Surveys in the Chrysanthemum Production Areas of Brazil and Colombia Reveal That Weeds Are Potential Reservoirs of Chrysanthemum Stunt Viroid

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
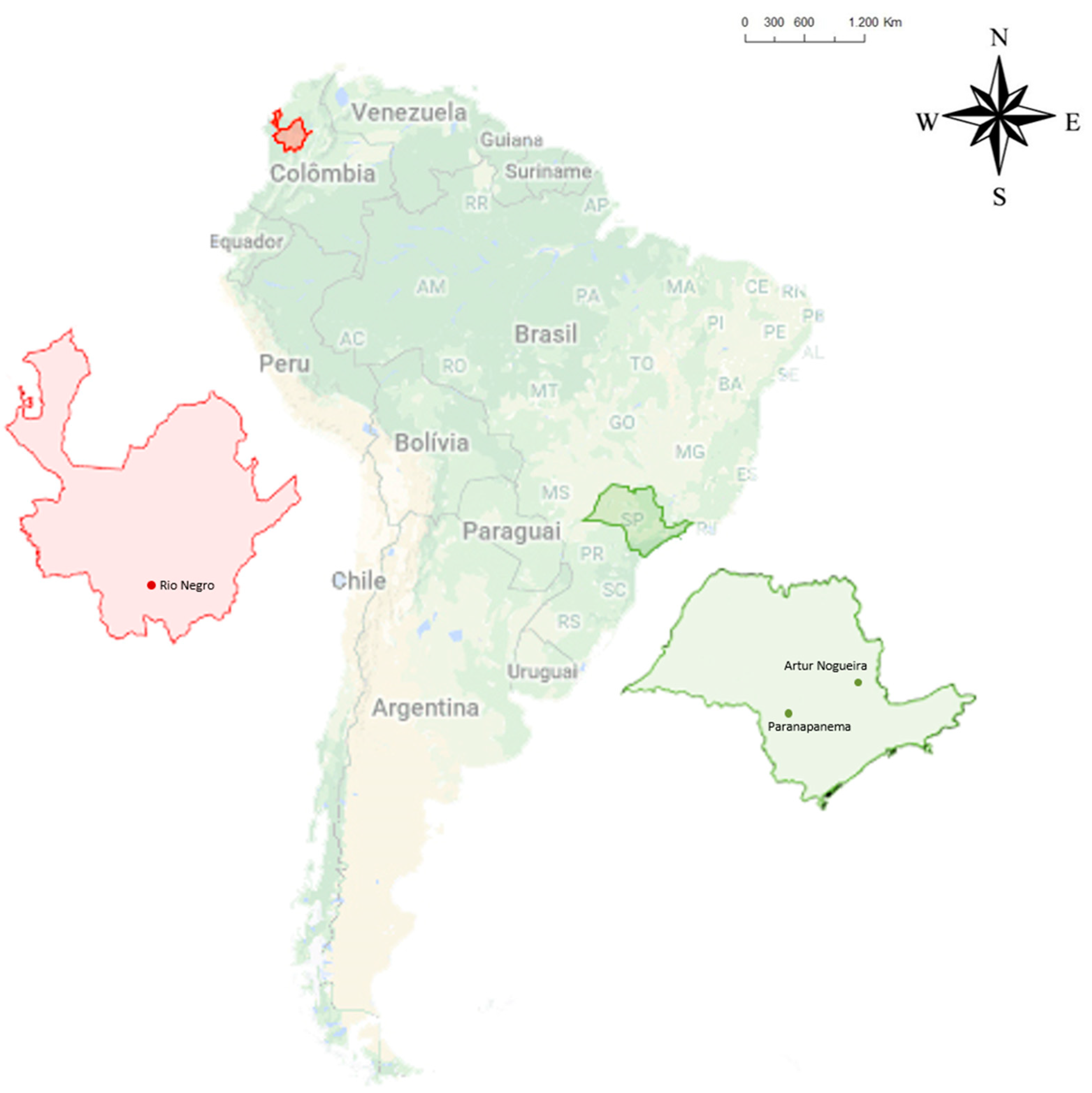
2.1. Phytosociological Survey in Fields of Vase and Cut Chrysanthemums
2.2. Mechanical Inoculation and Experimental Host Range
2.3. RNA Purification for the RT-PCR Analysis
2.4. RNA Purification for Sequential Polyacrylamide Gel Electrophoresis (sPAGE)
2.5. RT-PCR
2.6. Sequencing, Sequence Analyses and Phylogeny
3. Results and Discussion
3.1. Detection of Chrysanthemum Stunt Viroid (CSVd) in Broadleaf Woodsorrel and Chrysanthemum in Colombia
3.2. Experimental Host Range of the Brazilian CSVd Isolate
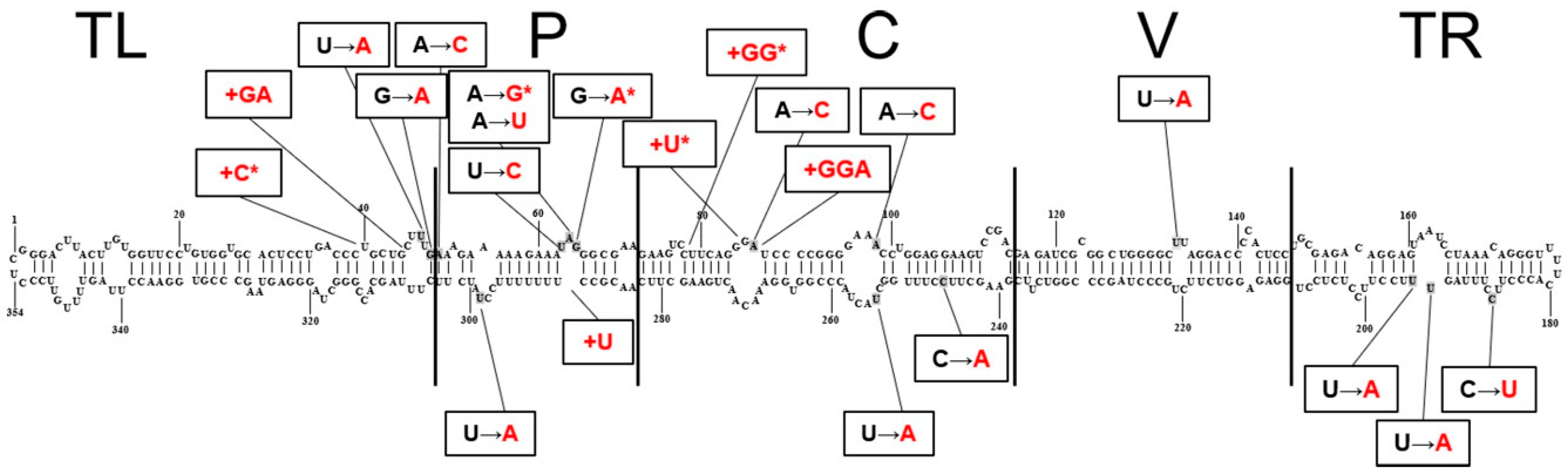
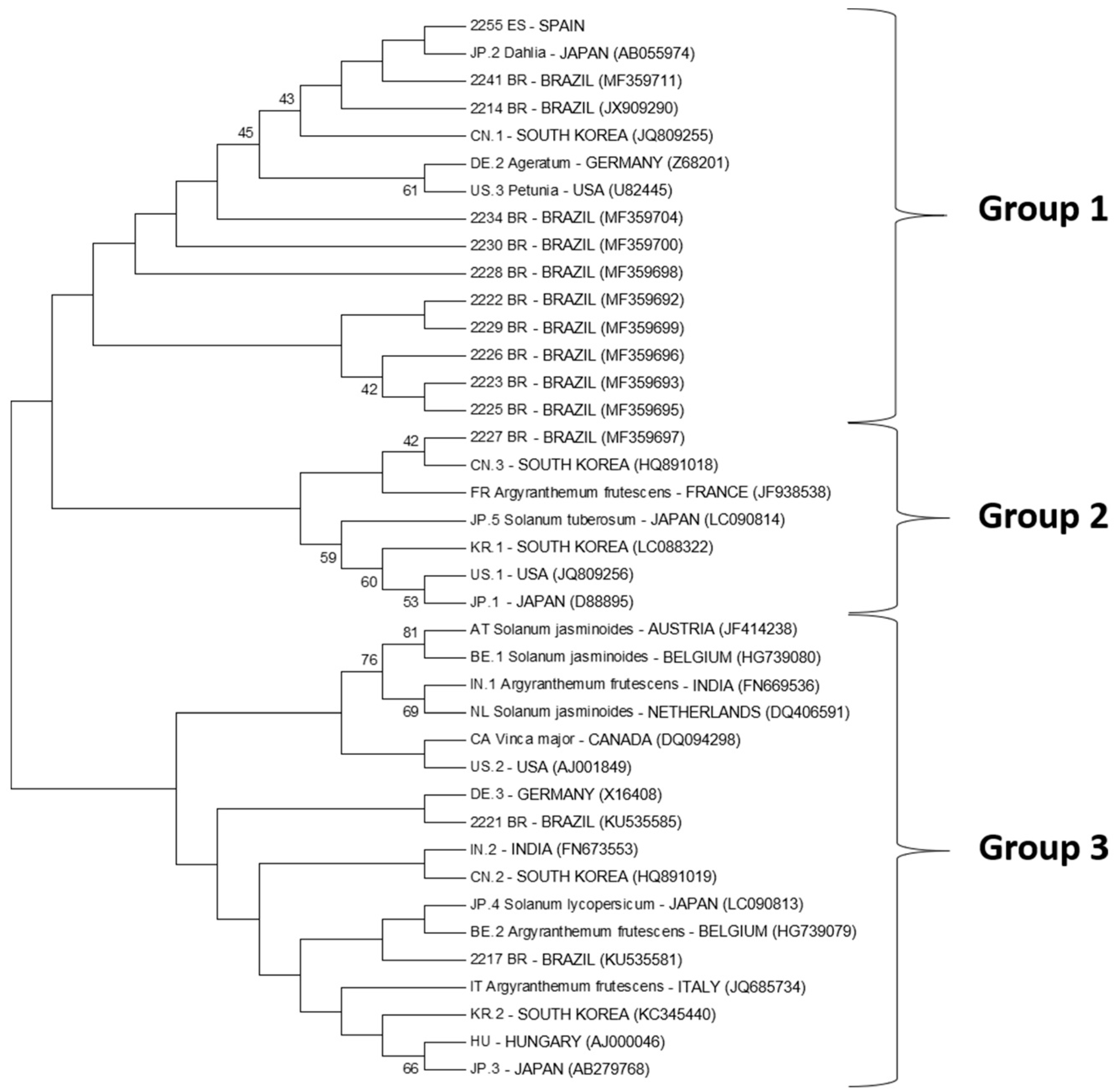
3.3. Diversity of CSVd Isolates: Sequence Analyses and Phylogeny
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ding, B. The biology of viroid-host interactions. Ann. Rev. Phytopath. 2009, 47, 105–131. [Google Scholar] [CrossRef]
- Flores, R.; Serra, P.; Minoia, S.; Di Serio, F.; Navarro, B. Viroids: from genotype to phenotype just relying on RNA sequence and structural motifs. Front. Microbiol. 2012, 3, 217. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Gago-Zachert, S.; Serra, P.; Sanjuan, R.; Elena, S.F. Viroids: Survivors from the RNA World? Annu. Rev. Microbiol. 2014, 68, 395–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, R.; Owens, R.A.; Taylor, J. Pathogenesis by subviral agents: Viroids and hepatits delta virus. Curr. Opin. Virol. 2016, 17, 87–94. [Google Scholar] [CrossRef]
- Steger, G.; Perreault, J.P. Structure and associated biological functions of viroids. Adv. Virus Res. 2016, 94, 141–172. [Google Scholar] [CrossRef]
- Steger, G.; Riesner, D. Viroid research and its significance for RNA technology and basic biochemistry. Nucleic Acids Res. 2018, 46, 10563–10576. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A.; Flores, R.; Randles, J.W.; Palukaitis, P. (Eds.) Viroids and Satellites; Elsevier: London, UK, 2017; ISBN 9780128014981. [Google Scholar]
- Gobatto, D.; Chaves, A.L.R.; Harakava, R.; Marque, J.M.; Daròs, J.A.; Eiras, M. Chrysanthemum stunt viroid in Brazil: Survey, identification, biological and molecular characterization and detection methods. J. Plant Pathol. 2014, 96, 111–119. [Google Scholar] [CrossRef]
- Hammond, R.W. Economic significance of viroids in vegetable and field crops. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Elsevier: London, UK, 2017; pp. 5–13. ISBN 9780128014981. [Google Scholar]
