
Influence of African Swine Fever Virus on Host Gene Transcription within Peripheral Blood Mononuclear Cells from Infected Pigs
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. RNA Purification from PBMCs and Gene Expression Analysis
2.2. Mapping of Sequence Reads to the Pig Genome
2.3. Differential Gene Expression Analyses
2.4. High-Throughput RT-qPCR
3. Results
3.1. Principal Component Analysis (PCA)
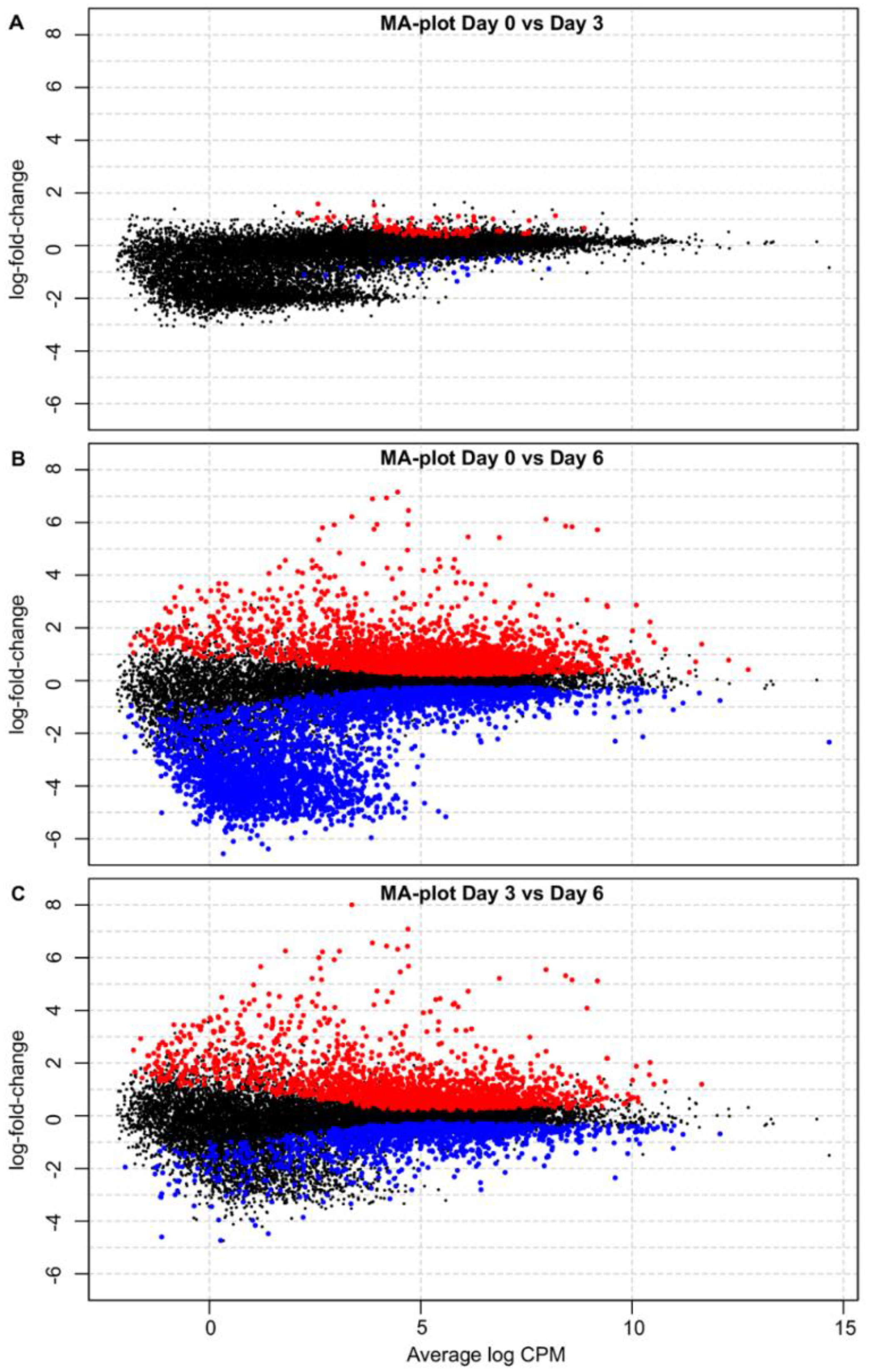
3.2. Differential Gene Expression Analysis
3.3. Changes in Host Gene Expression in PBMCs from ASFV-Infected Pigs
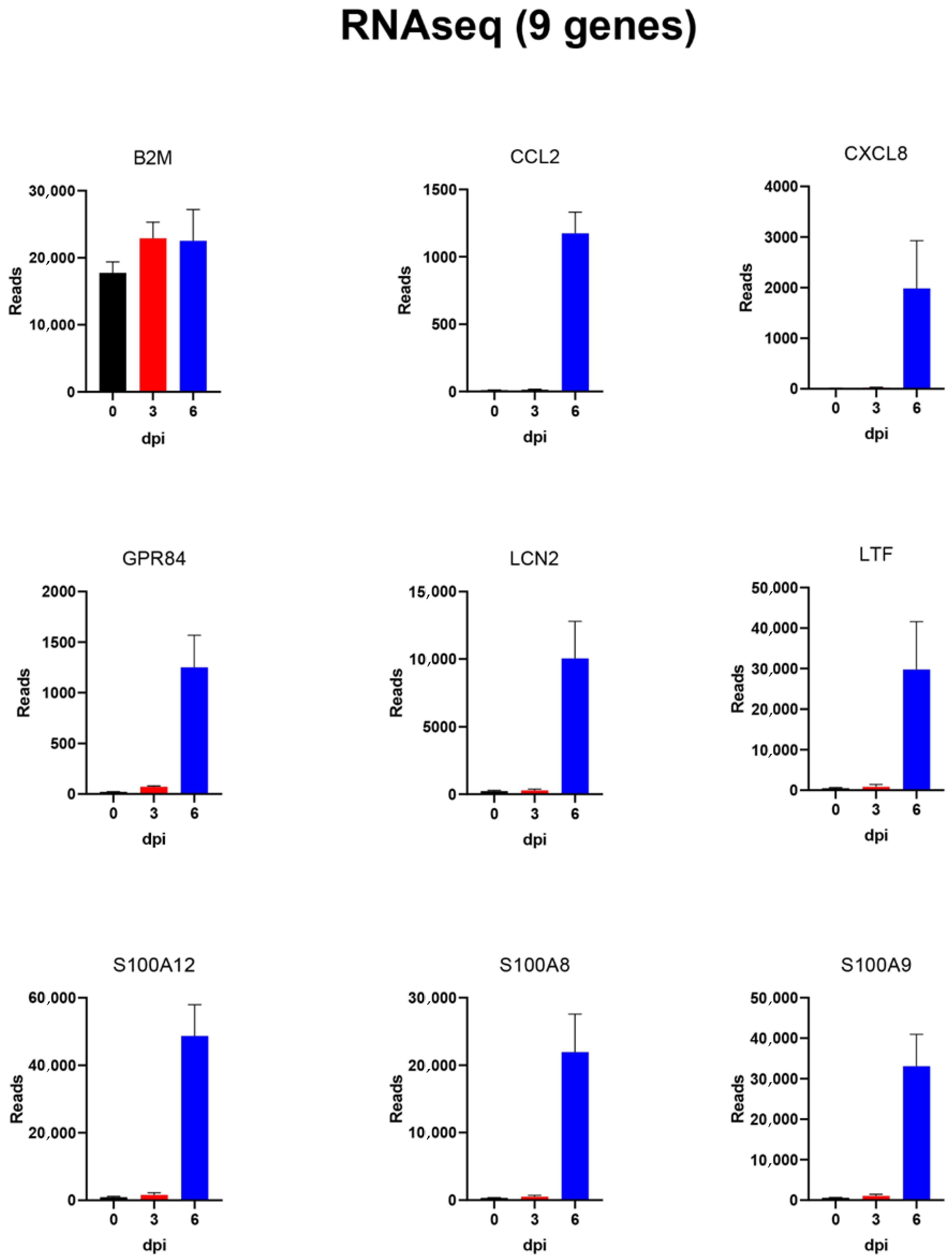
3.4. Analysis of the Expression of Individual Genes within PBMCs throughout the Course of ASFV Infection in Pigs
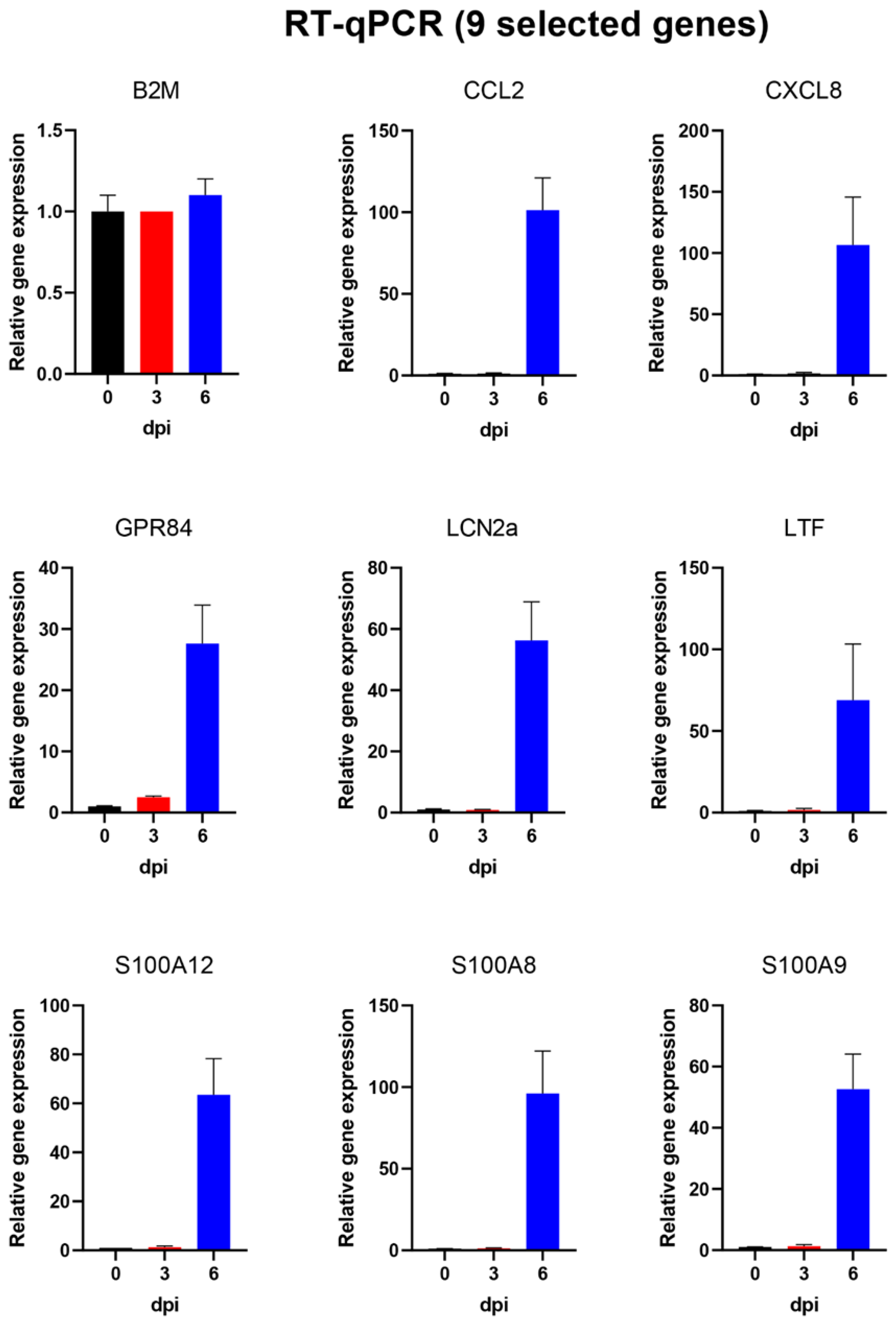
3.5. Changes in Selected Host Gene Expression in PBMCs from Pigs That Were Infected with ASFV
3.6. Analysis of Changes in Gene Expression in PBMCs Using High-Throughput Microfluidic qPCR Assays
