

SARS-CoV-2 at the Human–Animal Interface: Implication for Global Public Health from an African Perspective

 , , , ,
, , , ,
Abstract
:1. Introduction
2. The Probable Origin of SARS-CoV-2 Points to Animals
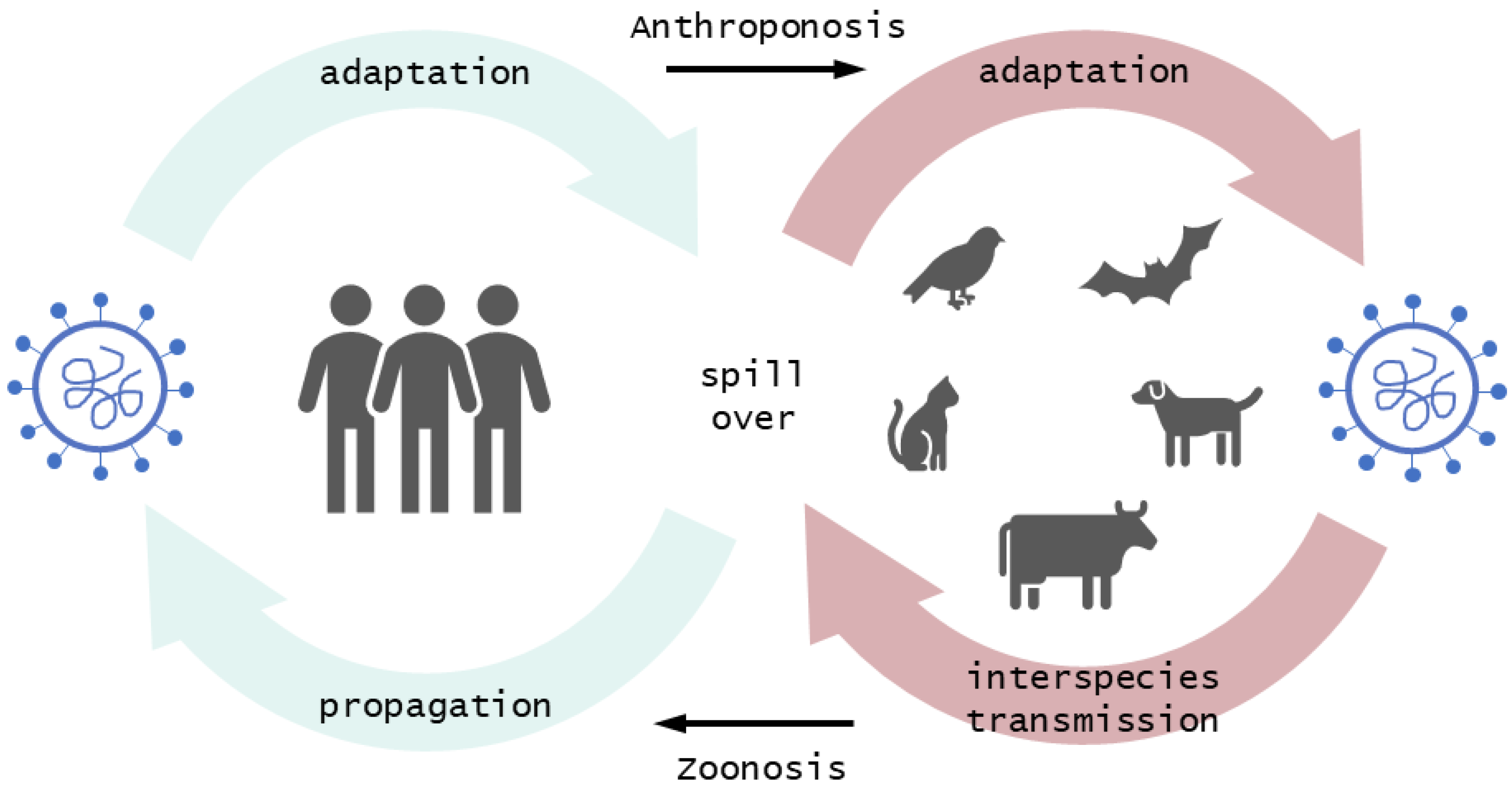
2.1. Mutations of SARS-CoV-2 Are a Common Feature Facilitating the Crossing of Interspecies Barriers
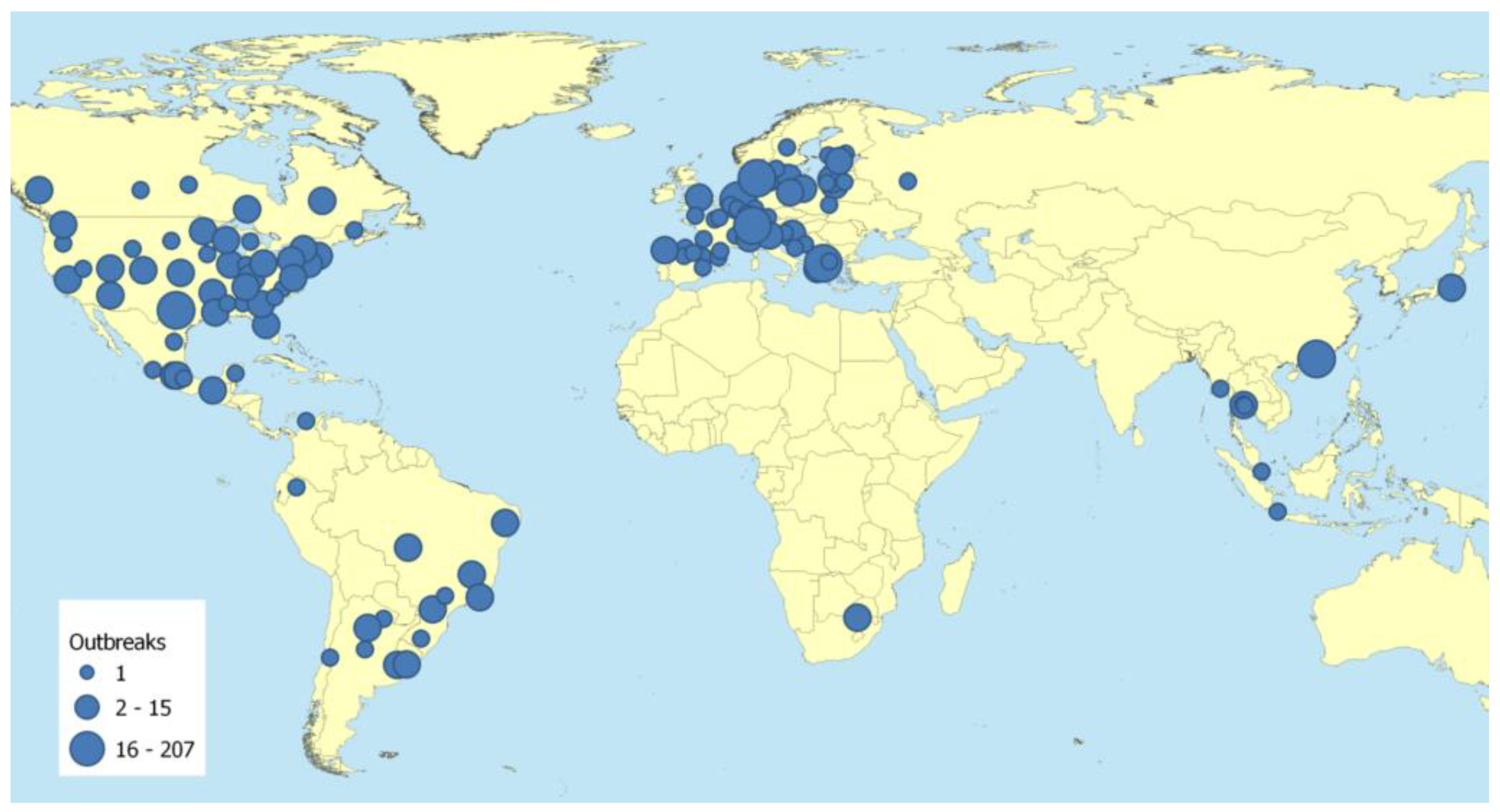
2.2. Frequent Transmissions of SARS-CoV-2 to Animals may Create Future Animal Reservoir Hosts
3. Effective Spillover at the Human–Animal Interface Marks the Entry Point of Novel Infectious Diseases
Spillover Transmission Is Driven by Human Activity
4. Compounding a Bad Situation: The Impact of Zoonotic Diseases in Africa with a Focus on Nigeria
4.1. SARS-CoV-2 Infection Detection Rate in the African Population Is Lower Compared to the Rest of the World
4.2. Socioeconomic and Ecological Factors in Africa Enhance the Probability of Anthropozoonotic Transmission of SARS-CoV-2
4.3. Limited Access to Vaccines Increases the Vulnerability of the Population
4.4. Human-to-Animal Transmission May Happen Undetected due to Limited Implementation of Biosecurity and Active Surveillance in Animal Holdings
5. Conclusions: Mitigating Future Respiratory Virus Pandemics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shang, Y.; Li, H.; Zhang, R. Effects of Pandemic Outbreak on Economies: Evidence from Business History Context. Front. Public Health 2021, 9, 632043. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO-Convened Global Study of Origins of SARS-CoV-2: China Part Joint WHO-China Study. Available online: https://www.who.int/publications/i/item/who-convened-global-study-of-origins-of-sars-cov-2-china-part (accessed on 17 July 2022).
- World Organization for Animal Health (WOAH). SARS-CoV-2 in Animals—Situation. 2022. Available online: https://www.woah.org/app/uploads/2022/02/sars-cov-2-situation-report-9.pdf (accessed on 3 August 2022).
- Fenollar, F.; Mediannikov, O.; Maurin, M.; Devaux, C.; Colson, P.; Levasseur, A.; Fournier, P.E.; Raoult, D. Mink, SARS-CoV-2, and the Human-Animal Interface. Front. Microbiol. 2021, 12, 663815. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H.; Latham, S.M.; Woolhouse, M.E. Risk factors for human disease emergence. Phil. Trans. R. Soc. Lond. B 2001, 356, 983–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, N.D.; Dunavan, C.P.; Diamond, J. Origins of major human infectious diseases. Nature 2007, 447, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Demmler, G.J.; Ligon, B.L. Severe acute respiratory syndrome (SARS): A review of the history, epidemiology, prevention, and concerns for the future. Semin. Pediatr. Infect. Dis. 2003, 14, 240–244. [Google Scholar] [CrossRef]
