
Full Genome Sequencing of Three Sedoreoviridae Viruses Isolated from Culicoides spp. (Diptera, Ceratopogonidae) in China

1
Yunnan Tropical and Subtropical Animal Virus Diseases Laboratory, Yunnan Animal Science and Veterinary Institute, Kunming 650224, China
2
Research Institute for the Environment and Livelihoods, Charles Darwin University, Darwin, NT 0909, Australia
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Viruses 2022, 14(5), 971; https://doi.org/10.3390/v14050971
Submission received: 13 April 2022
/
Revised: 2 May 2022
/
Accepted: 3 May 2022
/
Published: 5 May 2022
(This article belongs to the Special Issue Arbovirus Discovery)
Abstract
:Sedoreoviridae is a family of viruses belonging to the order Reovirales and comprises six genera, two of which, Orbivirus and Seadornavirus, contain arboviruses that cause disease in humans and livestock. Areas such as Yunnan Province in southwestern China, have high arboviral activity due in part to warm and wet summers, which support high populations of biting flies such as mosquitoes and Culicoides. Three viral isolates previously obtained from Culicoides collected at cattle farms in Shizong County of Yunnan Province, China, between 2019 and 2020 were completely sequenced and identified as Banna virus (BAV) genotype A of Seadornavirus and serotypes 1 and 7 of epizootic hemorrhagic disease virus (EHDV) of Orbivirus. These results suggest that Culicoides tainanus and C. orientalis are potential vectors of BAV and EHDV, respectively, and represent the first association of a BAV with C. tainanus and of an arbovirus with C. orientalis. Analysis using VP9 generally agreed with the current groupings within this genus based on VP12, although the classification for some strains should be corrected. Furthermore, the placement of Kadipiro virus (KDV) and Liao ning virus (LNV) in Seadornavirus may need confirmation as phylogenetic analysis placed these viruses as sister to other species in the genus.
1. Introduction
Viruses belonging to the Reovirales are characterized by possessing multiple genomic segments of dsRNA and a double layer of capsids [1]. Two families, Sedoreoviridae and Spinareoviridae, are recognized, with Sedoreoviridae composed of six genera including two, Orbivirus and Seadornavirus, which contain arboviruses that cause disease in humans or livestock [2]. The double layers of capsids help these arboviruses to infect diplex hosts, such as mammals and insects [1]. Bluetongue viruses (BTV, the prototype of genus Orbivirus), for example, possess an outer capsid protein VP2 and inner capsid protein VP7, which act as ligands binding to the cellular receptors of susceptible ruminant and insect cells, respectively [3,4]. BTV attaches to ruminant host cells via VP2 after they enter the blood through the bite of an infected Culicoides vector [3,5]. Following ingestion by the insect vector, virus in the blood meal infects Culicoides host cells via VP7 after the outer capsid is digested in the midgut [4,6,7]. Almost all species of Orbivirus and Seadornavirus are arboviruses [1,5,8,9,10].
Banna virus (BAV), the prototype species of Seadornavirus [11], was first isolated from sera and cerebrospinal fluid collected from human patients with encephalitis and fever in Xishuang-Banna of Yunnan Province, China in 1987 [12]. Subsequently, BAV strains have been isolated from cattle and swine [13], and have been reported in Indonesia and Vietnam in addition to China [8,13,14]. Thus far, BAV strains have been isolated from 10 mosquito species belonging to 3 genera (Aedes, Anopheles, and Culex) and unsorted Culicoides [13,15,16]. It is uncertain whether BAV can infect ticks, although isolation of BAV from ticks was reported in China in 1992 [17]. Phylogenetic analysis of BAV isolates recognizes four genotypes: A1, A2, B, and C [16,18].
Epizootic hemorrhagic disease virus (EHDV) belongs to the genus Orbivirus, and was first recognized from a highly fatal disease of wild white-tailed deer in north America in 1955 [19]. Closely related to BTV, EHDV mainly infects ruminants, especially deer, cattle, sheep and goats [5,20,21,22], and is transmitted by Culicoides [23]. EHDV has been reported from Africa, Asia, North America, South America, and Oceania [24]. Thus far, at least seven serotypes (EHDV-1, 2, 4, 5, 6, 7, and 8) are recognized by the World Organization for Animal Health (OIE) [25], while a further two serotypes, EHDV-10 [26,27] and an novel serotype isolated in China [28] reported recently, are yet to be formally recognized. Although EHDV infects bovine, sheep and goats, these hosts are usually asymptomatic or subsymptomatic. Some strains of EHDV-2, EHDV-6 and EHDV-7 have, however, caused several outbreaks of epizootic hemorrhagic disease in deer or cattle in North America and Asia [19,22,29,30,31].
Culicoides (Diptera: Ceratopogonidae) is a genus of small biting midges with more than 1340 described species [32]. At least 40 of these are associated with the transmission of approximately 50 animal-associated arboviruses belonging to the families Peribunyaviridae (previously known as Bunyaviridae), Sedoreoviridae, and Rhabdoviridae [23,33].
Yunnan Province in the southwest of China encompasses a broad range of climatic zones ranging from tropical areas bordering Myanmar, Laos and Vietnam in the south, to alpine climates bordering Tibet in the north. Warm and rainy summers throughout most of the province are conducive to populations of biting flies such as mosquitoes and Culicoides. These factors identify Yunnan as one of the most active provinces in China for insect-borne diseases, particularly if the threat of transboundary diseases from neighboring countries is considered. This high level of arboviral activity is evidenced by the initial discovery in Yunnan of BAV and Tibet Orbivirus (TIBOV) [12,34,35], the presence of a large number of strains of BTV and EHDV [28,36,37,38], and the high prevalence of dengue virus in southern Yunnan [39,40,41].
Shizong County of Yunnan Province was the site of the first domestic case of bluetongue disease in China, prompting a number of longitudinal studies into the ecology of this virus in the area [36,42,43]. Duan et al. [35] reported the isolation of three viruses from C. tainanus Kieffer and C. jacobsoni Macfie in one such study; however, they only identified one of these viruses. This paper reports full genome sequencing of the remaining two viruses and from a third virus isolated from a pool of unsorted Culicoides collected in Shizong in 2019 and 2020.
2. Materials and Methods
2.1. Virus Isolates
Insects were collected from cattle farms at Wulong Village (24.64° N, 104.29° E, 975 m a.s.l.) in Shizong County, then identified, sorted and processed for virus isolation as described previously [35]. Details of two isolates, YNV/01-1 and YNV/03-2, were reported previously [35], while the same methods were used to isolate a third virus from a pool of approximately 100 unidentified parous female Culicoides free from visible blood.
2.2. Cells
Baby hamster kidney cell line BHK-21 and Aedes albopictus cell line C6/36 were cultured in minimum essential medium (MEM) containing 5% fetal bovine serum (FBS), 100 U/mL penicillin and 100 μg/mL streptomycin (Gibco, Thermo Fisher Scientific, Grand Island, NY, USA), under an atmosphere of 95% relative humidity and 5% CO2. C6/36 cells were cultured at 28 °C and seeded with viral isolate YNV/01-1, while BHK-21 cells were cultured at 37 °C and seeded with the other two isolates. At viral isolation step performed previously, both cell lines were used to culture potential viruses, respectively.