- Rodriguez, M.; Vadamalai, G.; Randles, J.W. Economic Significance of in Palm Tree Viroids. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Elsevier: London, UK, 2017; pp. 27–38. ISBN 9780128014981. [Google Scholar]
- Verhoeven, J.T.J.; Hammond, R.W.; Stancanelli, G. Economic significance of viroids in ornamental crops. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Elsevier: London, UK, 2017; pp. 27–38. ISBN 9780128014981. [Google Scholar]
- Siddell, S.G.; Walker, P.J.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; et al. Additional changes to taxonomy ratified in a special vote by the International Committee on Taxonomy of Viruses (October 2018). Arch. Virol. 2019, 164, 943. [Google Scholar] [CrossRef]
- Di Serio, F.; Flores, R.; Verhoeven, J.T.J.; Li, S.-F.; Pallas, V.; Randles, J.W.; Sano, T.; Vidalakis, G.; Owens, R.A. Current status of viroid taxonomy. Arch. Virol. 2014, 159, 3467–3478. [Google Scholar] [CrossRef] [Green Version]
- Diener, T.O.; Raymer, W.B. Potato spindle tuber virus: A plant virus with properties of a free nucleic acid. Science 1967, 158, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Diener, T.O. Potato spindle tuber ‘virus’ IV. A replicating, low molecular weight RNA. Virology 1971, 45, 411–428. [Google Scholar] [CrossRef]
- Diener, T.O. Potato spindle tuber virus: A plant virus with properties of a free nucleic acid. III. Subcellular location of PSTVd-RNA and the question of weather virions exit in extracts or in situ. Virology 1971, 43, 75–89. [Google Scholar] [CrossRef]
- Semancik, J.S.; Weathers, L.G. Exocortis disease: Evidence for a new species of “infectious” low molecular weight RNA in plants. Nat. New Biol. 1972, 237, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Semancik, J.S.; Weathers, L.G. Exocortis virus: An infectious free-nucleic acid plant virus with unusual properties. Virology 1972, 47, 456–466. [Google Scholar] [CrossRef]
- Hadidi, A.; Flores, R.; Randles, J.W.; Semancik, J.S. (Eds.) Viroids; CSIRO Publishing: Collingwood, Australia, 2003; ISBN 0643067892. [Google Scholar]
- Dimock, A.W. Chrysanthemum stunt. N. Y. State Flower Grow. Bull. 1947, 26, 2. [Google Scholar]
- Diener, T.O.; Lawson, R.H. Chrysanthemum stunt: A viroid disease. Virology 1973, 51, 94–101. [Google Scholar] [CrossRef]
- Hollings, M.; Stone, O.M. Some properties of chrysanthemum stunt, a virus with the characteristics of an uncoated ribonucleic acid. Ann. Appl. Biol. 1973, 74, 333–348. [Google Scholar] [CrossRef]