- Stawicki, S.P.; Arquilla, B.; Galwankar, S.C.; Hoey, B.A.; Jahre, J.A.; Kalra, S.; Kelkar, D.; Papadimos, T.J.; Sabol, D.; Sharpe, R.P. The emergence of ebola as a global health security threat: From ‘lessons learned’ to coordinated multilateral containment efforts. J. Glob. Infect. Dis. 2014, 6, 164–177. [Google Scholar] [CrossRef]
- Ligon, B.L. Monkeypox: A review of the history and emergence in the Western hemisphere. Semin. Pediatr. Infect. Dis. 2004, 15, 280–287. [Google Scholar] [CrossRef]
- Epstein, J.H.; Field, H.E.; Luby, S.; Pulliam, J.R.; Daszak, P. Nipah virus: Impact, origins, and causes of emergence. Curr. Infect. Dis. Rep. 2006, 8, 59–65. [Google Scholar] [CrossRef]
- Morens, D.M.; Fauci, A.S. Emerging infectious diseases: Threats to human health and global stability. PLoS Pathog. 2013, 9, e1003467. [Google Scholar] [CrossRef] [Green Version]
- Gu, D.; Andreev, K.; Dupre, M.E. Major Trends in Population Growth Around the World. China CDC Wkly. 2021, 3, 604–613. [Google Scholar] [CrossRef]
- Byrnes, B.; Bumb, B. Population Growth, Food Production and Nutrient Requirements. J. Crop. Prod. 2008, 1, 1–27. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife—Threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Johnson, C.K.; Hitchens, P.L.; Evans, T.S.; Goldstein, T.; Thomas, K.; Clements, A.; Joly, D.O.; Wolfe, N.D.; Daszak, P.; Karesh, W.B.; et al. Spillover and pandemic properties of zoonotic viruses with high host plasticity. Sci. Rep. 2015, 5, 14830. [Google Scholar] [CrossRef] [Green Version]
- Cuadros, F.D.; Xiao, Y.; Mukandavire, Z.; Correa-Agudelo, E.; Hernández, A.; Kim, H.; MacKinnon, N.J. Spatiotemporal transmission dynamics of the COVID-19 pandemic and its impact on critical healthcare capacity. Health Place 2020, 64, 102404. [Google Scholar] [CrossRef]
- Li, C.; Ji, F.; Wang, L.; Wang, L.; Hao, J.; Dai, M.; Liu, Y.; Pan, X.; Fu, J.; Li, L.; et al. Asymptomatic and Human-to-Human Transmission of SARS-CoV-2 in a 2-Family Cluster, Xuzhou, China. Emerg. Infect. Dis. 2020, 26, 1626–1628. [Google Scholar] [CrossRef]
- Burrell, C.J.; Howard, C.R.; Murphy, F.A. Epidemiology of Viral Infections. Fenner White’s Med. Virol. 2017, 5, 185–203. [Google Scholar] [CrossRef]
- Lytras, S.; Xia, W.; Hughes, J.; Jiang, X.; Robertson, D.L. The animal origin of SARS-CoV-2. Science 2021, 373, 968–970. [Google Scholar] [CrossRef]
- Coronaviridae. Chapter Version: ICTV Ninth Report; 2009 Taxonomy Release. Available online: https://ictv.global/report_9th/RNApos/Nidovirales/Coronaviridae (accessed on 1 August 2022).