2.3. Extracting Viral Nucleic Acid for Test
An aliquot of 150 μL of supernatant from cell cultures exhibiting obvious cytopathic effect (CPE), was subjected to viral nucleic acid extraction using a MagMAX™-96 viral RNA Isolation kit (Am1836; Ambion, Austin, TX, USA) and a MagMAX™ Express machine (Applied Biosystems, Foster City, CA, USA) following the manufacturers’ directions.
2.4. RT-qPCR
RNA samples were denatured at 95 °C and quickly cooled on ice, and then subjected to testing by reverse transcript-quantitative polymerase chain reaction (RT-qPCR). For each sample, 20 μL of reaction solution was prepared using One Step PrimeScriptTM RT-PCR kit (Takara, Dalian, China) according to the manufacturer’s instructions, and 4 μL of RNA template, 0.4 μL of each primer and 0.8 μL of probe were added (Table S1). These primers and probes were against Akabane virus (AKAV), BAV, BTV, EHDV, Palyam virus (PALV), and TIBOV, respectively (Table S1). The RT-qPCR was performed on a 7500 Fast Real-time PCR machine (Applied Biosystems, Carlsbad, CA, USA) under the following cycling conditions: 45 °C, 5 min; 95 °C, 10 s; then 95 °C/5 s, 60 °C/34 s for 40 cycles. Fluorescence was measured at the end of each extension step.
2.5. Preparing Viral Genomic RNA
Each virus was inoculated to a T75 flask of monolayer cells. YNV/KM3 and YNV/03-2 were inoculated to BHK-21, while YNV/01-1 was inoculated to C6/36. When CPE appeared in 90% of cells at approximately 6 days post-infection, flasks were removed to −80 °C until use.
Cell pellets were collected after scraping cells and centrifugation at 360× g for 5 min. Viral genomes were extracted and prepared by methods modified from references [35,44,45]. Briefly, RNA was extracted from cell pellets using RNAiso-plus kit (Takara) according to the manufacturer’s instructions. Air dried RNA precipitant was dissolved by 100 μL of RNase free water, and then an equal volume of 4 M LiCl (Sigma-Aldrich, St. Louis, MO, USA) was added. Single-stranded RNA was removed by precipitation with 2 M LiCl at 4 °C overnight, followed by centrifugation at 15,000× g for 2 min. Nearly 200 μL of supernatant was transferred to a new tube and mixed with 500 μL of isopropanol, and then stored at −20 °C for 2 h. The dsRNA was pelleted by centrifugation at 15,000× g for 15 min, washed with 1 mL of 75% ethanol, air dried and suspended in 50 μL of RNase free water.
2.6. Amplifying Viral Genome
Complete viral genomic cDNA was synthesized by full-length amplification of cDNAs (FLAC) following the methods of Maan et al. [37,46]. Briefly, the 3′ end of the viral dsRNA was covalently linked with an anchor primer by T4 RNA ligase 1 (NEB, USA) at 16 °C overnight. The anchored dsRNA was purified by a MiniBEST Universal RNA Extraction kit (Takara) and used to synthesize cDNA with PrimeScriptTM II Reverse Transcriptase kit (Takara) following the manufacturer’s instructions.
2.7. DNA Electrophoresis
Viral genomic segments (RT-PCR produced dsDNA) were separated by electrophoresis (90 V for 2–3 h) in 1% agarose gel with dye Goldview II (Solarbio, Beijing, China). Fluorescent bands were screened by a Gel Doc™ XR+ System with Image Lab™ software (Bio-Rad, Hercules, CA, USA).
2.8. Complete Sequencing of Viral Genomes
Prepared viral genomic DNA samples were completely sequenced by the MAGIGEN Company (Guangzhou, China) using a HiSeq 2000 system (Illumina, San Diego, CA, USA) and software SOAPdenovo. Sequence data for each segment of each isolate were stored independently on Genbank.
2.9. Sequence Data and Phylogenetic Analysis
Public nucleic acid sequences used for analysis in this study were downloaded from GenBank and are listed in Table S2. Complete coding sequence (CDS) regions without stop codons were prepared for phylogenetic analysis. Prepared sequences were aligned by MUSCLE (Codons) with default parameters, and phylogenetic trees were built by Neighbor-Joining (NJ) algorithm (bootstrap = 1000, Model = Kimura 2, d: Transitins + Transversions, Gap Treatment=Pairwise deletion, Codon Position=1) or Maximum Parsimony (MP) algorithm (bootstrap = 1000, Gap treatment = Use all sites, Codon Position = 1) with default parameters. All above operations for analysis were finished using MEGA-11 software. All the phylogenetic trees in this study were constructed by the CDS sequences within viral genomic segments that code corresponding proteins.
3. Results
3.1. Primary Identification
RNA samples extracted from the supernatants of viral isolates infected cells (YNV/01-1, YNV/KM3 and YNV/03-2), were tested by RT-qPCR using six pairs of primers and probes, respectively (Table S1). As a result, the three viruses were identified as a single BAV and two EHDV (Table 1).
DNA samples produced by FLAC were subjected to electrophoresis. Segments of BAV (YNV/01-1) were evenly distributed generally, but the 5th band (seg5/seg6), the 6th band (seg7/seg8/seg9), and the 8th band (seg11/seg12) were composed of multiple segments (Figure 1). EHDV isolates YNV/KM3 and YNV/03-2 showed a 3-3-3-1 pattern (Figure 1).
3.2. Complete Genome Sequences
Sequence details for each segment of each virus and relevant GenBank accession numbers are provided in Table 2. Sequence data from each virus are provided in Tables S3–S5, respectively. Viral isolates YNV/01-1, YNV/KM3, and YNV/03-2 were identified as BAV, EHDV-1 and EHDV-7 according to their gene sequences (Table 2 and Tables S3–S5). The relationships between viral strains and Culicoides hosts are summarized in Table 3.
3.3. Phylogenetic Analysis to Confirm Virus Status
To classify the status of genera and viral species, three relatively conservative homofunctional proteins, namely VP1 functioning as a structural protein and RNA dependent RNA polymerase (RdRP), as well as the two major inner capsid proteins on a T = 2 lattice (T2) and a T = 13 lattice (T13), were used for phylogenetic analysis. Difficulties were encountered using NJ and ML algorithms to construct phylogenetic trees of the T13 therefore a MP algorithm was adopted.
In the phylogenetic trees of VP1, T2 proteins, and T13 proteins, Orbivirus and Seadornavirus and the different virus species were reciprocally monophyletic (Figure 2 and Figure 3 and Figure S1). The status of the BAV (YNV/01-1) and two EHDV (YNV/KM3 and YNV/03-2) isolates reported in this study were confirmed by these phylogenetic trees (Figure 2 and Figure 3, and Figure S1).
The VP1 genetic distance between the nodes of Orbivirus and Seadornavirus was 0.838, and the shortest distance between an Orbivirus (BTV or EHDV) and a Seadornavirus (BAV) was 1.316 (Figure 2). The VP1 distances between the clades of Orbivirus ranged from 0.192 to 0.692, and the distances between the clades of Seadornavirus ranged from 0.301 to 0.896 (Figure 2, Tables S6 and S7).
