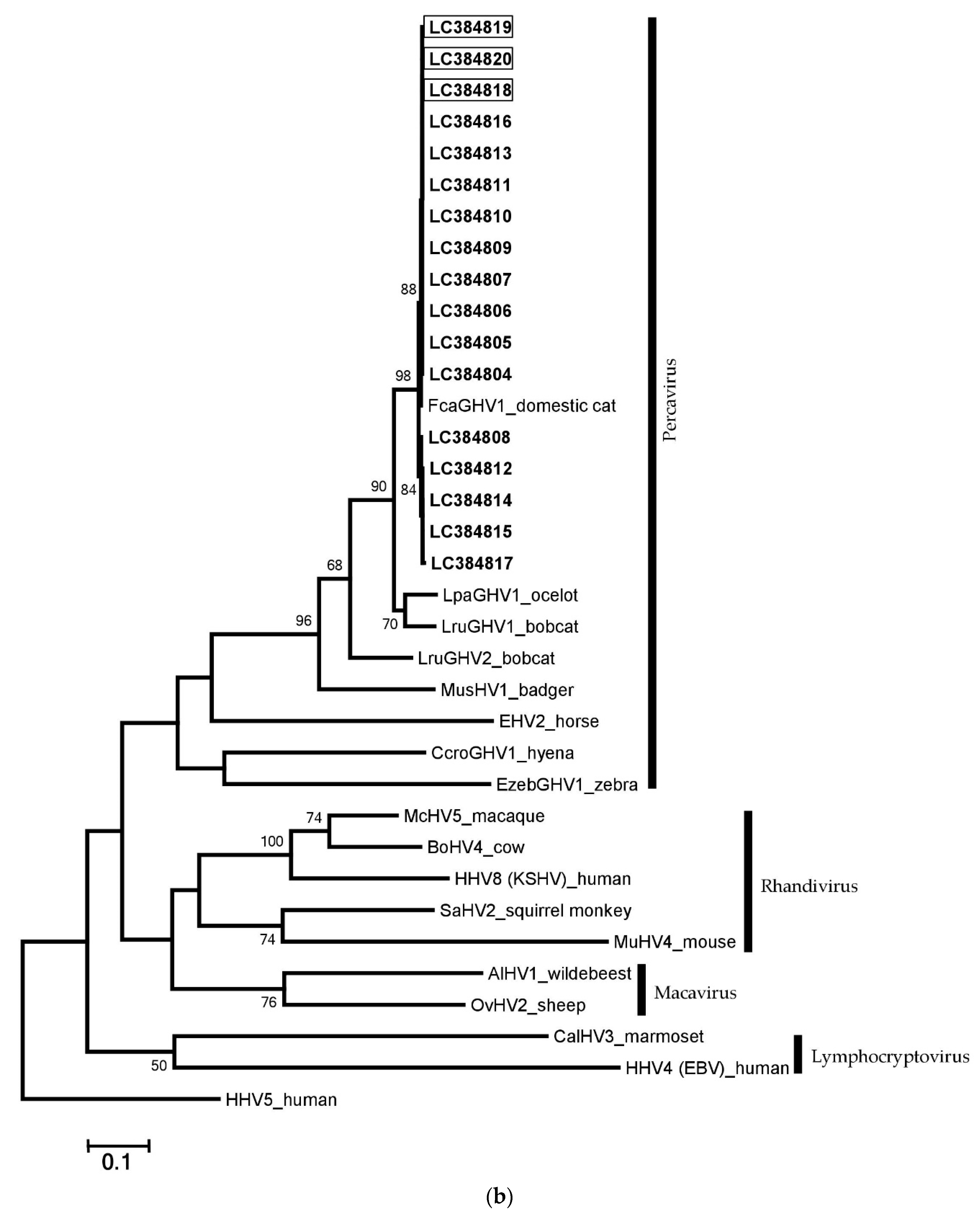
3.5. Phylogenetic Analyses and Comparison with Other GHVs
3.6. Nucleotide Sequence Accession Numbers
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tateno, M.; Nishio, T.; Matsuo, T.; Sakuma, M.; Nakanishi, N.; Izawa, M.; Asari, Y.; Okamura, M.; Shimokawa-Miyama, T.; Setoguchi, A.; et al. Epidemiological survey of tick-borne protozoal infection in iriomote cats and Tsushima leopard cats in Japan. J. Vet. Med. Sci. 2013, 75, 985–989. [Google Scholar] [CrossRef] [PubMed]
- Mitani, N.; Ishii, S.; Koike, H. Clues to the cause of the Tsushima leopard cat (Prionailurus bengalensis euptilura) decline from isotopic measurements in three species of Carnivora. Ecol. Res. 2009, 24, 897–908. [Google Scholar] [CrossRef]
- Tateno, M.; Nishio, T.; Sakuma, M.; Nakanishi, N.; Izawa, M.; Asari, Y.; Okamura, M.; Maruyama, S.; Miyama, T.S.; Setoguchi, A.; et al. Molecular epidemiologic survey of Bartonella, Ehrlichia, and Anaplasma infections in Japanese Iriomote and Tsushima leopard cats. J. Wildl. Dis. 2013, 49, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Hayama, Y.; Yamamoto, H.; Nakanishi, S.; Hiyama, T.; Murayama, A.; Mori, H.; Sugitani, A.; Fujiwara, S. Risk analysis of feline immunodeficiency virus infection in Tsushima leopard Cats (Prionailurus bengalensis euptilurus) and domestic cats using a Geographic Information System. J. Vet. Med. Sci. 2010, 9, 1113–1118. [Google Scholar] [CrossRef]
- Nishimura, Y.; Goto, Y.; Yoneda, K.; Endo, Y.; Mizuno, T.; Hamachi, M.; Maruyama, H.; Kinoshita, H.; Koga, S.; Watari, T.; et al. Interspecies transmission of feline immunodeficiency virus from the domestic cat to the Tsushima cat (Felis bengalensis euptilura) in the wild. J. Virol. 1999, 73, 7916–7921. [Google Scholar] [PubMed]
- Makundi, I.; Koshida, Y.; Kuse, K.; Hiratsuka, T.; Ito, J.; Baba, T.; Watanabe, S.; Kawamura, M.; Odahara, Y.; Miyake, A.; et al. Epidemiologic survey of feline leukemia virus in domestic cats on Tsushima Island, Japan: Management strategy for Tsushima leopard cats. J. Vet. Diagn. Investig. 2017, 29, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Barton, E.; Mandal, P.; Speck, S.H. Pathogenesis and host control of gammaherpesviruses: Lessons from the mouse. Annu. Rev. Immunol. 2011, 29, 351–397. [Google Scholar] [CrossRef] [PubMed]