- Leopardi, S.; Oluwayelu, D.; Meseko, C.; Marciano, S.; Tassoni, L.; Bakarey, S.; Monne, I.; Cattoli, G.; De Benedictis, P. The close genetic relationship of lineage D Betacoronavirus from Nigerian and Kenyan straw-colored fruit bats (Eidolon helvum) is consistent with the existence of a single epidemiological unit across sub-Saharan Africa. Virus Genes 2016, 52, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.-Y.; Wang, N.; Zhang, W.; Hu, B.; Li, B.; Zhang, Y.-Z.; Zhou, J.-H.; Luo, C.-M.; Yang, X.-L.; Wu, L.-J.; et al. Coexistence of multiple coronaviruses in several bat colonies in an abandoned mineshaft. Virol. Sin. 2016, 31, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.S. The high diversity of SARS-CoV-2-related coronaviruses in pangolins alerts potential ecological risks. Zool. Res. 2021, 42, 834–844. [Google Scholar] [CrossRef]
- Temmam, S.; Vongphayloth, K.; Baquero, E.; Munier, S.; Bonomi, M.; Regnault, B.; Douangboubpha, B.; Karami, Y.; Chrétien, D.; Sanamxay, D.; et al. Bat coronaviruses related to SARS-CoV-2 and infectious for human cells. Nature 2022, 604, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Allam, Z. The first 50 days of COVID-19: A detailed chronological timeline and extensive review of literature documenting the pandemic. In Surveying the Covid-19 Pandemic and its Implications. Urban Health, Data Technology and Political Economy; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–7. [Google Scholar] [CrossRef]
- McCarthy, S. Coronavirus: Thousands of Wild Animals Were Sold at Wuhan Markets in Months before COVID-19 Outbreak, New Report Finds. South China Morning Post, 2021. Available online: https://www.scmp.com/news/china/science/article/3136674/thousands-wild-animals-were-sold-wuhan-markets-months-covid-19(accessed on 23 November 2021).
- Center for Genomic Regulation. Ferrets, cats and civets most susceptible to coronavirus infection after humans: Ducks, rats, mice, pigs and chickens had lower or no susceptibility to infection. Science Daily, 2021. Available online: www.sciencedaily.com/releases/2020/12/201210112147.htm(accessed on 11 September 2021).
- Porter, S.M.; Hartwig, A.E.; Bielefeldt-Ohmann, H.; Bosco-Lauth, A.M.; Root, J.J. Susceptibility of wild canids to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). BioRxiv 2022. preprint. [Google Scholar] [CrossRef]
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Munnink, B.B.O.; Der Honing, R.W.H.-V.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.S.; et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Euro Surveill. 2020, 25, 2001005. [Google Scholar] [CrossRef] [PubMed]
- Freuling, C.M.; Breithaupt, A.; Müller, T.; Sehl, J.; Balkema-Buschmann, A.; Rissmann, M.; Klein, A.; Wylezich, C.; Höper, D.; Wernike, K.; et al. Susceptibility of Raccoon Dogs for Experimental SARS-CoV-2 Infection. Emerg. Infect. Dis. 2020, 26, 2982–2985. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Liu, W.; Liu, P.; Lei, W.; Jia, Z.; He, X.; Liu, L.; Shi, W.; Tan, Y.; Zou, S.; et al. Surveillance of SARS-CoV-2 in the environment and animal samples of the Huanan Seafood Market. Res. Sq. 2022. Available online: https://assets.researchsquare.com/files/rs-1370392/v1_covered.pdf?c=1645813311 (accessed on 17 March 2021).