The T2 protein genetic distance between the nodes of Orbivirus and Seadornavirus was 0.920, and the shortest distance between an Orbivirus (BTV) and a Seadornavirus (BAV) was 1.654 (Figure 3). The T2 protein distances between the clades of Orbivirus ranged from 0.149 to 0.903, and the distances between the clades of Seadornavirus ranged from 0.441 to 1.369 (Figure 3, Tables S8 and S9). Sequences from Kadipiro (KDV) and Liao ning viruses (LNV) were found to be quite distant from other members of Seadornavirus such as Balaton virus, BAV, and Mangshi virus (MSV), with genetic distances greater than 1.0 (Table S9). The MP tree of T13 protein (Figure S1) exhibited similar topology to the NJ tree of T2 protein (Figure 3).
3.4. Phylogenetic Analysis for Viral Serotypes
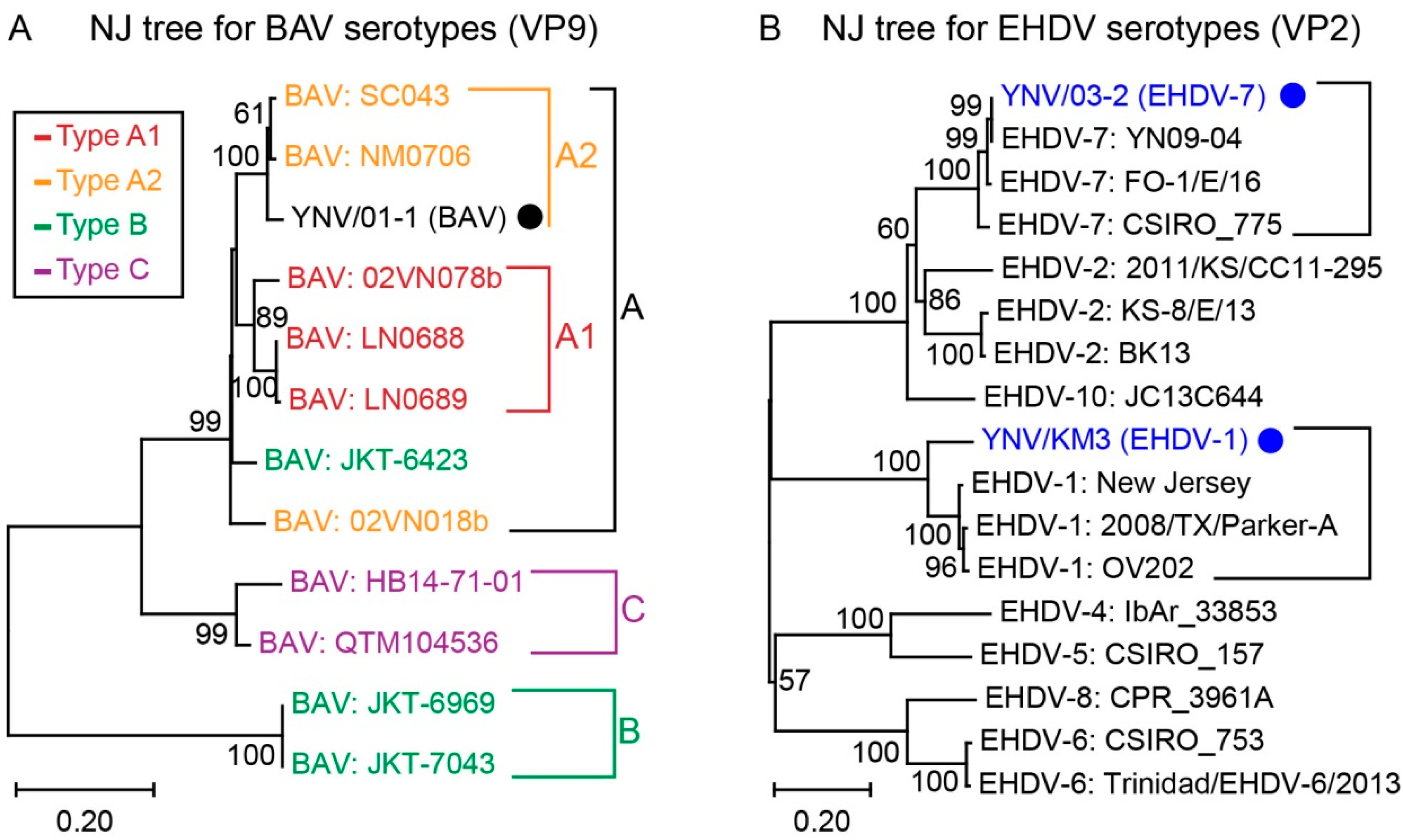
The phylogenetic tree to classify genotypes of BAV was constructed using VP9 sequences including 11 other BAV belonging to the 4 known genotypes (Figure 4A). In our phylogenetic tree, BAV were grouped into 3 major genotypes (A, B, and C) with VP9 genetic distances between genotypes of 0.719 (A to B), 0.266 (A to C), and 0.727 (B to C), respectively (Figure 4A). There were no clear clades within genotype A, although the so called type A1 and type A2 had a genetic distance of 0.070 (Figure 4A). According to this phylogenetic tree, the novel BAV strain YNV/01-1 reported in this study was grouped with genotype A or A2, however two publicly listed strains, JKT-6423 previously placed within genotype B and 02VN018b previously placed within genotype A2, were in a group that was sister to both the A1 and A2 genotypes (Figure 4A). Extended phylogenetic trees for Seadornavirus genotypes, in which VP10 of Balaton virus and LNV as well as VP9 of MSV were added, were constructed by NJ algorithm and MP algorithm, respectively. Both trees supported the above conclusions (Figure S2).
4. Discussion
Three viruses isolated from Culicoides without blood meals collected from Shizong County of Yunnan Province, China, were identified by full genome sequencing in this study. Viral strain YNV/KM3, isolated from unsorted Culicoides species was identified as EHDV, while strains YNV/01-1 and YNV/03-2 isolated from C. tainanus and C. orientalis, respectively [35], were identified as BAV (genotype A) and EHDV-7, respectively, in this study. The latter two results confirm that BAV and EHDV can infect C. tainanus and C. orientalis, respectively, and provide the first evidence that these species are potential vectors of these respective viruses.
Culicoides tainanus is a widespread species in Yunnan Province, China, and has recently been implicated as a potential important vector of BTV in Asia [47,48,49]. Some evidence has also been reported associating C. tainanus with TIBOV, although more evidence is required to confirm this association [35]. The isolation of BAV from this species represents both the third known association with an arbovirus and the first association of BAV with a known species of Culicoides.
Culicoides orientalis was reported to be a potential vector of Onchocerca gibsoni by Buckley [50], but has not previously been associated with the transmission of any virus. In this study, C. orientalis was confirmed to be infected by EHDV-7 in the field making this the first report associating C. orientalis with EHDV.