- Lawson, R.H. Chrysantemum Stunt. In The viroids; Diener, T.O., Ed.; Plenum Press: New York, NY, USA, 1987; pp. 247–259. ISBN 9781461318552. [Google Scholar]
- Palukaitis, P.; Symons, R.H. Purification and characterization of the circular and linear forms of chrysanthemum stunt viroid. J. Gen. Virol. 1980, 46, 477–489. [Google Scholar] [CrossRef]
- Haseloff, J.; Symons, R.H. Chrysantemum stunt viroid: Primary sequence and secondary structure. Nucleic Acids Res. 1981, 9, 2741–2752. [Google Scholar] [CrossRef]
- Gross, H.J.; Krupp, G.; Domdey, H.; Raba, M.; Jank, P.; Lossow, C.; Alberty, H.; Ramm, K.; Sanger, H.L. Nucleotide sequence and secondary structure of citrus exocortis and Chrysanthemum stunt viroid. Eur. J. Biochem. 1982, 121, 249–257. [Google Scholar] [CrossRef]
- Cho, W.K.; Jo, Y.; Jo, K.M.; Kim, K.H. A current overview of two viroids that infect chrysanthemums: Chrysanthemum stunt viroid and Chrysanthemum chlorotic mottle viroid. Viruses 2013, 5, 1099–1113. [Google Scholar] [CrossRef]
- Palukaitis, P. Chrysanthemum stunt viroid. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Elsevier: London, UK, 2017; pp. 181–190. ISBN 9780128014981. [Google Scholar]
- Agribusiness intelligence/informa (IEG, FNP, Agrianual 2016). Available online: http://www.informafnpstore.com.br/agrianual-2016-pr-181-349650.htm (accessed on 16 March 2019).
- Flores e plantas ornamentais do Brasil (SEBRAE), v.1, Série Estudos Mercadológicos. Available online: http://www.bibliotecas.sebrae.com.br/chronus/ARQUIVOS_CHRONUS/bds/bds.nsf/7ed114f4eace9ea970dadf63bc8baa29/$File/5518.pdf (accessed on 10 March 2019).
- Bouwen, I.; Van Zaayen, A. Chrysanthemum Stunt Viroid. In Viroids; Hadidi, A., Flores, R., Randles, J.W., Semancik, J.S., Eds.; CSIRO Publishing: Collingwood, Australia, 2003; pp. 218–223. ISBN 0643067892. [Google Scholar]
- Horst, R.K.; Langhana, R.W.; Smith, S.H. Effects of chrysanthemum stunt, chlorotic mottle, aspermy and mosaic on flowering and rooting of chrysanthemums. Phytopathology 1977, 67, 9–14. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Baek, E.; Palukaitis, P. Are there strains of Chrysanthemum stunt viroid? J. Plant Pathol. 2012, 94, 697–701. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Palukaitis, P. Sequence comparisons of global Chrysanthemum stunt viroid variants: Multiple polymorphic positions scattered through the viroid genome. Virus Genes 2013, 46, 97–104. [Google Scholar] [CrossRef]
- Flores, R. Viroids in ornamentals. Acta Hortic. 2011, 901, 23–34. [Google Scholar] [CrossRef]