- Allen, T.; Murray, K.A.; Zambrana-Torrelio, C.; Morse, S.S.; Rondinini, C.; Di Marco, M.; Breit, N.; Olival, K.J.; Daszak, P. Global hotspots and correlates of emerging zoonotic diseases. Nat. Commun. 2017, 8, 1124. [Google Scholar] [CrossRef] [Green Version]
- Houe, H.; Nielsen, S.S.; Nielsen, L.R.; Ethelberg, S.; Mølbak, K. Opportunities for improved disease surveillance and control by use of integrated data on animal and human health. Front. Vet. Sci. 2019, 6, 301. [Google Scholar] [CrossRef]
- Grange, S.L.; Goldstein, T.; Johnson, C.K.; Anthony, S.; Gilardi, K.; Daszak, P.; Olival, K.J.; O’Rourke, T.; Murray, S.; Olson, S.H.; et al. Ranking the risk of animal-to-human spillover for newly discovered viruses. Proc. Natl. Acad. Sci. USA 2021, 118, e2002324118. [Google Scholar] [CrossRef]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Li, X.; Luk, H.K.; Lau, S.P.; Woo, P.C. Human Coronaviruses: General Features. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Amer, H.M. Bovine-like coronaviruses in domestic and wild ruminants. Anim. Health Res. Rev. 2018, 19, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Mousavizadeh, L.; Ghasemi, S. Genotype and phenotype of COVID-19: Their roles in pathogenesis. J. Microbiol. Immunol. Infect. 2021, 54, 159–163. [Google Scholar] [CrossRef]
- Ji, W.; Wang, W.; Zhao, X.; Zai, J.; Li, X. Cross-species transmission of the newly identified coronavirus 2019-nCoV. J. Med. Virol. 2020, 92, 433–440. [Google Scholar] [CrossRef]
- Lau, S.K.P.; Luk, H.K.H.; Wong, A.C.P.; Li, K.S.M.; Zhu, L.; He, Z.; Fung, J.; Chan, T.T.Y.; Fung, K.S.C.; Woo, P.C.Y. Possible Bat Origin of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1542–1547. [Google Scholar] [CrossRef]
- Pfefferle, S.; Oppong, S.; Drexler, J.F.; Gloza-Rausch, F.; Ipsen, A.; Seebens, A.; Müller, M.A.; Annan, A.; Vallo, P.; Adu-Sarkodie, Y.; et al. Distant relatives of severe acute respiratory syndrome coronavirus and close relatives of human coronavirus 229E in bats, Ghana. Emerg. Infect. Dis. 2009, 15, 1377–1384. [Google Scholar] [CrossRef]
- Quan, P.-L.; Firth, C.; Street, C.; Henriquez, J.A.; Petrosov, A.; Tashmukhamedova, A.; Hutchison, S.K.; Egholm, M.; Osinubi, M.O.V.; Niezgoda, M.; et al. Identification of a severe acute respiratory syndrome coronavirus-like virus in a leaf-nosed bat in Nigeria. mBio 2010, 1, e00208-10. [Google Scholar] [CrossRef] [Green Version]
- Parrish, C.R.; Holmes, E.C.; Morens, D.M.; Park, E.-C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-species virus transmission and the emergence of new epidemic diseases. Microbiol. Mol. Biol. Rev. 2008, 72, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Cleaveland, S.; Laurenson, M.K.; Taylor, L.H. Diseases of humans and their domestic mammals: Pathogen characteristics, host range and the risk of emergence. Philos. Trans. R. Soc. B 2001, 356, 991–999. [Google Scholar] [CrossRef]
- Woolhouse, M.E. Population biology of emerging and re-emerging pathogens. Trends Microbiol. 2002, 10, S3–S7. [Google Scholar] [CrossRef]
- Rendon-Marin, S.; Martinez-Gutierrez, M.; Whittaker, G.R.; James, J.A.; Ruiz-Saenz, J. SARS CoV-2 spike protein in silico interaction with ACE2 receptors from wild and domestic species. Front. Genet. 2021, 12, 571707. [Google Scholar] [CrossRef]
- Van Aart, A.E.; Velkers, F.C.; Fischer, E.A.; Broens, E.M.; Egberink, H.; Zhao, S.; Engelsma, M.; Hakze-van der Honing, R.W.; Harders, F.; de Rooij, M.M.; et al. SARS-CoV-2 infection in cats and dogs in infected mink farms. Transbound Emerg. Dis. 2021, 69, 3001–3007. [Google Scholar] [CrossRef]
- Gusev, E.; Sarapultsev, A.; Solomatina, L.; Chereshnev, V. SARS-CoV-2-Specific Immune Response and the Pathogenesis of COVID-19. Int. J. Mol. Sci. 2022, 23, 1716. [Google Scholar] [CrossRef]
- Nelson, D.D.; Duprau, J.L.; Wolff, P.L.; Evermann, J.F. Persistent Bovine Viral Diarrhea Virus Infection in Domestic and Wild Small Ruminants and Camelids Including the Mountain Goat (Oreamnos americanus). Front. Microbiol. 2016, 6, 1415. [Google Scholar] [CrossRef]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.-E.; Kavanagh Williamson, M.; Antón-Plágaro, C.; Shoemark, D.K.; Simón-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; Van Der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Wang, K.; Chen, W.; Zhang, Z.; Deng, Y.; Lian, J.-Q.; Du, P.; Wei, D.; Zhang, Y.; Sun, X.-X.; Gong, L.; et al. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct. Target. Ther. 2020, 5, 283. [Google Scholar] [CrossRef] [PubMed]
- Strumillo, S.T.; Kartavykh, D.; de Carvalho, F.F., Jr.; Cruz, N.C.; de Souza Teodoro, A.C.; Sobhie, D.R.; Curcio, M.F. Host-virus interaction and viral evasion. Cell Biol. Int. 2021, 45, 1124–1147. [Google Scholar] [CrossRef] [PubMed]