4. Discussion
4.1. Key Changes in Host Gene Expression
4.2. Different Approaches to Assessing Changes in Host Gene Expression in Cells following ASFV Infection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Urbano, A.; Forth, J.H.; Olesen, A.S.; Dixon, L.; Rasmussen, T.B.; Cackett, G.; Werner, F.; Karger, A.; Andrés, G.; Wang, X.; et al. African swine fever virus: Cellular and molecular aspects. In Understanding and Combatting African Swine Fever; Iacolina, L., Penrith, M.-L., Bellini, S., Chenais, E., Jori, F., Montoya, M., Ståhl, K., Gavier-Widén, D., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2021; pp. 25–61. [Google Scholar] [CrossRef]
- Dixon, L.K.; Sun, H.; Roberts, H. African swine fever. Antivir. Res. 2019, 165, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Olesen, A.S.; Belsham, G.J.; Rasmussen, T.B.; Lohse, L.; Bødker, R.; Halasa, T.; Boklund, A.; Bøtner, A. Potential routes for indirect transmission of African swine fever virus into domestic pig herds. Transbound. Emerg. Dis. 2020, 67, 1472–1484. [Google Scholar] [CrossRef]
- Achenbach, J.E.; Gallardo, C.; Nieto-Pelegrín, E.; Rivera-Arroyo, B.; Degefa-Negi, T.; Arias, M.; Jenberie, S.; Mulisa, D.D.; Gizaw, D.; Gelaye, E.; et al. Identification of a new genotype of African swine fever virus in domestic pigs from Ethiopia. Transbound. Emerg. Dis. 2017, 64, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.D.; Penrith, M.L.; Crucière, C.; Edrich, J.L.; Hutchings, G.; Roger, F.; Couacy-Hymann, E.R.; Thomson, G. Genotyping field strains of African swine fever virus by partial p72 gene characterisation. Arch. Virol. 2003, 148, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Boshoff, C.I.; Bastos, A.D.S.; Gerber, L.J.; Vosloo, W. Genetic characterisation of African swine fever viruses from outbreaks in southern Africa (1973–1999). Vet. Microbiol. 2007, 121, 45–55. [Google Scholar] [CrossRef]
- Lubisi, B.A.; Bastos, A.D.S.; Dwarka, R.M.; Vosloo, W. Molecular epidemiology of African swine fever in East Africa. Arch. Virol. 2005, 150, 2439–2452. [Google Scholar] [CrossRef]
- Quembo, C.J.; Jori, F.; Vosloo, W.; Heath, L. Genetic characterization of African swine fever virus isolates from soft ticks at the wildlife/domestic interface in Mozambique and identification of a novel genotype. Transbound. Emerg. Dis. 2018, 65, 420–431. [Google Scholar] [CrossRef]
- Costard, S.; Wieland, B.; de Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.U.; Dixon, L.K. African swine fever: How can global spread be prevented? Philos. Trans. R Soc. Lond. B Biol. Sci. 2009, 364, 2683–2696. [Google Scholar] [CrossRef]
- OIE WAHIS Interface. 2021. Available online: https://wahis.oie.int/#/dashboards/country-or-disease-dashboard (accessed on 19 August 2021).
