Critical Role of an MHC Class I-Like/Innate-Like T Cell Immune Surveillance System in Host Defense against Ranavirus (Frog Virus 3) Infection



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and methods:
2.1. Animals
2.2. Frog Virus 3 Stocks and Infection
2.3. Quantitative Gene Expression Analyses
2.4. Viral Load Quantification by qPCR and Plaque Assay
2.5. XNC10 Tetramer Production
2.6. Statistical Analysis
3. Results
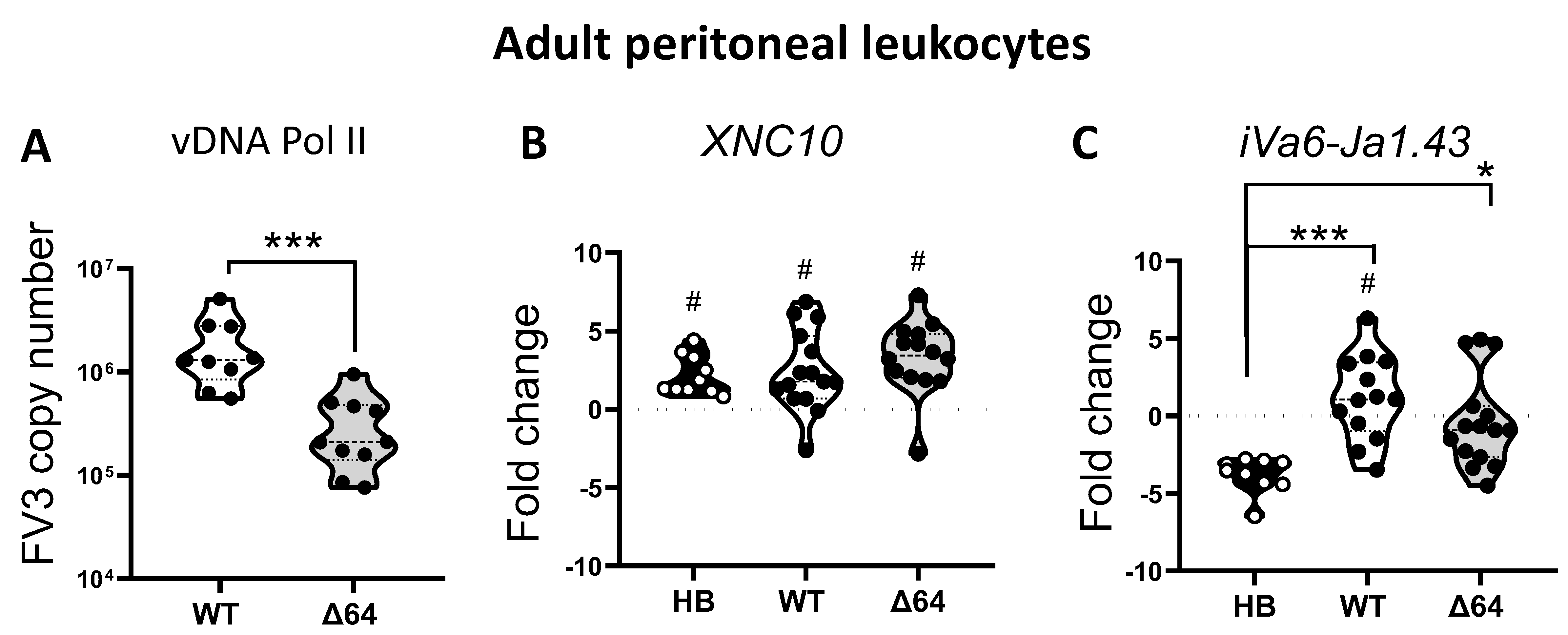
3.1. Relationships Between FV3 Infection Magnitude and iVα6 T cell Response in Adult X. Laevis
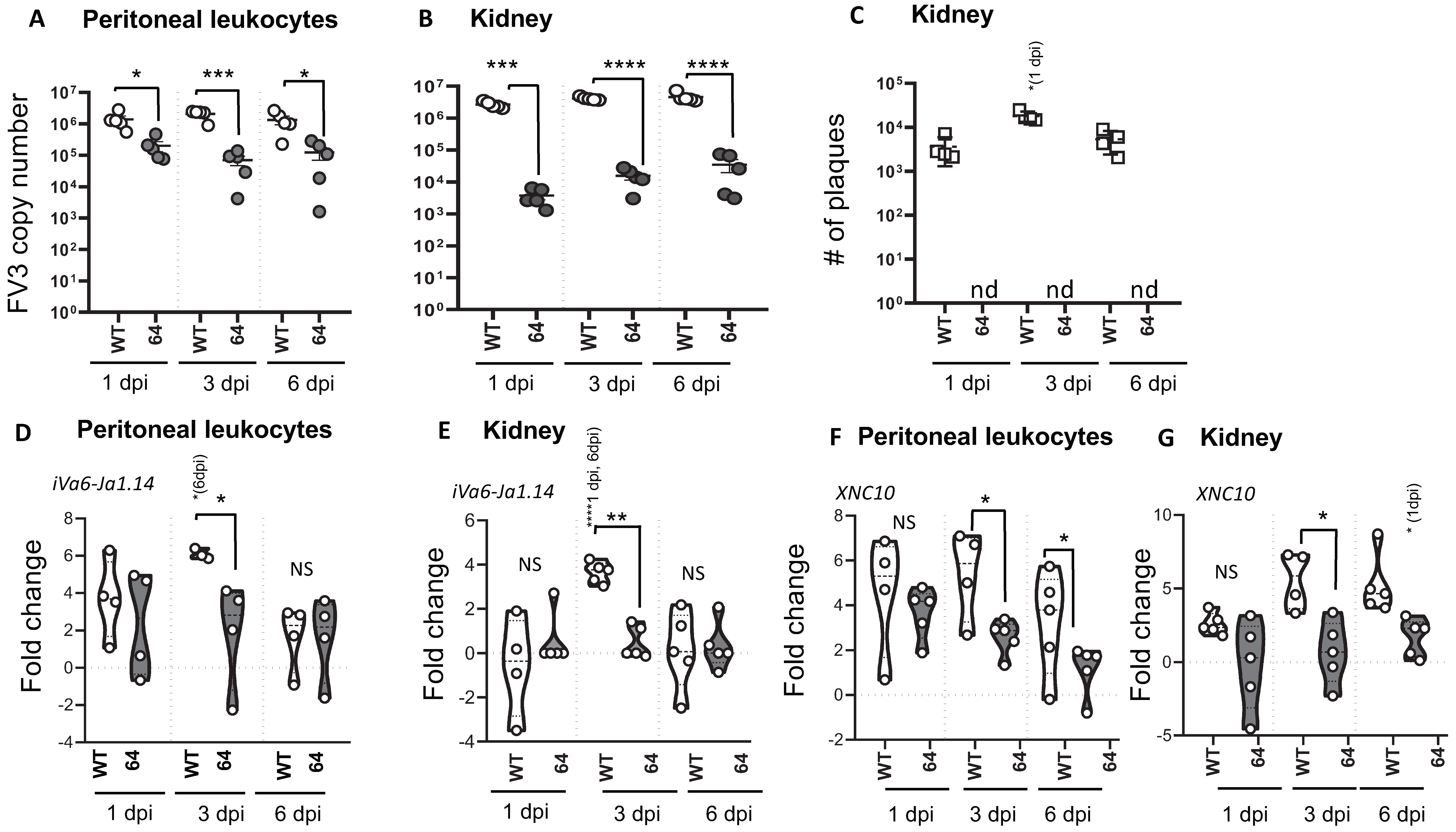
3.2. Relationships Between FV3 Infection Magnitude and iVα6 T Cell Response in Tadpoles
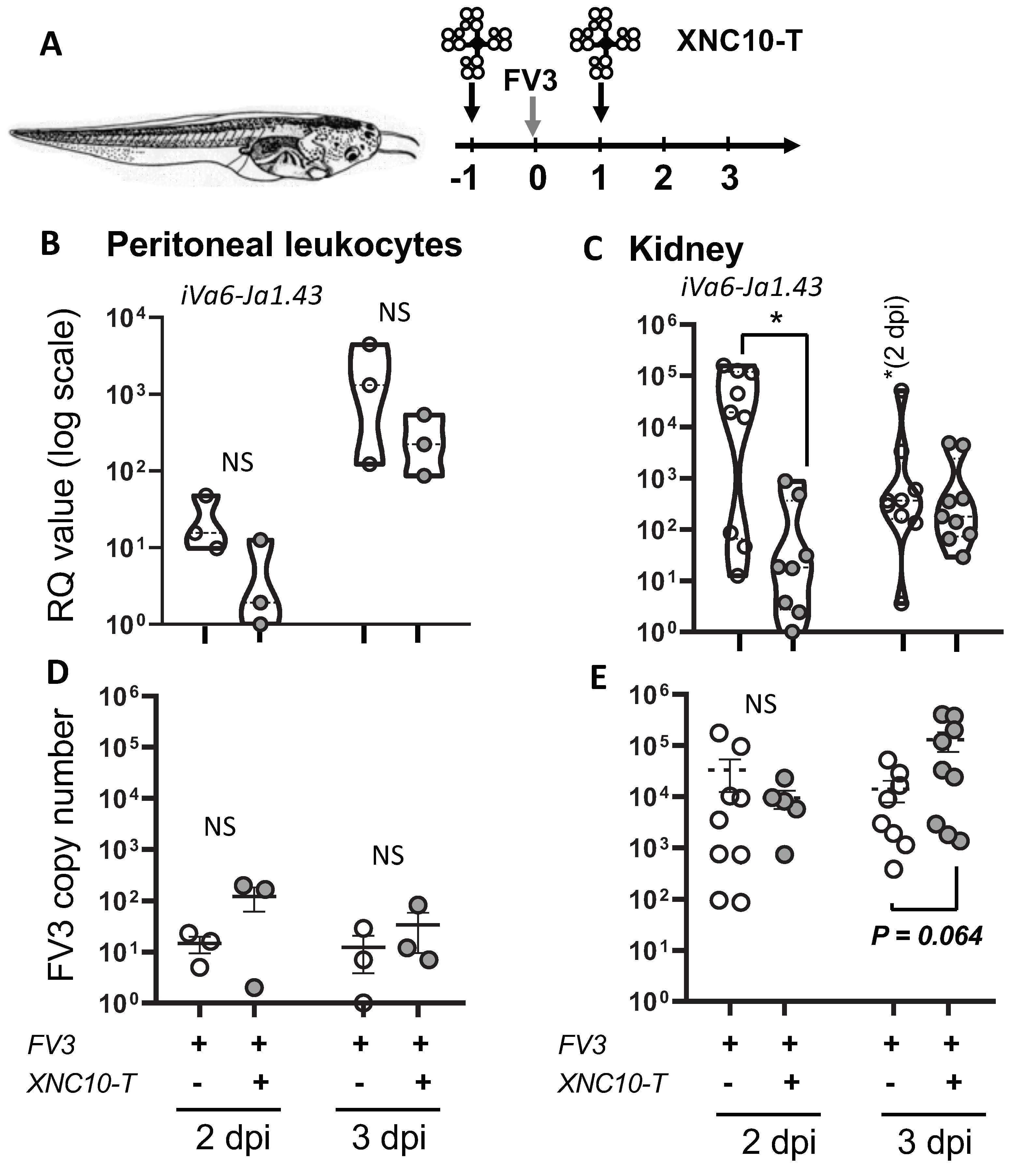
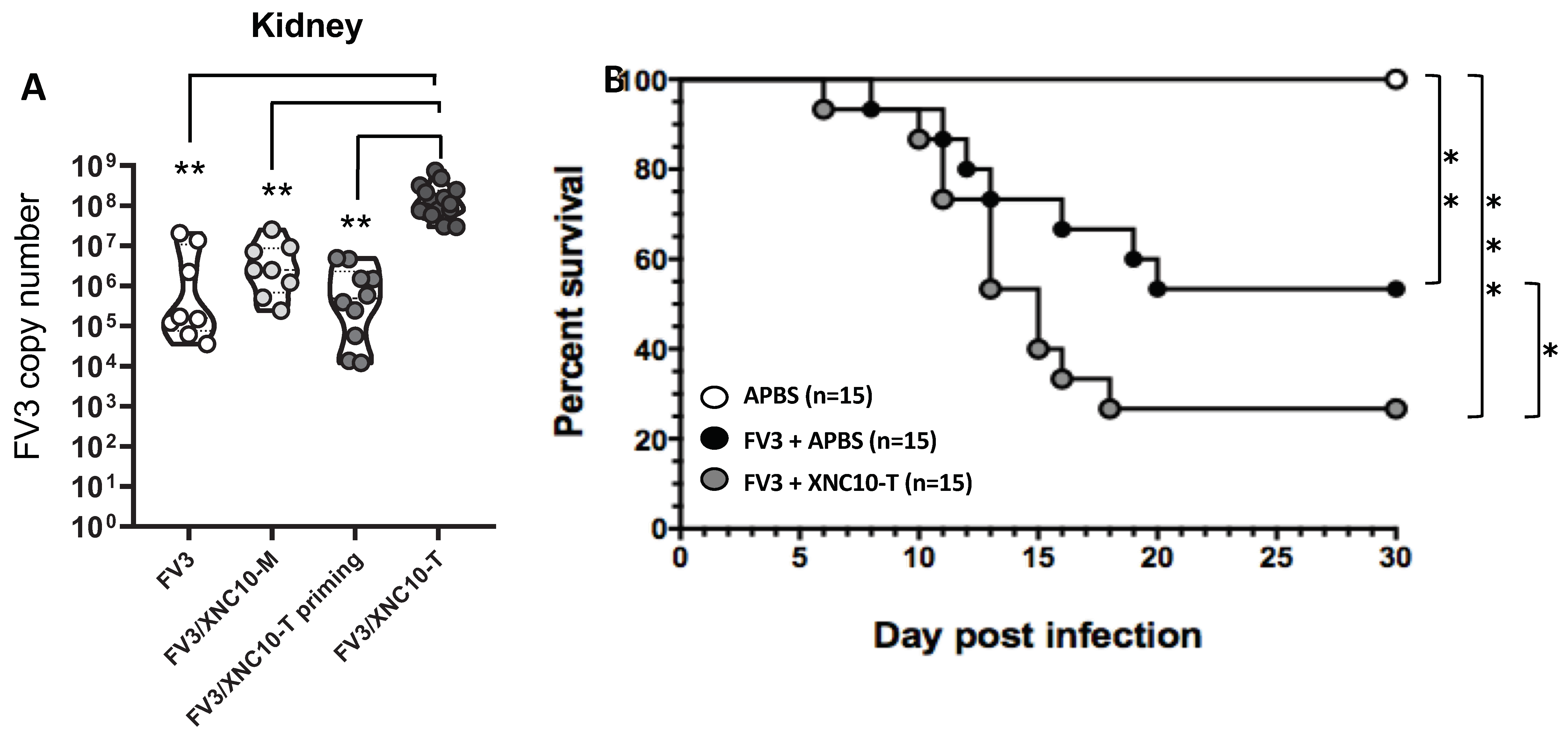
3.3. Targeted iVα6T cell depletion in vivo
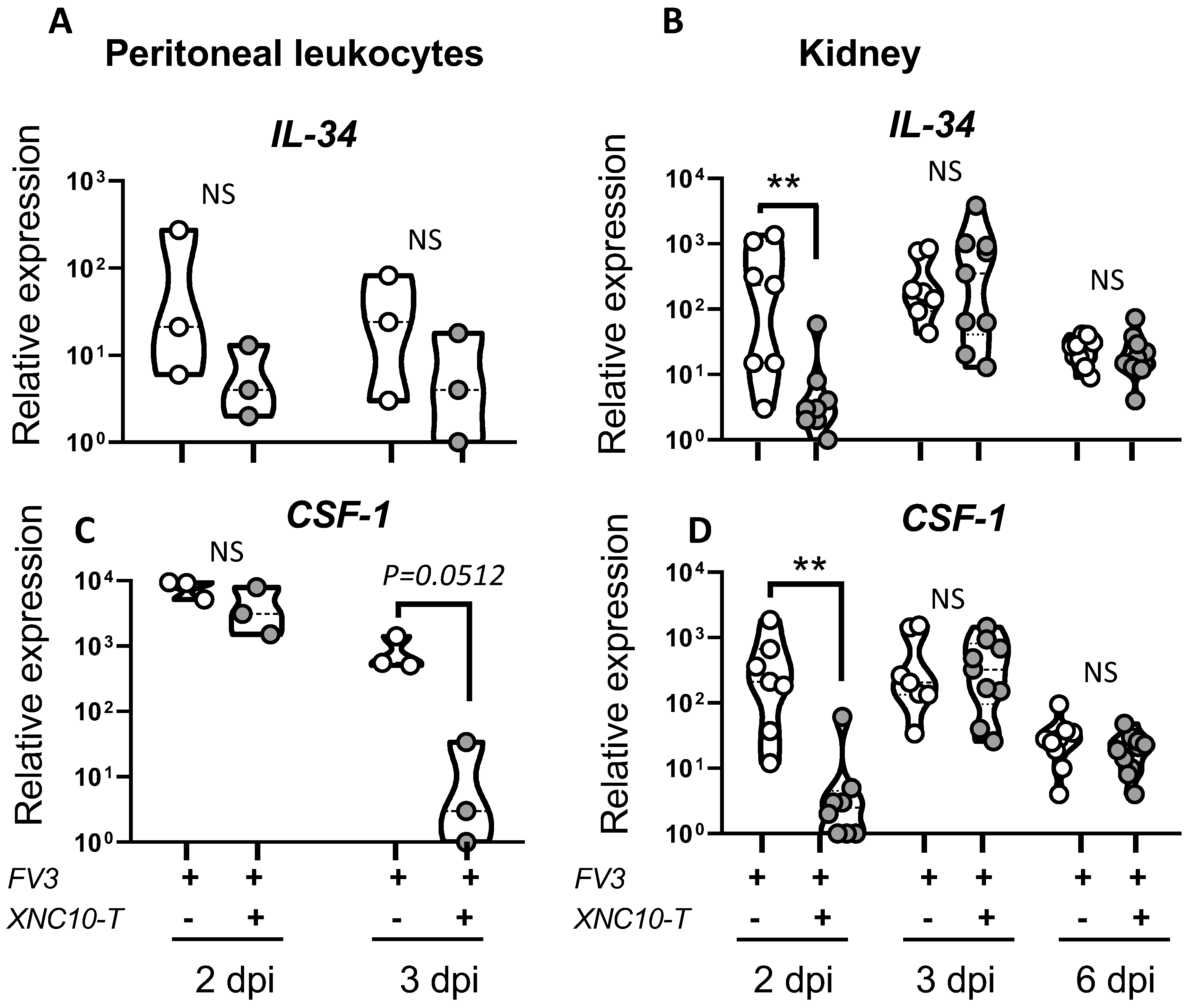
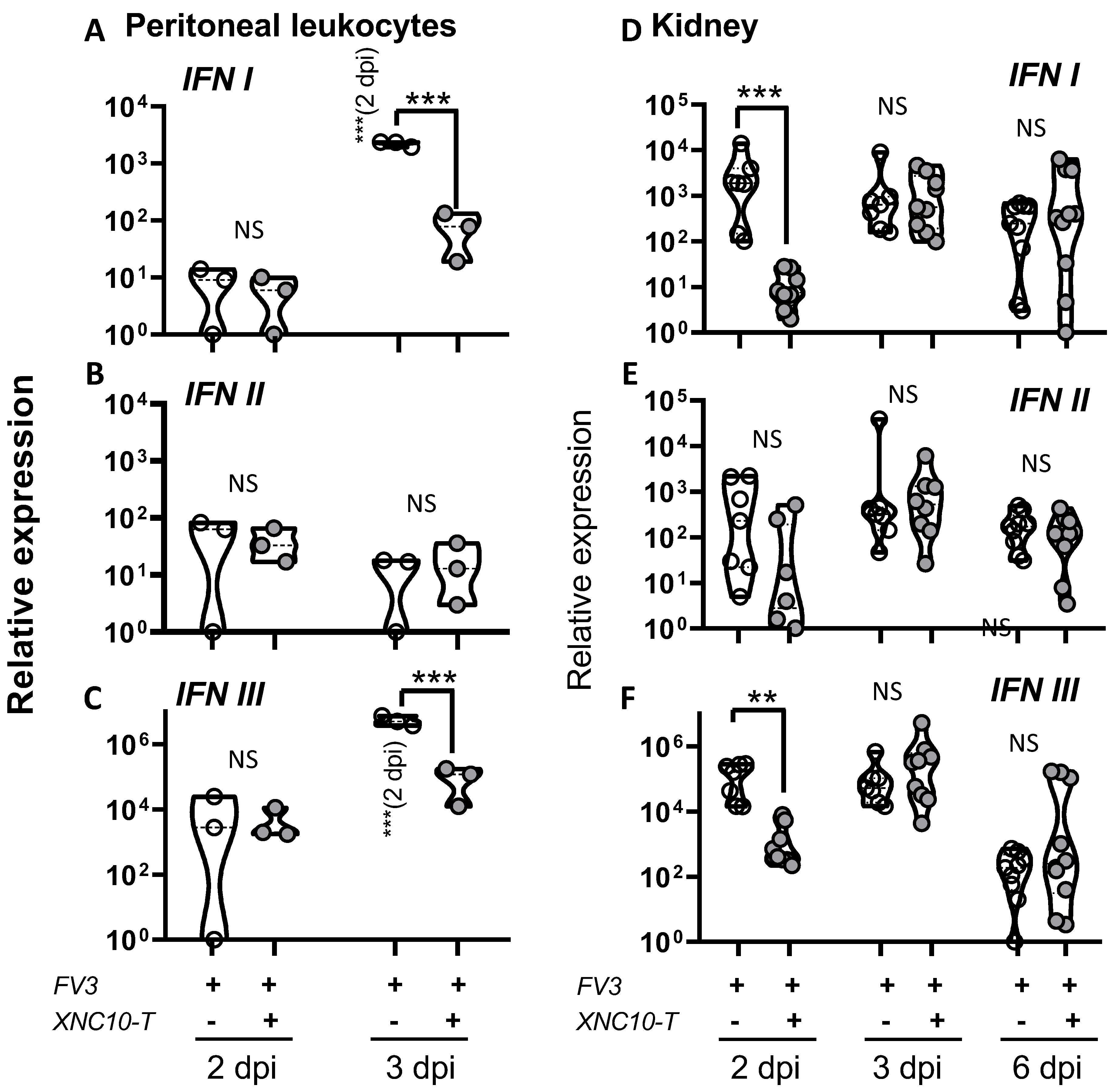
3.4. Effects of iVα6T cell depletion on PLs and kidney antiviral responses in tadpoles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, E.J.; Luoma, A.M. The adaptable major histocompatibility complex (MHC) fold: Structure and function of nonclassical and MHC class I-like molecules. Annu. Rev. Immunol. 2013, 31, 529–561. [Google Scholar] [CrossRef]