- Račnik, J.; Kočevar, A.; Slavec, B.; Korva, M.; Rus, K.R.; Zakotnik, S.; Zorec, T.M.; Poljak, M.; Matko, M.; Rojs, O.Z.; et al. Transmission of SARS-CoV-2 from Human to Domestic Ferret. Emerg. Infect. Dis. 2021, 27, 2450–2453. [Google Scholar] [CrossRef]
- Salguero, F.J.; White, A.D.; Slack, G.S.; Fotheringham, S.A.; Bewley, K.R.; Gooch, K.E.; Longet, S.; Humphries, H.E.; Watson, R.J.; Hunter, L.; et al. Comparison of rhesus and cynomolgus macaques as an infection model for COVID-19. Nat. Commun. 2021, 12, 1260. [Google Scholar] [CrossRef] [PubMed]
- Hale, V.L.; Dennis, P.M.; McBride, D.S.; Nolting, J.M.; Madden, C.; Huey, D.; Ehrlich, M.; Grieser, J.; Winston, J.; Lombardi, D.; et al. SARS-CoV-2 infection in free-ranging white-tailed deer. Nature 2022, 602, 481–486. [Google Scholar] [CrossRef]
- Mykytyn, A.Z.; Lamers, M.M.; Okba, N.M.A.; Breugem, T.I.; Schipper, D.; Doel, P.B.V.D.; van Run, P.; van Amerongen, G.; de Waal, L.; Koopmans, M.P.G.; et al. Susceptibility of rabbits to SARS-CoV-2. Emerg. Microbes Infect. 2021, 10, 1–7. [Google Scholar] [CrossRef]
- Francis, M.E.; Goncin, U.; Kroeker, A.; Swan, C.; Ralph, R.; Lu, Y.; Etzioni, A.L.; Falzarano, D.; Gerdts, V.; Machtaler, S.; et al. SARS-CoV-2 infection in the Syrian hamster model causes inflammation as well as type I interferon dysregulation in both respiratory and non-respiratory tissues including the heart and kidney. PLoS Pathog. 2021, 17, e1009705. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Sia, S.F.; Yan, L.-M.; Chin, A.W.H.; Fung, K.; Choy, K.-T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.A.P.M.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef]
- De Morais, H.A.; dos Santos, A.P.; do Nascimento, N.C.; Kmetiuk, L.B.; Barbosa, D.S.; Brandão, P.E.; Guimarães, A.M.S.; Pettan-Brewer, C.; Biondo, A.W. Natural Infection by SARS-CoV-2 in Companion Animals: A Review of Case Reports and Current Evidence of Their Role in the Epidemiology of COVID-19. Front. Vet. Sci. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Koeppel, K.N.; Mendes, A.; Strydom, A.; Rotherham, L.; Mulumba, M.; Venter, M. SARS-CoV-2 Reverse Zoonoses to Pumas and Lions, South Africa. Viruses 2022, 14, 120. [Google Scholar] [CrossRef]
- Palermo, P.M. SARS-CoV-2 neutralizing antibodies in white-tailed deer from Texas. Vector Borne Zoonotic Dis. 2022, 22, 62–64. [Google Scholar] [CrossRef]
- Yen, H.L.; Sit, T.H.; Brackman, C.J.; Chuk, S.S.; Gu, H.; Tam, K.W.; Law, P.Y.; Leung, G.M.; Peiris, M.; Poon, L.L.; et al. Transmission of SARS-CoV-2 delta variant (AY.127) from pet hamsters to humans, leading to onward human-to-human transmission: A case study. Lancet 2022, 399, 1070–1078. [Google Scholar] [CrossRef]
- Pickering, B.; Lung, O.; Maguire, F.; Kruczkiewicz, P.; Kotwa, J.D.; Buchanan, T.; Gagnier, M.; Guthrie, J.L.; Jardine, C.M.; Marchand-Austin, A.; et al. Highly divergent white-tailed deer SARS-CoV-2 with potential deer-to-human transmission. BioRxiv 2022. preprint. [Google Scholar] [CrossRef]
- Tan, C.C.S.; Lam, S.D.; Richard, D.; Owen, C.J.; Berchtold, D.; Orengo, C.; Nair, M.S.; Kuchipudi, S.V.; Kapur, V.; van Dorp, L.; et al. Transmission of SARS-CoV-2 from humans to animals and potential host adaptation. Nat. Commun. 2022, 13, 2988. [Google Scholar] [CrossRef]
- Ellwanger, J.H.; Chies, J.A.B. Zoonotic spillover: Understanding basic aspects for better prevention. Genet. Mol. Biol. 2021, 44 (Suppl. S1). [Google Scholar] [CrossRef]
- Wang, N.; Li, S.-Y.; Yang, X.-L.; Huang, H.-M.; Zhang, Y.-J.; Guo, H.; Luo, C.-M.; Miller, M.; Zhu, G.; Chmura, A.A.; et al. Serological evidence of bat SARS-related coronavirus infection in humans, China. Virol. Sin. 2018, 33, 104–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Mendelsohn, E.; Zong, C.; Zhang, W.; Hagan, E.; Wang, N.; Li, S.; Yan, H.; Huang, H.; Zhu, G.; et al. Human-animal interactions and bat coronavirus spill-over potential among rural residents in Southern China. Biosaf. Health 2019, 1, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Hassell, J.M. Urbanization and Disease Emergence: Dynamics at the Wildlife–Livestock–Human Interface. Trends Ecol. Evol. 2017, 32, 55–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.L.; Leach, M.; Waldman, L.; MacGregor, H.; Fooks, A.R.; Jones, K.E. A framework for the study of zoonotic disease emergence and its drivers: Spill-over of bat pathogens as a case study. Phil. Trans. R. Soc. B 2012, 367, 2881–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Health Organization (WHO). Ebola Virus Disease. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 4 December 2021).
- World Health Organization (WHO). Marburg Virus Disease. 2021. Available online: https://www.who.int/health-topics/marburg-virus-disease (accessed on 4 December 2021).
- World Health Organization (WHO). Hendra Virus Disease. 2021. Available online: https://www.who.int/health-topics/hendra-virus-disease (accessed on 4 December 2021).