- FAO Website. Agriculture and Consumer Protection Department, Animal Production and Health. ASF Situation in Asia Update. 2020. Available online: http://www.fao.org/ag/againfo/programmes/en/empres/ASF/situation_update.html (accessed on 3 March 2020).
- Busch, F.; Haumont, C.; Penrith, M.-L.; Laddomada, A.; Dietze, K.; Globig, A.; Guberti, V.; Zani, L.; Depner, K. Evidence-based African swine fever policies: Do we address virus and host adequately? Front. Vet. Sci. 2021, 8, 637487. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Montoya, M.; Reis, A.L.; Dixon, L.K. African swine fever: A reemerging viral disease threatening the global pig industry. Vet. J. 2018, 233, 41–48. [Google Scholar] [CrossRef]
- Anon. Vaccine targets African swine fever. Science 2022, 376, 1140. [Google Scholar]
- Gómez-Villamandos, J.C.; Bautista, M.J.; Sánchez-Cordón, P.J.; Carrasco, L. Pathology of African swine fever: The role of monocyte-macrophage. Virus Res. 2013, 173, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Olesen, A.S.; Kodama, M.; Lohse, L.; Accensi, F.; Rasmussen, T.B.; Lazov, C.M.; Limborg, M.T.; Gilbert, M.T.P.; Bøtner, A.; Belsham, G.J. Identification of African Swine Fever Virus Transcription within Peripheral Blood Mononuclear Cells of Acutely Infected Pigs. Viruses 2021, 13, 2333. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Warr, A.; Affara, N.; Aken, B.; Beiki, H.; Bickhart, D.M.; Billis, K.; Chow, W.; Eory, L.; Finlayson, H.A.; Flicek, P.; et al. An improved pig reference genome sequence to enable pig genetics and genomics research. GigaScience 2020, 9, giaa051. [Google Scholar] [CrossRef]
- Smid, M.; van de Werken, H.J.G.; van Riet, J.; van Galen, A.; de Weerd, V.; van der Vlugt-Daane, M.; Bril, S.I.; Lalmahomed, Z.S.; Kloosterman, W.P.; Wilting, S.M.; et al. Gene length corrected trimmed mean of M-values (GeTMM) processing of RNA-seq data performs similarly in intersample analyses while improving intrasample comparisons. BMC Bioinform. 2018, 19, 236. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From reads to genes to pathways: Differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 2016, 5, 1438. [Google Scholar]