- Speck, S.H.; Ganem, D. Viral latency and its regulation: Lessons from the gamma-herpesviruses. Cell Host Microbe 2010, 8, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, B.; Dural, G.; Yasmum, N.; Lembo, T.; De Thoisy, B.; Ryser-Degiorgis, M.P.; Ulrich, R.G.; Mcgeoch, D.J. Novel mammalian herpesviruses and lineages within the Gammaherpesvirinae: Cospeciation and interspecies transfer. J. Virol. 2008, 82, 3509–3516. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.M.; Venta, P.J.; Swenson, C.L.; Syring, R.; Gibbons-Burgener, S.N.; Richter, M.; Maes, R.K. Prevalence of bovine herpesvirus-4 infection in cats in Central Michigan. J. Vet. Intern. Med. 2000, 14, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Troyer, R.M.; Beatty, J.A.; Stutzman-Rodriguez, K.R.; Carver, S.; Lozano, C.C.; Lee, J.S.; Lappin, M.R.; Riley, S.P.D.; Serieys, L.E.K.; Logan, K.A.; et al. Novel gammaherpesviruses in north American domestic cats, bobcats, and pumas: Identification, prevalence, and risk factors. J. Virol. 2014, 88, 3914–3924. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.C.; Sweanor, L.L.; Wilson-Henjum, G.; Kays, R.W.; Moreno, R.; VandeWoude, S.; Troyer, R.M. Identification of novel gammaherpesviruses in Ocelots (Leopardus pardalis) and Bobcats (Lynx rufus) in Panama and Colorado, USA. J. Wildl. Dis. 2015, 51, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M. Pathogenesis of gammaherpesvirus infections. Vet. Microbiol. 2006, 113, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J.A.; Troyer, R.M.; Carver, S.; Barrs, V.R.; Espinasse, F.; Conradi, O.; Stutzman-Rodriguez, K.; Chan, C.C.; Tasker, S.; Lappin, M.R.; et al. Felis catus gammaherpesvirus 1; a widely endemic potential pathogen of domestic cats. Virology 2014, 460–461, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Ertl, R.; Korb, M.; Langbein-Detsch, I.; Klein, D. Prevalence and risk factors of gammaherpesvirus infection in domestic cats in Central Europe. Virol. J. 2015, 12, 146. [Google Scholar] [CrossRef] [PubMed]
- Tateno, M.; Takahashi, M.; Miyake, E.; Nishigaki, K.; Tsujimoto, H.; Endo, Y. Molecular epidemiological study of gammaherpesvirus in domestic cats in Japan. J. Vet. Med. Sci. 2017, 79, 1735–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurissio, J.K.; Rodrigues, M.V.; Taniwaki, S.A.; Zanutto, M.S.; Filoni, C.; Galdino, M.V.; Junior, J.P.A. Felis catus gammaherpesvirus 1 (FcaGHV1) and coinfections with feline viral pathogens in domestic cats in Brazil. Ciênc. Rural 2018, 48. [Google Scholar] [CrossRef]