Three relatively conservative homofunctional proteins were used for genetic analysis between Orbivirus and Seadornavirus. VP1 with RdRP function was an essential protein in all viruses analyzed. The other two proteins analyzed, T2 and T13, are the major components of the inner capsid and were both found to provide good resolution to separate viral species within genus Orbivirus [51,52]. Primarily, the structure analysis of BTV inner capsid suggested that the icosahedral lattice of inner capsid was constructed by 120 copies of VP3 and 780 copies of VP7, corresponding to a VP3:VP7 composition ratio of 2:13 [53,54,55]. In this study, we tried to construct phylogenetic trees using the homogenous proteins of BTV VP3 (T2) and VP7 (T13) proteins (Figure 3, Figure S1 and Table S2). We, however, encountered problems with constructing both NJ and ML algorithm-based phylogenetic trees for the segments encoding the T13 protein when trying to encompass both the Orbivirus and Seadornavirus genera. This likely reflected the high variability in structure and function of this protein within each group. T13 proteins of BTV provide the ligand for binding to cellular receptors of susceptible insect cells [4,7] and are consequently likely to evolve to avoid immune responses of insects much the same, as the proteins that provide the ligand for attachment to vertebrate cells are serotype specific, although insect did not produce antibodies [56,57]. Given this selection pressure, genes that code for these proteins in other viruses are also likely to be informative in separating strains of virus, as shown in Figure S1.
Thus far, only three viruses, BAV, KDV, and LNV, have been placed into genus Seadornavirus by ICTV [11], and a BAV-like virus (Balaton virus) was reported and considered as a novel Seadornavirus by Reuter et al. in 2013 [58]. In the phylogenetic trees of VP1, T2 and T13, KDV and LNV were genetically distant from other species of Seadornavirus suggesting that KDV and LNV might be better placed in a novel genus or perhaps, subgenus [8,9,10,58,59].
Existing strains of BAV have traditionally been grouped into four genotypes (A1, A2, B, and C) through phylogenetic analysis of the 12th segment encoding a non-structural protein [13,16,18,60]. However, it is not reliable to classify genotypes or serotypes by non-structural proteins. VP9 of BAV is known as the ligand binding to the cellular receptor of susceptive mammal cells [58,60] so BAV serotypes should be determined by VP9, which is subjected to evolutionary stress from mammalian immune systems such as neutralizing antibodies. This is also more consistent with the method of classifying other arthropod-borne Reoviruses like BTV [61]. Our phylogenetic analysis of BAV VP9 found that BAV strains were in general agreement with the system based on VP12, excepting that two viruses were sister to rather than belonging to the A1/A2 clades, and one of these, strain JKT-6423, was placed into genotype A rather than genotype B as previously reported (Figure 4A). However, the VP9/9th segment sequences were lacking for some BAV isolates [13], which compromised the genotype investigation. We encourage future workers to include VP9 data in their descriptions of novel BAV viruses, as this will allow more comprehensive comparison between the classification provided by VP9 and VP12 for serotyping these viruses.
All the known serotypes of EHDV except for EHDV-2 and EHDV-4 have been previously reported in China [28,62,63,64], and seropositive rates of EHDV antibodies are high in cattle in southern China, including Yunnan Province (Duan et al., under review), but no outbreak of clinical disease has yet been reported in Yunnan Province. The only documented outbreak of this disease in mainland China was eight cattle with Ibaraki disease-like symptoms reported in 1987 [65]. In East Asia, a large epidemic of Ibaraki disease in cattle, caused by the Ibaraki strain of EHDV-2, occurred in Japan between 1959 and 1960 [66]; an outbreak of Ibaraki-like disease in cattle in south Japan in 1997 was caused by EHDV-7 strains [26,47].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/v14050971/s1, Figure S1: Phylogenetic tree of representative species of Orbivirus and Seadornavirus based on T13 core capsid protein genes. Figure S2: Phylogenetic trees for genotype analysis of representative Seadornavirus. Table S1: Primers and probes used for RT-qPCR tests. Table S2: Data of sequences downloaded from GenBank and used in this study. Table S3: The general genome data of BAV strain YNV/01-1. Table S4: The general genome data of EHDV-1 strain YNV/KM3. Table S5: The general genome data of EHDV-7 strain YNV/03-2. Table S6: VP1 genetic distances between some species of Orbivirus. Table S7: VP1 genetic distances between some species of Seadornavirus. Table S8: T2 protein genetic distances between some species of Orbivirus. Table S9: T2 protein genetic distances between some species of Seadornavirus.
Author Contributions
Conceptualization, Y.D., L.L.; methodology, Y.D., Z.Y.; software, Y.D., Z.Y.; validation, Y.D., Z.Y.; formal analysis, Y.D., Z.Y.; investigation, Y.D., Z.Y., J.X., L.L.; resources, L.L.; data curation, Y.D., Z.Y., L.L.; writing—original draft preparation, Y.D., G.B.; writing—review and editing, Y.D., G.B.; supervision, L.L.; project administration, L.L.; visualization, Y.D.; funding acquisition, Y.D., L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32160846), and the Foreign Experts Project of Yunnan Province (YNZ2019002 and YNZ2020015).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequence data are openly available in GenBank (https://www.ncbi.nlm.nih.gov/, accessed on: 22 February 2022).
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schiff, L.A.; Nibert, M.L.; Tyler, K.L. Orthoreoviruses and Their Replication. In Fields Virology, 5th ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2388–2471. [Google Scholar]
- ICTV Virus Taxonomy: 2019 Release. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 9 May 2020).
- Hassan, S.S.; Roy, P. Expression and functional characterization of bluetongue virus VP2 protein: Role in cell entry. J. Virol. 1999, 73, 9832–9842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.-H.; Nason, E.; Staeuber, N.; Jiang, W.; Monastryrskaya, K.; Roy, P. RGD Tripeptide of Bluetongue Virus VP7 Protein Is Responsible for Core Attachment to Culicoides Cells. J. Virol. 2001, 75, 3352–3362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, P. Orbiviruses. In Fields Virology, 5th ed.; Fields, B.N., Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer/Lippincott Williams & Wilkins Health: Philadelphia, PA, USA, 2007; Volume 2, pp. 2541–2568. [Google Scholar]
- Mills, M.; Michel, K.; Pfannenstiel, R.S.; Ruder, M.G.; Veronesi, E.; Nayduch, D. Culicoides–virus interactions: Infection barriers and possible factors underlying vector competence. Curr. Opin. Insect Sci. 2017, 22, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Mertens, P.; Burroughs, J.; Walton, A.; Wellby, M.; Fu, H.; O’Hara, R.; Brookes, S.; Mellor, P. Enhanced Infectivity of Modified Bluetongue Virus Particles for Two Insect Cell Lines and for Two Culicoides Vector Species. Virology 1996, 217, 582–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attoui, H.; De Micco, P.; De Lamballerie, X.; Billoir, F.; Biagini, P. Complete sequence determination and genetic analysis of Banna virus and Kadipiro virus: Proposal for assignment to a new genus (Seadornavirus) within the family Reoviridae. J. Gen. Virol. 2000, 81, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Jaafar, F.M.; Belhouchet, M.; Tao, S.; Chen, B.; Liang, G.; Tesh, R.B.; De Micco, P.; De Lamballerie, X. Liao ning virus, a new Chinese Seadornavirus that replicates in transformed and embryonic mammalian cells. J. Gen. Virol. 2006, 87, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, H.; He, Y.; Zhou, Y.; Meng, J.; Zhu, W.; Chen, H.; Liao, D.; Man, Y. Isolation and Genetic Characterization of Mangshi Virus: A Newly Discovered Seadornavirus of the Reoviridae Family Found in Yunnan Province, China. PLoS ONE 2015, 10, e0143601. [Google Scholar] [CrossRef]
- ICTV ICTV Master Species List 2020.v1. Available online: https://talk.ictvonline.org/files/master-species-lists/m/msl/12314 (accessed on 21 February 2022).