- Ritchie, M.E.; Phipson, B.; Wu, D.I.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Brogaard, L.; Heegaard, P.M.; Larsen, L.E.; Mortensen, S.; Schlegel, M.; Dürrwald, R.; Skovgaard, K. Late regulation of immune genes and microRNAs in circulating leukocytes in a pig model of influenza A (H1N2) infection. Sci. Rep. 2016, 6, 21812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skovgaard, K.; Cirera, S.; Vasby, D.; Podolska, A.; Breum, S.Ø.; Dürrwald, R.; Schlegel, M.; Heegaard, P.M. Expression of innate immune genes, proteins and microRNAs in lung tissue of pigs infected experimentally with influenza virus (H1N2). Innate Immun. 2013, 19, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Cackett, G.; Portugal, R.; Matelska, D.; Dixon, L.; Werner, F. African swine fever virus and host response: Transcriptome profiling of the Georgia 2007/1 strain and porcine macrophages. J. Virol. 2022, 96, e0193921. [Google Scholar] [CrossRef] [PubMed]
- Jaing, C.; Rowland, R.R.R.; Allen, J.E.; Certoma, A.; Thissen, J.B.; Bingham, J.; Rowe, B.; White, J.R.; Wynne, J.W.; Johnson, D.; et al. Gene expression analysis of whole blood RNA from pigs infected with low and high pathogenic African swine fever viruses. Sci. Rep. 2017, 7, 10115. [Google Scholar] [CrossRef] [PubMed]
- Fishbourne, E.; Hutet, E.; Abrams, C.; Cariolet, R.; Le Potier, M.-F.; Takamatsu, H.-H.; Dixon, L.K. Increase in chemokines CXCL10 and CCL2 in blood from pigs infected with high compared to low virulence African swine fever virus isolates. Vet. Res. 2013, 44, 87. [Google Scholar] [CrossRef]
- Afonso, C.L.; Piccone, M.E.; Zaffuto, K.M.; Neilan, J.; Kutish, G.F.; Lu, Z.; Balinsky, C.A.; Gibb, T.R.; Bean, T.J.; Zsak, L.; et al. African swine fever virus multigene family 360 and 530 genes affect host interferon response. J. Virol. 2004, 78, 1858–1864. [Google Scholar] [CrossRef]
- Gil, S.; Sepúlveda, N.; Albina, E.; Leitão, A.; Martins, C. The low-virulent African swine fever virus (ASFV/NH/P68) induces enhanced expression and production of relevant regulatory cytokines (IFNalpha, TNFalpha and IL12p40) on porcine macrophages in comparison to the highly virulent ASFV/L60. Arch. Virol. 2008, 153, 1845–1854. [Google Scholar] [CrossRef]
- Karalyan, Z.; Zakaryan, H.; Sargsyan, K.; Voskanyan, H.; Arzumanyan, H.; Avagyan, H.; Karalova, E. Interferon status and white blood cells during infection with African swine fever virus in vivo. Vet. Immunol. Immunopathol. 2012, 145, 551–555. [Google Scholar] [CrossRef]
- Golding, J.P.; Goatley, L.; Goodbourn, S.; Dixon, L.K.; Taylor, G.; Netherton, C.L. Sensitivity of African swine fever virus to type I interferon is linked to genes within multigene families 360 and 505. Virology 2016, 493, 154–161. [Google Scholar] [CrossRef]
- Cackett, G.; Matelska, D.; Sýkora, M.; Portugal, R.; Malecki, M.; Bähler, J.; Dixon, L.; Werner, F. The African Swine Fever Virus Transcriptome. J. Virol. 2020, 94, e00119–e00120. [Google Scholar] [CrossRef]