- Borg, N.A.; Wun, K.S.; Kjer-Nielsen, L.; Wilce, M.C.; Pellicci, D.G.; Koh, R.; Besra, G.S.; Bharadwaj, M.; Godfrey, D.I.; McCluskey, J.; et al. CD1d-lipid-antigen recognition by the semi-invariant NKT T-cell receptor. Nature 2007, 448, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Rossjohn, J.; Pellicci, D.G.; Patel, O.; Gapin, L.; Godfrey, D.I. Recognition of CD1d-restricted antigens by natural killer T cells. Nat. Rev. Immunol. 2012, 12, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Wesley, J.D.; Tessmer, M.S.; Chaukos, D.; Brossay, L. NK Cell–Like Behavior of Vα14i NK T Cells during MCMV Infection. PLoS Pathog. 2008, 4, e1000106. [Google Scholar] [CrossRef] [PubMed]
- Diana, J.; Griseri, T.; Lagaye, S.; Beaudoin, L.; Autrusseau, E.; Gautron, A.-S.; Tomkiewicz, C.; Herbelin, A.; Barouki, R.; von Herrath, M.; et al. NKT Cell-Plasmacytoid Dendritic Cell Cooperation via OX40 Controls Viral Infection in a Tissue-Specific Manner. Immunity 2009, 30, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Raftery, M.J.; Winau, F.; Giese, T.; Kaufmann, S.H.; Schaible, U.E.; Schonrich, G. Viral danger signals control CD1d de novo synthesis and NKT cell activation. Eur. J. Immunol. 