- McLuckie, A.J.; Barrs, V.R.; Lindsay, S.; Aghazadeh, M.; Sangster, C.; Beatty, J.A. Molecular diagnosis of Felis catus gammaherpesvirus 1 (FcaGHV1) infection in cats of known retrovirus status with and without Lymphoma. Viruses 2018, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- McLuckie, A.J.; Barrs, V.R.; Wilson, B.; Westman, M.E.; Beatty, J.A. Felis catus gammaherpesvirus 1 DNAemia in whole blood from therapeutically immunosuppressed or retrovirus-infected cats. Vet. Sci. 2017, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Stutzman-Rodriguez, K.; Rovnak, J.; VandeWoude, S.; Troyer, R.M. Domestic cats seropositive for Felis catus gammaherpesvirus 1 are often qPCR negative. Virology 2016, 498, 23–30. [Google Scholar] [CrossRef] [PubMed]
- McLuckie, A.; Tasker, S.; Dhand, N.K.; Spencer, S.; Beatty, J.A. High prevalence of Felis catus gammaherpesvirus 1 infection in haemoplasma-infected cats supports co-transmission. Vet. J. 2016, 214, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acid Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Sleeman, J.M.; Keane, J.M.; Johnson, J.S.; Brown, R.J.; VandeWoude, S. Feline Leukemia Virus in a Captive Bobcat. J. Wildl. Dis. 2001, 37, 194–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Carver, S.; Beatty, J.A.; Troyer, R.M.; Harris, R.L.; Stutzman-Rodriguez, K.; Barrs, V.R.; Chan, C.C.; Tasker, S.; Lappin, M.R.; VandeWoude, S. Closing the gap on causal processes of infection risk from cross-sectional data: Structural equation models to understand infection and co-infection. Parasites Vectors 2015, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Primer a | Sequence (5′ to 3′) | Target Gene | Product Size (bp) | Reference |
|---|---|---|---|---|
| FcaGHV1gB–1s | GACCTGCACCAGAGCATGAG | |||
| FcaGHV1gB–1as | AGGATCCCTGGCAGATTGGT | 715 | ||
| FcaGHV1gB–2s | TGCACCAGAGCATGAGAGTT | |||
| FcaGHV1gB–2as | TCCCCCGAGAGGGTTTTTGA | gB | 580 | This study |
| FcaGHV1pol–1s | GGTGTTAATGGAAGCCCTGTG | |||
| FcaGHV1pol–1as | TTAGTCAGCCTTGGCATTGC | 818 | ||
| FcaGHV1pol–2s | ATGGAAGCCCTGTGAAGTTT | |||
| FcaGHV1pol–2as | CAGTGTCTCATTGCTTGCTGT | DNApol | 568 | This study |