- Gobatto, D.; Eiras, M. Viroides em Ornamentais (Chapter 4). In Plantas Ornamentais: Doenças e Pragas; Alexandre, M.A.V., Duarte, L.M.L., Campos, A.E.C., Eds.; 1ª Edição. Devir Livraria: São Paulo, Brazil, 2017; pp. 171–199. ISBN 9788575326428. (In Portuguese) [Google Scholar]
- Matsushita, Y.; Tsukiboshi, T.; Ito, Y.; Chikuo, Y. Nucleotide sequences and distribution of Chrysanthemum stunt viroid in Japan. J. Jpn. Soc. Hort. Sci. 2007, 75, 333–337. [Google Scholar] [CrossRef]
- Menzel, W.; Maiss, E. Detection of chrysanthemum stunt viroid (CSVd) in cultivars of Argyranthemum frutescens by RT-PCR-ELISA. J. Plant Dis. Prot. 2000, 107, 548–552. [Google Scholar]
- Nakashima, A.; Hosokawa, M.; Maeda, S.; Yazawa, S. Natural infection of chrysanthemum stunt viroid in dahlia plants. J. Gen. Plant Pathol. 2007, 73, 225–227. [Google Scholar] [CrossRef]
- Verhoeven, J.T.J.; Arts, M.S.J.; Owens, R.A.; Roenhorst, J.W. Natural infection of petunia by chrysanthemum stunt viroid. Eur. J. Plant Pathol. 1998, 104, 383–386. [Google Scholar] [CrossRef]
- Runia, W.T.H.; Peters, D. The response of plant species used in agriculture and horticulture to viroid infections. Neth. J. Plant Pathol. 1980, 86, 135–146. [Google Scholar] [CrossRef]
- Barba, M.; Hadidi, A. Quarantine and certification for viroids and viroid diseases. In Viroids and Satellites; Hadidi, A., Flores, R., Randles, J.W., Palukaitis, P., Eds.; Elsevier: London, UK, 2017; pp. 415–423. ISBN 9780128014981. [Google Scholar]
- Alvarez-Díaz, J.C.; Ortiz-Echeverry, B.A.; Velásquez, N. Duplex RT-PCR assay for simultaneous detection of TSWV and CSVd in chrysanthemum. J. Virol. Methods 2019, 266, 41–48. [Google Scholar] [CrossRef]
- Brierley, P.; Smith, F.F. Chrysantemum stunt. Control measures affective against virus in florists’ crops. Flor. Rev. 1951, 107, 27–30. [Google Scholar]
- Song, A.; You, Y.; Chen, F.; Li, P.; Jiang, J.; Chen, S. A multiplex RT-PCR for rapid and simultaneous detection of viruses and viroids in chrysanthemum. Lett. Appl. Microbiol. 2012, 56, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Toguri, T.; Kudoh, H.; Okamura, M.; Momma, T.; Yoshika, M.; Kato, K.; Hagiwara, Y.; Sano, T. Double stranded RNA-specific ribonuclease confers tolerance against Chrysanthemum stunt viroid and Tomato spotted wilt virus in transgenic chrysanthemum plants. Breed. Sci. 2005, 55, 49–55. [Google Scholar] [CrossRef]
- Jo, K.M.; Jo, Y.; Choi, H.; Chu, H.; Lian, S.; Yoon, J.Y.; Choi, S.K.; Kim, K.H.; Cho, W.K. Development of genetically modified chrysanthemums resistant to Chrysanthemum stunt viroid using sense and antisense RNAs. Sci. Hortic. 2015, 195, 17–24. [Google Scholar] [CrossRef]