- World Health Organization (WHO). Nipah Virus. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/nipah-virus (accessed on 4 December 2021).
- Shope, R.E. Rabies-related viruses. Yale J. Biol. Med. 1982, 55, 271–275. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Arenaviruses. 2021. Available online: https://www.cdc.gov/vhf/virus-families/arenaviridae.html (accessed on 17 March 2022).
- Kurpiers, L.A.; Schulte-Herbrüggen, B.; Ejotre, I.; Reeder, D.M. Bushmeat and Emerging Infectious Diseases: Lessons from Africa. Probl. Wildl. 2015, 21, 507–551. [Google Scholar] [CrossRef]
- Saéz, A.M.; Weiss, S.; Nowak, K.; Lapeyre, V.; Zimmermann, F.; Düx, A.; Kühl, H.S.; Kaba, M.; Regnaut, S.; Merkel, K.; et al. Investigating the zoonotic origin of the West African Ebola epidemic. EMBO Mol. Med. 2015, 7, 17–23. [Google Scholar] [CrossRef]
- Guan, Y.J.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Butt, K.M.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, C.A.; Li, H.; Phelps, K.L.; Zambrana-Torrelio, C.; Wang, L.-F.; Zhou, P.; Shi, Z.-L.; Olival, K.J.; Daszak, P. A strategy to assess spillover risk of bat SARS-related coronaviruses in Southeast Asia. Nat. Commun. 2022, 13, 4380. [Google Scholar] [CrossRef]
- World Health Organization. Transmission of SARS-CoV-2: Implications for Infection Prevention Precautions. Sci. Brief. 2020. Available online: https://www.who.int/news-room/commentaries/detail/transmission-of-sars-cov-2-implications-for-infection-prevention-precautions (accessed on 11 July 2022).
- Bonwitt, J.; Saez, A.M.; Lamin, J.; Ansumana, R.; Dawson, M.; Buanie, J.; Lamin, J.; Sondufu, D.; Borchert, M.; Sahr, F.; et al. At Home with Mastomys and Rattus: Human-Rodent Interactions and Potential for Primary Transmission of Lassa Virus in Domestic Spaces. Am. J. Trop. Med. Hyg. 2017, 96, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Gurley, E.S.; Hegde, S.T.; Hossain, K.; Sazzad, H.M.S.; Hossain, M.J.; Rahman, M.; Sharker, M.A.Y.; Salje, H.; Islam, M.S.; Epstein, J.H.; et al. Convergence of Humans, Bats, Trees, and Culture in Nipah Virus Transmission, Bangladesh. Emerg. Infect. Dis. 2017, 23, 1446–1453. [Google Scholar] [CrossRef] [Green Version]
- Altizer, S.; Becker, D.J.; Epstein, J.H.; Forbes, K.M.; Gillespie, T.R.; Hall, R.J.; Hawley, D.M.; Hernandez, S.M.; Martin, L.B.; Plowright, R.K.; et al. Food for contagion: Synthesis and future directions for studying host-parasite responses to resource shifts in anthropogenic environments. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2018, 373, 20170102. [Google Scholar] [CrossRef]
- Shervani, Z.; Khan, I.; Siddiqui, N.Y.; Khan, T.; Qazi, U.Y. Risk of SARS-CoV-2 transmission from humans to pets and vice versa. Eur. J. Med. Health Sci. 2021, 1, 34–38. [Google Scholar] [CrossRef]
- Cadar, D.; Allendorf, V.; Schulze, V.; Ulrich, R.G.; Schlottau, K.; Ebinger, A.; Hoffmann, B.; Hoffmann, D.; Rubbenstroth, D.; Ismer, G.; et al. Introduction and spread of variegated squirrel bornavirus 1 (VSBV-1) between exotic squirrels and spill-over infections to humans in Germany. Emerg. Microbes Infect. 2021, 10, 602–611. [Google Scholar] [CrossRef]
- Reynolds, M.G.; Davidson, W.B.; Curns, A.T.; Conover, C.S.; Huhn, G.; Davis, J.P.; Wegner, M.; Croft, D.R.; Newman, A.; Obiesie, N.N.; et al. Spectrum of infection and risk factors for human monkeypox, United States, 2003. Emerg. Infect. Dis. 2007, 13, 1332–1339. [Google Scholar] [CrossRef]
- Kampf, G.; Lemmen, S.; Suchomel, M. Ct values and infectivity of SARS-CoV-2 on surfaces. Lancet Infect. Dis. 2021, 21, e141. [Google Scholar] [CrossRef]
- Fadare, O.O.; Okoffo, E.D. COVID-19 face masks: A potential source of microplastic fibers in the environment. Sci. Total Environ. 2020, 737, 140279. [Google Scholar] [CrossRef]
- Magwedere, K.; Bishi, A.; Tjipura-Zaire, G.; Eberle, G.; Hemberger, Y.; Hoffman, L.C.; Dziva, F. Brucellae through the food chain: The role of sheep, goats and springbok (Antidorcus marsupialis) as sources of human infections in Namibia. J. S. Afr. Vet. Assoc. 2011, 82, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Buchholz, U.; Bernard, H.; Werber, D.; Böhmer, M.M.; Remschmidt, C.; Wilking, H.; Deleré, Y.; der Heiden, M.A.; Adlhoch, C.; Dreesman, J.; et al. German outbreak of Escherichia coli O104:H4 associated with sprouts. N. Engl. J. Med. 2011, 365, 1763–1770. [Google Scholar] [CrossRef] [PubMed]
- Sophia, G.; de Vries, B.J.; Visser, I.M.; Nagel, M.G.A.; Goris, R.A.; Hartskeerl, M.P.G. Leptospirosis in Sub-Saharan Africa: A systematic review. Inter. J. Infect. Dis. 2014, 28, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Berthe, F.C.J.; Bouley, T.; Karesh, W.B.; Le Gall, F.G.; Machalaba, C.C.; Plante, C.A.; Seifman, R.M. Operational Framework for Strengthening Human, Animal and Environmental Public Health Systems at Their Interface (English); World Bank Group: Washington, DC, USA, 2018; Available online: http://documents.worldbank.org/curated/en/703711517234402168/Operational-framework-for-strengthening-human-animal-and-environmental-public-health-systems-at-their-interface (accessed on 7 July 2022).