| Host Species | No. of Samples | No. of FcaGHV1 Positive | %Positive (95% CI) |
|---|---|---|---|
| Leopard cat | 89 | 3 | 3.37 (0.70–9.54) |
| Domestic cat | 215 | 28 | 13.02 (8.83–18.27) |
| Variables | Categories | FcaGHV1 Status | Total | % Positive | |
|---|---|---|---|---|---|
| Positive | Negative | ||||
| Male | 22 | 72 | 94 | 23 | |
| Sex | Female | 6 | 115 | 121 | 5 |
| Kamijima | 22 | 145 | 167 | 13 | |
| Region | Shimojima | 6 | 42 | 48 | 12 |
| Negative | 11 | 107 | 118 | 9 | |
| FIV infection | Positive | 17 | 80 | 97 | 17 |
| Negative | 22 | 165 | 187 | 12 | |
| FeLV infection 1 | Positive | 6 | 22 | 28 | 21 |
| Variables | Categories | b 1 | SE 2 | Odds-Ratio | 95% CI | p-Value |
|---|---|---|---|---|---|---|
| Intercept | Male vs. female | −0.58 | 0.64 | |||
| Sex | 1.77 | 0.48 | 5.86 | 2.27–15.14 | <0.0001 | |
| Intercept | Kamijima vs. Shimojima | 2.13 | 1.87 | |||
| Region | −0.06 | 0.49 | 0.94 | 0.36–2.47 | 0.903 | |
| Intercept | Positive vs. negative | −2.08 | 2.25 | |||
| FIV infection | 0.73 | 0.41 | 2.07 | 0.92–4.66 | 0.080 | |
| Intercept | Positive vs. negative | −3.71 | 4.01 | |||
| FeLV infection | 0.72 | 0.51 | 2.05 | 0.75–5.60 | 0.163 |
| Variables | Categories | b 1 | SE 2 | Odds-Ratio | 95% CI | p-Value |
|---|---|---|---|---|---|---|
| Intercept | −11.02 | 5.87 | ||||
| Sex | Male vs. female | 1.64 | 0.49 | 5.17 | 1.95–13.70 | 0.001 |
| FIV infection | Positive vs. negative | 0.57 | 0.47 | 1.77 | 0.71–4.44 | 0.223 |
| FeLV infection | Positive vs. negative | 0.96 | 0.58 | 2.60 | 0.84–8.09 | 0.099 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Makundi, I.; Koshida, Y.; Endo, Y.; Nishigaki, K. Identification of Felis catus Gammaherpesvirus 1 in Tsushima Leopard Cats (Prionailurus bengalensis euptilurus) on Tsushima Island, Japan. Viruses 2018, 10, 378. https://doi.org/10.3390/v10070378
Makundi I, Koshida Y, Endo Y, Nishigaki K. Identification of Felis catus Gammaherpesvirus 1 in Tsushima Leopard Cats (Prionailurus bengalensis euptilurus) on Tsushima Island, Japan. Viruses. 2018; 10(7):378. https://doi.org/10.3390/v10070378
Chicago/Turabian StyleMakundi, Isaac, Yushi Koshida, Yasuyuki Endo, and Kazuo Nishigaki. 2018. "Identification of Felis catus Gammaherpesvirus 1 in Tsushima Leopard Cats (Prionailurus bengalensis euptilurus) on Tsushima Island, Japan" Viruses 10, no. 7: 378. https://doi.org/10.3390/v10070378