- Xu, P.T.; Wang, Y.M.; Zuo, J.M.; Lin, J.W.; Xu, P.M. New Orbiviruses isolated from patients with unknown fever and encephalitis in Yunnan Province Chin. J. Virol. 1990, 6, 27–33. [Google Scholar]
- Liu, H.; Li, M.H.; Zhai, Y.G.; Meng, W.S.; Sun, X.H.; Cao, Y.X.; Fu, S.H.; Wang, H.Y.; Xu, L.H.; Tang, Q.; et al. Banna virus, China, 1987–2007. Emerg. Infect. Dis. 2010, 16, 514–517. [Google Scholar] [CrossRef]
- Nabeshima, T.; Nga, P.T.; Guillermo, P.; Parquet MD, C.; Yu, F.; Thuy, N.T.; Trang, B.M.; Hien, N.T.; Nam, V.S.; Inoue, S.; et al. Isolation and Molecular Characterization of Banna Virus from Mosquitoes, Vietnam. Emerg. Infect. Dis. 2008, 14, 1276–1279. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Gao, X.-Y.; Fu, S.-H.; Li, M.-H.; Zhai, Y.-G.; Meng, W.-S.; Sun, X.-H.; Lv, Z.; Wang, H.-Y.; Shen, X.-X.; et al. Molecular evolution of emerging Banna virus. Infect. Genet. Evol. 2016, 45, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Li, Y.; Fu, S.; Lei, W.; Guo, X.; Feng, Y.; Gao, X.; Li, X.; Yang, Z.; Xu, Z.; et al. Genome sequencing and phylogenetic analysis of Banna virus (genus Seadornavirus, family Reoviridae) isolated from Culicoides. Sci. China Life Sci. 2017, 60, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.P.; Xie, X.C.; Zhi, Q.; Ma, L.; Liu, Y.D.; Dangzheng, W.; Abdureciti, A.; Hashan, Y.; Zhang, Y.J.; Wang, C.; et al. First isolation of new Orbivirus (Banna) from ticks and infected cattle sera in Xinjiang. Endem. Dis. Bull. 1992, 7, 64–69. [Google Scholar]
- Xia, H.; Liu, H.; Zhao, L.; Atoni, E.; Wang, Y.; Yuan, Z. First Isolation and Characterization of a Group C Banna Virus (BAV) from Anopheles sinensis Mosquitoes in Hubei, China. Viruses 2018, 10, 555. [Google Scholar] [CrossRef] [Green Version]
- Shope, R.E.; MacNamara, L.G.; Mangold, R. A virus-induced epizootic hemorrhagic disease of the virginia white-tailed deer (Odocoileus virginianus). J. Exp. Med. 1960, 111, 155–170. [Google Scholar] [CrossRef]
- MacLachlan, N.; Zientara, S.; Savini, G.; Daniels, P. Epizootic haemorrhagic disease. Rev. Sci. Et Tech. l’OIE 2015, 34, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Casey, C.; Rathbun, S.; Stallknecht, D.; Ruder, M. Spatial Analysis of the 2017 Outbreak of Hemorrhagic Disease and Physiographic Region in the Eastern United States. Viruses 2021, 13, 550. [Google Scholar] [CrossRef]
- Cêtre-Sossah, C.; Roger, F.; Sailleau, C.; Rieau, L.; Zientara, S.; Bréard, E.; Viarouge, C.; Beral, M.; Esnault, O.; Cardinale, E. Epizootic haemorrhagic disease virus in Reunion Island: Evidence for the circulation of a new serotype and associated risk factors. Veter. Microbiol. 2014, 170, 383–390. [Google Scholar] [CrossRef]
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides Biting Midges: Their Role as Arbovirus Vectors. Annu. Rev. Èntomol. 2000, 45, 307–340. [Google Scholar] [CrossRef]
- Savini, G.; Afonso, A.; Mellor, P.; Aradaib, I.; Yadin, H.; Sanaa, M.; Wilson, W.; Monaco, F.; Domingo, M. Epizootic heamorragic disease. Res. Vet. Sci. 2011, 91, 1–17. [Google Scholar] [CrossRef]
- OIE Epizootic Haemorrhagic Disease (Infection with Epizootic Hemorrhagic Disease Virus). Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.07_EHD.pdf (accessed on 30 November 2021).
- Shirafuji, H.; Kato, T.; Yamakawa, M.; Tanaka, T.; Minemori, Y.; Yanase, T. Characterization of genome segments 2, 3 and 6 of epizootic hemorrhagic disease virus strains isolated in Japan in 1985–2013: Identification of their serotypes and geographical genetic types. Infect. Genet. Evol. 2017, 53, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Hiromatsu, R.; Kaida, M.; Kato, T.; Yanase, T.; Shirafuji, H. Isolation of epizootic hemorrhagic disease virus serotype 7 from cattle showing fever in Japan in 2016 and improvement of a reverse transcription-polymerase chain reaction assay to detect epizootic hemorrhagic disease virus. J. Veter. Med. Sci. 2021, 83, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, Z.; Wang, J.; Li, Z.; Yang, Z.; Liao, D.; Zhu, J.; Li, H. Novel Serotype of Epizootic Hemorrhagic Disease Virus, China. Emerg. Infect. Dis. 2020, 26, 3081–3083. [Google Scholar] [CrossRef] [PubMed]
- Yanase, T.; Murota, K.; Hayama, Y. Endemic and Emerging Arboviruses in Domestic Ruminants in East Asia. Front. Veter- Sci. 2020, 7, 168. [Google Scholar] [CrossRef]
- Cottingham, S.; White, Z.; Wisely, S.; Campos-Krauer, J. A Mortality-Based Description of EHDV and BTV Prevalence in Farmed White-Tailed Deer (Odocoileus virginianus) in Florida, USA. Viruses 2021, 13, 1443. [Google Scholar] [CrossRef]
- Temizel, E.M.; Yesilbag, K.; Batten, C.; Senturk, S.; Maan, N.S.; Mertens, P.; Batmaz, H. Epizootic Hemorrhagic Disease in Cattle, Western Turkey. Emerg. Infect. Dis. 2009, 15, 317–319. [Google Scholar] [CrossRef]
- Borkent, A.; Dominiak, P. Catalog of the Biting Midges of the World (Diptera: Ceratopogonidae). Zootaxa 2020, 4787, 1–377. [Google Scholar] [CrossRef]
- ICTV ICTV Taxonomy History: Peribunyaviridae. Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=202000081 (accessed on 21 February 2022).