- World Health Organization (WHO). In Africa, 63% Jump in Diseases Spread from Animals to People Seen in Last Decade. 2022. Available online: https://www.afro.who.int/news/africa-63-jump-diseases-spread-animals-people-seen-last-decade (accessed on 12 August 2022).
- Mableson, H.E.; Okello, A.; Picozzi, K.; Welburn, S.C. Neglected zoonotic diseases-the long and winding road to advocacy. PLoS Negl. Trop. Dis. 2014, 8, e2800. [Google Scholar] [CrossRef] [PubMed]
- Grace, D.; Songe, M.; Knight-Jones, T. Impact of Neglected Diseases on Animal Productivity and Public Health in Africa. Africa—OIE Regional Commission. 2015. Available online: https://www.oie.int/app/uploads/2021/03/2015-afr1-grace-a.pdf (accessed on 17 August 2022).
- Elelu, N.; Aiyedun, J.O.; Mohammed, I.G.; Oludairo, O.; Odetokun, I.A.; Mohammed, K.M.; Bale, J.O.; Nuru, S. Neglected zoonotic diseases in Nigeria: Role of the public health veterinarian. Pan Afr. Med. J. 2019, 32, 36. [Google Scholar] [CrossRef] [PubMed]
- Rist, C.L.; Arriola, C.S.; Rubin, C. Prioritizing zoonoses: A proposed One Health tool for collaborative decision-making. PLoS ONE 2014, 9, e109986. [Google Scholar] [CrossRef]
- Grace, D.; Mutua, F.; Ochungo, P.; Kruska, R.; Jones, K.; Brierley, L.; Lapar, L.; Said, M.; Herrero, M.; Phuc, P.M.; et al. Mapping of Poverty and Likely Zoonoses Hotspots; Zoonoses Project 4, Report to the UK Department for International Development; ILRI: Nairobi, Kenya, 2012. [Google Scholar]
- World Health Organization. New WHO Estimates: Up to 190,000 People Could Die of COVID-19 in Africa If Not Controlled. 2020. Available online: https://www.afro.who.int/news/new-who-estimates-190-000-people-could-die-covid-19-africa-if-not-controlled (accessed on 12 November 2021).
- Cabore, J.W.; Karamagi, H.C.; Kipruto, H.K.; Mungatu, J.K.; Asamani, J.A.; Droti, B.; Titi-Ofei, R.; Seydi, A.B.W.; Kidane, S.N.; Balde, T.; et al. COVID-19 in the 47 countries of the WHO African region: A modelling analysis of past trends and future patterns. Lancet Glob. Health 2022, 10, e1099–e1114. [Google Scholar] [CrossRef]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. USA 2013, 110, 8399–8404. [Google Scholar] [CrossRef] [Green Version]
- Kapata, N.; Ihekweazu, C.; Ntoumi, F.; Raji, T.; Chanda-Kapata, P.; Mwaba, P.; Mukonka, V.; Bates, M.; Tembo, J.; Corman, V.; et al. Is Africa prepared for tackling the COVID-19 (SARS-CoV-2) epidemic. Lessons from past outbreaks, ongoing pan-African public health efforts, and implications for the future. Int. J. Infect. Dis. 2020, 93, 233–236. [Google Scholar] [CrossRef]
- Agusi, E.R.; Ijoma, S.I.; Nnochin, C.S.; Njoku-Achu, N.O.; Nwosuh, C.I.; Meseko, C.A. The COVID-19 pandemic and social distancing in Nigeria: Ignorance or defiance. Pan Afr. Med. 2020, 35 (Suppl. S2), 52. [Google Scholar] [CrossRef]
- Torti, C.; Mazzitelli, M.; Trecarichi, E.M.; Darius, O. Potential implications of SARS-CoV-2 epidemic in Africa: Where are we going from now? BMC Infect. Dis. 2022, 20, 412. [Google Scholar] [CrossRef]
- Nordling, L. Africa’s pandemic puzzle: Why so few cases and deaths? Science 2020, 369, 756–757. [Google Scholar] [CrossRef]
- Soko, N.D.; Dlamini, S.; Ntsekhe, M.; Dandara, C. The COVID-19 Pandemic and Explaining Outcomes in Africa: Could Genomic Variation Add to the Debate? OMICS J. Integr. Biol. 2022. [Google Scholar] [CrossRef]
- Altable, M.; de la Serna, J.M. Protection against COVID-19 in African population: Immunology, genetics, and malaria clues for therapeutic targets. Virus Res. 2021, 299, 198347. [Google Scholar] [CrossRef]
- The World Bank. Nigeria’s Economy Faces Worst Recession in Four Decades, Says New World Bank Report. 2020. Available online: https://www.worldbank.org/en/news/press-release/2020/06/25/nigerias-economy-faces-worst-recession-in-four-decades-says-new-world-bank-report (accessed on 23 May 2022).
- World Health Organization. Achieving 70% COVID-19 Immunization Coverage by Mid-2022. 2021. Available online: https://www.who.int/news/item/23-12-2021-achieving-70-covid-19-immunization-coverage-by-mid-2022 (accessed on 19 November 2021).