2008, 38, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Juno, J.A.; Keynan, Y.; Fowke, K.R. Invariant NKT cells: Regulation and function during viral infection. PLoS Pathog. 2012, 8, e1002838. [Google Scholar] [CrossRef]
- Abboud, G.; Tahiliani, V.; Desai, P.; Varkoly, K.; Driver, J.; Hutchinson, T.E.; Salek-Ardakani, S. Natural Killer Cells and Innate Interferon Gamma Participate in the Host Defense against Respiratory Vaccinia Virus Infection. J. Virol. 2016, 90, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Edholm, E.S.; Banach, M.; Robert, J. Evolution of innate-like T cells and their selection by MHC class I-like molecules. Immunogenetics 2016, 68, 525–536. [Google Scholar] [CrossRef]
- Edholm, E.S.; Albertorio Saez, L.M.; Gill, A.L.; Gill, S.R.; Grayfer, L.; Haynes, N.; Myers, J.R.; Robert, J. Nonclassical MHC class I-dependent invariant T cells are evolutionarily conserved and prominent from early development in amphibians. Proc. Natl. Acad. Sci. USA 2013, 110, 14342–14347. [Google Scholar] [CrossRef]
- Price, S.J.; Garner, T.W.; Nichols, R.A.; Balloux, F.; Ayres, C.; Mora-Cabello de Alba, A.; Bosch, J. Collapse of amphibian communities due to an introduced Ranavirus. Curr. Biol. CB 2014, 24, 2586–2591. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J.; Chinchar, V.G. Ranaviruses; Springer Open: Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherland; London, UK, 2015. [Google Scholar]
- Edholm, E.S.; Grayfer, L.; De Jesus Andino, F.; Robert, J. Nonclassical MHC-Restricted Invariant Valpha6 T Cells Are Critical for Efficient Early Innate Antiviral Immunity in the Amphibian Xenopus laevis. J. Immunol. 2015, 195, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Nieuwkoop, P.; Faber, J. Normal Table of Xenopus Laevis (Daudin): A Systematical and Chronological Survey of the Development from the Fertilized Egg Till the End of Metamorphosis, 2nd ed.; North Holland: Amsterdam, The Netherland, 1967. [Google Scholar]
- Andino Fde, J.; Grayfer, L.; Chen, G.; Gregory Chinchar, V.; Edholm, E.S.; Robert, J. Characterization of Frog Virus 3 knockout mutants lacking putative virulence genes. Virology 2015, 485, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ward, B.M.; Yu, K.H.; Chinchar, V.G.; Robert, J. Improved