- Nabeshima, T.; Matsushita, Y.; Hosokawa, M. Chrysanthemum stunt viroid resistance in Chrysanthemum. Viruses 2018, 10, 719. [Google Scholar] [CrossRef]
- Iraklis, B.; Kanda, H.; Nabeshima, T.; Onda, M.; Ota, N.; Koeda, S.; Hosokawa, M. Digestion of chrysanthemum stunt viroid by leaf extracts of Capsicum chinense indicates strong RNA-digesting activity. Plant Cell Rep. 2016, 35, 1617–1628. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, Y.; Penmetcha, K.K.R. In vitro-transcribed Chrysanthemum stunt viroid RNA is infectiuos to Chrysanthemum and other plants. Phytopathology 2009, 99, 58–66. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Fitosociologia: Bases Para el Estudio de las Comunidades Vegetales, 3rd ed.; H. Blume: Madrid, Spain, 1979; ISBN 8472141748. (In Spanish) [Google Scholar]
- Horst, R.K.; Nelson, P.E. Compendium of Chrysanthemum Diseases; APS Press: St. Paul, MN, USA, 1997; ISBN 0890541779. [Google Scholar]
- Salzman, R.A.; Fujita, T.; Zhusalzman, K.; Hasegawa, P.M.; Bressan, R.A. An improved RNA isolation method for plant tissues containing high levels of phenolic compounds or carbohydrates. Plant Mol. Biol. Rep. 1999, 17, 11–17. [Google Scholar] [CrossRef]
- Codoñer, F.M.; Daròs, J.A.; Solé, R.V.; Elena, S.F. The fittest versus the flattest: Experimental confirmation of the quasispecies effect with subviral pathogens. PLoS Pathog. 2006, 2, e136. [Google Scholar] [CrossRef]
- Sanger, F.; Niclken, S.; Coulson, A.R. DNA sequencing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Zuker, M. On finding all suboptimal foldings of an RNA molecule. Science 1989, 244, 48–52. [Google Scholar] [CrossRef]
- Lorenzi, H. Plantas Daninhas do Brasil: Terrestres, Aquáticas, Parasitas e Tóxicas, 3rd ed.; Instituto Plantarium: Nova Odessa, Brazil, 2000; ISBN 8586714097. (In Portuguese) [Google Scholar]
- Colariccio, A.; Chaves, A.L.R.; Castro, L.C.; Oliveira, J.M. Natural occurrence of tospovirus on Emilia sagitatta in São Paulo, Brazil. Fitopatol. Bras. 1996, 21, 423. [Google Scholar]
- Brierley, P. Some experimental hosts of the chrysanthemum stunt virus. Plant Dis. Report. 1953, 37, 343–345. [Google Scholar]
- Franco, D.A.S.; Rozanski, A.; Almeida, S.D.B.; Matallo, M.B. Levantamento fitossociológico do agrião amargo (Cardamine bonariensis Pers.) em sistema de produção de crisântemo. O Biológico (Brazil) 2006, 68, 564–565. (In Portuguese) [Google Scholar]
- Henkel, J.; Sanger, H.L. Genbank Accession Z68201. 1995. Available online: https://www.ncbi.nlm.nih.gov/nuccore/Z68201 (accessed on 16 March 2019).