- Xing, S.; Guo, X.; Zhang, X.; Zhao, Q.; Li, L.; Zuo, S.; An, X.; Pei, G.; Sun, Q.; Cheng, S.; et al. A novel mosquito-borne reassortant Orbivirus isolated from Xishuangbanna, China. Virol. Sin. 2016, 32, 159–162. [Google Scholar] [CrossRef]
- Duan, Y.L.; Yang, Z.X.; Bellis, G.; Li, L. Additional file 2 of Isolation of Tibet Orbivirus from Culicoides jacobsoni (Diptera, Ceratopogonidae) in China. Parasites Vectors 2021, 14, 1–11. [Google Scholar] [CrossRef]
- Zhang, N.; Li, Z.; Zhang, F.; Zhu, J. Studies on bluetongue disease in the People’s Republic of China. Veter. Ital. 2010, 40, 51–56. [Google Scholar]
- Duan, Y.L.; Miao, H.S.; Liao, D.F.; Kou, M.L.; Li, Z.H.; Wang, Z.; Li, H.C.; Li, L. The serologic investigation and viral isolation of bluetongue virus in Shangri-La in Southwest China. Transbound. Emerg. Dis. 2019, 66, 2353–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Wang, F.; Chang, J.; Zhang, Y.; Zhu, J.; Li, H.; Yu, L. Identification and complete-genome phylogenetic analysis of an epizootic hemorrhagic disease virus serotype 7 strain isolated in China. Arch. Virol. 2019, 164, 3121–3126. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Lun, Z.R.; James, A.A.; Chen, X.G. Review: Dengue Fever in mainland China. Am. J. Trop Med. Hyg. 2010, 83, 664–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.-C.; Zhao, H.; Li, L.-H.; Jiang, T.; Hong, W.-X.; Wang, J.; Zhao, L.-Z.; Yang, H.-Q.; Ma, D.-H.; Bai, C.-H.; et al. Severe dengue outbreak in Yunnan, China, 2013. Int. J. Infect. Dis. 2014, 27, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, Y.; Lu, S.; Dong, J.; Xu, H.; Zhang, Q.; Weng, R.; Yin, Y.; He, R.; Fang, P.; et al. Epidemiological survey and screening strategy for dengue virus in blood donors from Yunnan Province. BMC Infect. Dis. 2021, 21, 1–7. [Google Scholar] [CrossRef]
- Zhang, N.Z.; Zhang, K.L.; Li, Z.H.; Chan-yu, N.C.; Hu, Y.L.; Li, G.; Zhao, K.; Zou, F.Z.; Xu, W.Z.; Li, S.X.; et al. A Report of the Investigation and Research of Bluetongue on Sheep. Yunnan J. Anim. Sci. Vet. Med. 1989, 18, 3–13. [Google Scholar]
- Kirkland, P.D.; Zhang, N.; Hawkes, R.A.; Li, Z.; Zhang, F.; Davis, R.J.; Sanders, D.A.; Li, H.; Zhang, K.; Jiang, B.F.; et al. Studies on the epidemiology of bluetongue virus in China. Epidemiol. Infect. 2002, 128, 257–263. [Google Scholar] [CrossRef]
- Attoui, H.; Jaafar, F.M.; Belhouchet, M.; Biagini, P.; Cantaloube, J.-F.; de Micco, P.; de Lamballerie, X. Expansion of family Reoviridae to include nine-segmented dsRNA viruses: Isolation and characterization of a new virus designated Aedes pseudoscutellaris reovirus assigned to a proposed genus (Dinovernavirus). Virology 2005, 343, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Belaganahalli, M.N.; Maan, S.; Maan, N.S.; Brownlie, J.; Tesh, R.; Attoui, H.; Mertens PP, C. Genetic Characterization of the Tick-Borne Orbiviruses. Viruses 2015, 7, 2185–2209. [Google Scholar] [CrossRef]
- Maan, S.; Rao, S.; Maan, N.S.; Anthony, S.J.; Attoui, H.; Samuel, A.R.; Mertens PP, C. Rapid cDNA synthesis and sequencing techniques for the genetic study of bluetongue and other dsRNA viruses. J. Virol. Methods 2007, 143, 132–139. [Google Scholar] [CrossRef]
- Kato, T.; Shirafuji, H.; Tanaka, S.; Sato, M.; Yamakawa, M.; Tsuda, T.; Yanase, T. Bovine Arboviruses in Culicoides Biting Midges and Sentinel Cattle in Southern Japan from 2003 to 2013. Transbound. Emerg. Dis. 2015, 63, e160–e172. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.L.; Bellis, G.; Li, L.; Li, H.C.; Miao, H.S.; Kou, M.L.; Liao, D.F.; Wang, Z.; Gao, L.; Li, J.Z. Potential vectors of bluetongue virus in high altitude areas of Yunnan Province, China. Parasites Vectors 2019, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.L.; Li, L.; Bellis, G.; Yang, Z.X.; Li, H.C. Detection of bluetongue virus in Culicoides spp. in southern Yunnan Province, China. Parasites Vectors 2021, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.J.C. On Culicoides as a Vector of Onchocerca gibsoni (Cleland & Johnston, 1910). J. Helminthol. 1938, 16, 121–158. [Google Scholar] [CrossRef]
- Attoui, H.; Stirling, J.M.; Munderloh, U.G.; Billoir, F.; Brookes, S.M.; Burroughs, J.N.; De Micco, P.; Mertens, P.; De Lamballerie, X. Complete sequence characterization of the genome of the St Croix River virus, a new Orbivirus isolated from cells of Ixodes scapularis. J. Gen. Virol. 2001, 82, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Jaafar, F.M.; Belhouchet, M.; Aldrovandi, N.; Tao, S.; Chen, B.; Liang, G.; Tesh, R.B.; De Micco, P.; De Lamballerie, X. Yunnan Orbivirus, a new Orbivirus species isolated from Culex tritaeniorhynchus mosquitoes in China. J. Gen. Virol. 2005, 86, 3409–3417. [Google Scholar] [CrossRef] [PubMed]
- Grimes, J.M.; Burroughs, J.N.; Gouet, P.; Diprose, J.M.; Malby, R.L.; Ziéntara, S.; Mertens, P.; Stuart, D. The atomic structure of the bluetongue virus core. Nature 1998, 395, 470–478. [Google Scholar] [CrossRef]