knockout methodology reveals that frog virus 3 mutants lacking either the 18K immediate-early gene or the truncated vIF-2alpha gene are defective for replication and growth in vivo. J. Virol. 2011, 85, 11131–11138. [Google Scholar] [CrossRef]
- Jacques, R.; Edholm, E.S.; Jazz, S.; Odalys, T.L.; Francisco, J.A. Xenopus-FV3 host-pathogen interactions and immune evasion. Virology 2017, 511, 309–319. [Google Scholar] [CrossRef]
- Morales, H.D.; Abramowitz, L.; Gertz, J.; Sowa, J.; Vogel, A.; Robert, J. Innate immune responses and permissiveness to ranavirus infection of peritoneal leukocytes in the frog Xenopus laevis. J. Virol. 2010, 84, 4912–4922. [Google Scholar] [CrossRef]
- Grayfer, L.; Robert, J. Divergent antiviral roles of amphibian (Xenopus laevis) macrophages elicited by colony-stimulating factor-1 and interleukin-34. J. Leukoc. Biol. 2014, 96, 1143–1153. [Google Scholar] [CrossRef]
- Grayfer, L.; Robert, J. Colony-Stimulating Factor-1-Responsive Macrophage Precursors Reside in the Amphibian (Xenopus laevis) Bone Marrow Rather than the Hematopoietic Sub-Capsular Liver. J. Innate Immun. 2013, 5, 531–542. [Google Scholar] [CrossRef]