- Matsushita, Y. Chrysanthemum stunt viroid. Jpn. Agric. Res. Q. 2013, 47, 237–242. [Google Scholar] [CrossRef]
- Nie, X.Z.; Singh, R.P.; Bostan, H. Molecular cloning, secondary structure, and phylogeny of three pospiviroids from ornamental plants. Can. J. Plant Pathol. 2005, 27, 592–602. [Google Scholar] [CrossRef]
- Torchetti, E.M.; Navarro, B.; Trisciuzzi, V.N.; Nuccitelli, L.; Silletti, M.R.; Di Serio, F. First report of Chrysanthemum stunt viroid in Argyranthemum frutescens in Italy. J. Plant Pathol. 2012, 94, 451–454. [Google Scholar] [CrossRef]
- Mackie, A.E.; Rodoni, B.C.; Barbetti, M.J.; Mckirdy, S.J.; Jones, R.A.C. Potato spindle tuber viroid: Alternative host reservoirs and strain found in a remote subtropical irrigation area. Eur. J. Plant Pathol. 2016, 145, 433–446. [Google Scholar] [CrossRef]
- Adkar-Purushothama, C.R.; Chennappa, G.; Poornachandra Rao, K.; Sreenivasa, M.Y.; Maheshwar, P.K.; Nagendra Prasad, M.N.; Sano, T. Molecular diversity among viroids infecting chrysanthemum in India. Virus Genes 2017, 53, 636–642. [Google Scholar] [CrossRef]
- Choi, H.; Jo, Y.; Yoon, J.Y.; Choi, S.K.; Cho, W.K. Sequence variability of chrysanthemum stunt viroid in different chrysanthemum cultivars. PeerJ 2017, 5, e2933. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Family | Common Name | Infected/Inoculated Plants | RT-PCR | Symptoms * |
|---|---|---|---|---|
| Aeschynomene rudis/Fabaceae | Zigzag jointvetch | 0/4 | - | A |
| Ageratum conyzoides/Asteraceae | Billygoat-weed | 0/4 | - | A |
| Amaranthus deflexus/Amaranthaceae | Large-fruit amaranth | 0/4 | - | A |
| A. viridis/Amaranthaceae | Green amaranth | 2/4 | + | P (stunt) |
| Antirrhinum majus/Plantaginaceae | Snapdragon | 0/4 | - | A |
| Arachis pintoi/Fabaceae | Perennial Peanut | 0/4 | - | A |
| Bidens pilosa/Asteraceae | Hairy bidens | 0/4 | - | A |
| Brassica napus/Brassicacea | Summer rape | 0/4 | - | A |
| Cardamine bonariensis/Brassicacea | Cardamine (cress) | 4/4 | + | A |
| Cassia occidentalis/Fabaceae | Coffee senna | 0/4 | - | A |
| Catharanthus roseus/Apocynaceae | Rose periwinkle | 3/4 | + | A |
| Chamaesyce hirta/Euphorbiaceae | Snakeweed | 3/4 | + | A |
| C. prostata/Euphorbiaceae | Prostrate spurge | 0/4 | - | A |
| Commelina benghalensis/Commelinaceae | Benghal dayflower | 0/4 | - | A |
| Conyza bonariensis/Asteraceae | Hairy fleabane | 4/4 | + | A |
| Chrysanthemum carinatum/Asteraceae | Tricolor daisy | 4/4 | + | A |
| Cyperus rotundus/Cyperaceae | Nut Sedge | 0/4 | - | A |
| Digitaria insularis/Poaceae | Sourgrass | 0/4 | - | A |
| D. sanguinalis/Poaceae | Hairy crabgrass | 4/4 | + | A |
| Eclipta prostrata/Asteraceae | Eclipta | 0/4 | - | A |
| Eleusine indica/Poaceae | Indian goosegrass | 0/4 | - | A |
| Emilia sagittata/Asteraceae | Tasselflower | 4/4 | + | P (stunt) |
| Eschscholzia californica/Papaveraceae | California poppy | 0/4 | - | A |
| Fragaria vesca/Rosaceae | Woodland strawberry | 0/4 | - | A |
| Gomphrena globosa/Amaranthaceae | Globe Amaranth | 4/4 | + | A |
| Godetia grandiflora/Ericaceae | Clarkia amoena | 0/4 | - | A |
| Gypsophila elegans/Caryophyllaceae | Baby’s breath | 0/4 | - | A |