- Xia, C.; Braunstein, Z.; Toomey, A.C.; Zhong, J.; Rao, X. S100 proteins as an important regulator of macrophage inflammation. Front. Immunol. 2018, 8, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberdi, A.; Andersen, S.B.; Limborg, M.T.; Dunn, R.R.; Gilbert, M.T.P. Disentangling host–microbiota complexity through hologenomics. Nat. Rev. Genet. 2022, 23, 281–297. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Mean Reads 0 dpi | SEM | Mean Reads 3 dpi | SEM | Mean Reads 6 dpi | SEM |
|---|---|---|---|---|---|---|
| B2M | 17,770 | 1617 | 22,921 | 2379 | 22,529 | 4663 |
| C1QB | 34 | 10 | 43 | 10 | 476 | 85 |
| C1QBP | 1554 | 194 | 2175 | 216 | 3049 | 249 |
| C1QC | 40 | 10 | 39 | 9 | 581 | 89 |
| CCL2 | 11 | 3 | 15 | 3 | 1174 | 158 |
| CCL4 | 413 | 37 | 417 | 43 | 1269 | 65 |
| CD101 | 204 | 22 | 218 | 22 | 478 | 120 |
| CD14 | 1462 | 234 | 2491 | 535 | 4529 | 989 |
| CD163 | 1348 | 282 | 2176 | 482 | 9191 | 2069 |
| CD68 | 497 | 101 | 603 | 103 | 1936 | 382 |
| CXCL8 | 13 | 1 | 26 | 4 | 1984 | 946 |
| CXCL9 | 9 | 4 | 10 | 2 | 87 | 23 |
| CXCR2 | 340 | 106 | 501 | 177 | 4429 | 1481 |
| RIG-I (DDX58) | 1139 | 204 | 3195 | 507 | 9131 | 1674 |
| GPR84 | 23 | 2 | 73 | 9 | 1251 | 317 |
| IFNG | 58 | 8 | 87 | 11 | 202 | 33 |
| IL1A | 31 | 8 | 17 | 3 | 318 | 32 |
| IL1RAP | 559 | 44 | 571 | 59 | 5142 | 1596 |
| IRF3 | 1476 | 155 | 2004 | 183 | 2137 | 157 |
| IRF7 | 2176 | 549 | 5297 | 1006 | 11,558 | 1163 |
| ISG15 | 1868 | 748 | 3332 | 632 | 14,387 | 1723 |
| ISG20 | 580 | 155 | 1417 | 240 | 5877 | 1077 |
| LCN2 | 233 | 56 | 297 | 79 | 10,041 | 2749 |
| LTF | 516 | 182 | 847 | 601 | 29,764 | 11,781 |
| MX1 | 9184 | 2963 | 21,212 | 3847 | 68,391 | 7402 |
| MX2 | 1845 | 604 | 2820 | 554 | 8547 | 1365 |
| OAS1 | 1681 | 693 | 3532 | 950 | 16,605 | 2317 |
| OAS2 | 6640 | 1780 | 11,464 | 1791 | 24,732 | 4024 |
| OASL | 90 | 11 | 114 | 17 | 659 | 111 |
| PPIA | 26,158 | 3235 | 33,908 | 3636 | 43,087 | 3728 |
| RPL13A | 46,135 | 4072 | 55,115 | 4428 | 34,299 | 1795 |
| S100A12 | 946 | 213 | 1612 | 605 | 48,702 | 9254 |
| S100A8 | 311 | 80 | 524 | 190 | 21,969 | 5629 |
| S100A9 | 577 | 116 | 1039 | 380 | 33,094 | 7872 |
| TNF | 109 | 11 | 184 | 21 | 453 | 86 |
| VWF | 1083 | 272 | 1332 | 426 | 3722 | 1223 |
| Gene 1 | Mean RE 0 dpi | SEM | Mean RE 3 dpi | SEM | Mean RE 6 dpi | SEM |
|---|---|---|---|---|---|---|
| B2M | 1.0 | 0.1 | 1.0 | 0 | 1.1 | 0.1 |
| C1QB | 1.0 | 0.2 | 1.4 | 0.4 | 7.2 | 1.4 |
| C1QBP | 1.0 | 0.1 | 1.1 | 0 | 1.3 | 0.1 |
| C1QC | 1.0 | 0.1 | 1.0 | 0 | 7.8 | 2.2 |