- Barbarin, A.; Cayssials, E.; Jacomet, F.; Nunez, N.G.; Basbous, S.; Lefevre, L.; Abdallah, M.; Piccirilli, N.; Morin, B.; Lavoue, V.; et al. Phenotype of NK-Like CD8(+) T Cells with Innate Features in Humans and Their Relevance in Cancer Diseases. Front. Immunol. 2017, 8, 316. [Google Scholar] [CrossRef]
- Huber, C.M.; Doisne, J.M.; Colucci, F. IL-12/15/18-preactivated NK cells suppress GvHD in a mouse model of mismatched hematopoietic cell transplantation. Eur. J. Immunol. 2015, 45, 1727–1735. [Google Scholar] [CrossRef]
- Zajonc, D.M.; Girardi, E. Recognition of Microbial Glycolipids by Natural Killer T Cells. Front. Immunol. 2015, 6, 400. [Google Scholar] [CrossRef]
- Kohlgruber, A.C.; Donado, C.A.; LaMarche, N.M.; Brenner, M.B.; Brennan, P.J. Activation strategies for invariant natural killer T cells. Immunogenetics 2016, 68, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Morales, H.D.; Robert, J. Characterization of primary and memory CD8 T-cell responses against ranavirus (FV3) in Xenopus laevis. J. Virol. 2007, 81, 2240–2248. [Google Scholar] [CrossRef]