- Food and Agriculture Organization of the United Nations. The State of Food Security and Nutrition in the World; FAO: Rome, Italy, 2021. [Google Scholar]
- Orjuela-Grimm, M.; Deschak, C.; Gama, C.A.A.; Carreño, S.B.; Hoyos, L.; Mundo, V.; Bojorquez, I.; Carpio, K.; Quero, Y.; Xicotencatl, A.; et al. Migrants on the Move and Food (In)security: A Call for Research. J. Immigr Minor. Health 2022, 24, 1318–1327. [Google Scholar] [CrossRef]
- Holder, J. Tracking Coronavirus Vaccinations Around the World. New York Times Newspaper, 2022. Available online: https://www.nytimes.com/interactive/2021/world/covid-vaccinations-tracker.html(accessed on 5 September 2022).
- World Health Organization. Less than 10% of African Countries to Hit Key COVID-19 Vaccination Goal. 2021. Available online: https://www.afro.who.int/news/less-10-african-countries-hit-key-covid-19-vaccination-goal (accessed on 19 November 2021).
- Mutombo, P.N.; Fallah, M.P.; Munodawafa, D.; Kabel, A.; Houeto, D.; Goronga, T.; Mweemba, O.; Balance, G.; Onya, H.; Kamba, R.S.; et al. COVID-19 vaccine hesitancy in Africa: A call to action. Lancet Glob. Health 2022, 10, e320–e321. [Google Scholar] [CrossRef]
- Nature Editorial. Africa is bringing vaccine manufacturing home. Nature 2022, 602, 184. [Google Scholar] [CrossRef]
- Chan, O.S.K.; Bradley, K.C.F.; Grioni, A.; Lau, S.K.P.; Li, W.-T.; Magouras, I.; Naing, T.; Padula, A.; To, E.M.W.; Tun, H.M.; et al. Veterinary Experiences can Inform One Health Strategies for Animal Coronaviruses. Ecohealth 2021, 18, 301–314. [Google Scholar] [CrossRef]
- Ducrot, C.; Bed’Hom, B.; Béringue, V.; Coulon, J.-B.; Fourichon, C.; Guérin, J.-L.; Krebs, S.; Rainard, P.; Schwartz-Cornil, I.; Torny, D.; et al. Issues and special features of animal health research. Vet. Res. 2011, 42, 96. [Google Scholar] [CrossRef] [Green Version]
- The Food and Agriculture Organization of the United Nations. The Impact of Disasters and Crises on Agriculture and Food Security; FAO: Rome, Italy, 2021. [Google Scholar] [CrossRef]
- He, S.; Han, J.; Lichtfouse, E. Backward transmission of COVID-19 from humans to animals may propagate reinfections and induce vaccine failure. Environ. Chem. Lett. 2021, 19, 763–768. [Google Scholar] [CrossRef]
- Ajayi, E.A.; Olatumile, A. Indigenous folklores as a tool of transformative learning for environmental sustainability in Nigeria. Andragoške Studije 2018, 11, 29–44. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Measure | Explanation |
|---|---|
| Enactments | As a precaution, the public is urged to keep safe distances from wildlife, particularly species that are known to be susceptible to SARS-CoV-2 infections. This can be achieved through enactments such as emergency orders and temporary bans on hunting, trading, and non-essential contact with wildlife. Relatedly, prohibitive measures may be taken to discourage bushmeat hunting and the consumption of raw, unprocessed meat. In all of these, alternative sources of low-cost nutrition should be provided in resource limited communities in order to spare wildlife hunting. |
| Education | To highlight issues of public health concern, awareness must be raised among the population regarding the possibility of disease transmission from animals to humans and vice versa. While some cultural or religious traditions may predispose to direct contact with wildlife, there is a need to effectively communicate the risks of disease transmission. It may be helpful to recall traditional folk tales that highlight the inherent danger in handling (deceased) wildlife as a learning tool in local public teaching. Many such folklores abound among the native Yorubas of Nigeria and may be applicable in other tribal settings in Africa [123]. Furthermore, early education of pupils on the dangers of emerging or neglected zoonotic diseases and the precautionary measures to be taken can be a significant way to bridge the knowledge gap. |
| National One Health Strategic Plan | A national one health strategic plan should be actively adopted across the different organizational frontiers (in Africa) to safeguard a holistic impact. From joint (national) research projects to research projects by independent donors, combined efforts must achieve the objective of containing virus spread. The capabilities of self-determined and independent, top-level research should be continuously increased via access to funding, training, and education. This enables local and independent monitoring and rapid identification of novel emerging pathogens. |
| Vaccine equity | The autonomous development and production of vaccines in Africa must be actively supported. This will create the enabling conditions to achieve a rapid response to emerging or pandemic pathogens and increase the global vaccination coverage while continually developing vaccines for mutant strains of the virus [117]. |
| Heightened Monitoring | Gathering data on human practices as well as contact with animals in settings with diverse host assemblages will ensure effective analysis regarding potential spillover risks. Environmental, veterinary and medical scientists may further investigate the pathogens present and exchanged at the identified human–animal interfaces of risk. This will give an informed direction towards critical points for disease control and behavior change interventions aimed at prevention. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agusi, E.R.; Allendorf, V.; Eze, E.A.; Asala, O.; Shittu, I.; Dietze, K.; Busch, F.; Globig, A.; Meseko, C.A. SARS-CoV-2 at the Human–Animal Interface: Implication for Global Public Health from an African Perspective. Viruses 2022, 14, 2473. https://doi.org/10.3390/v14112473
Agusi ER, Allendorf V, Eze EA, Asala O, Shittu I, Dietze K, Busch F, Globig A, Meseko CA. SARS-CoV-2 at the Human–Animal Interface: Implication for Global Public Health from an African Perspective. Viruses. 2022; 14(11):2473. https://doi.org/10.3390/v14112473
Chicago/Turabian StyleAgusi, Ebere Roseann, Valerie Allendorf, Emmanuel Aniebonam Eze, Olayinka Asala, Ismaila Shittu, Klaas Dietze, Frank Busch, Anja Globig, and Clement Adebajo Meseko. 2022. "SARS-CoV-2 at the Human–Animal Interface: Implication for Global Public Health from an African Perspective" Viruses 14, no. 11: 2473. https://doi.org/10.3390/v14112473