| Helianthus annuus/Asteraceae | Sunflower | 3/4 | + | A |
| Hemiscola aculeata/Cleomaceae | Prickly spider flower | 0/4 | - | A |
| Lobelia erinus/Campanulaceae | Lobelia | 0/4 | - | A |
| Lupinus polyphyllus/Fabaceae | Lupine | 3/4 | + | P (stunt) |
| Melampodium perfoliatum/Asteraceae | Blackfoots flower | 0/4 | - | A |
| Mirabilis jalapa/Nyctaginaceae | Four-clock flower | 3/4 | + | P (stunt) |
| Nicandra physalodes/Solanaceae | Shoo-fly plant | 0/4 | - | A |
| Nicotiana benthamiana/Solanaceae | Tobacco | 0/4 | - | A |
| N. megalosiphon/Solanaceae | Tobacco | 0/4 | - | A |
| N. occidentalis/Solanaceae | Nightshade | 0/4 | - | A |
| N. rustica/Solanaceae | Aztec tobacco | 0/4 | - | A |
| N. tabacum “Samsun”/Solanaceae | Tobacco | 0/4 | - | A |
| N. tabacum “Xanthi”/Solanaceae | Tobacco | 0/4 | - | A |
| Oxalis corniculata/Oxalidaceae | Creeping woodsorrel | 0/4 | - | A |
| O. latifolia/Oxalidaceae | Broadleaf woodsorrel | 4/4 | + | P (mosaic) |
| Portulaca oleracea/Portulacaceae | Common purslane | 4/4 | + | A |
| Raphanus raphanistrum/Brassicaceae | Wild radish | 0/4 | - | A |
| Ruta graveolens/Rutaceae | Rue | 0/4 | - | A |
| Sagina chilensis/Caryophyllaceae | Pearlworts | 0/4 | - | A |
| Senecio cruentus/Asteraceae | Cineraria | 2/4 | + | A |
| Solanum americanum/Solanaceae | Black nightshade | 0/4 | - | A |
| S. lycopersicum “Rutgers”/Solanaceae | Tomato | 4/4 | + | P (stunt) |
| S. tuberosum “Ágata”/Solanaceae | Potato | 4/4 | + | A |
| Sonchus oleraceus/Asteraceae | Sowthistle | 0/4 | - | A |
| Tropaeolum majus/Tropaeolaceae | Garden nasturtium | 0/4 | - | A |
| Viola tricolor/Violaceae | Johnny jump-ups | 0/4 | - | A |
| Xanthium strumarium/Asteraceae | Rough cocklebur | 0/4 | - | A |
| Zinnia elegans/Asteraceae | Common zinnia | 0/4 | - | A |
| Z. elegans “Lilliput”/Asteraceae | Zinnia | 0/4 | - | A |
| Name | Origin | Genbank Code | Genome Size |
|---|---|---|---|
| 2215-BR | Artur Nogueira, SP, Brazil | KU535579 | 354 |
| 2216-BR | Artur Nogueira, SP, Brazil | KU535580 | 354 |
| 2217-BR | Artur Nogueira, SP, Brazil | KU535581 | 354 |
| 2218-BR | Artur Nogueira, SP, Brazil | KU535582 | 354 |
| 2219-BR | Artur Nogueira, SP, Brazil | KU535583 | 354 |
| 2220-BR | Artur Nogueira, SP, Brazil | KU535584 | 354 |
| 2221-BR | Artur Nogueira, SP, Brazil | KU535585 | 354 |
| 2222-BR | Paranapanema, SP, Brazil | MF359692 | 354 |
| 2223-BR | Artur Nogueira, SP, Brazil | MF359693 | 355 |
| 2224-BR | Artur Nogueira, SP, Brazil | MF359694 | 354 |
| 2225-BR | Artur Nogueira, SP, Brazil | MF359695 | 356 |
| 2226-BR | Artur Nogueira, SP, Brazil | MF359696 | 356 |
| 2227-BR | Artur Nogueira, SP, Brazil | MF359697 | 354 |
| 2228-BR | Artur Nogueira, SP, Brazil | MF359698 | 354 |
| 2229-BR | Artur Nogueira, SP, Brazil | MF359699 | 354 |
| 2230-BR | Artur Nogueira, SP, Brazil | MF359700 | 356 |
| 2231-BR | Artur Nogueira, SP, Brazil | MF359701 | 354 |
| 2232-BR | Artur Nogueira, SP, Brazil | MF359702 | 354 |
| 2233-BR | Artur Nogueira, SP, Brazil | MF359703 | 354 |
| 2234-BR | Artur Nogueira, SP, Brazil | MF359704 | 354 |
| 2235-BR | Artur Nogueira, SP, Brazil | MF359705 | 354 |
| 2236-BR | Artur Nogueira, SP, Brazil | MF359706 | 354 |