- Edholm, E.S.; Goyos, A.; Taran, J.; De Jesus Andino, F.; Ohta, Y.; Robert, J. Unusual evolutionary conservation and further species-specific adaptations of a large family of nonclassical MHC class Ib genes across different degrees of genome ploidy in the amphibian subfamily Xenopodinae. Immunogenetics 2014, 66, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Goyos, A.; Sowa, J.; Ohta, Y.; Robert, J. Remarkable conservation of distinct nonclassical MHC class I lineages in divergent amphibian species. J. Immunol. 2011, 186, 372–381. [Google Scholar] [CrossRef]
- De Jesús Andino, F.; Chen, G.; Li, Z.; Grayfer, L.; Robert, J. Susceptibility of Xenopus laevis tadpoles to infection by the ranavirus Frog-Virus 3 correlates with a reduced and delayed innate immune response in comparison with adult frogs. Virology 2012, 432, 435–443. [Google Scholar]
- Grayfer, L.; De Jesus Andino, F.; Robert, J. Prominent amphibian (Xenopus laevis) tadpole type III interferon response to the frog virus 3 ranavirus. J. Virol. 2015, 89, 5072–5082. [Google Scholar] [CrossRef]
- Droin, N.; Solary, E. Editorial: CSF1R, CSF-1, and IL-34, a "menage a trois" conserved across vertebrates. J. Leukoc. Biol. 2010, 87, 745–747. [Google Scholar] [CrossRef]