- Grimes, J.M.; Jakana, J.; Ghosh, M.; Basak, A.K.; Roy, P.; Chiu, W.I.; Stuart, D.; Prasad, B.V. An atomic model of the outer layer of the bluetongue virus core derived from X-ray crystallography and electron cryomicroscopy. Structure 1997, 5, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Burroughs, J.; Grimes, J.; Mertens, P.; Stuart, D. Crystallization and Preliminary X-ray Analysis of the Core Particle of Bluetongue Virus. Virology 1995, 210, 217–220. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.R. The insect cellular immune response. Insect. Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Mukherjee, D.; Das, S.; Begum, F.; Mal, S.; Ray, U. The Mosquito Immune System and the Life of Dengue Virus: What We Know and Do Not Know. Pathogens 2019, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, G.; Boros, Á.; Delwart, E.; Pankovics, P. Novel Seadornavirus (family Reoviridae) related to Banna virus in Europe. Arch. Virol. 2013, 158, 2163–2167. [Google Scholar] [CrossRef] [PubMed]
- Prow, N.A.; Mah, M.G.; Deerain, J.M.; Warrilow, D.; Colmant AM, G.; O’Brien, C.A.; Harrison, J.J.; McLean, B.J.; Hewlett, E.K.; Piyasena TB, H.; et al. New genotypes of Liao ning virus (LNV) in Australia exhibit an insect-specific phenotype. J. Gen. Virol. 2018, 99, 596–609. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, F.M.; Attoui, H.; Mertens PP, C.; De Micco, P.; De Lamballerie, X. Structural organization of an encephalitic human isolate of Banna virus (genus Seadornavirus, family Reoviridae). J. Gen. Virol. 2005, 86, 1147–1157. [Google Scholar] [CrossRef]
- Maan, S.; Maan, N.S.; Nomikou, K.; Veronesi, E.; Bachanek-Bankowska, K.; Belaganahalli, M.N.; Attoui, H.; Mertens PP, C. Complete Genome Characterisation of a Novel 26th Bluetongue Virus Serotype from Kuwait. PLoS ONE 2011, 6, e26147. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wu, J.; Zhu, J.; Wang, J.; Lyu, M.; Xiao, L.; Yang, Z.; Liao, D.; Li, H.; Yang, H. Isolation and genetic characterization of epizootic hemorrhagic disease virus serotype 1 strains prevalent in China from 2013 to 2019. J. South. Agric. 2021, 52, 2043–2062. [Google Scholar]
- Zhang, Y.X.; Lin, J.; Cao, Y.Y.; Zhu, J.B.; Du, Y.C.; Yang, Z.X.; Yao, J.; Li, H.C.; Wu, J.M. Investigation of the serotypes of epizootic hemorrhagic disease virus and analysis of their distribution in Guangxi. Shanghai J. Anim. Husb. Vet. Med. 2016, 61, 19–21. [Google Scholar]
- Kou, M.; Yang, Z.; Li, L.; Zhu, J.; Gao, L.; Miao, H. Serological investigation and serotype identification of epidemic hemorrhagic virus in Yunnan border area. China Anim. Husb. Vet. Med. 2019, 46, 3065–3074. [Google Scholar]
- Xue, D.M.; Zhang, G.X.; Liu, A.D.; Zhang, Q.Y.; Li, D.C.; Guo, Y.Z.; Huang, D.J.; Yang, B.S. Suspicious Ibaraki disease in cattle. J. Anim. Sci. Vet. Med. 1987, 6, 43–44. [Google Scholar]
- Omori, T.; Inaba, Y.; Morimoto, T.; Tanaka, Y.; Ishitani, R. Ibaraki virus, an agent of epizootic disease of cattle resembling bluetongue. I. Epidemiologic, clinical and pathologic observations and experimental transmission to calves. Jpn. J. Microbiol. 1969, 13, 139–157. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Electrophoresis gel displaying genomic fragments of three viruses isolated from Culicoides spp. in Yunnan Province, China. DNA marker and DNA samples of YNV/01-1 (BAV), YNV/03-2 (EHDV-7), and YNV/KM3 (EHDV-1) were separated by electrophoresis in 1% agarose gel. The EHDV segments are annotated on the right.
Figure 1.
Electrophoresis gel displaying genomic fragments of three viruses isolated from Culicoides spp. in Yunnan Province, China. DNA marker and DNA samples of YNV/01-1 (BAV), YNV/03-2 (EHDV-7), and YNV/KM3 (EHDV-1) were separated by electrophoresis in 1% agarose gel. The EHDV segments are annotated on the right.

Figure 2.
Phylogenetic NJ tree of representative species of Orbivirus and Seadornavirus based on VP1 (RdRP) genes. Viral species and voucher names are provided; novel isolates reported in this study are highlighted in blue circles. Bootstrap values < 50% omitted.
Figure 2.
Phylogenetic NJ tree of representative species of Orbivirus and Seadornavirus based on VP1 (RdRP) genes. Viral species and voucher names are provided; novel isolates reported in this study are highlighted in blue circles. Bootstrap values < 50% omitted.

Figure 3.
Phylogenetic NJ tree of representative species of Orbivirus and Seadornavirus based on T2 core capsid protein genes. Viral species and voucher names are provided; novel isolates reported in this study are highlighted in blue circles. Bootstrap values less than 50% are not shown.
Figure 3.
Phylogenetic NJ tree of representative species of Orbivirus and Seadornavirus based on T2 core capsid protein genes. Viral species and voucher names are provided; novel isolates reported in this study are highlighted in blue circles. Bootstrap values less than 50% are not shown.
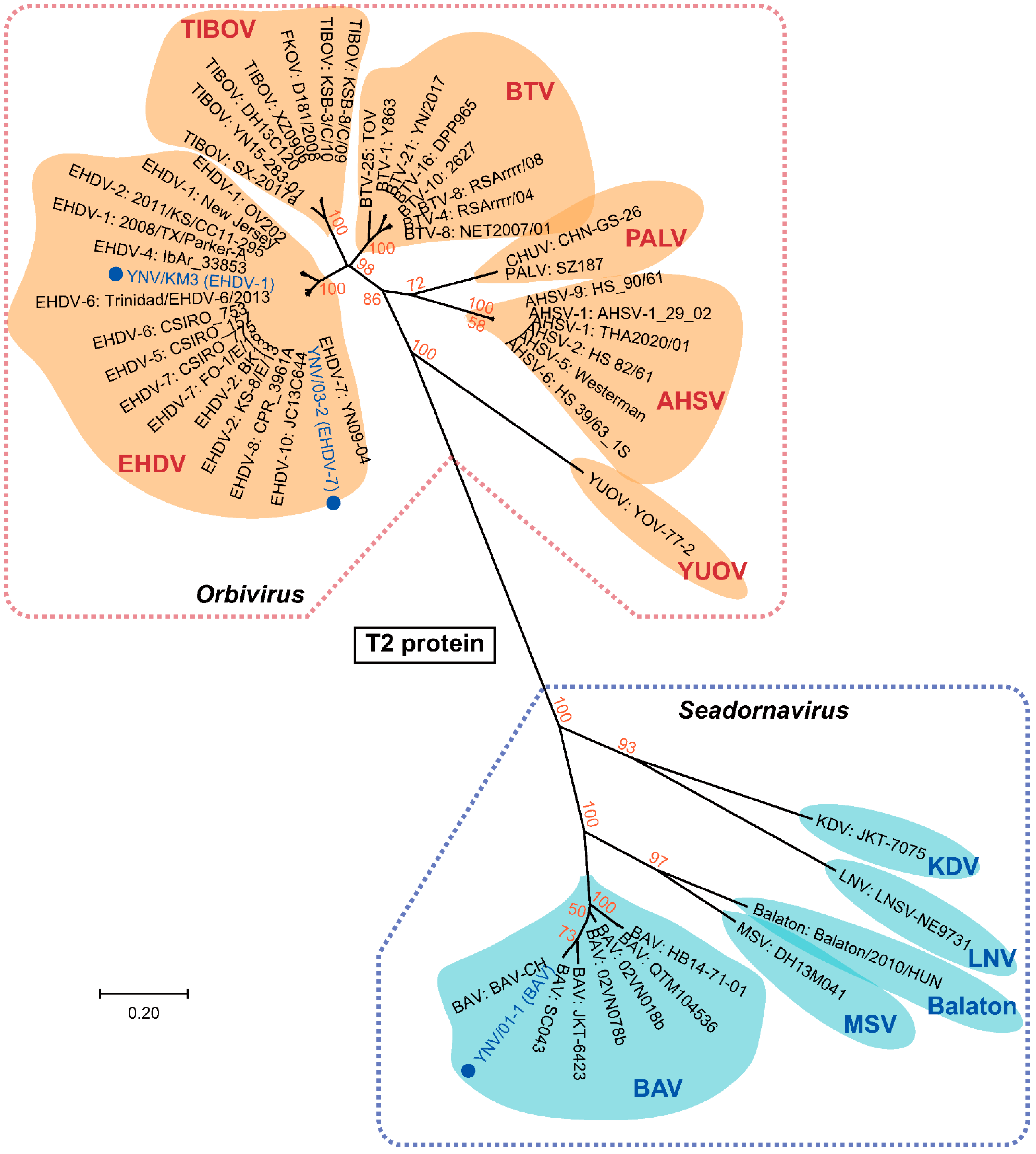
Figure 4.