| 2237-BR | Artur Nogueira, SP, Brazil | MF359707 | 354 |
| 2238-BR | Artur Nogueira, SP, Brazil | MF359708 | 354 |
| 2239-BR | Artur Nogueira, SP, Brazil | MF359709 | 354 |
| 2240-BR | Artur Nogueira, SP, Brazil | MF359710 | 354 |
| 2241-BR | Artur Nogueira, SP, Brazil | MF359711 | 354 |
| 2242-CO | Rionegro, Antioquia, Colombia | MF359712 | 354 |
| 2243-CO | Rionegro, Antioquia, Colombia | MF359713 | 354 |
| 2244-CO | Rionegro, Antioquia, Colombia | MF359714 | 354 |
| 2245-CO | Rionegro, Antioquia, Colombia | MF359715 | 354 |
| 2246-CO | Rionegro, Antioquia, Colombia | MF359716 | 354 |
| 2247-CO * | Rionegro, Antioquia, Colombia | MF359717 | 354 |
| Sequence Variants and Access Code in Genbank | Identical Sequence Variants |
|---|---|
| JP.2 (AB055974) | None |
| 2241-BR (MF359711) | 2248/2250/2251/2252/2253 |
| 2214-BR (JX909290) | None |
| CN.1 (JQ809255) | None |
| DE.2 (Z68201) | None |
| US.3 (U82445) | None |
| 2234-BR (MF359704) | 2235/2236/2237/2238/2239/2240/2242/2243/2244/2245/2246/2247 |
| 2230-BR (MF359700) | 2231/2232 |
| 2228-BR (MF359698) | None |
| 2222-BR (MF359706) | None |
| 2229-BR (MF359699) | None |
| 2226-BR (MF359696) | None |
| 2223-BR (MF359693) | None |
| 2225-BR (MF359695) | None |
| 2227-BR (MF359697) | 2218/2219/2220/2233 |
| CN.3 (HQ891018) | CN.4 |
| FR (JF938538) | None |
| JP.5 (LC090814) | None |
| KR.1 (LC088322) | None |
| US.1 (JQ895296) | US.3 a US.40 |
| JP.1 (D88895) | JP.6/JP.7 |
| AT (JF414238) | None |
| BE.1 (HG739080) | None |
| IN.1 (FN669536) | IN.3/IN.4/IN.5/IN.6 |
| NL (DQ406591) | None |
| CA (DQ094298) | None |
| US.2 (AJ001849) | US.41 a US.64 |
| DE.3 (X16408) | DE.1/DE.4 |
| 2221-BR (KU535585) | None |
| IN.2 (FN673553) | IN.7/IN.8/IN.9/IN.10/IN.11/IN.12/IN.13/IN.14 |
| CN.2 (HQ891019) | Nenhum |
| JP.4 (LC090813) | JP.8/JP.9/JP.10/JP.11/JP.12 |
| BE.2 (HG739079) | BE.3/BE.4 |
| 2217-BR (KU535581) | None |
| IT.1 (JQ685734) | IT.2/IT.3/IT.4/IT.5/IT.6/IT.7/IT.8/IT.9/IT.10/AU.1/AU.2/AU.3 |
| KR.2 (KC345440) | KR.3/KR.4 |
| HU (AJ000046) | None |
| JP.3 (AB279768) | JP.13/JP.14/JP.15/JP.16/JP.17 |
| 2255-ES (not deposited in Genbank) | None |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gobatto, D.; de Oliveira, L.A.; de Siqueira Franco, D.A.; Velásquez, N.; Daròs, J.-A.; Eiras, M. Surveys in the Chrysanthemum Production Areas of Brazil and Colombia Reveal That Weeds Are Potential Reservoirs of Chrysanthemum Stunt Viroid. Viruses 2019, 11, 355. https://doi.org/10.3390/v11040355
Gobatto D, de Oliveira LA, de Siqueira Franco DA, Velásquez N, Daròs J-A, Eiras M. Surveys in the Chrysanthemum Production Areas of Brazil and Colombia Reveal That Weeds Are Potential Reservoirs of Chrysanthemum Stunt Viroid. Viruses. 2019; 11(4):355. https://doi.org/10.3390/v11040355
Chicago/Turabian StyleGobatto, Danielle, Lucas Araújo de Oliveira, Daniel Andrade de Siqueira Franco, Nubia Velásquez, José-Antonio Daròs, and Marcelo Eiras. 2019. "Surveys in the Chrysanthemum Production Areas of Brazil and Colombia Reveal That Weeds Are Potential Reservoirs of Chrysanthemum Stunt Viroid" Viruses 11, no. 4: 355. https://doi.org/10.3390/v11040355