- Grayfer, L.; Robert, J. Distinct functional roles of amphibian (Xenopus laevis) colony-stimulating factor-1- and interleukin-34-derived macrophages. J. Leukoc. Biol. 2015, 98, 641–649. [Google Scholar] [CrossRef]
- Yasuda, K.; Nakanishi, K.; Tsutsui, H. Interleukin-18 in Health and Disease. Int. J. Mol. Sci. 2019, 20, E649. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edholm, E.-S.I.; De Jesús Andino, F.; Yim, J.; Woo, K.; Robert, J. Critical Role of an MHC Class I-Like/Innate-Like T Cell Immune Surveillance System in Host Defense against Ranavirus (Frog Virus 3) Infection. Viruses 2019, 11, 330. https://doi.org/10.3390/v11040330
Edholm E-SI, De Jesús Andino F, Yim J, Woo K, Robert J. Critical Role of an MHC Class I-Like/Innate-Like T Cell Immune Surveillance System in Host Defense against Ranavirus (Frog Virus 3) Infection. Viruses. 2019; 11(4):330. https://doi.org/10.3390/v11040330
Chicago/Turabian StyleEdholm, Eva-Stina Isabella, Francisco De Jesús Andino, Jinyeong Yim, Katherine Woo, and Jacques Robert. 2019. "Critical Role of an MHC Class I-Like/Innate-Like T Cell Immune Surveillance System in Host Defense against Ranavirus (Frog Virus 3) Infection" Viruses 11, no. 4: 330. https://doi.org/10.3390/v11040330