| CCL2 | 1.0 | 0.3 | 1.2 | 0.4 | 101.3 | 19.8 |
| CCL4 | 1.0 | 0.4 | 0.6 | 0.1 | 1.5 | 0.1 |
| CD101 | 1.0 | 0.2 | 1.3 | 0.1 | 1.3 | 0.2 |
| CD14 | 1.0 | 0.1 | 1.4 | 0.3 | 2.4 | 0.3 |
| CD163 | 1.0 | 0.2 | 1.5 | 0.4 | 6.2 | 1.2 |
| CD68a * | 1.0 | 0.1 | 1.1 | 0.2 | 4.9 | 0.7 |
| CD68b * | 1.0 | 0.2 | 1.2 | 0.2 | 5.2 | 0.8 |
| CXCL8 | 1.0 | 0.2 | 1.7 | 0.8 | 106.6 | 39.1 |
| CXCL9 | 1.0 | 0.2 | 0.6 | 0.2 | 10.8 | 2.5 |
| CXCR2 | 1.0 | 0.3 | 1.0 | 0.3 | 8.5 | 1.9 |
| RIG-I (DDX58) | 1.0 | 0.4 | 2.9 | 0.5 | 13.5 | 2.5 |
| GPR84 | 1.0 | 0.1 | 2.5 | 0.2 | 27.6 | 6.3 |
| IFNG | 1.0 | 0.2 | 1.2 | 0.1 | 2.9 | 0.3 |
| IL1A | 1.0 | 0.3 | 0.4 | 0.2 | 8.2 | 1.7 |
| IL1RAP | 1.0 | 0.2 | 1.2 | 0.1 | 9.1 | 1.2 |
| IRF3 | 1.0 | 0.3 | 0.9 | 0.1 | 1.1 | 0.1 |
| IRF7 | 1.0 | 0.4 | 2.0 | 0.4 | 4.5 | 0.5 |
| ISG15 | 1.0 | 0.4 | 1.8 | 0.4 | 9.1 | 1.3 |
| ISG20 | 1.0 | 0.4 | 2.1 | 0.4 | 12.4 | 2.0 |
| LCN2a * | 1.0 | 0.2 | 0.8 | 0.2 | 43.2 | 8.1 |
| LCN2b * | 1.0 | 0.2 | 0.8 | 0.2 | 56.3 | 12.6 |
| LTF | 1.0 | 0.3 | 1.5 | 1.1 | 68.9 | 34.4 |
| MX1 | 1.0 | 0.5 | 1.8 | 0.2 | 4.9 | 0.7 |
| MX2 | 1.0 | 0.4 | 1.5 | 0.2 | 2.1 | 0.7 |
| OAS1 | 1.0 | 0.5 | 1.7 | 0.3 | 9.9 | 2.1 |
| OAS2 | 1.0 | 0.4 | 1.8 | 0.1 | 2.2 | 0.5 |
| OASL | 1.0 | 0.2 | 1.3 | 0.4 | 2.7 | 0.3 |
| PPIA | 1.0 | 0.1 | 1.0 | 0.1 | 1.3 | 0.1 |
| RPL13A | 1.0 | 0 | 0.9 | 0.1 | 0.4 | 0.1 |
| S100A12 | 1.0 | 0 | 1.3 | 0.5 | 63.5 | 14.8 |
| S100A8 | 1.0 | 0.1 | 1.2 | 0.4 | 96.0 | 26.0 |
| S100A9 | 1.0 | 0.1 | 1.3 | 0.5 | 52.6 | 11.5 |
| TNF | 1.0 | 0.2 | 1.3 | 0.2 | 1.2 | 0.2 |
| VWF | 1.0 | 0.4 | 0.9 | 0.3 | 1.7 | 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olesen, A.S.; Kodama, M.; Skovgaard, K.; Møbjerg, A.; Lohse, L.; Limborg, M.T.; Bøtner, A.; Belsham, G.J. Influence of African Swine Fever Virus on Host Gene Transcription within Peripheral Blood Mononuclear Cells from Infected Pigs. Viruses 2022, 14, 2147. https://doi.org/10.3390/v14102147
Olesen AS, Kodama M, Skovgaard K, Møbjerg A, Lohse L, Limborg MT, Bøtner A, Belsham GJ. Influence of African Swine Fever Virus on Host Gene Transcription within Peripheral Blood Mononuclear Cells from Infected Pigs. Viruses. 2022; 14(10):2147. https://doi.org/10.3390/v14102147
Chicago/Turabian StyleOlesen, Ann Sofie, Miyako Kodama, Kerstin Skovgaard, Ask Møbjerg, Louise Lohse, Morten T. Limborg, Anette Bøtner, and Graham J. Belsham. 2022. "Influence of African Swine Fever Virus on Host Gene Transcription within Peripheral Blood Mononuclear Cells from Infected Pigs" Viruses 14, no. 10: 2147. https://doi.org/10.3390/v14102147