Phylogenetic analysis of BAV genotypes and EHDV serotypes. Phylogenetic trees constructed by NJ algorithm using complete CDS of BAV VP9 (A) and EHDV VP2 (B), respectively. Viral species and voucher numbers are provided; novel BAV isolate and EHDV isolates reported in this study are labeled by black solid circle and blue solid circles respectively. Bootstrap values < 50% are omitted. In tree (A), previously reported genotypes are highlighted by red (type A1), orange (type A2), green (type B), and violet (type C), respectively.
Figure 4.
Phylogenetic analysis of BAV genotypes and EHDV serotypes. Phylogenetic trees constructed by NJ algorithm using complete CDS of BAV VP9 (A) and EHDV VP2 (B), respectively. Viral species and voucher numbers are provided; novel BAV isolate and EHDV isolates reported in this study are labeled by black solid circle and blue solid circles respectively. Bootstrap values < 50% are omitted. In tree (A), previously reported genotypes are highlighted by red (type A1), orange (type A2), green (type B), and violet (type C), respectively.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Quantification cycle (Cq) values of RT-qPCR tests of three viruses isolated from Culicoides spp. in Yunnan Province, China.
Table 1.
Quantification cycle (Cq) values of RT-qPCR tests of three viruses isolated from Culicoides spp. in Yunnan Province, China.
| Isolate | Cell | Target Virus Species | |||||
|---|---|---|---|---|---|---|---|
| AKAV | BAV | BTV | EHDV | PALV | TIBOV | ||
| YNV/01-1 | C6/36 | NA | 16.2 | NA | NA | NA | NA |
| YNV/KM3 | BHK-21 | NA | NA | NA | 26.3 | NA | NA |
| YNV/03-2 | BHK-21 | NA | NA | NA | 27.4 | NA | NA |
Note: NA means no signal detected, therefore no Cq value.
Table 2.
Genomic segment lengths and Genbank accession numbers of three viruses isolated from Culicoides spp. in Yunnan Province, China.
Table 2.
Genomic segment lengths and Genbank accession numbers of three viruses isolated from Culicoides spp. in Yunnan Province, China.
| Seg | YNV/01-1 (BAV) | YNV/KM3 (EHDV-1) | YNV/03-2 (EHDV-7) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Length (bp) | Gene | GenBank No. | Length (bp) | Gene | GenBank No. | Length (bp) | Gene | GenBank No. | |
| 1 | 3762 | VP1 | OM953801 | 3942 | VP1 | OM953791 | 3942 | VP1 | OM953813 |
| 2 | 3050 | VP2 (T2) | OM953802 | 2968 | VP2 | OM953792 | 3002 | VP2 | OM953814 |
| 3 | 2399 | VP3 | OM953803 | 2768 | VP3 (T2) | OM953793 | 2768 | VP3 (T2) | OM953815 |
| 4 | 2032 | VP4 | OM953804 | 1984 | VP4 | OM953794 | 1984 | VP4 | OM953816 |
| 5 | 1686 | VP5 | OM953805 | 1769 | NS1 | OM953795 | 1769 | NS1 | OM953817 |
| 6 | 1671 | VP6 | OM953806 | 1640 | VP5 | OM953796 | 1641 | VP5 | OM953818 |
| 7 | 1137 | VP7 | OM953807 | 1162 | VP7 (T13) | OM953797 | 1162 | VP7 (T13) | OM953819 |
| 8 | 1119 | VP8 (T13) | OM953808 | 1192 | NS2 | OM953798 | 1192 | NS2 | OM953820 |
| 9 | 1100 | VP9 | OM953809 | 1071 | VP6 | OM953799 | 1074 | VP6 | OM953821 |
| 10 | 977 | VP10 | OM953810 | 810 | NS3 | OM953800 | 810 | NS3 | OM953822 |
| 11 | 867 | VP11 | OM953811 | ||||||
| 12 | 861 | VP12 | OM953812 | ||||||
Table 3.
Isolation details of three viruses isolated from Culicoides collected from Wulong village in Shizong County, Yunnan Province, China.
Table 3.
Isolation details of three viruses isolated from Culicoides collected from Wulong village in Shizong County, Yunnan Province, China.
| Isolates | Identification | Hosts | Collection Date | Cells for Isolation | |
|---|---|---|---|---|---|
| C6/36 | BHK-21 | ||||
| YNV/KM3 | EHDV-1 | Culicoides spp. | 20 August 2019 | + | + |
| YNV/01-1 | BAV | C. tainanus | 20 May2020 | + | - |
| YNV/03-2 | EHDV-7 | C. orientalis | 9 June 2020 | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Duan, Y.; Yang, Z.; Bellis, G.; Xie, J.; Li, L. Full Genome Sequencing of Three Sedoreoviridae Viruses Isolated from Culicoides spp. (Diptera, Ceratopogonidae) in China. Viruses 2022, 14, 971. https://doi.org/10.3390/v14050971
AMA Style
Duan Y, Yang Z, Bellis G, Xie J, Li L. Full Genome Sequencing of Three Sedoreoviridae Viruses Isolated from Culicoides spp. (Diptera, Ceratopogonidae) in China. Viruses. 2022; 14(5):971. https://doi.org/10.3390/v14050971
Chicago/Turabian StyleDuan, Yingliang, Zhenxing Yang, Glenn Bellis, Jiarui Xie, and Le Li. 2022. "Full Genome Sequencing of Three Sedoreoviridae Viruses Isolated from Culicoides spp. (Diptera, Ceratopogonidae) in China" Viruses 14, no. 5: 971. https://doi.org/10.3390/v14050971
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.